
Radiotherapy Combined with PD-1 Inhibition Increases NK Cell Cytotoxicity towards Nasopharyngeal Carcinoma Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Patient-Derived Xenograft
2.3. Animal Studies
2.4. Isolation of Primary Human NK Cells
2.5. Reagents
2.6. Cell Viability Assay
2.7. Cell Cycle Analysis
2.8. Determination of Radiation Dose
2.9. Flow Cytometry
2.10. Calcein Release Assay
2.11. Statistical Analysis
3. Results
3.1. Radiotherapy Sensitizes NPC Cells to Killing by NK Cells
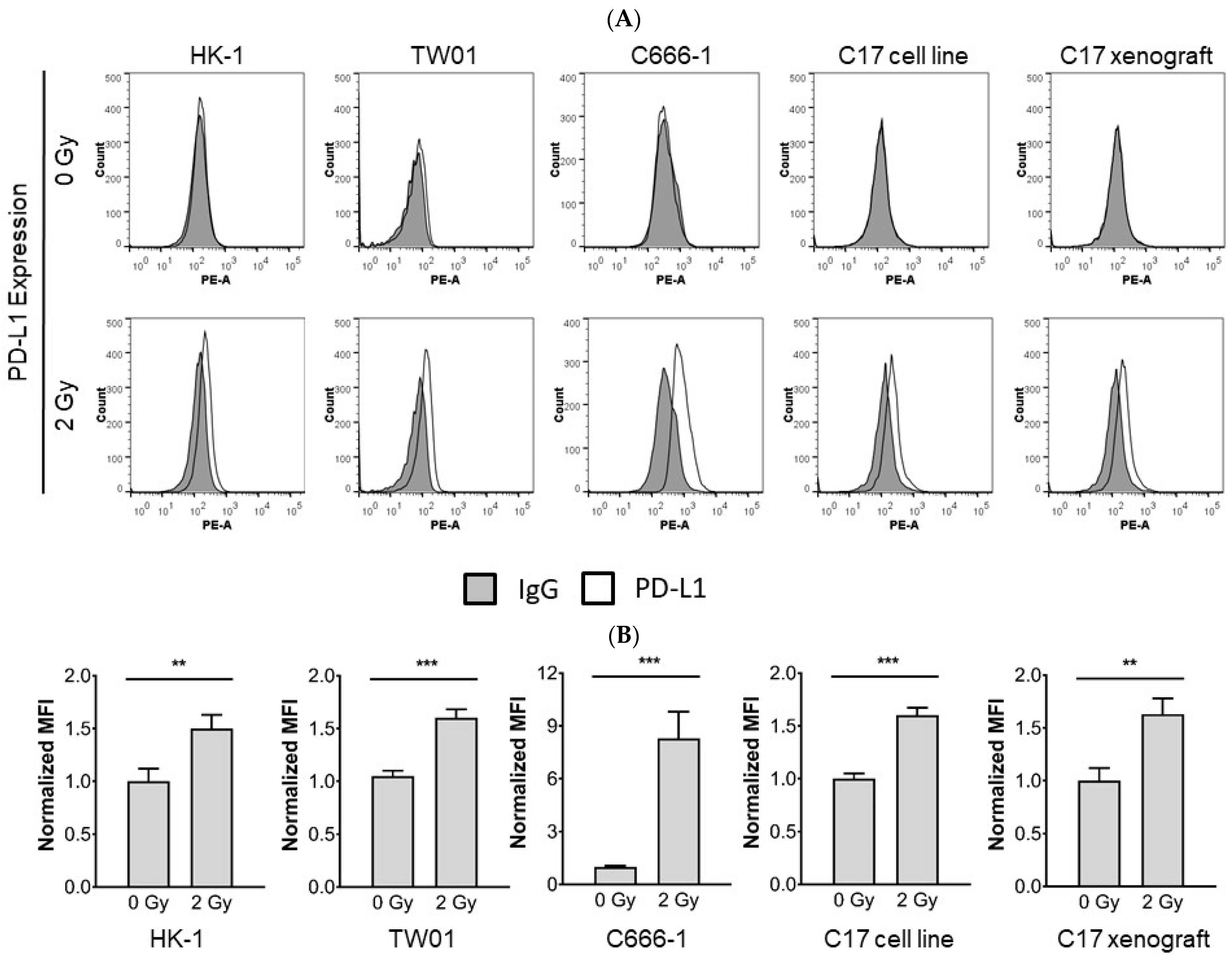
3.2. Radiotherapy Induces Upregulation of PD-L1 in NPC Cells
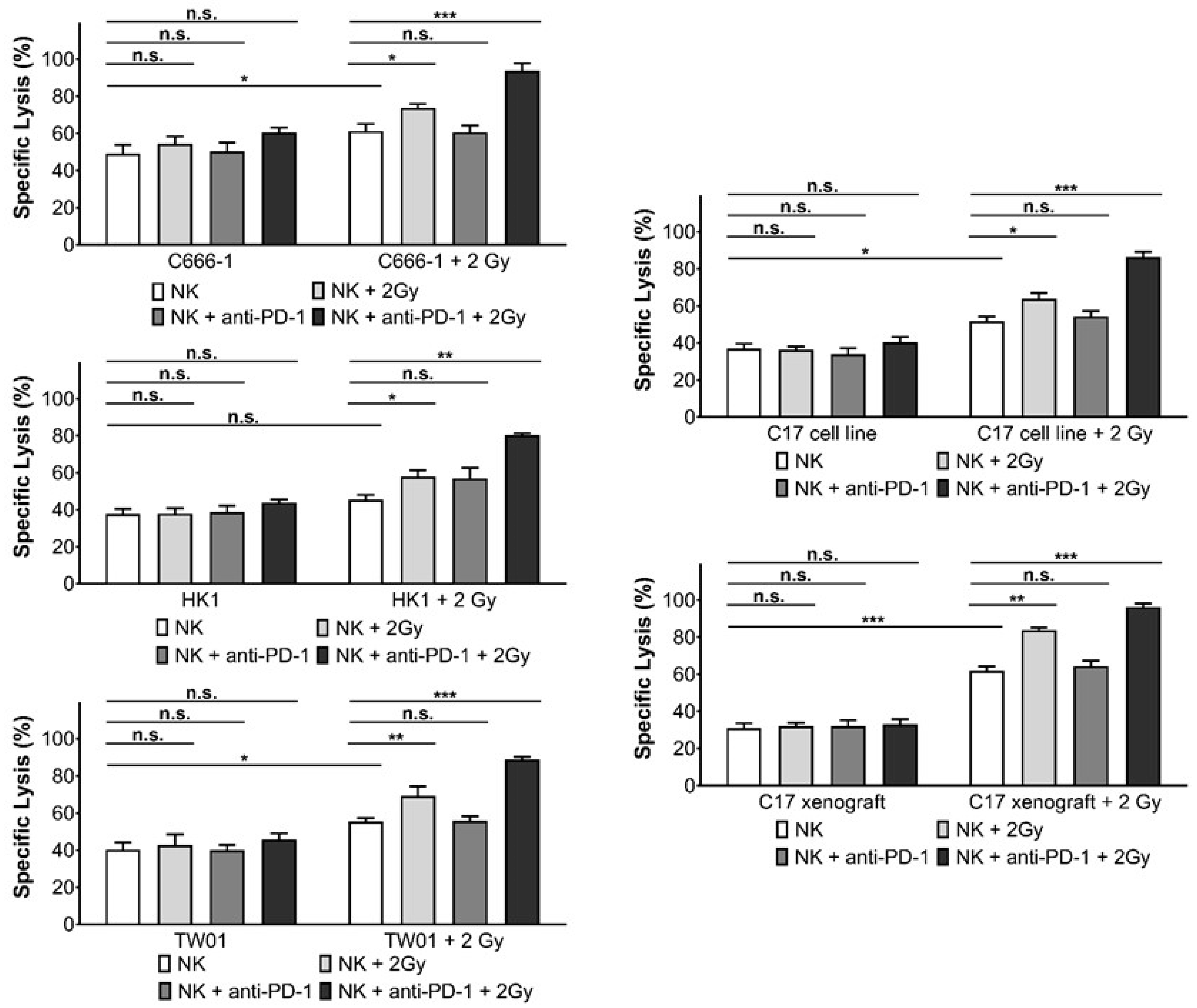
3.3. Inhibition of PD-1 Increases Killing of NPC Cells by Activated NK Cells
3.4. Radiotherapy Induces PD-1 Expression in NK Cells
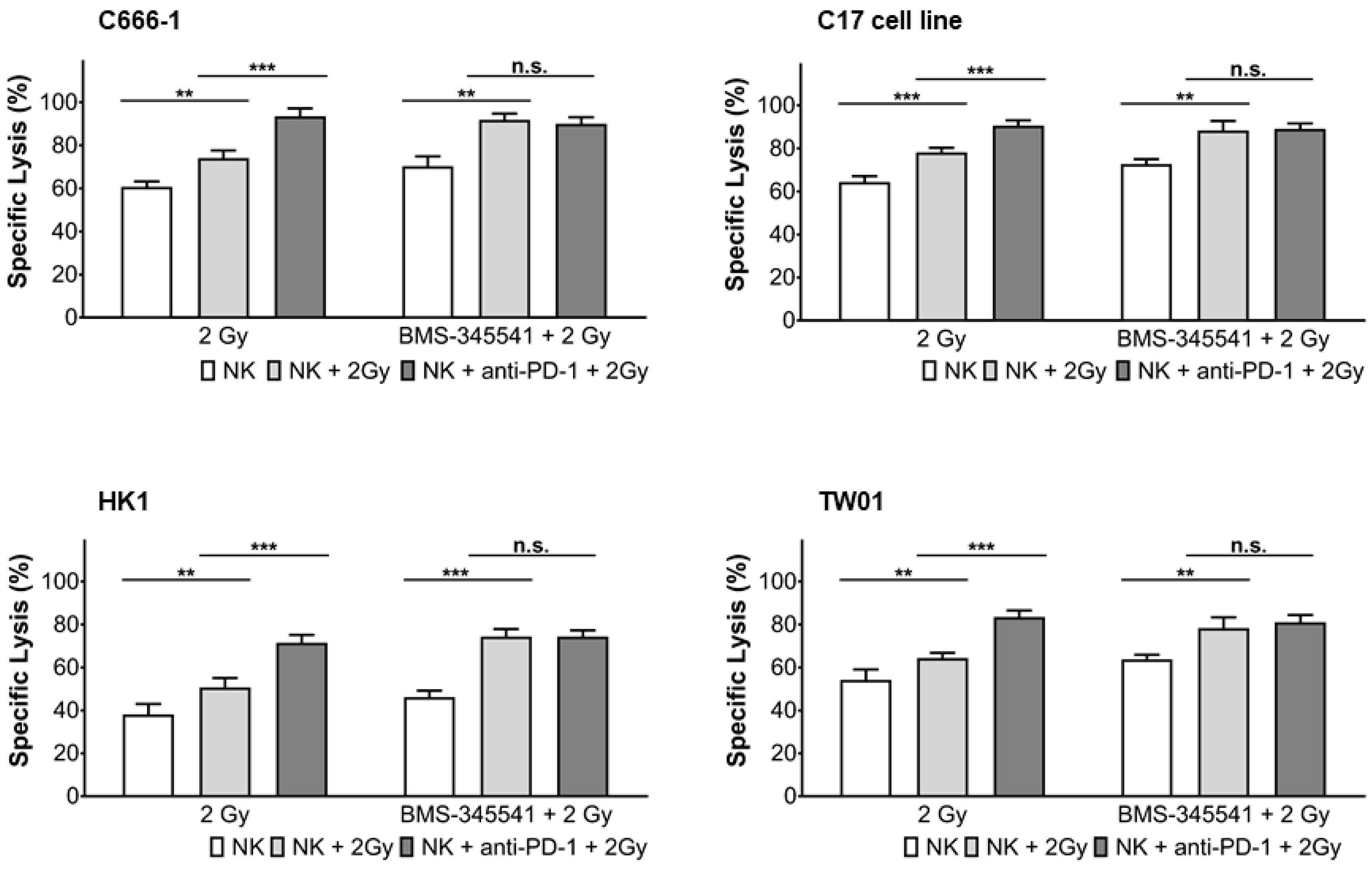
3.5. PD-L1 Expression in NPC Cells and PD-1 Expression in NK Cells by Radiotherapy Is Induced through the NF-κB Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raab-Traub, N. Epstein-Barr virus in the pathogenesis of NPC. Semin. Cancer Biol. 2002, 12, 431–441. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.; Forman, D.; Bray, F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide. Int. J. Cancer 2015, 136, e359–e386. [Google Scholar] [CrossRef]
- Chua, M.; Wee, J.; Hui, E.; Chan, A. Nasopharyngeal carcinoma. Lancet 2016, 387, 1012–1024. [Google Scholar] [CrossRef]
- Wei, W.; Sham, J. Nasopharyngeal carcinoma. Lancet 2005, 365, 41–54. [Google Scholar] [CrossRef]
- Lee, A.; Chan, W.; Sze, H.; Chan, C.; Lam, T. The battle against nasopharyngeal cancer. Radiother. Oncol. 2012, 104, 272–278. [Google Scholar] [CrossRef]
- Rodriguez-Galindo, C.; Wofford, M.; Castleberry, R.; Swanson, G.; London, W.; Fontanesi, J.; Pappo, A.; Douglass, E. Preradiation chemotherapy with methotrexate, cisplatin, 5-fluorouracil, and leucovorin for pediatric nasopharyngeal carcinoma. Cancer 2005, 103, 850–857. [Google Scholar] [CrossRef]
- Buehrlen, M.; Zwaan, C.; Granzen, B.; Lassay, L.; Deutz, P.; Vorwerk, P.; Staatz, G.; Gademann, G.; Christiansen, H.; Oldenburger, F.; et al. Multimodal treatment, including interferon beta, of nasopharyngeal carcinoma in children and young adults: Preliminary results from the prospective, multicenter study NPC-2003-GPOH/DCOG. Cancer 2012, 118, 4892–4900. [Google Scholar] [CrossRef]
- Casanova, M.; Bisogno, G.; Gandola, L.; Cecchetto, G.; Di Cataldo, A.; Basso, E.; Indolfi, P.; D’Angelo, P.; Favini, F.; Collini, P.; et al. Rare Tumors in Pediatric Age Group. A prospective protocol for nasopharyngeal carcinoma in children and adolescents: The Italian Rare Tumors in Pediatric Age (TREP) project. Cancer 2012, 118, 2718–2725. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.; Garden, A.; Ang, K.; O’Daniel, J.; Wang, H.; Court, L.; Morrison, W.; Rosenthal, D.; Chao, K.; Tucker, S.; et al. Quantification of volumetric and geometric changes occuring during fractionated radiotherapy for head and neck cancer using an integrated CT/linear accelerator system. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 960–970. [Google Scholar] [CrossRef]
- Yeh, S.; Tang, Y.; Lui, C.; Huang, Y.; Huang, E. Treatment outcomes and late complications of 849 patients with nasopharyngeal carcinoma treated with radiotherapy alone. Int. J. Radiat. Oncol. Biol. Phys. 2005, 62, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.; Huang, T.; Liao, K.; Chuang, H.; Lin, Y.; Lee, T.; Huang, H.; Fang, F. Impact of late toxicities on quality of life for survivors of nasopharyngeal carcinoma. BMC Cancer 2014, 14, 856. [Google Scholar] [CrossRef] [Green Version]
- Tsukuda, M.; Sawaki, S.; Yanoma, S. Suppressed cellular immunity in patients with nasopharyngeal carcinoma. J. Cancer Res. Clin. Oncol. 1993, 120, 115–118. [Google Scholar] [CrossRef]
- Lakhdar, M.; Oueslati, R.; Ellouze, R.; Thameur, H.; Cammoun, M.; Khedhiri, N.; Kastally, R. High interferon titer and defective NK-cell activity in the circulation of nasopharyngeal carcinoma patients. Int. J. Cancer 1989, 43, 543–548. [Google Scholar] [CrossRef]
- Lu, J.; Chen, X.; Huang, H.; Zhao, F.; Wang, F.; Liu, X.; Li, X. Detailed analysis of inflammatory cell infiltration and the prognostic impact on nasopharyngeal carcinoma. Head Neck 2018, 40, 1245–1253. [Google Scholar] [CrossRef]
- Makowska, A.; Braunschweig, T.; Denecke, B.; Shen, L.; Baloche, V.; Busson, P.; Kontny, U. Interferon β and Anti-PD-1/PD-L1 Checkpoint Blockade Cooperate in NK Cell-Mediated Killing of Nasopharyngeal Carcinoma Cells. Transl. Oncol. 2019, 12, 1237–1256. [Google Scholar] [CrossRef]
- Makowska, A.; Franzen, S.; Braunschweig, T.; Denecke, B.; Shen, L.; Baloche, V.; Busson, P.; Kontny, U. Interferon beta increases NK cell cytotoxicity against tumor cells in patients with nasopharyngeal carcinoma via tumor necrosis factor apoptosis-inducing ligand. Cancer Immunol. Immunother. 2019, 68, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.; Tao, J.; Mengqian, L. Synregistic effect of immunotherapy and radiotherapy in non-small cell lung cancer: Current clinical trials and prospective challenges. Precis. Clin. Med. 2019, 2, 57–70. [Google Scholar]
- Cheung, S.; Huang, D.; Hui, A.; Lo, K.; Ko, C.; Tsang, Y.; Wong, N.; Whitney, B.; Lee, J. Nasopharyngeal carcinoma cell line (C666–1) consistently harbouring Epstein-Barr virus. Int. J. Cancer 1999, 83, 121–126. [Google Scholar] [CrossRef]
- Yip, Y.; Lin, W.; Deng, W.; Jia, L.; Lo, K.; Busson, P.; Vérillaud, B.; Liu, X.; Tsang, C.; Lung, M.; et al. Establishment of a nasopharyngeal carcinoma cell line capable of undergoing lytic Epstein-Barr virus reactivation. Lab. Investig. 2018, 98, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y. Establishment of an epitheloid cell line and a fusiform cell line from a patient with nasopharyngeal carcinoma. Sci. Sin. 1978, 21, 127–134. [Google Scholar]
- Makowska, A.; Wahab, L.; Braunschweig, T.; Kapetanakis, N.; Vokuhl, C.; Denecke, B.; Shen, L.; Busson, P.; Kontny, U. Interferon beta induces apoptosis in nasopharyngeal carcinoma cells via the TRAIL-signaling pathway. Oncotarget 2018, 9, 14228–14250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gressette, M.; Vérillaud, B.; Jimenez-Pailhès, A.; Lelièvre, H.; Lo, K.; Ferrand, F.; Gattolliat, C.; Bescond, A.; Kraus-Berthier, L.; Depil, S.; et al. Treatment of nasopharyngeal carcinoma cells with the histone-deacetylase inhibitor abexinostat: Cooperative effects with cis-platin and radiotherapy on patient-derived xenografts. PLoS ONE 2014, 9, e91325. [Google Scholar]
- Park, B.; Yee, C.; Lee, K. The Effect of Radiation on the Immune Response to Cancers. Int. J. Mol. Sci. 2014, 15, 927–943. [Google Scholar] [CrossRef] [Green Version]
- Makowska, A.; Meier, S.; Shen, L.; Busson, P.; Baloche, V.; Kontny, U. Anti-PD-1 antibody increases NK cell cytotoxicity towards nasopharyngeal carcinoma cells in the context of chemotherapy-induced upregulation of PD-1 and PD-L1. Cancer Immunol. Immunother. 2021, 70, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Dovedi, S.; Adlard, A.; Lipowska-Bhalla, G.; McKenna, C.; Jones, S.; Cheadle, E.; Stratford, I.; Poon, E.; Morrow, M.; Stewart, R.; et al. Acquired Resistance to Fractionated Radiotherapy Can Be Overcome by Concurrent PD-L1 Blockade. Cancer Res. 2014, 74, 5458–5468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Deng, L.; Chmura, S.; Burnette, B.; Liadis, N.; Darga, T.; Beckett, M.; Lingen, M.; Witt, M.; Weichselbaum, R.; et al. Radiation-induced equilibrium is a balance between tumor cell proliferation and T cell-mediated killing. J. Immunol. 2013, 190, 5874–5881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Liang, H.; Burnette, B.; Beckett, M.; Darga, T.; Weichselbaum, R.; Fu, Y. Irradiation and anti–PD-L1 treatment synergistically promote antitumor immunity in mice. J. Clin. Investig. 2014, 124, 687–695. [Google Scholar] [CrossRef]
- Fiorica, F.; Belluomini, L.; Stefanelli, A.; Santini, A.; Urbini, B.; Giorgi, C.; Frassoldati, A. Immune Checkpoint Inhibitor Nivolumab and Radiotherapy in Pretreated Lung Cancer Patients Efficacy and Safety of Combination. Am. J. Clin. Oncol. 2018, 41, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, Y. Targeting NK Cell Checkpoint Receptors or Molecules for Cancer Immunotherapy. Front. Immunol. 2020, 11, 1295. [Google Scholar] [CrossRef]
- Portella, L.; Scala, S. Ionizing radiation effects on the tumor microenvironment. Semin. Oncol. 2019, 46, 254–260. [Google Scholar] [CrossRef]
- Makowska, A.; Eble, M.; Prescher, K.; Hoß, M.; Kontny, U. Chloroquine Sensitizes Nasopharyngeal Carcinoma Cells but Not Nasoepithelial Cells to Irradiation by Blocking Autophagy. PLoS ONE 2016, 11, e0166766. [Google Scholar] [CrossRef] [PubMed]
- Lauber, K.; Ernst, A.; Orth, M.; Herrmann, M.; Belka, C. Dying cell clearance and its impact on the outcome of tumor radiotherapy. Front. Oncol. 2012, 2, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, H.; Paget, J.; Khan, A.; Harrington, K. The tumour microenvironment after radiotherapy: Mechanisms of resistance and recurrence. Nat. Rev. Cancer 2015, 15, 409–425. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.; Huang, J. CD8+ T Cells and NK Cells: Parallel and Complementary Soldiers of Immunotherapy. Curr. Opin. Chem. Eng. 2018, 19, 9–20. [Google Scholar] [CrossRef]
- Caligiuri, M. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Ljunggren, H.; Malmberg, K. Prospects for the use of NK cells in immunotherapy of human cancer. Nat. Rev. Immunol. 2007, 7, 329–339. [Google Scholar] [CrossRef]
- Liu, W.; Chen, G.; Zhang, C.; Liao, X.; Xie, J.; Liang, T.; Liao, W.; Song, L.; Zhang, X. Prognostic significance of tumor-infiltrating lymphocytes and macrophages in nasopharyngeal carcinoma: A systematic review and meta-analysis. Eur. Arch. Otorhinolaryngol. 2021. [CrossRef]
- Zou, Z.; Ha, Y.; Liu, S.; Huang, B. Identification of tumor-infiltrating immune cells and microenvironment-relevant genes in nasopharyngeal carcinoma based on gene expression profiling. Life Sci. 2020, 263, 118620. [Google Scholar] [CrossRef]
- Zhang, L.; MacIsaac, K.; Zhou, T.; Huang, P.; Xin, C.; Dobson, J.; Yu, K.; Chiang, D.; Fan, Y.; Pelletier, M.; et al. Genomic Analysis of Nasopharyngeal Carcinoma Reveals TME-Based Subtypes. Mol. Cancer Res. 2017, 15, 1722–1732. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zeng, X.; Mo, H.; Rolén, U.; Gao, Y.; Zhang, X.; Chen, Q.; Zhang, L.; Zeng, M.; Li, M.; et al. Functional inactivation of EBV-specific T-lymphocytes in nasopharyngeal carci-noma: Implications for tumor immunotherapy. PLoS ONE 2007, 2, e1122. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yin, J.; Li, W.; Li, H.; Chen, D.; Zhang, P.; Lv, J.; Wang, Y.; Li, X.; Li, J.; et al. Single-cell transcriptomics reveals regulators underlying immune cell diversi-ty and immune subtypes associated with prognosis in nasopharyngeal carcinoma. Cell Res. 2020, 30, 1024–1042. [Google Scholar] [CrossRef]
- Liu, H.; Wang, S.; Xin, J.; Wang, J.; Yao, C.; Zhang, Z. Role of NKG2D and its ligands in cancer immunotherapy. Am. J. Cancer Res. 2019, 9, 2064–2078. [Google Scholar] [PubMed]
- Screpanti, V.; Wallin, R.; Ljunggren, H.; Grandien, A. A central role for death receptor-mediated apoptosis in the rejection of tumors by NK cells. J. Immunol. 2001, 167, 2068–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, S.; Singh, T.; Srivastava, R. Ionizing radiation enhances the therapeutic potential of TRAIL in prostate cancer in vitro and in vivo: Intracellular mechanisms. Prostate 2004, 61, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Marini, P.; Schmid, A.; Jendrossek, V.; Faltin, H.; Daniel, P.; Budach, W.; Belka, C. Irradiation specifically sensitises solid tumour cell lines to TRAIL mediated apoptosis. BMC Cancer 2005, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, M.; Abrams, S.; Camphausen, K.; Liu, K.; Scott, T.; Coleman, C.; Hodge, J. Irradiation of tumor cells up-regulates fas and enhances CTL lytic activity and CTL adoptive immunotherapy. J. Immunol. 2003, 170, 6338–6347. [Google Scholar] [CrossRef] [Green Version]
- Cacan, E.; Greer, S.; Garnett-Benson, C. Radiation-induced modulation of immunogenic genes in tumor cells is regulated by both histone deacetylases and DNA methyltransferases. Int. J. Oncol. 2015, 4, 2264–2275. [Google Scholar] [CrossRef]
- Horton, J.; Reihani, S.; Lee, C.; Zhou, Y.; Chen, W.; Geradts, J.; Fels, D.; Hoang, P.; Ashcraft, K.; Groth, J.; et al. FAS death receptor: A breast cancer subtype-specific radiation response biomarker and potential therapeutic target. Radiat. Res. 2015, 184, 456–469. [Google Scholar] [CrossRef]
- Yang, G.; Kong, Q.; Wang, G.; Jin, H.; Zhou, L.; Yu, D.; Niu, C.; Han, W.; Li, W.; Cui, J. Low-dose ionizing radiation induces direct activation of natural killer cells and provides a novel approach for adoptive cellular immunotherapy. Cancer Biother. Radiopharm. 2014, 29, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liu, X.; Zeng, Z.; Li, J.; Luo, Y.; Sun, W.; Gong, Y.; Zhang, J.; Wu, Q.; Xie, C. Immunomodulation of NK Cells by Ionizing Radiation. Front. Oncol. 2020, 10, 874. [Google Scholar] [CrossRef]
- Huang, C.; Bi, E.; Hu, Y.; Deng, W.; Tian, Z.; Dong, C.; Hu, Y.; Sun, B. A novel NF-κB binding site controls human granzyme B gene transcription. J. Immunol. 2006, 176, 4173–4181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Zhang, J.; Lichtenheld, M.; Meadows, G. A Role for NF-κB Activation in Perforin Expression of NK Cells Up-on IL-2 Receptor Signaling. J. Immunol. 2002, 169, 1319–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhry, M.A. Analysis of gene expression in normal and cancer cells exposed to gamma-radiation. J. Biomed. Biotechnol. 2008, 2018, 541678. [Google Scholar]
- Yoneda, K.; Kuwata, T.; Kanayama, M.; Mori, M.; Kawanami, T.; Yatera, K.; Ohguri, T.; Hisaoka, M.; Nakayama, T.; Tanaka, F. Alteration in tumoural PD-L1 expression and stromal CD8-positive tumour-infiltrating lymphocytes after concurrent chemo-radiotherapy for non-small cell lung cancer. Br. J. Cancer 2019, 121, 490–496. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makowska, A.; Lelabi, N.; Nothbaum, C.; Shen, L.; Busson, P.; Tran, T.T.B.; Eble, M.; Kontny, U. Radiotherapy Combined with PD-1 Inhibition Increases NK Cell Cytotoxicity towards Nasopharyngeal Carcinoma Cells. Cells 2021, 10, 2458. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092458
Makowska A, Lelabi N, Nothbaum C, Shen L, Busson P, Tran TTB, Eble M, Kontny U. Radiotherapy Combined with PD-1 Inhibition Increases NK Cell Cytotoxicity towards Nasopharyngeal Carcinoma Cells. Cells. 2021; 10(9):2458. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092458
Chicago/Turabian StyleMakowska, Anna, Nora Lelabi, Christina Nothbaum, Lian Shen, Pierre Busson, Tram Thi Bao Tran, Michael Eble, and Udo Kontny. 2021. "Radiotherapy Combined with PD-1 Inhibition Increases NK Cell Cytotoxicity towards Nasopharyngeal Carcinoma Cells" Cells 10, no. 9: 2458. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092458