
Disproportion in Pericyte/Endothelial Cell Proliferation and Mechanisms of Intussusceptive Angiogenesis Participate in Bizarre Vessel Formation in Glioblastoma

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Tissue Samples
2.2. Light Microscopy
2.3. Immunohistochemistry
2.4. Immunofluorescence in Confocal Microscopy
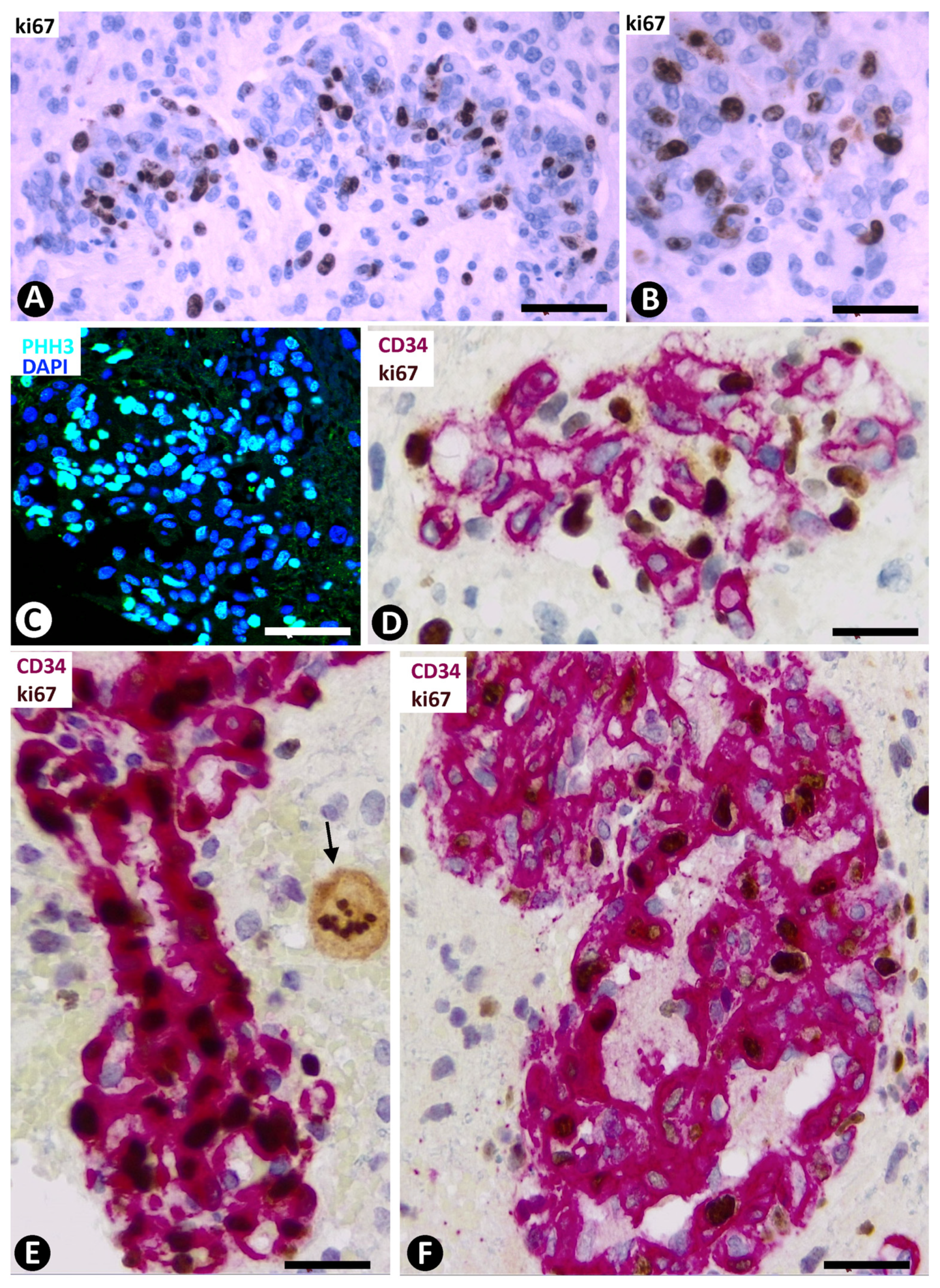
2.5. Evaluation of the PI in the BM of GBM
3. Results
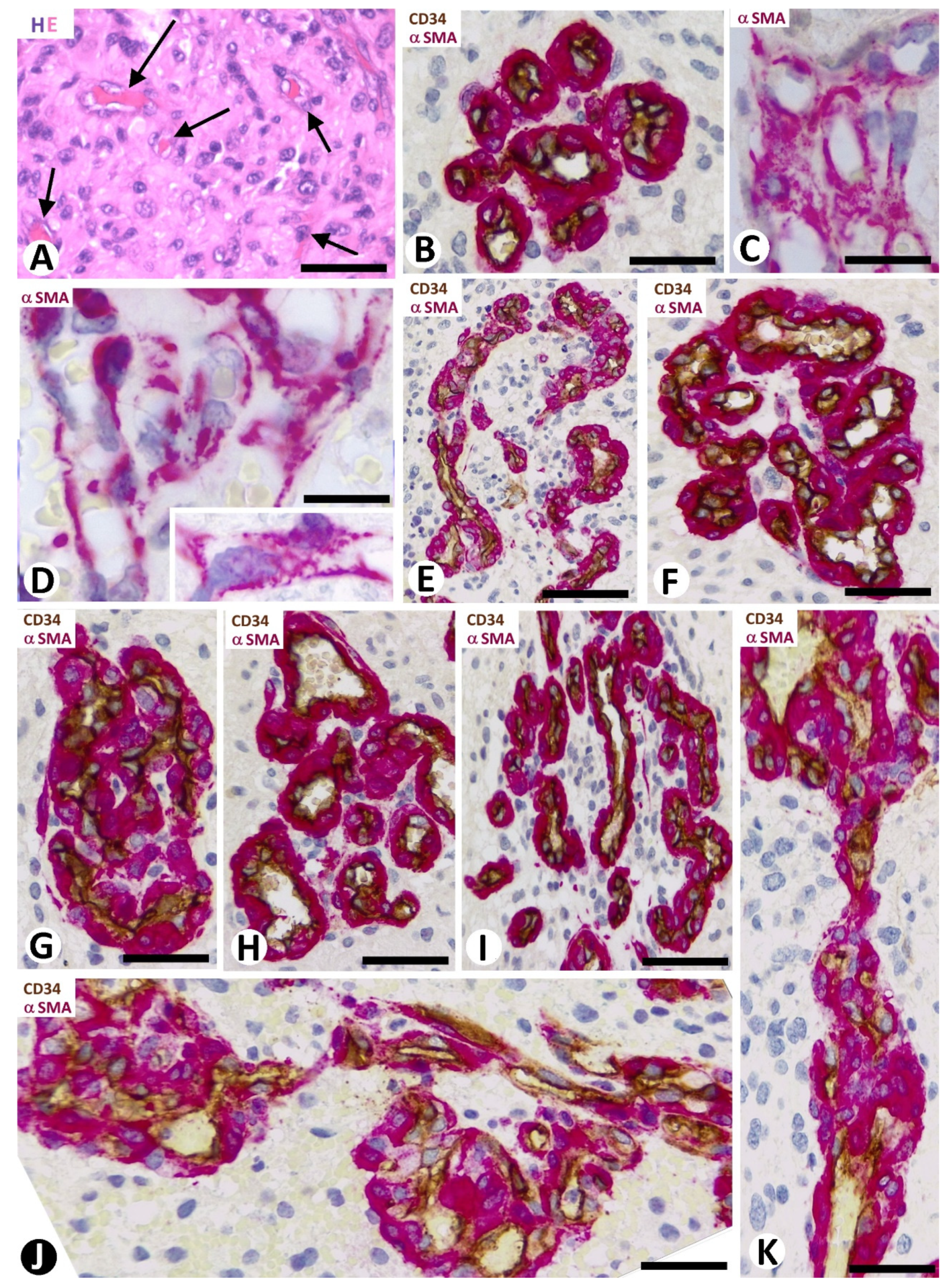
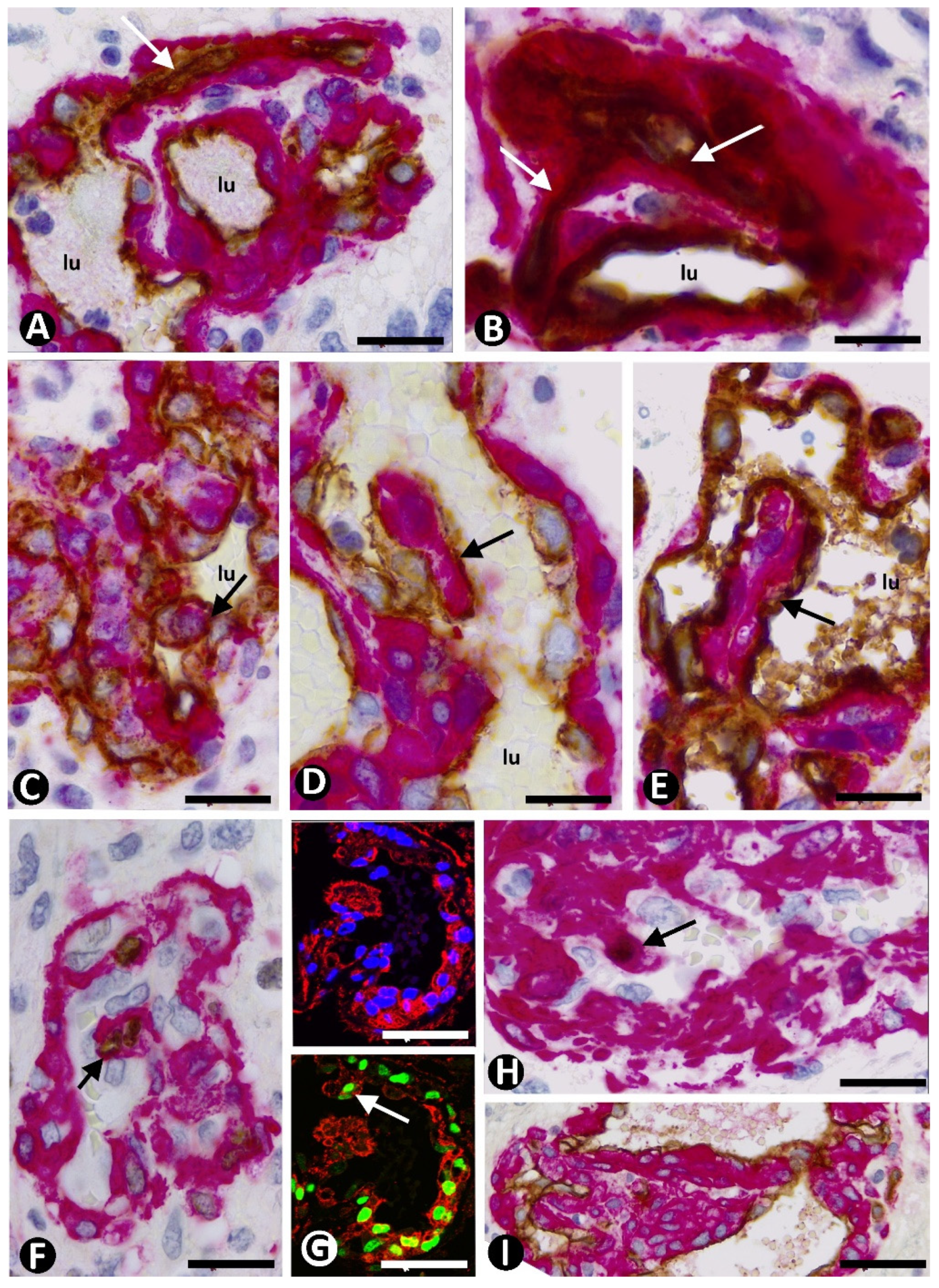
3.1. Microvascular Patterns in GBM
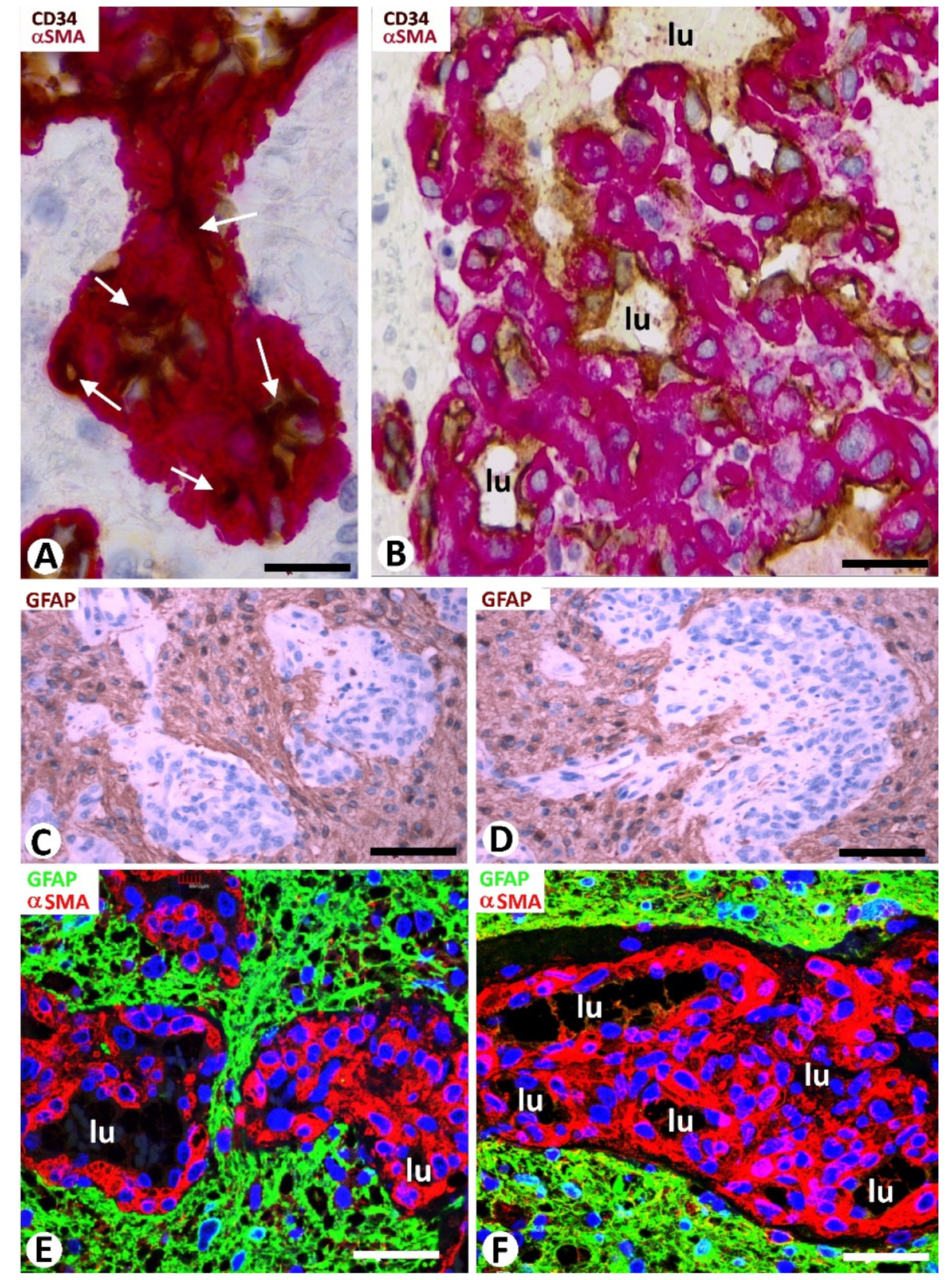
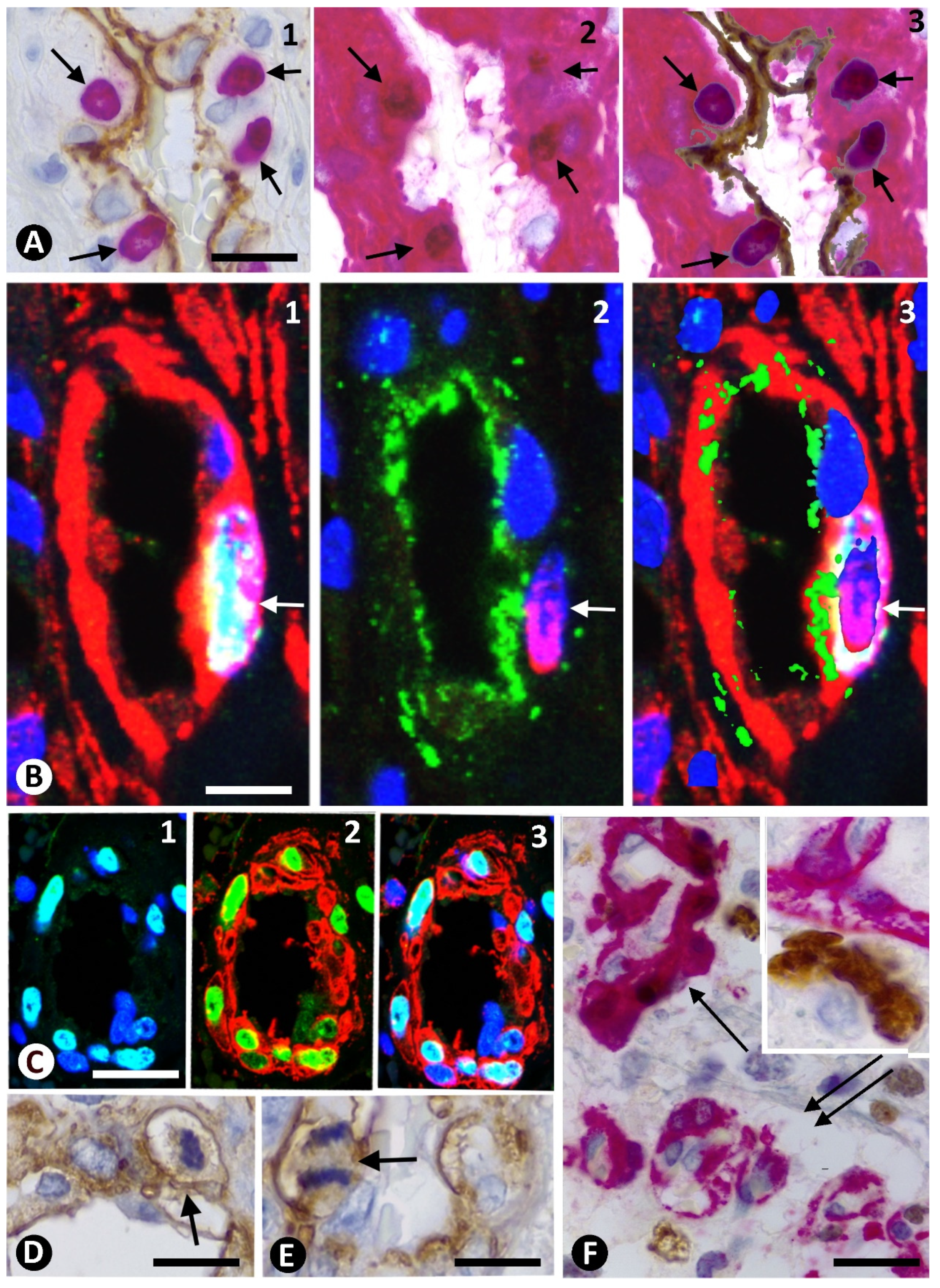
3.2. Cellular Components of Bizarre Vessels in GBM
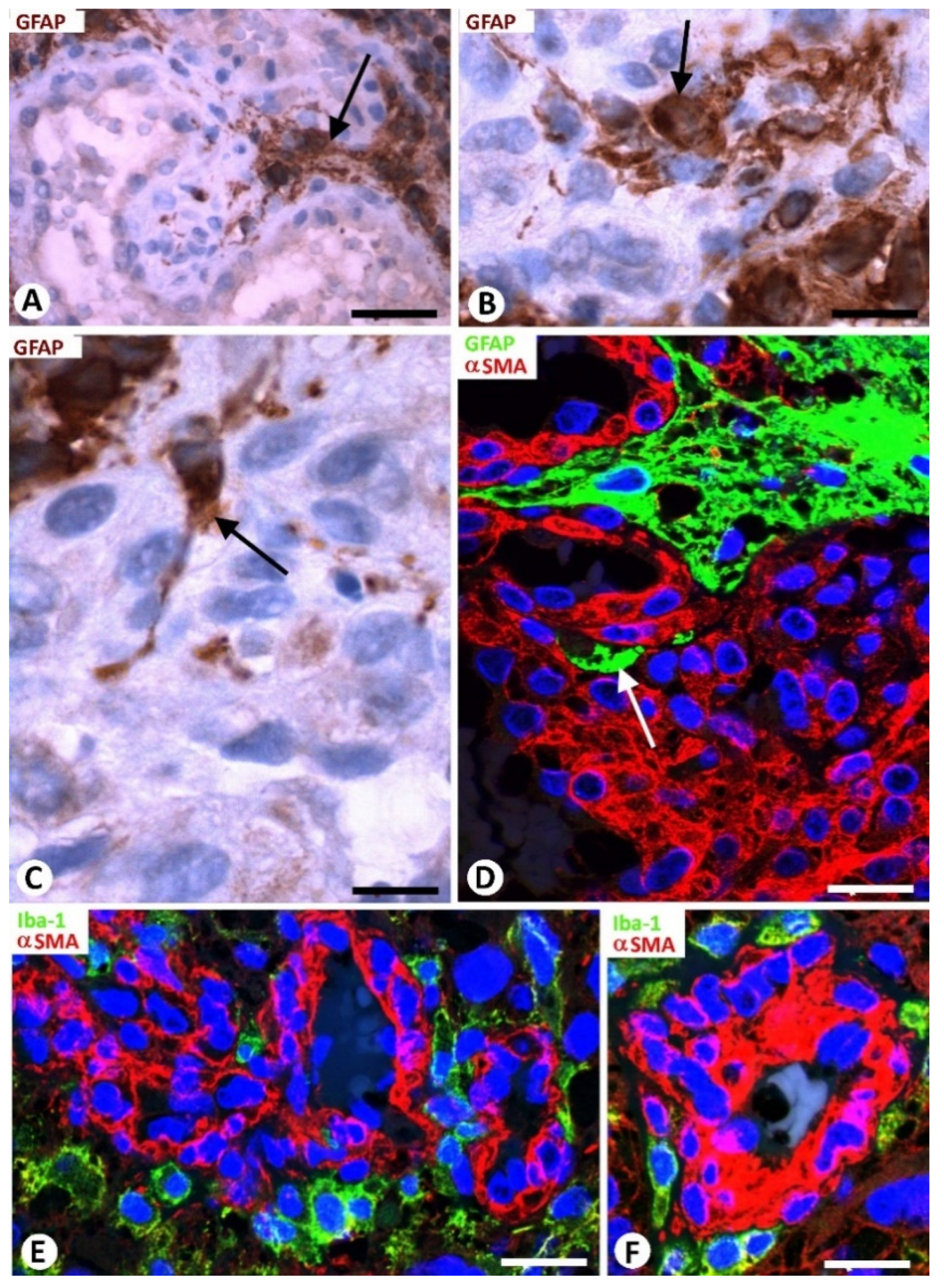
3.3. Neoplastic Glial Cells and BM in GBM
3.4. Microglia and BM in GBM
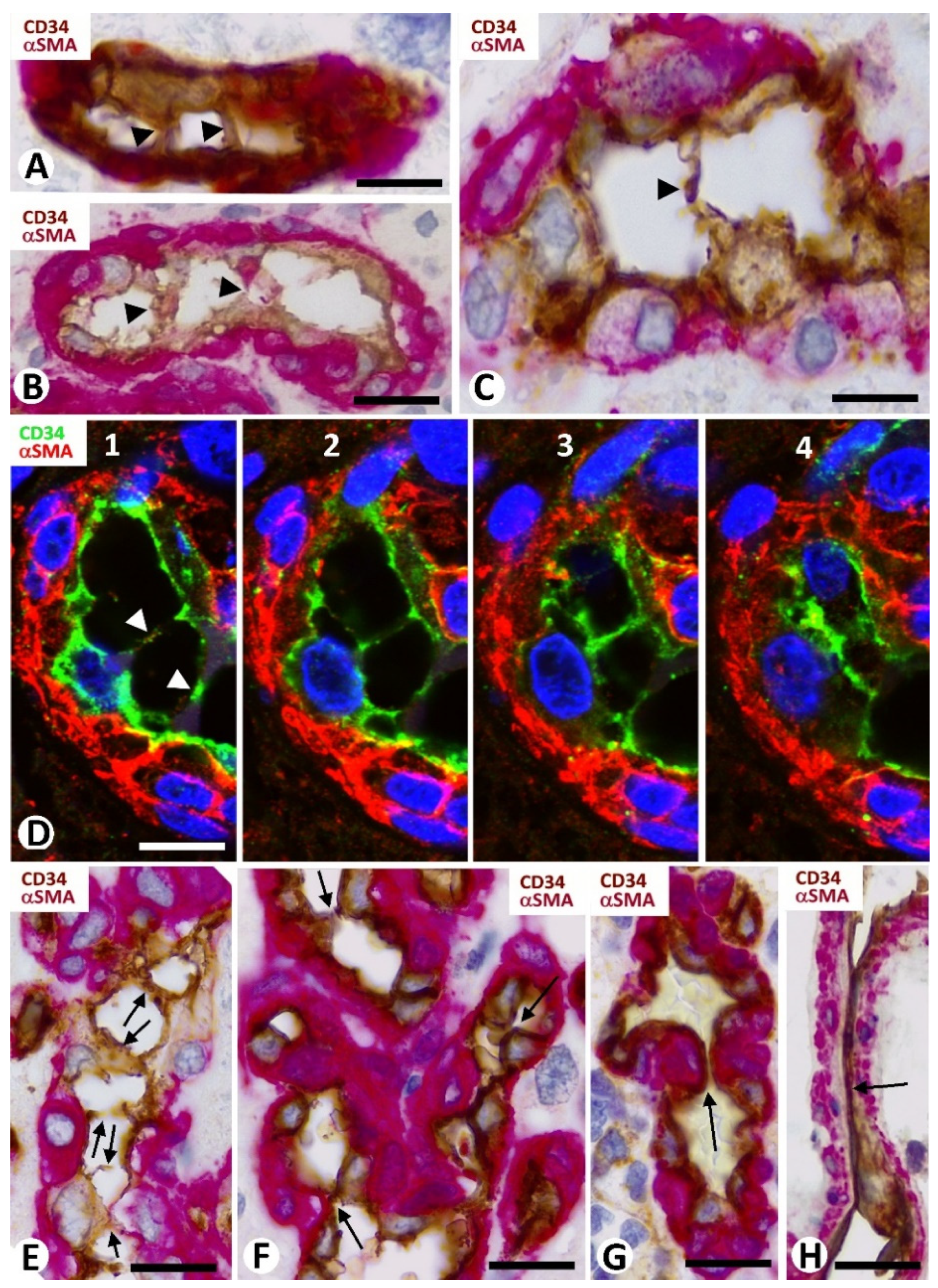
3.5. Pericyte and EC Proliferation in the BM
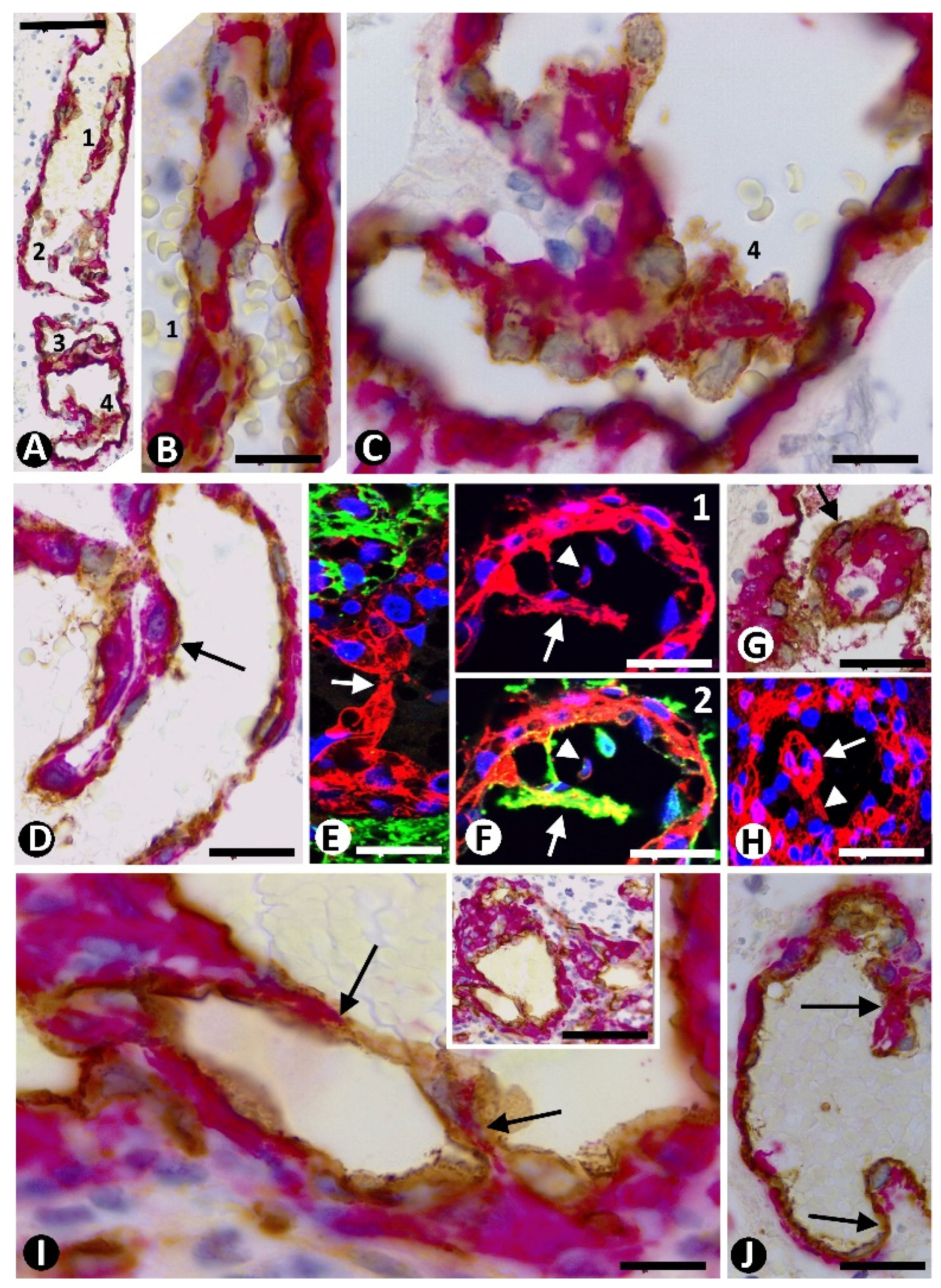
3.6. Intravascular Pillars
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Birner, P.; Piribauer, M.; Fischer, I.; Gatterbauer, B.; Marosi, C.; Ambros, P.F.; Ambros, I.M.; Bredel, M.; Oberhuber, G.; Roessler, K.; et al. Vascular Patterns in Glioblastoma Influence Clinical Outcome and Associate with Variable Expression of Angiogenic Proteins: Evidence for Distinct Angiogenic Subtypes. Brain Pathol. 2006, 13, 133–143. [Google Scholar] [CrossRef]
- Preusser, M.; Heinzl, H.; Gelpi, E.; Schonegger, K.; Haberler, C.; Birner, P.; Marosi, C.; Hegi, M.; Gorlia, T.; Hainfellner, J.A. Histopathologic assessment of Hot-Spot Microvessel Density and Vascular Patterns in Glioblastoma: Poor Observer Agreement Limits Clinical Utility as Prognostic Factors: A Translational Research Project of the European Organization for Research and Treatment of Cancer Brain Tumor Group. Cancer 2006, 107, 162–170. [Google Scholar] [PubMed]
- Chen, L.; Lin, Z.-X.; Lin, G.-S.; Zhou, C.-F.; Chen, Y.-P.; Wang, X.-F.; Zheng, Z.-Q. Classification of Microvascular Patterns via Cluster Analysis Reveals Their Prognostic Significance in Glioblastoma. Hum. Pathol. 2015, 46, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Mao, S.; Li, H.; Zheng, M.; Yi, L.; Lin, J.-M.; Lin, Z.-X. The Pathological Structure of the Perivascular Niche in Different Microvascular Patterns of Glioblastoma. PLoS ONE 2017, 12, e0182183. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Guo, D.; Su, Y.; Yu, D.; Wang, Q.; Wang, T.; Zhou, Q.; Ran, X.; Zou, Z. Hyperplasia of Pericytes Is One of the Main Characteristics of Microvascular Architecture in Malignant Glioma. PLoS ONE 2014, 9, e114246. [Google Scholar] [CrossRef]
- Haddad, S.F.; Moore, S.A.; Schelper, R.L.; Goeken, J.A. Vascular Smooth Muscle Hyperplasia Underlies the Formation of Glo-Meruloid Vascular Structures of Glioblastoma Multiforme. J. Neuropathol. Exp. Neurol. 1992, 51, 488–492. [Google Scholar] [CrossRef]
- Rojiani, A.M.; Dorovini-Zis, K. Glomeruloid Vascular Structures in Glioblastoma Multiforme: An Immunohistochemical and Ultrastructural Study. J. Neurosurg. 1996, 85, 1078–1084. [Google Scholar] [CrossRef]
- Ito, T.; Kitamura, H.; Nakamura, N.; Kameda, Y.; Kanisawa, M. A Comparative Study of Vascular Proliferation in Brain Me-Tastasis of Lung Carcinomas. Virchows Arch. A 1993, 423, 13–17. [Google Scholar] [CrossRef]
- Hardee, M.E.; Zagzag, D. Mechanisms of Glioma-Associated Neovascularization. Am. J. Pathol. 2012, 181, 1126–1141. [Google Scholar] [CrossRef] [Green Version]
- Balaziova, E.; Vymola, P.; Hrabal, P.; Mateu, R.; Zubal, M.; Tomas, R.; Netuka, D.; Kramar, F.; Zemanova, Z.; Svobodova, K.; et al. Fibroblast Activation Protein Expressing Mesenchymal Cells Promote Glioblastoma Angiogen-esis. Cancers 2021, 13, 3304. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Jain, R.K.; Batchelor, T.T. New Directions in Anti-Angiogenic Therapy for Glioblastoma. Neurotherapeutics 2017, 14, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Winkler, F.; Osswald, M.; Wick, W. Anti-Angiogenics: Their Role in the Treatment of Glioblastoma. Oncol. Res. Treat. 2017, 41, 181–186. [Google Scholar] [CrossRef]
- Ameratunga, M.; Pavlakis, N.; Wheeler, H.; Grant, R.; Simes, J.; Khasraw, M. Anti-angiogenic Therapy for High-Grade Glioma. Cochrane Database Syst. Rev. 2018, 11, CD008218. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Tao, W. Identification of Novel Transcriptome Signature as a Potential Prognostic Biomarker for Anti-Angiogenic Therapy in Glioblastoma Multiforme. Cancers 2021, 13, 1013. [Google Scholar] [CrossRef] [PubMed]
- Soda, Y.; Myskiw, C.; Rommel, A.; Verma, I.M. Mechanisms of Neovascularization and Resistance to Anti-Angiogenic Therapies in Glioblastoma Multiforme. J. Mol. Med. 2013, 91, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Rego, G.N.d.; Alves, A.d.; Nucci, M.P.; Mamani, J.B.; De Oliveira, F.A.; Gamarra, L.F. Antiangiogenic Targets for Glioblastoma Therapy from a Pre-Clinical Approach, Using Nanoformulations. Int. J. Mol. Sci. 2020, 21, 4490. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García-Suárez, M.P.; Sáez, F.J.; Gutiérrez, E.; Valladares, F.; Carrasco, J.L.; Díaz-Flores, L., Jr.; Madrid, J.F. Morphofunctional Basis of the Different Types of Angiogenesis and Formation of Postnatal Angiogenesis-Related Secondary Structures. Histol. Histopathol. 2017, 32, 1239–1279. [Google Scholar] [PubMed]
- Nico, B.; Crivellato, E.; Guidolin, D.; Annese, T.; Longo, V.; Finato, N.; Vacca, A.; Ribatti, D. Intussusceptive Microvascular Growth in Human Glioma. Clin. Exp. Med. 2009, 10, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Xuesong, D.; Du, X.; Wu, H.; Liu, H.; Xie, T.; Tong, H.; Chen, X.; Weiguo, Z.; Zhang, W. Aberrant Glioblastoma Neovascularization Patterns and Their Correlation with DCE-MRI-Derived Parameters Following Temozolomide and Bevacizumab Treatment. Sci. Rep. 2017, 7, 1–10. [Google Scholar]
- Caduff, J.H.; Fischer, L.C.; Burri, P.H. Scanning Electron Microscope Study of the Developing Microvasculature in the Postnatal Rat Lung. Anat. Rec.. 1986, 216, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Burri, P.H.; Djonov, V. Intussusceptive Angiogenesis-The Alternative to Capillary Sprouting. Mol. Aspects Med. 2002, 23, 1–27. [Google Scholar] [CrossRef]
- Burri, P.H.; Hlushchuk, R.; Djonov, V. Intussusceptive angiogenesis: Its emergence, its characteristics, and its significance. Dev. Dyn. 2004, 231, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Carrasco, J.L.; Madrid, J.F. Seg-mentation of Dilated Hemorrhoidal Veins in Hemorrhoidal Disease. Cells Tissues Organs 2018, 205, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Djonov, V.G.; Kurz, H.; Burri, P.H. Optimality in the Developing Vascular System: Branching Remodeling by Means of Intus-Susception as an Efficient Adaptation Mechanism. Dev. Dyn. 2002, 224, 391–402. [Google Scholar] [CrossRef]
- Patan, S.; Munn, L.L.; Tanda, S.; Roberge, S.; Jain, R.K.; Jones, R.C. Vascular Morphogenesis and Remodeling in a Model of Tissue Repair: Blood Vessel Formation and Growth in the Ovarian Pedicle After Ovariectomy. Circ. Res. 2001, 89, 723–731. [Google Scholar] [CrossRef]
- Patan, S.; Tanda, S.; Roberge, S.; Jones, R.C.; Jain, R.K.; Munn, L.L. Vascular Morphogenesis and Remodeling in a Human Tumor Xenograft: Blood Vessel Formation and Growth after Ovariectomy and Tumor Implantation. Circ. Res. 2001, 89, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Paku, S.; Dezső, K.; Bugyik, E.; Tóvári, J.; Tímár, J.; Nagy, P.; Laszlo, V.; Klepetko, W.; Dome, B. A New Mechanism for Pillar Formation during Tumor-Induced Intussusceptive Angiogenesis: Inverse Sprouting. Am. J. Pathol. 2011, 179, 1573–1585. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; Gayoso, S.; García, M.P.; González-Gómez, M.; Díaz-Flores, L., Jr.; Sánchez, R.; Carrasco, J.L.; Madrid, J.F. Intussusceptive Angiogenesis and Its Counterpart Intussusceptive Lymphangiogenesis. Histol. Histopathol. 2020, 35, 1083–1103. [Google Scholar]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.A.P.; Carrasco, J.L.; Díaz-Flores, L., Jr.; Madrid, J.F.; Álvarez-Argüelles, H. Participation of Intussusceptive Angiogenesis in the Morphogenesis of Lobular Capillary Hemangioma. Sci. Rep. 2020, 10, 4987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlatter, P.; König, M.F.; Karlsson, L.M.; Burri, P.H. Quantitative Study of Intussusceptive Capillary Growth in the Chori-Oallantoic Membrane (CAM) of the Chicken Embryo. Microvasc. Res. 1997, 54, 65–73. [Google Scholar] [CrossRef]
- Djonov, V.; Schmid, M.; Tschanz, S.A.; Burri, P.H. Intussusceptive Angiogenesis: Its Role in Embryonic Vascular Network For-Mation. Circ. Res. 2000, 86, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Kurz, H.; Burri, P.H.; Djonov, V.G. Angiogenesis and Vascular Remodeling by Intussusception: From Form to Function. Physiology 2003, 18, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, A.; Özen, I.; Genové, G.; Paul, G.; Bengzon, J. Endogenous Brain Pericytes Are Widely Activated and Contribute to Mouse Glioma Microvasculature. PLoS ONE 2015, 10, e0123553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohannon, D.; Long, D.; Kim, W.-K. Understanding the Heterogeneity of Human Pericyte Subsets in Blood–Brain Barrier Homeostasis and Neurological Diseases. Cells 2021, 10, 890. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; Madrid, J.F.; Varela, H.; Valladares, F.; Acosta, E.; Martín-Vasallo, P.; Díaz-Flores, L., Jr. Pericytes. Morphofunction, Interactions and Pathology in a Quiescent and Activated Mesenchymal Cell Niche. Histol. Histopathol. 2009, 24, 909–969. [Google Scholar]
- Wesseling, P.; Schlingemann, R.O.; Rietveld, F.J.; Link, M.; Burger, P.C.; Ruiter, D.J. Early and Extensive Contribution of Peri-Cytes/Vascular Smooth Muscle Cells to Microvascular Proliferation in Glioblastoma Multiforme: An Immuno-Light and Immu-No-Electron Microscopic Study. J. Neuropathol. Exp. Neurol. 1995, 54, 304–310. [Google Scholar] [CrossRef]
- Takeuchi, H.; Hashimoto, N.; Kitai, R.; Kubota, T.; Kikuta, K.-I. Proliferation of Vascular Smooth Muscle Cells in Glioblastoma Multiforme. J. Neurosurg. 2010, 113, 218–224. [Google Scholar] [CrossRef]
- Eberhard, A.; Kahlert, S.; Goede, V.; Hemmerlein, B.; Plate, K.H.; Augustin, H. Heterogeneity of Angiogenesis and Blood Vessel Maturation in Human Tumors: Implications for Antiangiogenic Tumor Therapies. Cancer Res. 2000, 60, 1388–1393. [Google Scholar]
- Gee, M.S.; Procopio, W.N.; Makonnen, S.; Feldman, M.D.; Yeilding, N.M.; Lee, W.M. Tumor Vessel Development and Maturation Impose Limits on the Effectiveness of Anti-Vascular Therapy. Am. J. Pathol. 2003, 162, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Sena, I.F.G.; Paiva, A.E.; Prazeres, P.H.D.M.; Azevedo, P.O.; Lousado, L.; Bhutia, S.K.; Salmina, A.B.; Mintz, A.; Birbrair, A. Glioblastoma-Activated Pericytes Support Tumor Growth via Immunosuppression. Cancer Med. 2018, 7, 1232–1239. [Google Scholar] [CrossRef]
- Zhang, X.-N.; Yang, K.-D.; Chen, C.; He, Z.-C.; Wang, Q.-H.; Feng, H.; Lv, S.-Q.; Wang, Y.; Mao, M.; Liu, Q.; et al. Pericytes Augment Glioblastoma Cell Resistance to Temozolomide through CCL5-CCR5 Paracrine Signaling. Cell Res. 2021, 1–16. [Google Scholar] [CrossRef]
- Sattiraju, A.; Mintz, A. Pericytes in Glioblastomas: Multifaceted Role within Tumor Microenvironments and Potential for Therapeutic Interventions. Adv. Exp. Med. Biol. 2019, 1147, 65–91. [Google Scholar]
- Valdor, R.; García-Bernal, D.; Riquelme, D.; Martinez, C.M.; Moraleda, J.M.; Cuervo, A.M.; Macian, F.; Martinez, S. Glio-Blastoma Ablates Pericytes Antitumor Immune Function through Aberrant Up-Regulation of Chaperone-Mediated Autophagy. Proc. Natl. Acad. Sci. USA 2019, 116, 20655–20665. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.; ElAli, A.; Virgintino, D.; Gilbert, M.R. Blood-Brain Barrier Pericyte Importance in Malignant Gliomas: What We Can Learn from Stroke and Alzheimer’s Disease. Neuro-Oncology 2017, 19, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Chen, C.; Shi, Y.; Wu, Q.; Gimple, R.C.; Fang, X.; Huang, Z.; Zhai, K.; Ke, S.Q.; Ping, Y.-F.; et al. Targeting Glioma Stem Cell-Derived Pericytes Disrupts the Blood-Tumor Barrier and Improves Chemotherapeutic Efficacy. Cell Stem Cell 2017, 21, 591–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, W.-K.; Yotsumoto, F.; Sakimura, K.; Adams, R.H.; Stallcup, W.B. NG2 Proteoglycan Promotes Tumor Vascularization via Integrin-Dependent Effects on Pericyte Function. Angiogenesis 2013, 17, 61–76. [Google Scholar] [CrossRef] [Green Version]
- Al-Mayhani, T.F.; Heywood, R.M.; Vemireddy, V.; Lathia, J.D.; Piccirillo, S.G.M.; Watts, C. A Non-hierarchical Organization of Tumorigenic NG2 Cells in Glioblastoma Promoted by EGFR. Neuro-Oncology 2018, 21, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Finocchiaro, G.; Pellegatta, S. NG2/CSPG4 in Glioblastoma: About Flexibility. Neuro-Oncology 2019, 21, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Krstulja, M.; Bonifačić, D.; Cvejanović, S. Proliferation of Endothelial Cells in Neoangiogenesis of Human Cortical Glioblastoma. Open Life Sci. 2008, 3, 113–119. [Google Scholar] [CrossRef]
- Burri, P.H.; Tarek, M.R. A Novel Mechanism of Capillary Growth in the Rat Pulmonary Microcirculation. Anat. Rec. 1990, 228, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Patan, S. Lycat and Cloche at the Switch between Blood Vessel Growth and Differentiation? Circ. Res. 2008, 102, 1005–1007. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.P.; Carrasco, J.L.; Díaz-Flores, L., Jr.; Madrid, J.F. Myriad Pillars Formed by Intussusceptive Angiogenesis as the Basis of Intravascular Papillary Endothelial Hyperplasia (IPEH). IPEH Is Intus-Susceptive Angiogenesis Made a Lesion. Histol. Histopathol. 2021, 36, 217–228. [Google Scholar]
- Dalemans, D.; Berendsen, S.; Draaisma, K.; Robe, P.; Snijders, T. Glioblastomas within the Subventricular Zone Are Region-Specific Enriched for Mesenchymal Transition Markers: An Intratumoral Gene Expression Analysis. Cancers 2021, 13, 3764. [Google Scholar] [CrossRef]
- Berendsen, S.; Van Bodegraven, E.; Seute, T.; Spliet, W.G.M.; Geurts, M.; Hendrikse, J.; Schoysman, L.; Huiszoon, W.B.; Varkila, M.; Rouss, S.; et al. Adverse Prognosis of Glioblastoma Contacting the Subventricular Zone: Biological Correlates. PLoS ONE 2019, 14, e0222717. [Google Scholar] [CrossRef] [Green Version]
- Arcella, A.; Sanchez, M. Natural Substances to Potentiate Canonical Glioblastoma Chemotherapy. J. Chemother. 2021, 33, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Goenka, A.; Tiek, D.; Song, X.; Huang, T.; Hu, B.; Cheng, S.-Y. The Many Facets of Therapy Resistance and Tumor Recurrence in Glioblastoma. Cells 2021, 10, 484. [Google Scholar] [CrossRef]
- Birzu, C.; French, P.; Caccese, M.; Cerretti, G.; Idbaih, A.; Zagonel, V.; Lombardi, G. Recurrent Glioblastoma: From Molecular Landscape to New Treatment Perspectives. Cancers 2020, 13, 47. [Google Scholar] [CrossRef]
- Lombardi, G.; Idbaih, A.; Le Rhun, E.; Preusser, M.; Zagonel, V.; French, P. A New Landscape for Systemic Pharmacotherapy of Recurrent Glioblastoma? Cancers 2020, 12, 3775. [Google Scholar] [CrossRef] [PubMed]
- Hlushchuk, R.; Riesterer, O.; Baum, O.; Wood, J.; Gruber, G.; Pruschy, M.; Djonov, V. Tumor Recovery by Angiogenic Switch from Sprouting to Intussusceptive Angiogenesis after Treatment with PTK787/ZK222584 or Ionizing Radiation. Am. J. Pathol. 2008, 173, 1173–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
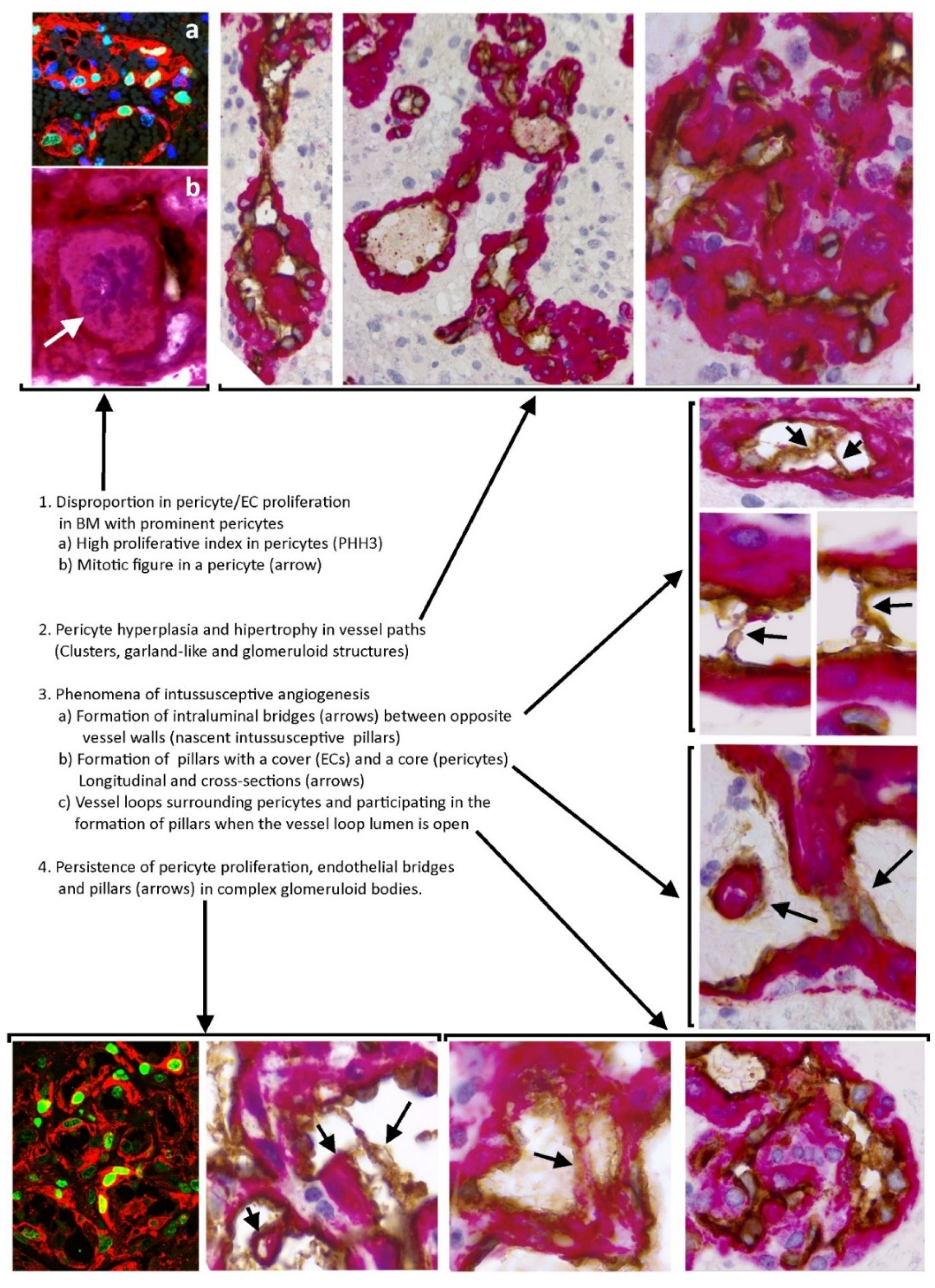{kind=link}
| Type I BM | Type II BM | |
|---|---|---|
| Overall PI | Mean 43.57 ± 9.5 | Mean 3.41 ± 0.76 |
| Pericyte PI | Mean 42.11 ± 5.76 | Mean 1.12 ± 0.21 |
| EC PI | Mean 0.88 ± 0.29 | Mean 2.21 ± 0.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.-d.-P.; Díaz-Flores, L., Jr.; González-Marrero, I.; Ávila, J.; Martín-Vasallo, P. Disproportion in Pericyte/Endothelial Cell Proliferation and Mechanisms of Intussusceptive Angiogenesis Participate in Bizarre Vessel Formation in Glioblastoma. Cells 2021, 10, 2625. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102625
Díaz-Flores L, Gutiérrez R, González-Gómez M, García M-d-P, Díaz-Flores L Jr., González-Marrero I, Ávila J, Martín-Vasallo P. Disproportion in Pericyte/Endothelial Cell Proliferation and Mechanisms of Intussusceptive Angiogenesis Participate in Bizarre Vessel Formation in Glioblastoma. Cells. 2021; 10(10):2625. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102625
Chicago/Turabian StyleDíaz-Flores, Lucio, Ricardo Gutiérrez, Miriam González-Gómez, María-del-Pino García, Lucio Díaz-Flores, Jr., Ibrahim González-Marrero, Julio Ávila, and Pablo Martín-Vasallo. 2021. "Disproportion in Pericyte/Endothelial Cell Proliferation and Mechanisms of Intussusceptive Angiogenesis Participate in Bizarre Vessel Formation in Glioblastoma" Cells 10, no. 10: 2625. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102625