
Evaluation of Immunomodulatory Responses and Changed Wound Healing in Type 2 Diabetes—A Study Exploiting Dermal Fibroblasts from Diabetic and Non-Diabetic Human Donors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Individuals
2.3. Isolation of Dermal Fibroblasts
2.4. Nuclear Count
2.5. Detection of Cell Proliferation by BrdU Assay
2.6. Reactive Oxygen Species
2.7. Glucose Uptake Assay
2.8. Insulin Receptor Detection
2.9. Detection of IL-6 in Supernatants of Dermal Fibroblasts
2.10. MIF Detection in Supernatants of Dermal Fibroblasts
2.11. MG-H1 Assay
2.12. Semi-Quantitative Real-Time PCR
2.13. Glyoxalase I Protein Expression
2.14. Glyoxalase I Activity
2.15. Procollagen Type I C-Peptide ELISA
2.16. Statistical Analysis
3. Results
3.1. Characteristics of the Donors
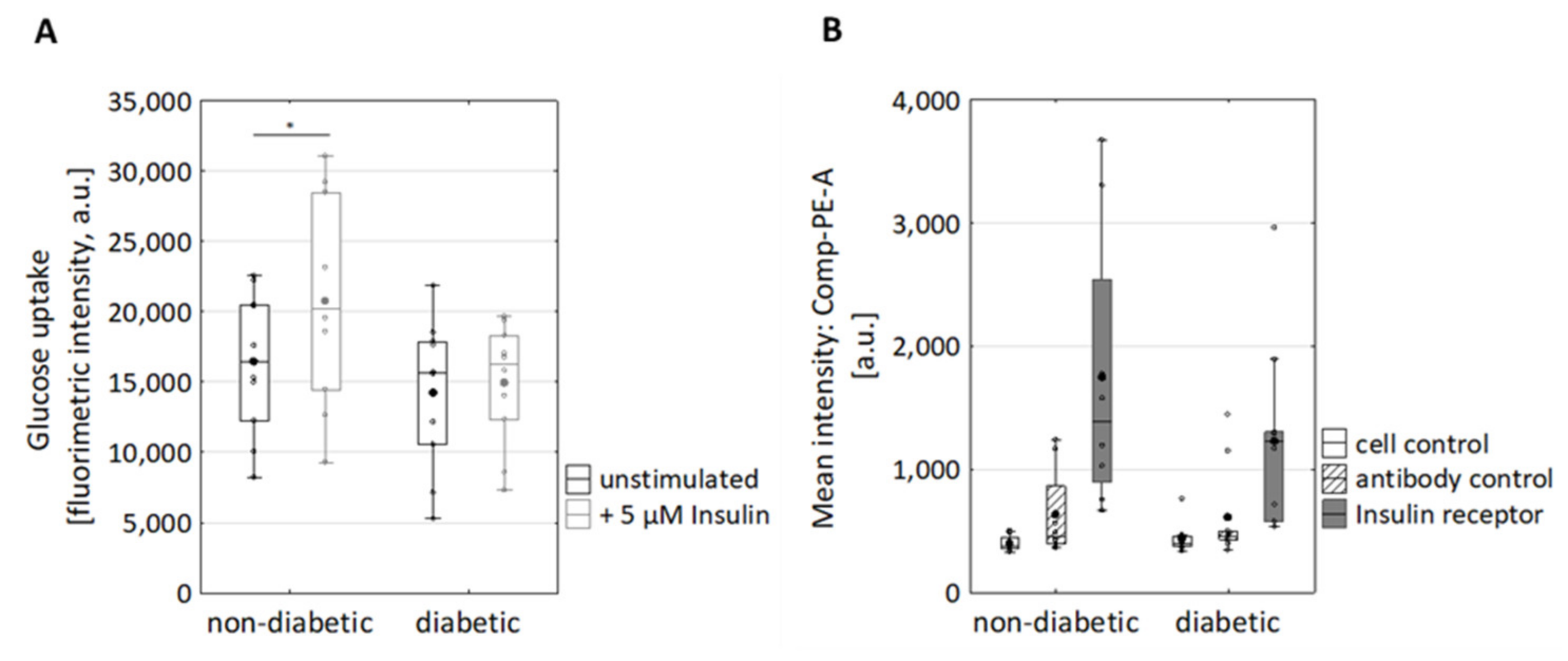
3.2. Reduced Insulin-Induced Glucose Uptake and Reduced Expression of the Insulin Receptor in Dermal Fibroblasts of Diabetic Donors
3.3. Changed Secretion of IL-6, Macrophage Migration Inhibitory Factor, and Procollagen Type I C-Peptide in Fibroblasts of Diabetic Donors
3.4. Effects of MGO Treatment on the Secretion of IL-6, Macrophage Migration Inhibitory Factor (MIF) and Procollagen Type I C-Peptide in Dermal Fibroblasts
3.5. MIF Treatment Results in Reduced Glyoxalase I Activity in Diabetic Fibroblasts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, E.; Raghavan, S. Getting under the skin of epidermal morphogenesis. Nat. Rev. Genet. 2002, 3, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, P. Dermatologie Und Venerologie Für Das Studium; Springer: Berlin, Germany, 2009. [Google Scholar]
- Kanitakis, J. Anatomy, histology and immunohistochemistry of normal human skin. Eur. J. Dermatol. 2002, 12, 390–401. [Google Scholar]
- Tobin, D.J. Biochemistry of human skin—Our brain on the outside. Chem. Soc. Rev. 2006, 35, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Urmacher, C. Histology of Normal Skin. Am. J. Surg. Pathol. 1990, 14, 671–686. [Google Scholar] [CrossRef]
- Saijo, Y.; Ida, T.; Iwazaki, H.; Shintate, R.; Sato, K.; Hiratsuka, T.; Yoshizawa, S.; Umemura, S.-I.; Miyajima, J.; Tang, H. Visualization of skin morphology and microcirculation with high frequency ultrasound and dual-wavelength photoacoustic microscope. In Proceedings of the Photons Plus Ultrasound: Imaging and Sensing 2019, San Francisco, CA, USA, 27 February 2019. [Google Scholar]
- Prost-Squarcioni, C.; Fraitag, S.; Heller, M.; Boehm, N. Histologie fonctionnelle du derme. Ann. Dermatol. Venereol. 2008, 135, 5–20. [Google Scholar] [CrossRef]
- Stunova, A.; Vistejnova, L. Dermal fibroblasts—A heterogeneous population with regulatory function in wound healing. Cytokine Growth Factor Rev. 2018, 39, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal fibroblast in cutaneous development and healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e307. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J.; Kivirikko, K.I. COLLAGENS: Molecular Biology, Diseases, and Potentials for Therapy. Annu. Rev. Biochem. 1995, 64, 403–434. [Google Scholar] [CrossRef] [PubMed]
- Francini, G.; Gonnelli, S.; Petrioli, R.; Bruni, S.; Marsili, S.; Aquino, A.; Camporeale, A. Procollagen type I carboxy-terminal pro-peptide as a marker of osteoblastic bone metastases. Cancer Epidemiol. Biomark. Prev. 1993, 2, 125–129. [Google Scholar]
- International Diabetes Federation. Diabetes Atlas 9th Edition. Available online: https://www.idf.org/e-library/epidemiology-research/diabetes-atlas/159-idf-diabetes-atlas-ninth-edition-2019.html (accessed on 16 August 2021).
- Van Belle, T.L.; Coppieters, K.T.; Von Herrath, M.G. Type 1 Diabetes: Etiology, Immunology, and Therapeutic Strategies. Physiol. Rev. 2011, 91, 79–118. [Google Scholar] [CrossRef]
- Behm, B.; Schreml, S.; Landthaler, M.; Babilas, P. Skin signs in diabetes mellitus. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.I.; Kohn, S.R. Cutaneous manifestations of diabetes mellitus. J. Am. Acad. Dermatol. 1994, 30, 519–531. [Google Scholar] [CrossRef]
- Wahid, Z.; Kanjee, A. Cutaneous manifestations of diabetes mellitus. J. Pak. Med. Assoc. 1998, 48, 304–305. [Google Scholar] [PubMed]
- Meurer, M.; Stumvoll, M.; Szeimies, R.-M. Hautveränderungen bei Diabetes mellitus. Der Hautarzt 2004, 55, 428–435. [Google Scholar] [CrossRef]
- Van Hattem, S.; Bootsma, A.H.; Thio, H.B. Skin manifestations of diabetes. Clevel. Clin. J. Med. 2008, 75, 772–787. [Google Scholar] [CrossRef] [PubMed]
- Allaman, I.; Bélanger, M.; Magistretti, P.J. Methylglyoxal, the dark side of glycolysis. Front. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, R.; Yan, S.F.; Schmidt, A.M. Receptor for AGE (RAGE): Signaling mechanisms in the pathogenesis of diabetes and its complications. Ann. N. Y. Acad. Sci. 2011, 1243, 88–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bierhaus, A.; Fleming, T.; Stoyanov, S.; Leffler, A.; Babes, A.; Neacsu, C.; Sauer, S.K.; Eberhardt, M.; Schnölzer, M.; Lasitschka, F.; et al. Methylglyoxal modification of Nav1.8 facilitates nociceptive neuron firing and causes hyperalgesia in diabetic neuropathy. Nat. Med. 2012, 18, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Roosterman, D.; Goerge, T.; Schneider, S.W.; Bunnett, N.W.; Steinhoff, M. Neuronal Control of Skin Function: The Skin as a Neuro-immunoendocrine Organ. Physiol. Rev. 2006, 86, 1309–1379. [Google Scholar] [CrossRef]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Jack, M.; Wright, D. Role of advanced glycation endproducts and glyoxalase I in diabetic peripheral sensory neuropathy. Transl. Res. 2012, 159, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Thornalley, P.J.; Dawczynski, J.; Franke, S.; Strobel, J.; Stein, G.; Haik, G.M. Methylglyoxal-Derived Hydroimidazolone Advanced Glycation End-Products of Human Lens Proteins. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5287–5292. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.; Akash, M.S.H.; Liaqat, A.; Kamal, S.; Qadir, M.I.; Rasul, A. Role of Interleukin-6 in Development of Insulin Resistance and Type 2 Diabetes Mellitus. Crit. Rev. Eukaryot. Gene Expr. 2017, 27, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.U.; Lee, W.-Y.; Kim, W.-S.; Lee, Y.-T.; Yoon, K.J. Expression of macrophage migration inhibitory factor in footpad skin lesions with diabetic neuropathy. Mol. Pain 2018, 14, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Zamora, Y.I.; Rodriguez-Sosa, M. The Role of MIF in Type 1 and Type 2 Diabetes Mellitus. J. Diabetes Res. 2014, 2014, 804519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubetsky, J.B.; Dios, A.; Han, J.; Aljabari, B.; Ruzsicska, B.; Mitchell, R.; Lolis, E.; Al-Abed, Y. The Tautomerase Active Site of Macrophage Migration Inhibitory Factor Is a Potential Target for Discovery of Novel Anti-inflammatory Agents. J. Biol. Chem. 2002, 277, 24976–24982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Brodsky, S.V.; Goligorsky, D.M.; Hampel, D.J.; Li, H.; Gross, S.S.; Goligorsky, M.S. Glycated Collagen I Induces Premature Senescence-Like Phenotypic Changes in Endothelial Cells. Circ. Res. 2002, 90, 1290–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pageon, H.; Bakala, H.; Monnier, V.M.; Asselineau, D. Collagen glycation triggers the formation of aged skin in vitro. Eur. J. Dermatol. 2007, 17. [Google Scholar] [CrossRef]
- Aronson, D. Cross-linking of glycated collagen in the pathogenesis of arterial and myocardial stiffening of aging and diabetes. J. Hypertens. 2003, 21, 3–12. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shi, H.; Cheng, Y.; Ye, J.; Cai, P.; Zhang, J.; Li, R.; Yang, Y.; Wang, Z.; Zhang, H.; Lin, C.; et al. bFGF Promotes the Migration of Human Dermal Fibroblasts under Diabetic Conditions through Reactive Oxygen Species Production via the PI3K/Akt-Rac1- JNK Pathways. Int. J. Biol. Sci. 2015, 11, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, J.; Fang, Q.; Liu, M.; Liu, X.; Jia, W.; Dong, L.Q.; Liu, F. Autophagy-Mediated Insulin Receptor Down-Regulation Contributes to Endoplasmic Reticulum Stress-Induced Insulin Resistance. Mol. Pharmacol. 2009, 76, 596–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loots, M.A.; Lamme, E.N.; Zeegelaar, J.; Mekkes, J.R.; Bos, J.D.; Middelkoop, E. Differences in Cellular Infiltrate and Extracellular Matrix of Chronic Diabetic and Venous Ulcers Versus Acute Wounds. J. Investig. Dermatol. 1998, 111, 850–857. [Google Scholar] [CrossRef] [Green Version]
- Boulton, A.J.M. The pathway to foot ulceration in diabetes. Med. Clin. 2013, 97, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Zakhaleva, J.; Chen, W. Cells and tissue interactions with glycated collagen and their relevance to delayed diabetic wound healing. Biomaterials 2009, 30, 1689–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahrani, A.A.; Zeng, W.; Shakher, J.; Piya, M.K.; Hughes, S.; Dubb, K.; Stevens, M.J. Cutaneous Structural and Biochemical Correlates of Foot Complications in High-Risk Diabetes. Diabetes Care 2012, 35, 1913–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maquart, F.X.; Monboisse, J.C. Extracellular matrix and wound healing. Pathol. Biol. 2014, 62, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Shu, J.; Lee, C.-T. Whole-genome expression analyses of type 2 diabetes in human skin reveal altered immune function and burden of infection. Oncotarget 2017, 8, 34601–34609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiglitz, B.M.; Keene, D.R.; Greenspan, D.S. PCOLCE2 Encodes a Functional Procollagen C-Proteinase Enhancer (PCPE2) That Is a Collagen-binding Protein Differing in Distribution of Expression and Post-translational Modification from the Previously Described PCPE1. J. Biol. Chem. 2002, 277, 49820–49830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichert, O.; Fleming, T.; Neufang, G.; Schmelz, M.; Genth, H.; Kaever, V.; Wenck, H.; Stäb, F.; Terstegen, L.; Kolbe, L.; et al. Impaired glyoxalase activity is associated with reduced expression of neurotrophic factors and pro-inflammatory processes in diabetic skin cells. Exp. Dermatol. 2017, 26, 44–50. [Google Scholar] [CrossRef]
- Sejersen, H.; Rattan, S.I.S. Dicarbonyl-induced accelerated aging in vitro in human skin fibroblasts. Biogerontology 2009, 10, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, N.; Abe, M.; Souma, T.; Tanemoto, M.; Abe, T.; Nakayama, M.; Ito, S. Methylglyoxal augments intracellular oxidative stress in human aortic endothelial cells. Free. Radic. Res. 2009, 44, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.M.O.; Villar-Delfino, P.H.; dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Yu, M.-R.; Yang, Y.; Jiang, Z.; Ha, H. Reactive Oxygen Species-Regulated Signaling Pathways in Diabetic Nephropathy. J. Am. Soc. Nephrol. 2003, 14, S241–S245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornalley, P.J. The glyoxalase system in health and disease. Mol. Asp. Med. 1993, 14, 287–371. [Google Scholar] [CrossRef]
- Skapare, E.; Konrade, I.; Liepinsh, E.; Strele, I.; Makrecka-Kuka, M.; Bierhaus, A.; Lejnieks, A.; Pirags, V.; Dambrova, M. Association of reduced glyoxalase 1 activity and painful peripheral diabetic neuropathy in type 1 and 2 diabetes mellitus patients. J. Diabetes Complicat. 2013, 27, 262–267. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Glyoxalase in diabetes, obesity and related disorders. Semin. Cell Dev. Biol. 2011, 22, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Farr, L.; Ghosh, S.; Moonah, S. Role of MIF Cytokine/CD74 Receptor Pathway in Protecting Against Injury and Promoting Repair. Front. Immunol. 2020, 11, 1273. [Google Scholar] [CrossRef]
- Bhogal, R.K.; Bona, C.A. Regulatory Effect of Extracellular Signal-Regulated Kinases (ERK) on Type I Collagen Synthesis in Human Dermal Fibroblasts Stimulated by IL-4 and IL-13. Int. Rev. Immunol. 2008, 27, 472–496. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1: Donors with T2D | Group 2: Donors without T2D | |
|---|---|---|
| n total n female n male | 10 3 7 | 10 3 7 |
| Age (y) | 54 (±9) | 54 (±9) |
| HbA1c (%) | 7.06 (54 mmol/mol) (±1.03) | 5.44 (36 mmol/mol) (±0.22) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nickel, K.; Wensorra, U.; Wenck, H.; Peters, N.; Genth, H. Evaluation of Immunomodulatory Responses and Changed Wound Healing in Type 2 Diabetes—A Study Exploiting Dermal Fibroblasts from Diabetic and Non-Diabetic Human Donors. Cells 2021, 10, 2931. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112931
Nickel K, Wensorra U, Wenck H, Peters N, Genth H. Evaluation of Immunomodulatory Responses and Changed Wound Healing in Type 2 Diabetes—A Study Exploiting Dermal Fibroblasts from Diabetic and Non-Diabetic Human Donors. Cells. 2021; 10(11):2931. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112931
Chicago/Turabian StyleNickel, Kimberly, Ursula Wensorra, Horst Wenck, Nils Peters, and Harald Genth. 2021. "Evaluation of Immunomodulatory Responses and Changed Wound Healing in Type 2 Diabetes—A Study Exploiting Dermal Fibroblasts from Diabetic and Non-Diabetic Human Donors" Cells 10, no. 11: 2931. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10112931