
Fingolimod (FTY720), a Sphinogosine-1-Phosphate Receptor Agonist, Mitigates Choroidal Endothelial Proangiogenic Properties and Choroidal Neovascularization

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Isolation and Characterization of Various Cells from Mouse Eyes
2.2. Expression of S1P Receptors
2.3. In Vitro Cell Viability
2.4. In Vitro Cell Migration Assay
2.5. In Vitro Capillary Morphogenesis Assays
2.6. Animals
2.7. Laser-Induced Choroidal Neovascularization (CNV)
2.8. Drug Treatment
2.9. Detection and Quantification of Choroidal Neovascularization
2.10. Statistical Analysis
3. Results
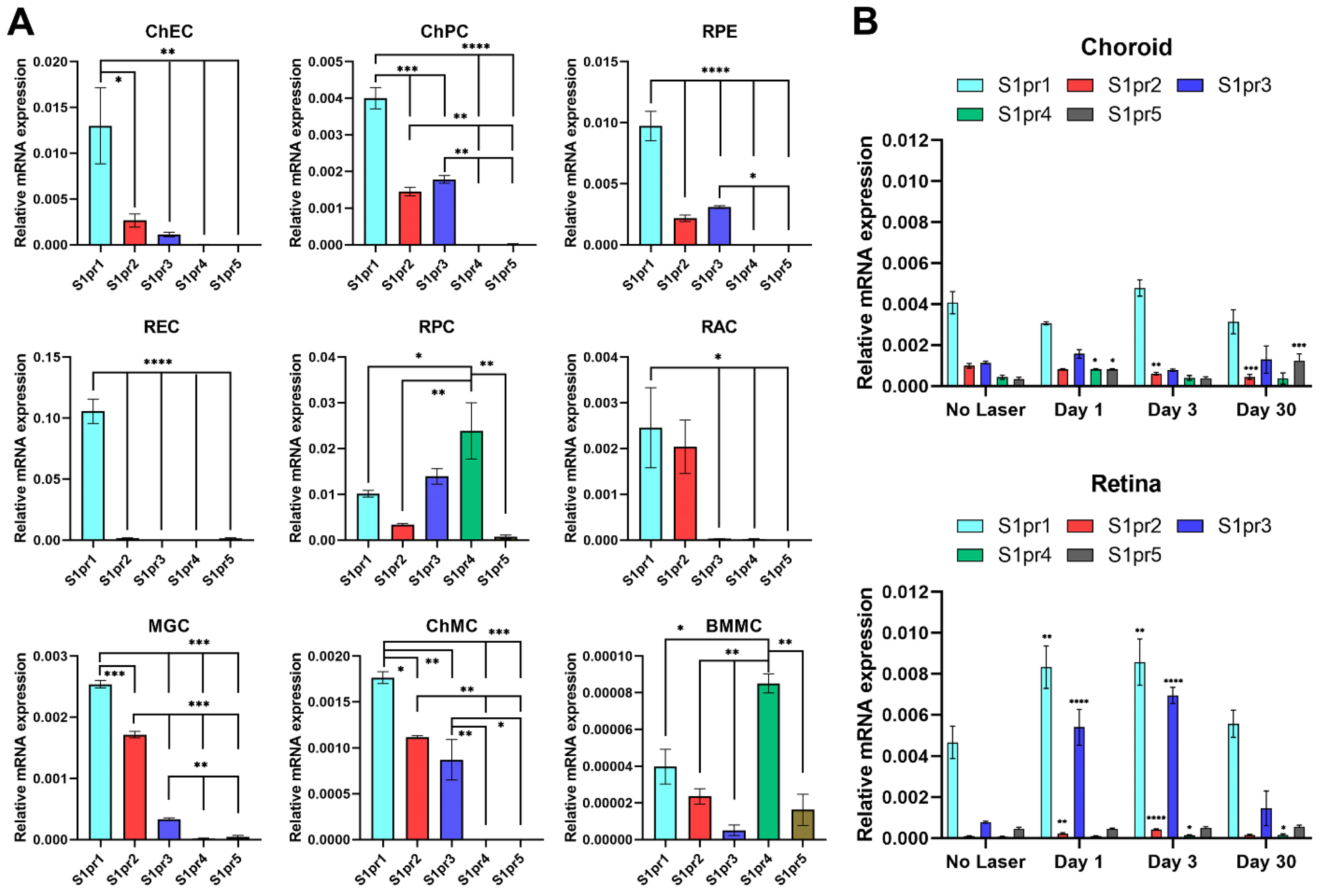
3.1. S1PR1 Is the Highest-Expressing Subtype in Choroidal Endothelial Cells
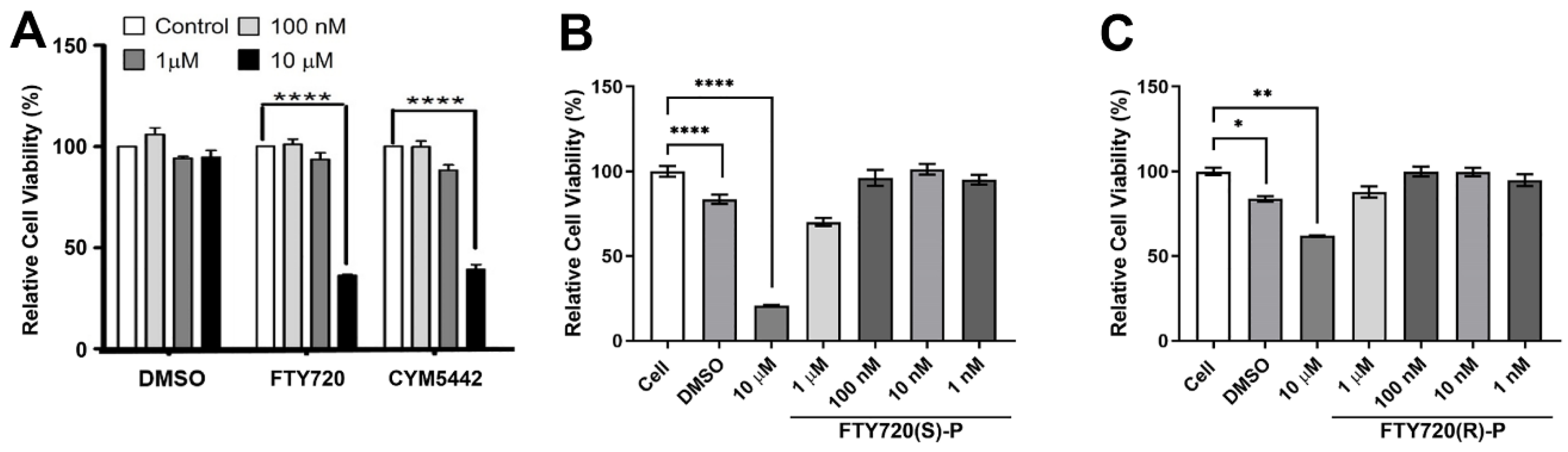
3.2. Decreased ChEC Viability with S1PR Agonists
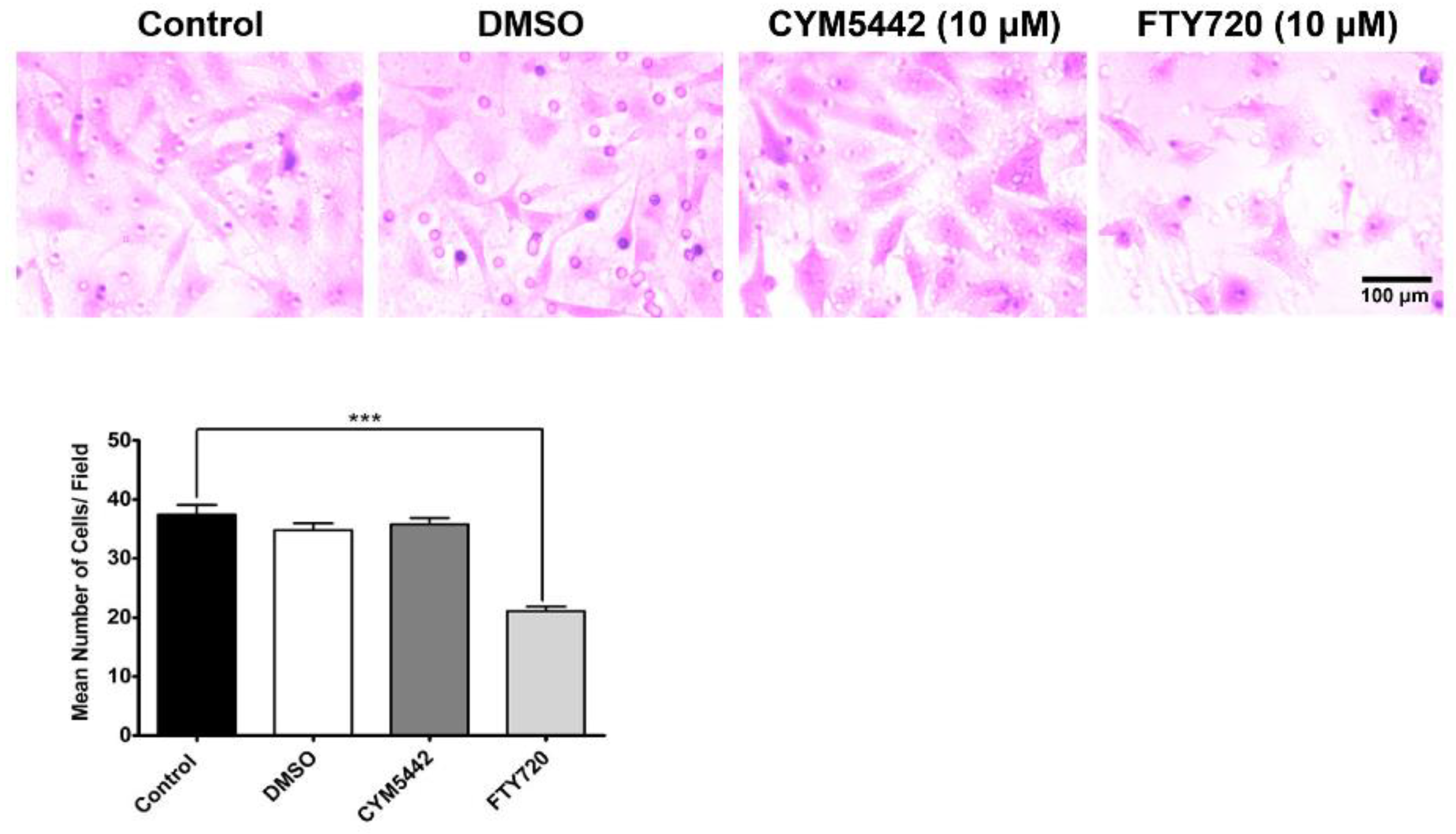
3.3. Agonism of S1PR Inhibits ChEC Migration
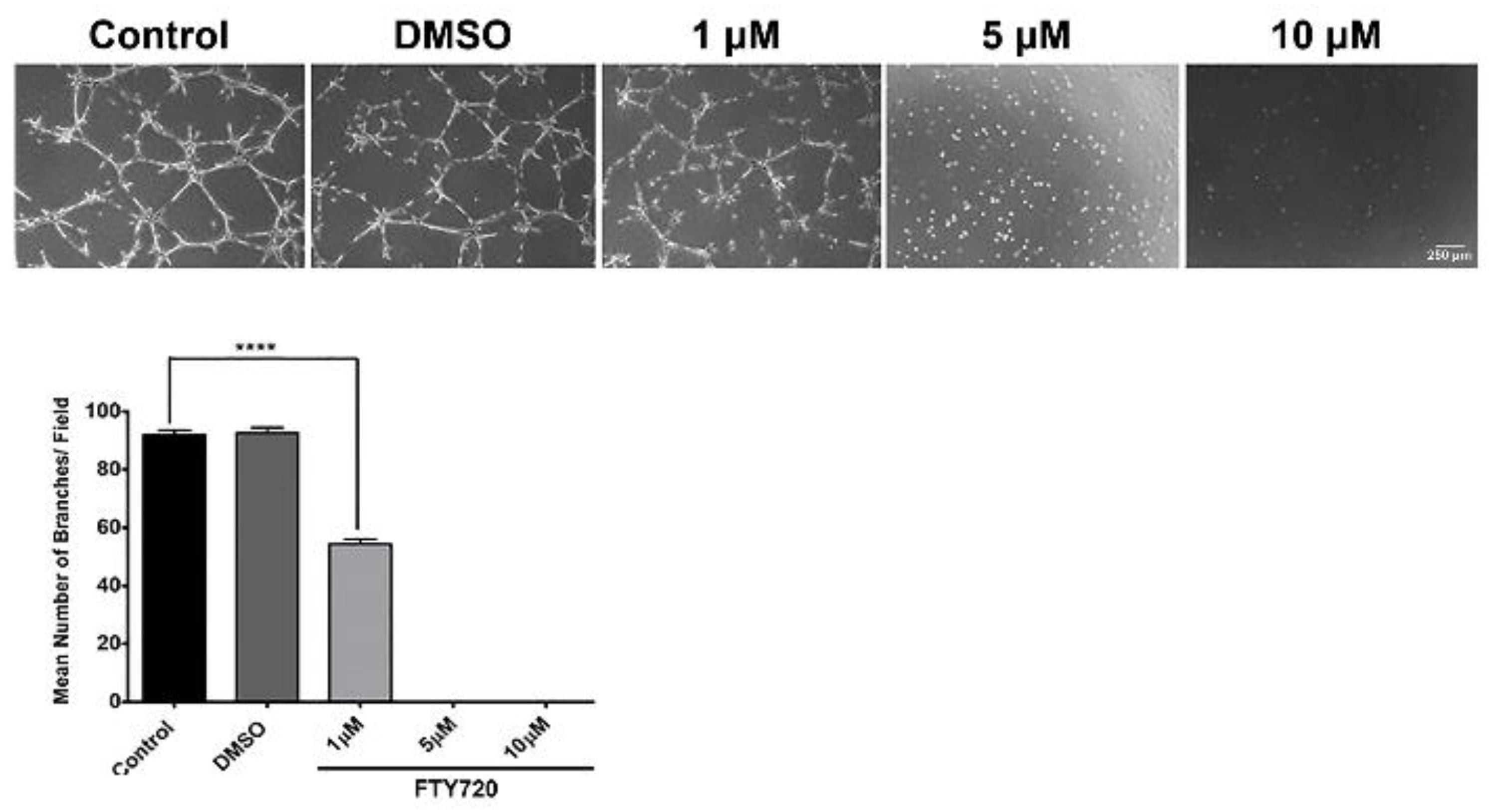
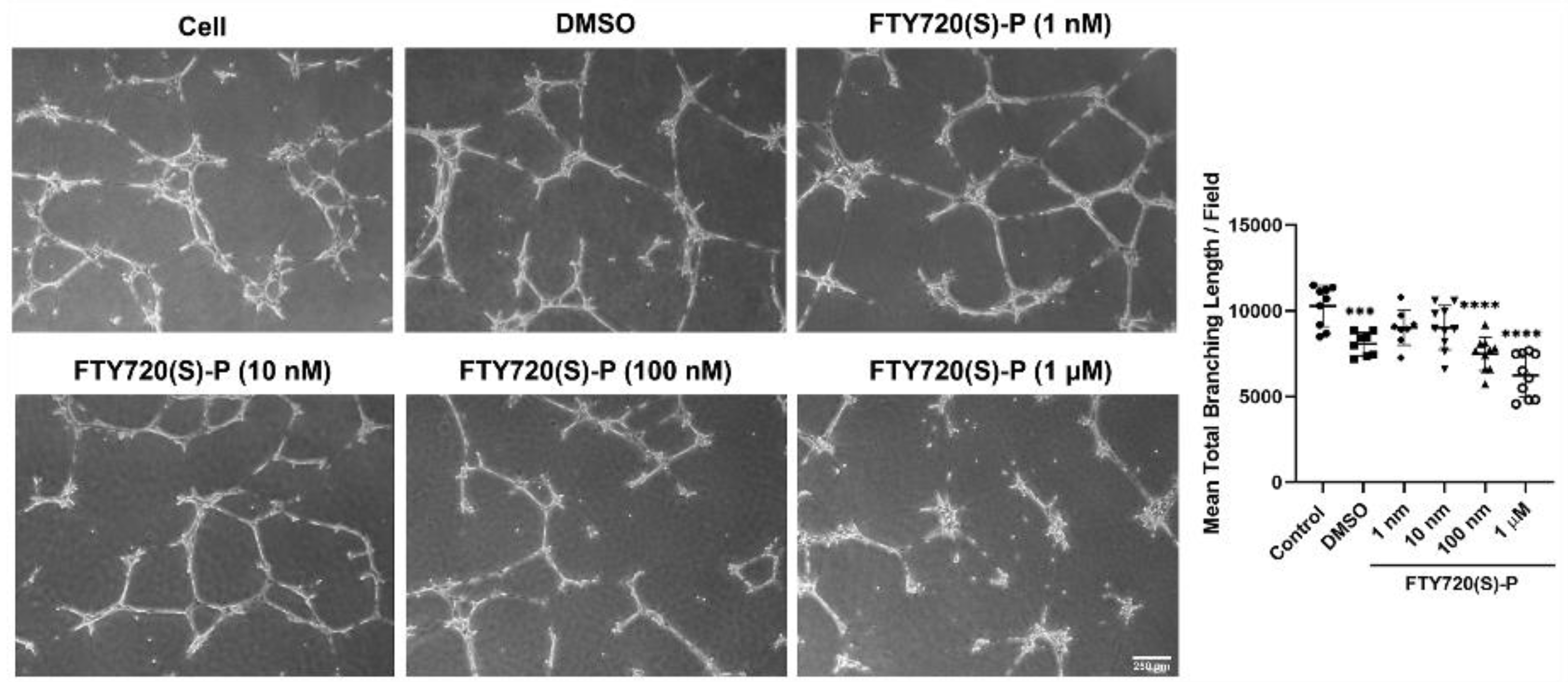
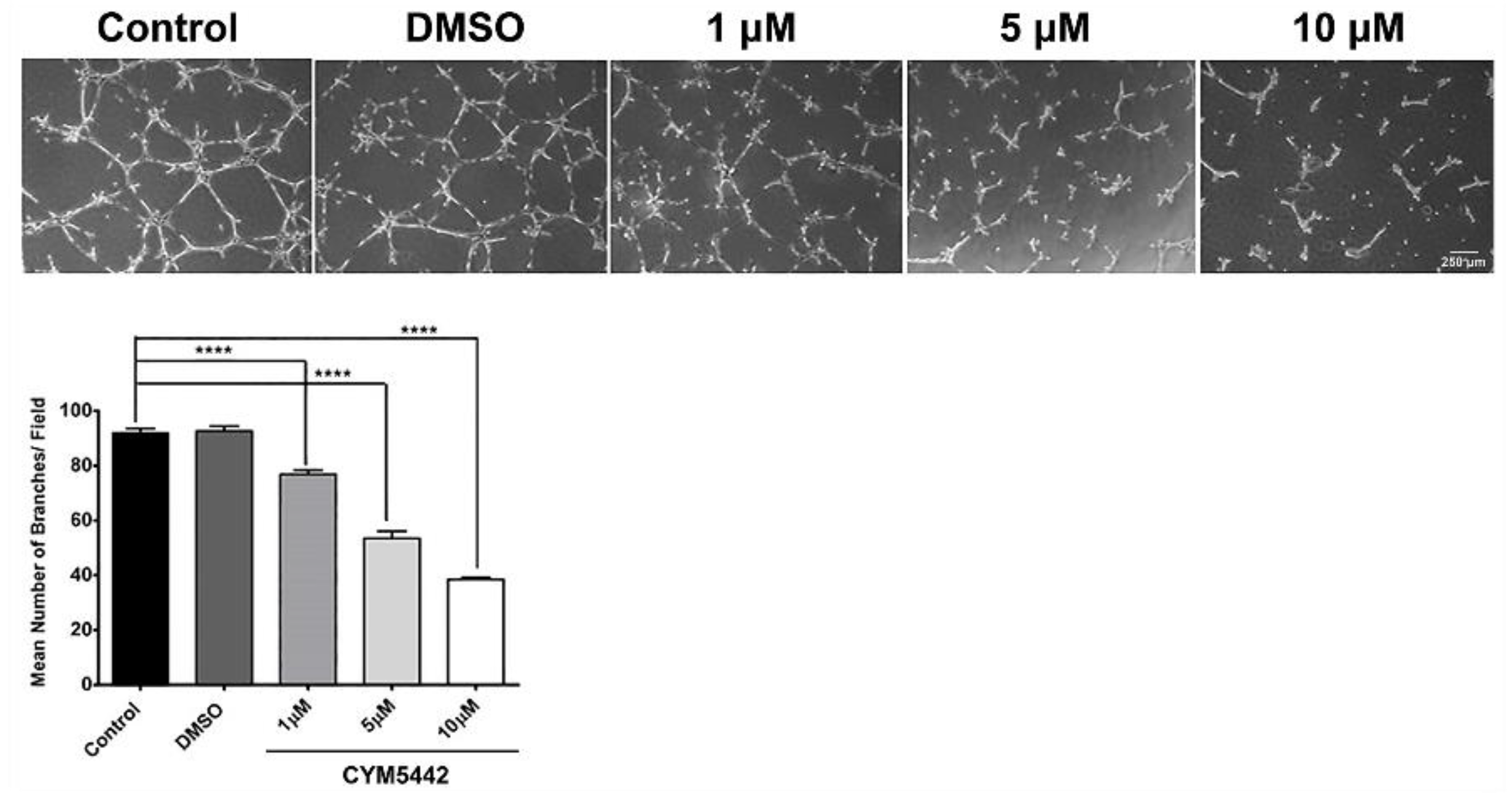
3.4. FTY720 and CYM5442 Inhibit ChEC Capillary Morphogenesis
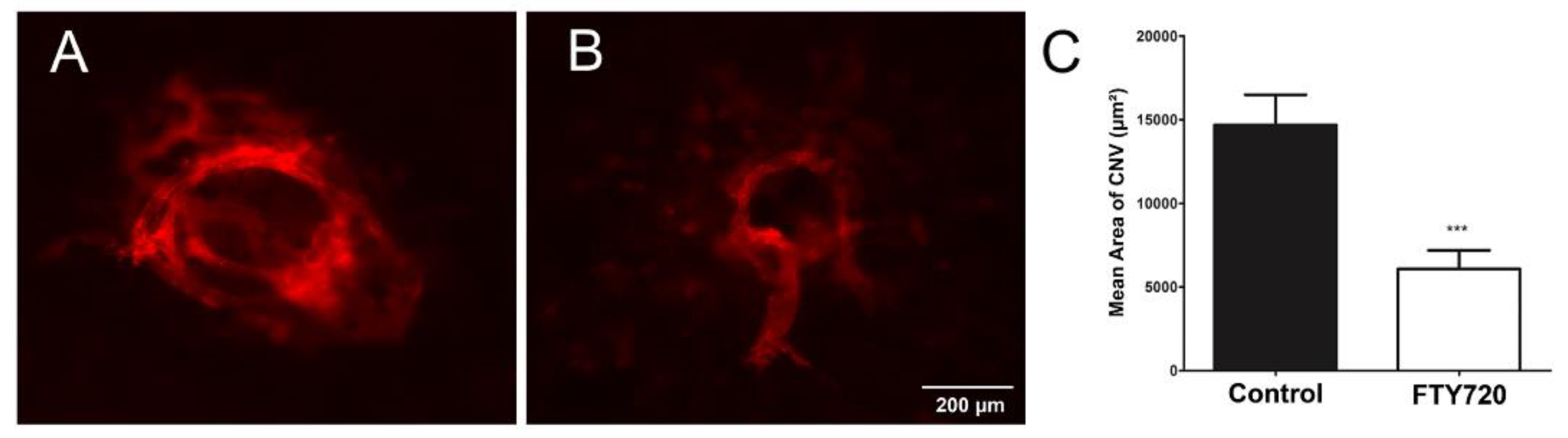
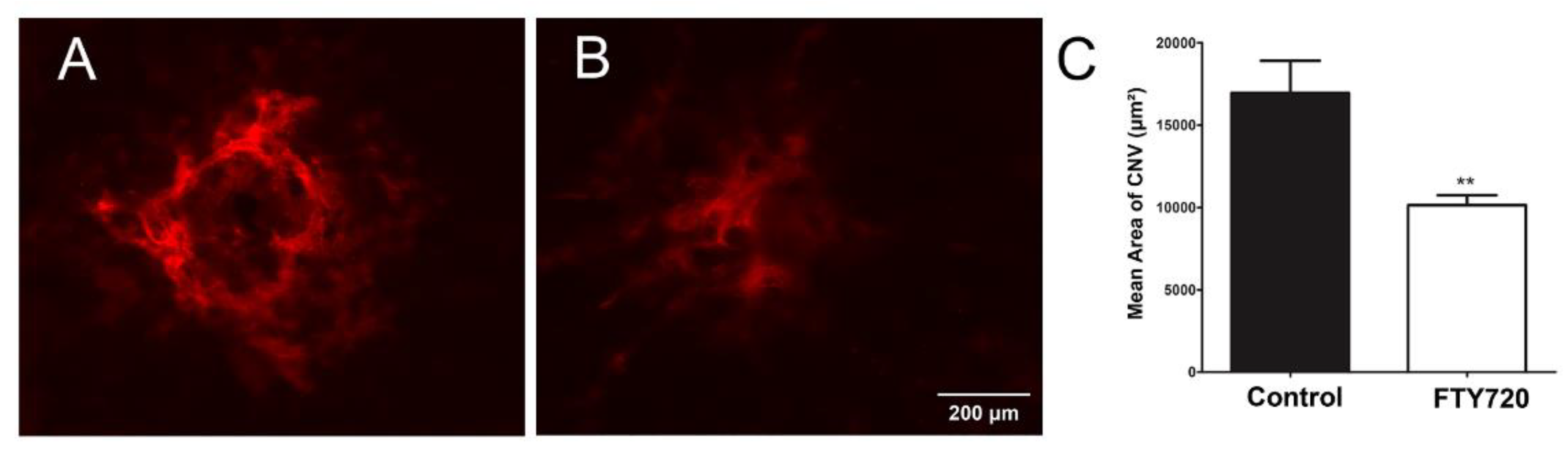
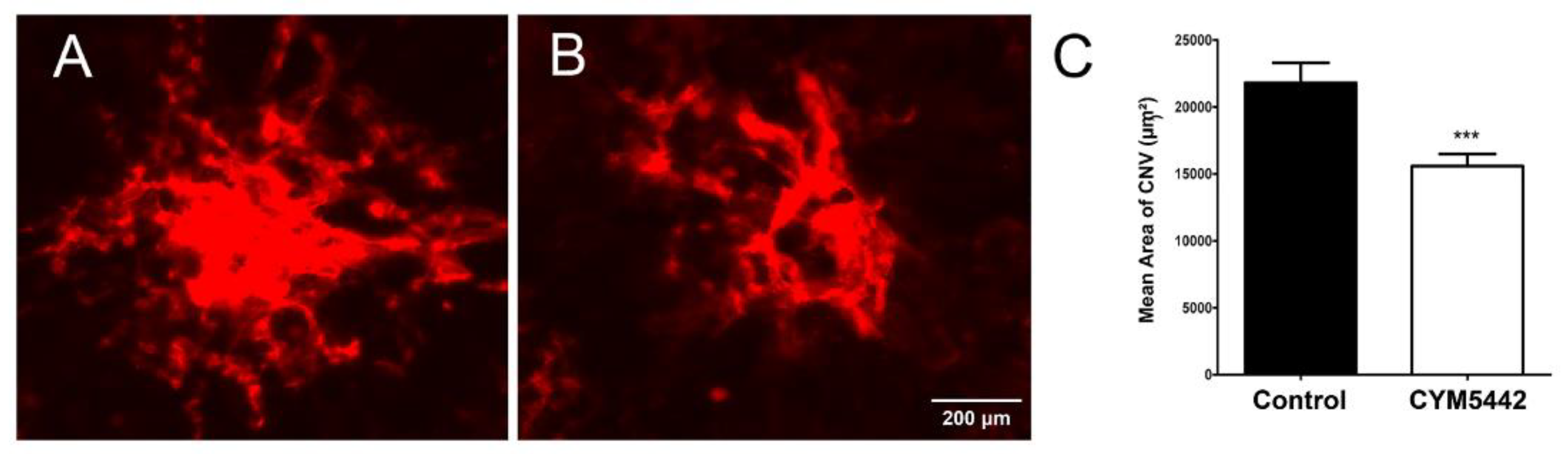
3.5. Agonism of S1PR Inhibits Choroidal Neovascularization In Vivo
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hernandez-Zimbron, L.F.; Zamora-Alvarado, R.; Ochoa-De la Paz, L.; Velez-Montoya, R.; Zenteno, E.; Gulias-Canizo, R.; Quiroz-Mercado, H.; Gonzalez-Salinas, R. Age-related macular degeneration: New paradigms for treatment and management of amd. Oxidative Med. Cell. Longev. 2018, 2018, 8374647. [Google Scholar] [CrossRef] [PubMed]
- Farnoodian, M.; Wang, S.; Dietz, J.; Nickells, R.W.; Sorenson, C.M.; Sheibani, N. Negative regulators of angiogenesis: Important targets for treatment of exudative amd. Clin. Sci. 2017, 131, 1763–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zając-Pytrus, H.M.; Pilecka, A.; Turno-Kręcicka, A.; Adamiec-Mroczek, J.; Misiuk-Hojło, M. The dry form of age-related macular degeneration (amd): The current concepts of pathogenesis and prospects for treatment. Adv. Clin. Exp. Med. 2015, 24, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Robredo, P.; Sancho, A.; Johnen, S.; Recalde, S.; Gama, N.; Thumann, G.; Groll, J.; García-Layana, A. Current treatment limitations in age-related macular degeneration and future approaches based on cell therapy and tissue engineering. J. Ophthalmol. 2014, 2014, 510285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hla, T.; Venkataraman, K.; Michaud, J. The vascular s1p gradient-cellular sources and biological significance. Biochim. Biophys. Acta 2008, 1781, 477–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappu, R.; Schwab, S.R.; Cornelissen, I.; Pereira, J.P.; Regard, J.B.; Xu, Y.; Camerer, E.; Zheng, Y.W.; Huang, Y.; Cyster, J.G.; et al. Promotion of lymphocyte egress into blood and lymph by distinct sources of sphingosine-1-phosphate. Science 2007, 316, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, K.; Lee, Y.M.; Michaud, J.; Thangada, S.; Ai, Y.; Bonkovsky, H.L.; Parikh, N.S.; Habrukowich, C.; Hla, T. Vascular endothelium as a contributor of plasma sphingosine 1-phosphate. Circ. Res. 2008, 102, 669–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kono, M.; Mi, Y.; Liu, Y.; Sasaki, T.; Allende, M.L.; Wu, Y.P.; Yamashita, T.; Proia, R.L. The sphingosine-1-phosphate receptors s1p1, s1p2, and s1p3 function coordinately during embryonic angiogenesis. J. Biol. Chem. 2004, 279, 29367–29373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terao, R.; Honjo, M.; Totsuka, K.; Miwa, Y.; Kurihara, T.; Aihara, M. The role of sphingosine 1-phosphate receptors on retinal pigment epithelial cells barrier function and angiogenic effects. Prostaglandins Other Lipid Mediat. 2019, 145, 106365. [Google Scholar] [CrossRef]
- Xie, B.; Shen, J.; Dong, A.; Rashid, A.; Stoller, G.; Campochiaro, P.A. Blockade of sphingosine-1-phosphate reduces macrophage influx and retinal and choroidal neovascularization. J. Cell. Physiol. 2009, 218, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Caballero, S.; Swaney, J.; Moreno, K.; Afzal, A.; Kielczewski, J.; Stoller, G.; Cavalli, A.; Garland, W.; Hansen, G.; Sabbadini, R.; et al. Anti-sphingosine-1-phosphate monoclonal antibodies inhibit angiogenesis and sub-retinal fibrosis in a murine model of laser-induced choroidal neovascularization. Exp. Eye Res. 2009, 88, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Billich, A.; Baumruker, T.; Heining, P.; Schmouder, R.; Francis, G.; Aradhye, S.; Burtin, P. Fingolimod (fty720): Discovery and development of an oral drug to treat multiple sclerosis. Nat. Rev. Drug Discov. 2010, 9, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Sorenson, C.M.; Sheibani, N. Isolation and characterization of murine retinal endothelial cells. Mol. Vis. 2003, 9, 171–178. [Google Scholar] [PubMed]
- Scheef, E.; Wang, S.; Sorenson, C.M.; Sheibani, N. Isolation and characterization of murine retinal astrocytes. Mol. Vis. 2005, 11, 613–624. [Google Scholar] [PubMed]
- Scheef, E.A.; Sorenson, C.M.; Sheibani, N. Attenuation of proliferation and migration of retinal pericytes in the absence of thrombospondin-1. Am. J. Physiol. Cell Physiol. 2009, 296, C724–C734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, P.; Zaitoun, I.; Farnoodian, M.; Fisk, D.L.; Wang, S.; Sorenson, C.M.; Sheibani, N. Expression of thrombospondin-1 modulates the angioinflammatory phenotype of choroidal endothelial cells. PLoS ONE 2014, 9, e116423. [Google Scholar] [CrossRef] [Green Version]
- Farnoodian, M.; Kinter, J.B.; Yadranji Aghdam, S.; Zaitoun, I.; Sorenson, C.M.; Sheibani, N. Expression of pigment epithelium-derived factor and thrombospondin-1 regulate proliferation and migration of retinal pigment epithelial cells. Physiol. Rep. 2015, 3, e12266. [Google Scholar] [CrossRef]
- Lavine, J.A.; Farnoodian, M.; Wang, S.; Darjatmoko, S.R.; Wright, L.S.; Gamm, D.M.; Ip, M.S.; Sorenson, C.M.; Sheibani, N. Beta2-adrenergic receptor antagonism attenuates cnv through inhibition of vegf and il-6 expression. Investig. Ophthalmol. Vis. Sci. 2017, 58, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, S.J. The development of an experimental model of subretinal neovascularization in disciform macular degeneration. Trans. Am. Ophthalmol. Soc. 1979, 77, 707–745. [Google Scholar]
- Wang, S.; Sorenson, C.M.; Sheibani, N. Lack of thrombospondin 1 and exacerbation of choroidal neovascularization. Arch. Ophthalmol. 2012, 130, 615–620. [Google Scholar] [CrossRef] [Green Version]
- Lavine, J.A.; Sang, Y.; Wang, S.; Ip, M.S.; Sheibani, N. Attenuation of choroidal neovascularization by beta2-adrenoreceptor antagonism. JAMA Ophthalmol. 2013, 131, 376–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lee, D.K. What is the proper way to apply the multiple comparison test? Korean J. Anesthesiol. 2018, 71, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.W.; Gardell, S.E.; Herr, D.R.; Rivera, R.; Lee, C.W.; Noguchi, K.; Teo, S.T.; Yung, Y.C.; Lu, M.; Kennedy, G.; et al. Fty720 (fingolimod) efficacy in an animal model of multiple sclerosis requires astrocyte sphingosine 1-phosphate receptor 1 (s1p1) modulation. Proc. Natl. Acad. Sci. USA 2011, 108, 751–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, R.; Hinterding, K.; Brinkmann, V.; Guerini, D.; Müller-Hartwieg, C.; Knecht, H.; Simeon, C.; Streiff, M.; Wagner, T.; Welzenbach, K.; et al. Novel immunomodulator fty720 is phosphorylated in rats and humans to form a single stereoisomer. Identification, chemical proof, and biological characterization of the biologically active species and its enantiomer. J. Med. Chem. 2005, 48, 5373–5377. [Google Scholar] [CrossRef] [PubMed]
- Morrison, M.E.; Palenski, T.L.; Jamali, N.; Sheibani, N.; Sorenson, C.M. Modulation of vascular cell function by bim expression. Int. J. Cell Biol. 2013, 2013, 297537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheibani, N.; Morrison, M.E.; Gurel, Z.; Park, S.; Sorenson, C.M. Bim deficiency differentially impacts the function of kidney endothelial and epithelial cells through modulation of their local microenvironment. Am. J. Physiol.-Ren. Physiol. 2012, 302, F809–F819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaengel, K.; Niaudet, C.; Hagikura, K.; Laviña, B.; Muhl, L.; Hofmann, J.J.; Ebarasi, L.; Nyström, S.; Rymo, S.; Chen, L.L.; et al. The sphingosine-1-phosphate receptor s1pr1 restricts sprouting angiogenesis by regulating the interplay between ve-cadherin and vegfr2. Dev. Cell 2012, 23, 587–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arya, M.; Sabrosa, A.S.; Duker, J.S.; Waheed, N.K. Choriocapillaris changes in dry age-related macular degeneration and geographic atrophy: A review. Eye Vis. 2018, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biesemeier, A.; Taubitz, T.; Julien, S.; Yoeruek, E.; Schraermeyer, U. Choriocapillaris breakdown precedes retinal degeneration in age-related macular degeneration. Neurobiol. Aging 2014, 35, 2562–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutty, G.A.; McLeod, D.S.; Bhutto, I.A.; Edwards, M.M.; Seddon, J.M. Choriocapillaris dropout in early age-related macular degeneration. Exp. Eye Res. 2020, 192, 107939. [Google Scholar] [CrossRef]
- Skoura, A.; Sanchez, T.; Claffey, K.; Mandala, S.M.; Proia, R.L.; Hla, T. Essential role of sphingosine 1-phosphate receptor 2 in pathological angiogenesis of the mouse retina. J. Clin. Investig. 2007, 117, 2506–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fettel, J.; Kühn, B.; Guillen, N.A.; Sürün, D.; Peters, M.; Bauer, R.; Angioni, C.; Geisslinger, G.; Schnütgen, F.; Meyer Zu Heringdorf, D.; et al. Sphingosine-1-phosphate (s1p) induces potent anti-inflammatory effects in vitro and in vivo by s1p receptor 4-mediated suppression of 5-lipoxygenase activity. FASEB J. 2019, 33, 1711–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Cao, C.; Chen, Z.; Bankaitis, V.; Tzima, E.; Sheibani, N.; Burridge, K. Pericytes regulate vascular basement membrane remodeling and govern neutrophil extravasation during inflammation. PLoS ONE 2012, 7, e45499. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Zhang, X.D.; Miao, W.Y.; Sun, Y.J.; Xiong, G.; Wu, Q.; Li, G.; Yang, P.; Yu, H.; Li, H.; et al. Pdgfrbeta cells rapidly relay inflammatory signal from the circulatory system to neurons via chemokine CCL2. Neuron 2018, 100, 183–200.e8. [Google Scholar] [PubMed] [Green Version]
- Allende, M.L.; Yamashita, T.; Proia, R.L. G-protein-coupled receptor s1p1 acts within endothelial cells to regulate vascular maturation. Blood 2003, 102, 3665–3667. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wada, R.; Yamashita, T.; Mi, Y.; Deng, C.X.; Hobson, J.P.; Rosenfeldt, H.M.; Nava, V.E.; Chae, S.S.; Lee, M.J.; et al. Edg-1, the g protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J. Clin. Investig. 2000, 106, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allende, M.L.; Proia, R.L. Sphingosine-1-phosphate receptors and the development of the vascular system. Biochim. Biophys. Acta 2002, 1582, 222–227. [Google Scholar] [CrossRef]
- McGuire, P.G.; Rangasamy, S.; Maestas, J.; Das, A. Pericyte-derived sphingosine 1-phosphate induces the expression of adhesion proteins and modulates the retinal endothelial cell barrier. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e107–e115. [Google Scholar] [CrossRef] [Green Version]
- Ben Shoham, A.; Malkinson, G.; Krief, S.; Shwartz, Y.; Ely, Y.; Ferrara, N.; Yaniv, K.; Zelzer, E. S1p1 inhibits sprouting angiogenesis during vascular development. Development 2012, 139, 3859–3869. [Google Scholar] [CrossRef] [Green Version]
- Kihara, Y.; Jonnalagadda, D.; Zhu, Y.; Ray, M.; Ngo, T.; Palmer, C.; Rivera, R.; Chun, J. Ponesimod inhibits astrocyte-mediated neuroinflammation and protects against cingulum demyelination via s1p(1)-selective modulation. FASEB J. 2022, 36, e22132. [Google Scholar] [CrossRef]
- Terao, R.; Honjo, M.; Aihara, M. Apolipoprotein m inhibits angiogenic and inflammatory response by sphingosine 1-phosphate on retinal pigment epithelium cells. Int. J. Mol. Sci. 2017, 19, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; DiMaio, T.A.; Scheef, E.A.; Sorenson, C.M.; Sheibani, N. Pecam-1 regulates proangiogenic properties of endothelial cells through modulation of cell-cell and cell-matrix interactions. Am. J. Physiol. Cell Physiol. 2010, 299, C1468–C1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Scheef, E.A.; Wang, S.; Sorenson, C.M.; Marcus, C.B.; Jefcoate, C.R.; Sheibani, N. Cyp1b1 expression promotes the proangiogenic phenotype of endothelium through decreased intracellular oxidative stress and thrombospondin-2 expression. Blood 2009, 113, 744–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Ma, S.; Lin, D.; Mei, Y.; Gong, H.; Lei, L.; Chen, Y.; Zhao, Y.; Hu, B.; Wu, Y.; et al. The s1p1 receptor-selective agonist cym-5442 reduces the severity of acute gvhd by inhibiting macrophage recruitment. Cell. Mol. Immunol. 2015, 12, 681–691. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Alshaker, H.; Cooper, C.; Winkler, M.; Pchejetski, D. The emerging role of fty720 (fingolimod) in cancer treatment. Oncotarget 2016, 7, 23106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pchejetski, D.; Bohler, T.; Brizuela, L.; Sauer, L.; Doumerc, N.; Golzio, M.; Salunkhe, V.; Teissié, J.; Malavaud, B.; Waxman, J.; et al. Fty720 (fingolimod) sensitizes prostate cancer cells to radiotherapy by inhibition of sphingosine kinase-1. Cancer Res. 2010, 70, 8651–8661. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ to 3′) | Reverse (5′ to 3′) |
|---|---|---|
| S1pr1 | CGGTGTAGACCCAGAGTCCT | AGCTTTTCCTTGGCTGGAG |
| S1pr2 | CCCAACTCCGGGACATAGA | ACAGCCAGTGGTTGGTTTTG |
| S1pr3 | TCTCCCAATTGTTCCCTGAA | ACTTCAACAGTCCACGAGAGG |
| S1pr4 | CGTGATGAATGTTTGGCAGA | CCCTTCAAGGCCCAGACT |
| S1pr5 | CAAGACTTCCCCACAACCTG | TATGGCTGCAGCAGAAATTG |
| Rpl13a | TCTCAAGGTTGTTCGGCTGAA | CCAGACGCCCCAGGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorenson, C.M.; Farnoodian, M.; Wang, S.; Song, Y.-S.; Darjatmoko, S.R.; Polans, A.S.; Sheibani, N. Fingolimod (FTY720), a Sphinogosine-1-Phosphate Receptor Agonist, Mitigates Choroidal Endothelial Proangiogenic Properties and Choroidal Neovascularization. Cells 2022, 11, 969. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060969
Sorenson CM, Farnoodian M, Wang S, Song Y-S, Darjatmoko SR, Polans AS, Sheibani N. Fingolimod (FTY720), a Sphinogosine-1-Phosphate Receptor Agonist, Mitigates Choroidal Endothelial Proangiogenic Properties and Choroidal Neovascularization. Cells. 2022; 11(6):969. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060969
Chicago/Turabian StyleSorenson, Christine M., Mitra Farnoodian, Shoujian Wang, Yong-Seok Song, Soesiawati R. Darjatmoko, Arthur S. Polans, and Nader Sheibani. 2022. "Fingolimod (FTY720), a Sphinogosine-1-Phosphate Receptor Agonist, Mitigates Choroidal Endothelial Proangiogenic Properties and Choroidal Neovascularization" Cells 11, no. 6: 969. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060969