
Gdnf Acts as a Germ Cell-Derived Growth Factor and Regulates the Zebrafish Germ Stem Cell Niche in Autocrine- and Paracrine-Dependent Manners

 , , , , ,
, , , , ,  ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Zebrafish Stocks
2.2. Sequence Analysis
2.3. Expression Profiling of Gdnf (gdnfa and gdnfb) and Gfrα1 (gfrα1a and gfrα1b) Transcripts in Zebrafish Testes
2.4. Differential Plating Method
2.5. Immunofluorescence and Western Blot
2.6. Recombinant Human GDNF
2.7. Testis Tissue Culture
2.8. In Silico Analysis of Putative Regulatory Sequences Upstream Human GDNF, Mouse Gdnf and Zebrafish Gdnfa
2.9. Statistical Analyses
3. Results
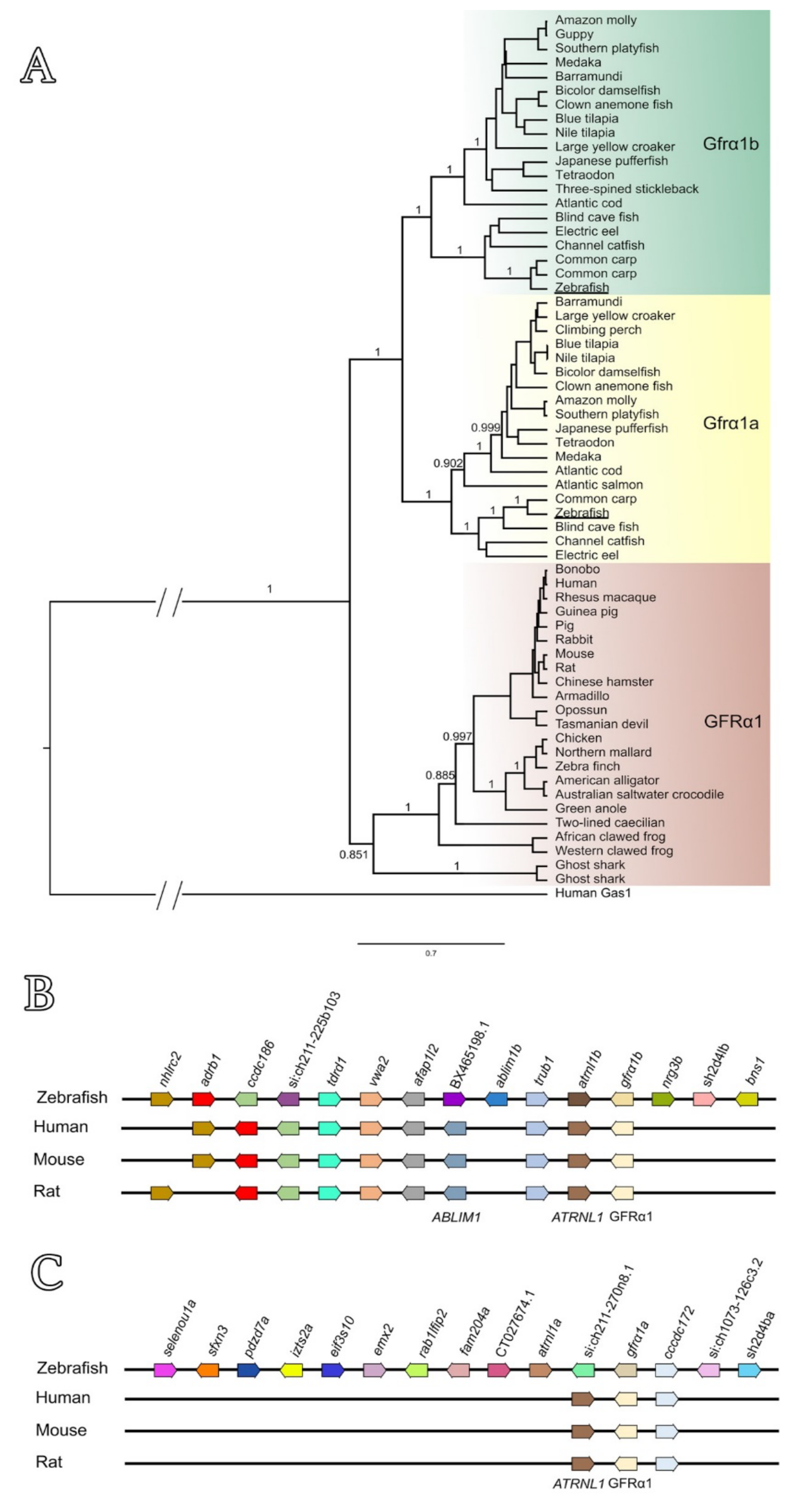
3.1. Sequence Analyses, Phylogenetic Tree and Genomic Organization of Zebrafish Gfrα1a and Gfrα1b
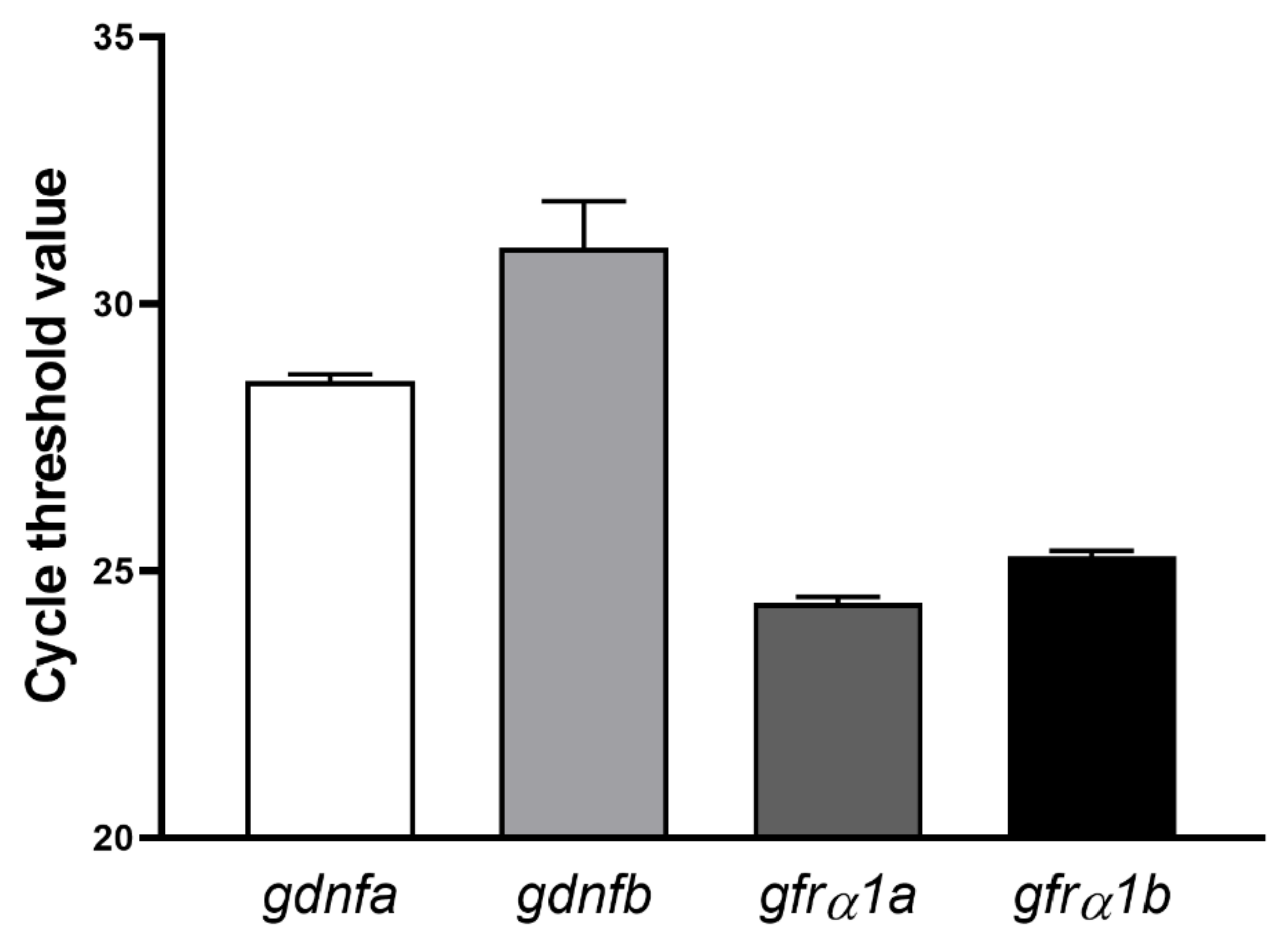
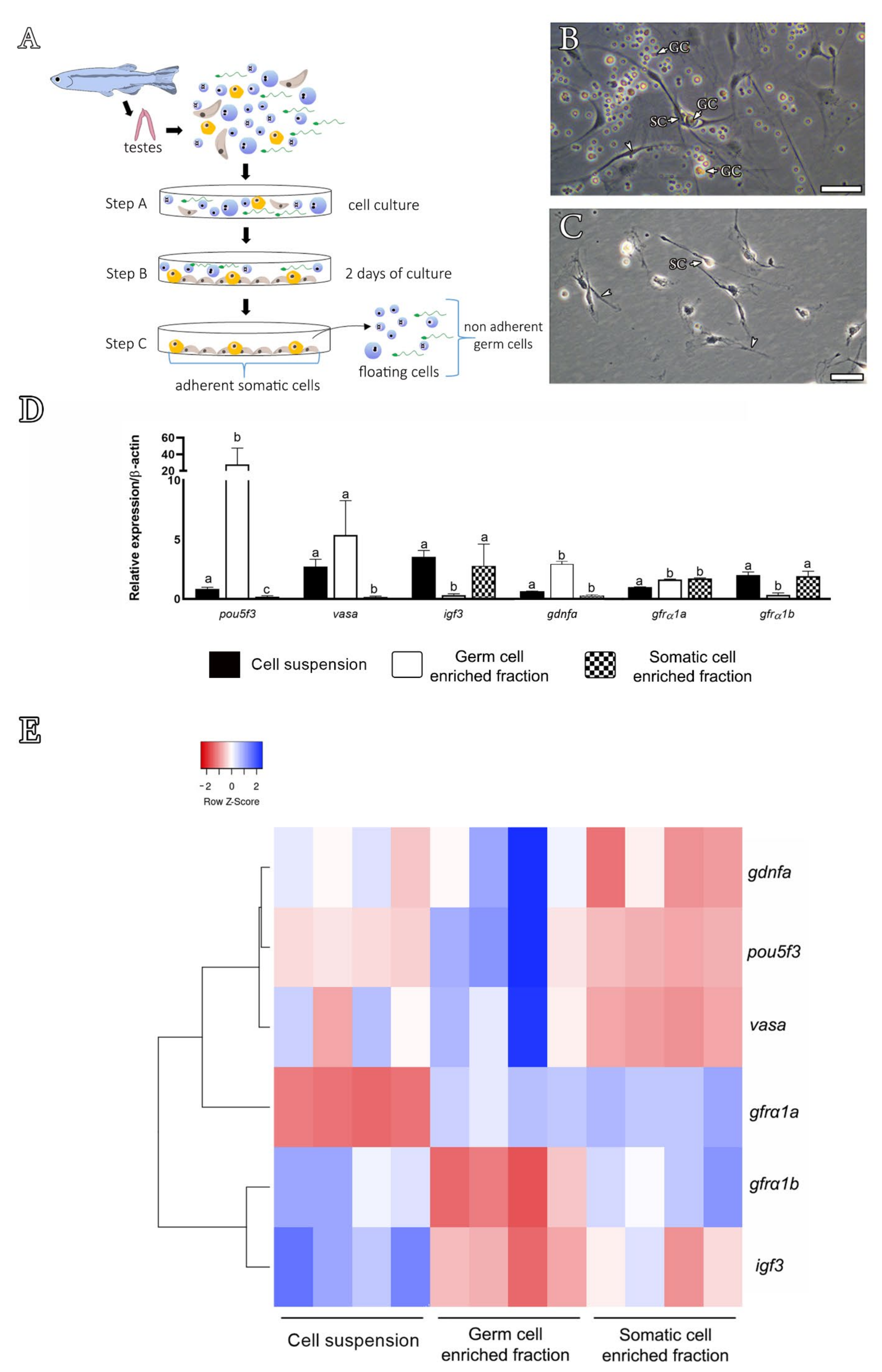
3.2. Expression Profiling in Zebrafish Testes and Identification of Gdnfa-, Gfrα1a- and Gfrα1b-Expressing Cells
3.3. Localization of Gfrα1a Protein in Zebrafish Testis
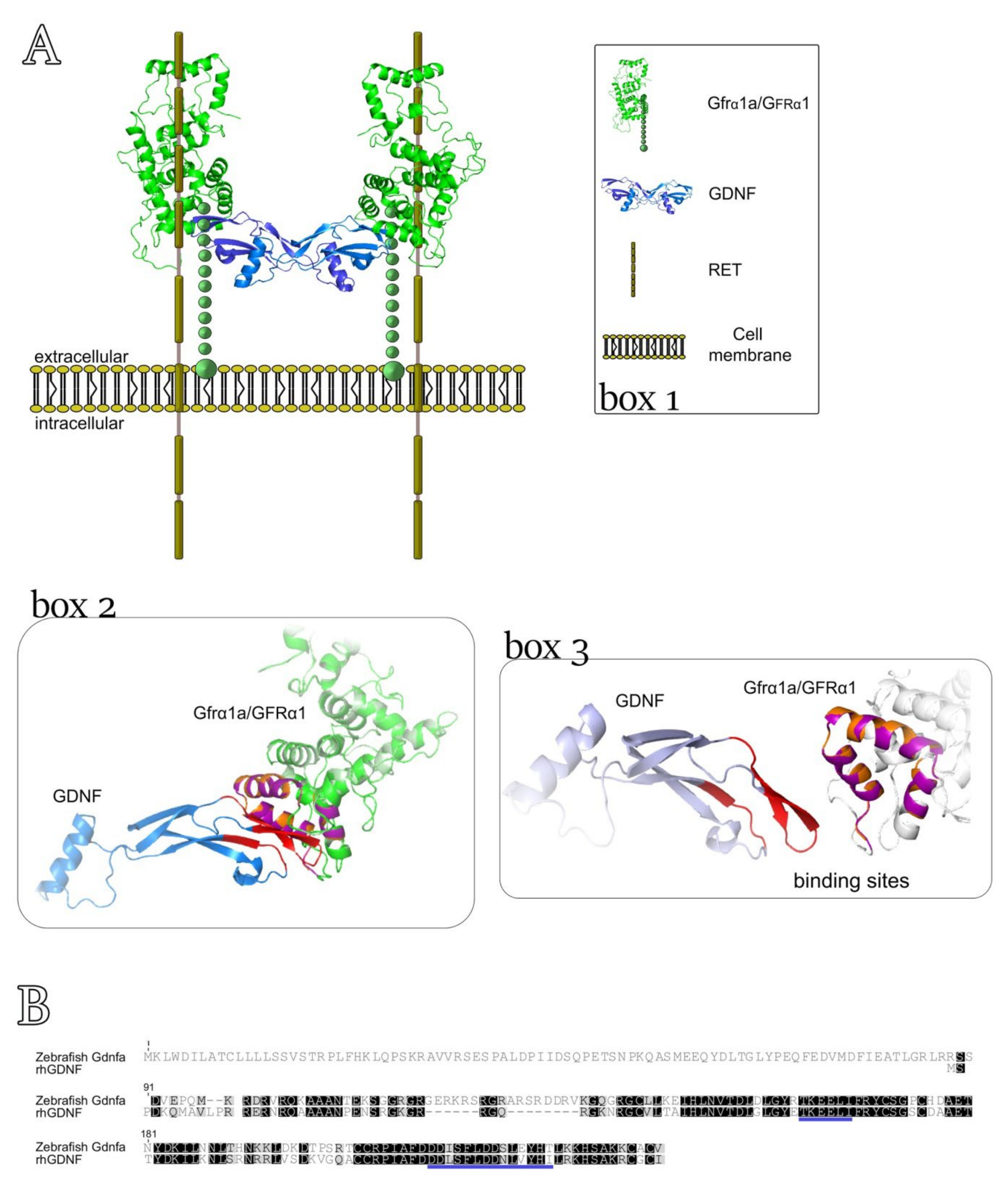
3.4. Three-Dimensional Model for Predicting the Interaction between rhGDNF and Zebrafish Gfrα1a
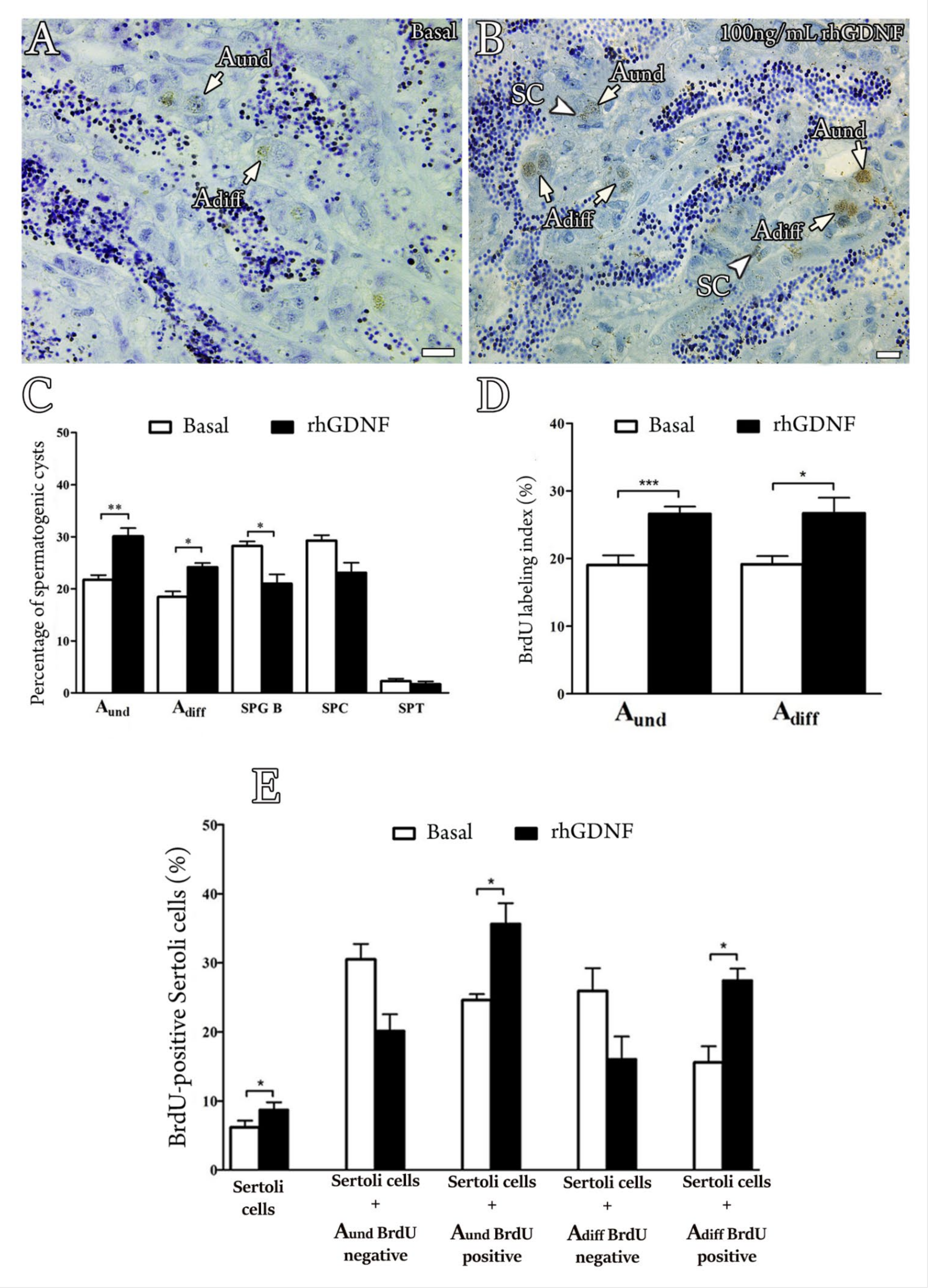
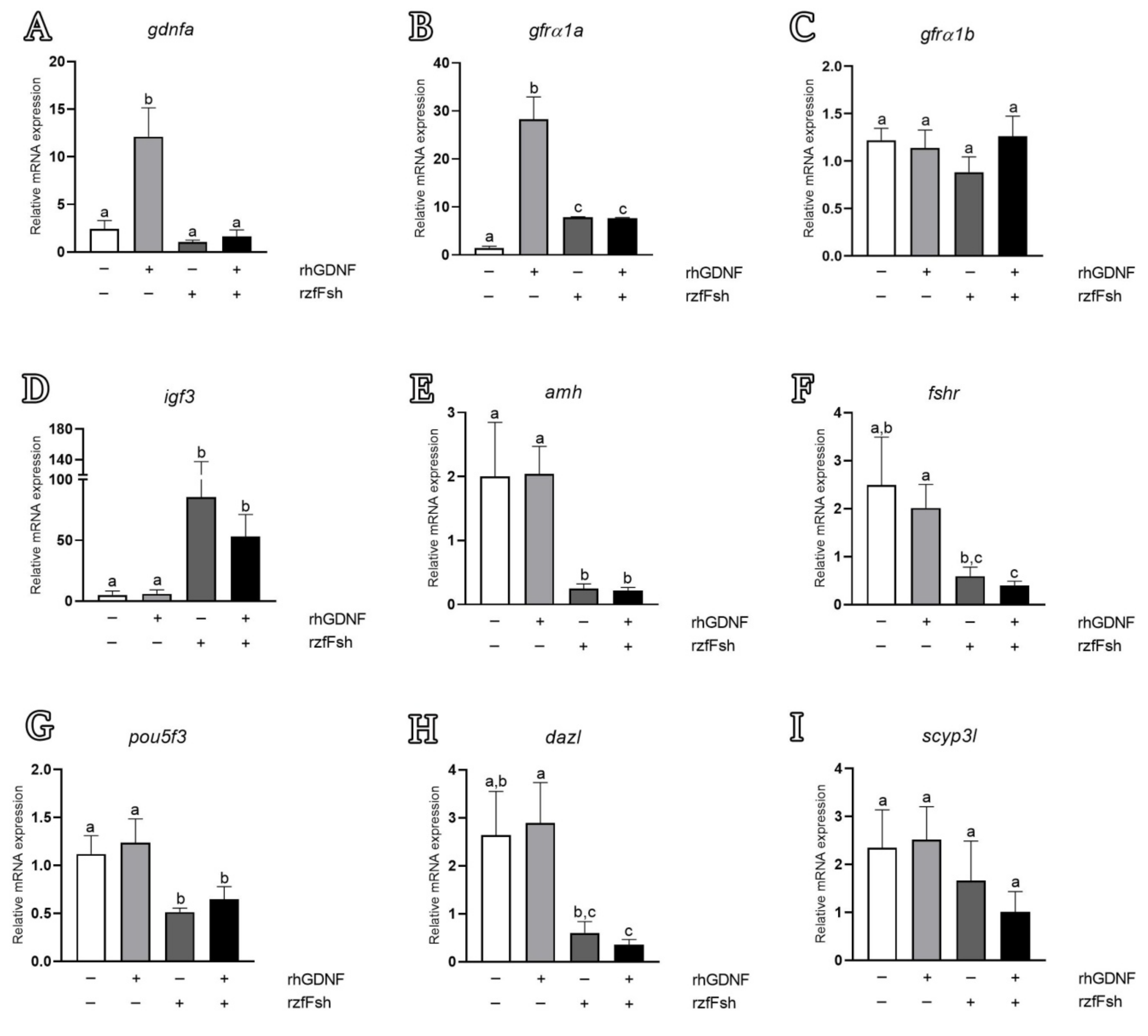
3.5. Biological Effects of rhGDNF
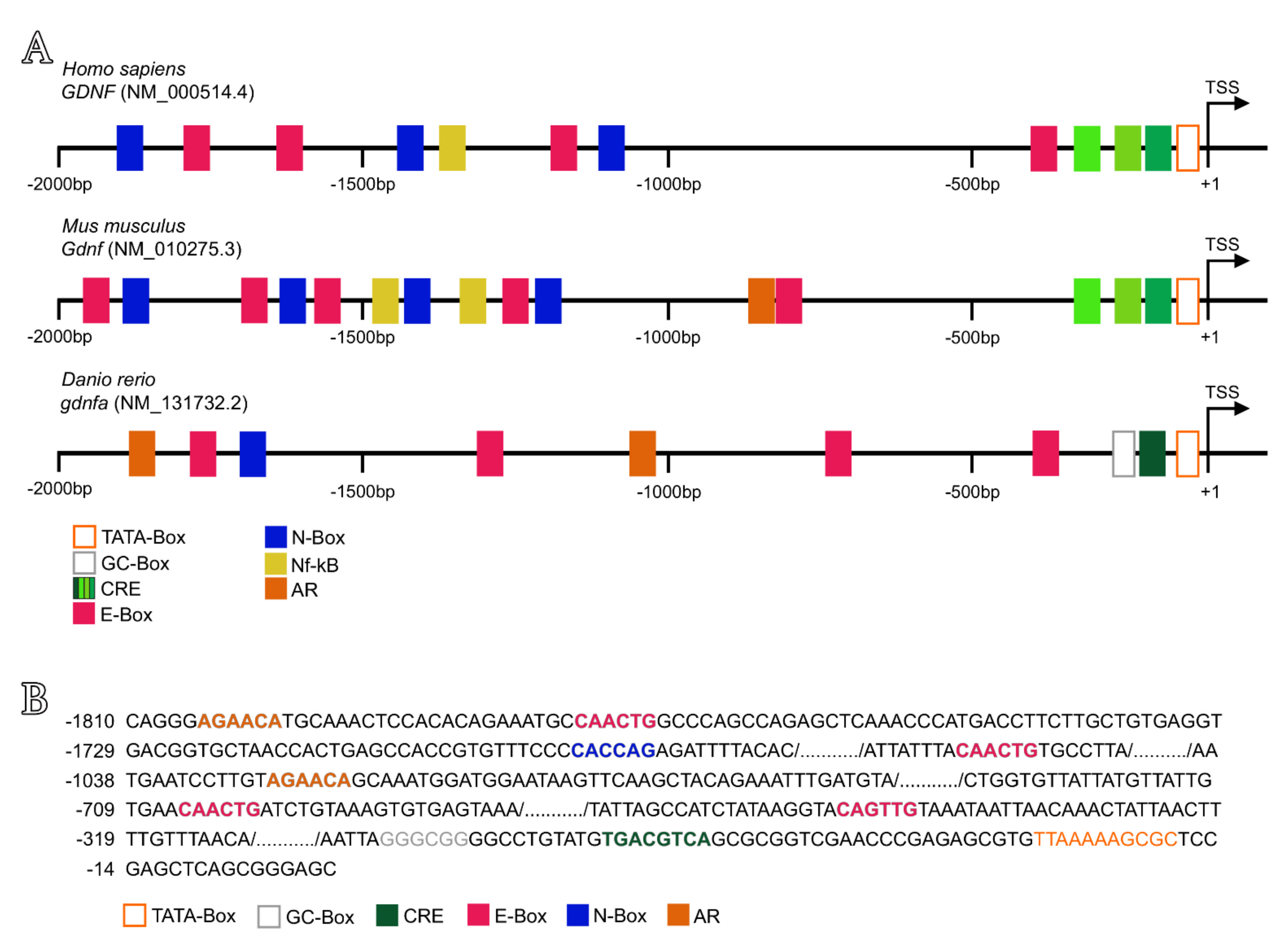
3.6. In Silico Analysis of Putative Regulatory Sequences Upstream of Human GDNF, Mouse Gdnf and Zebrafish Gdnfa
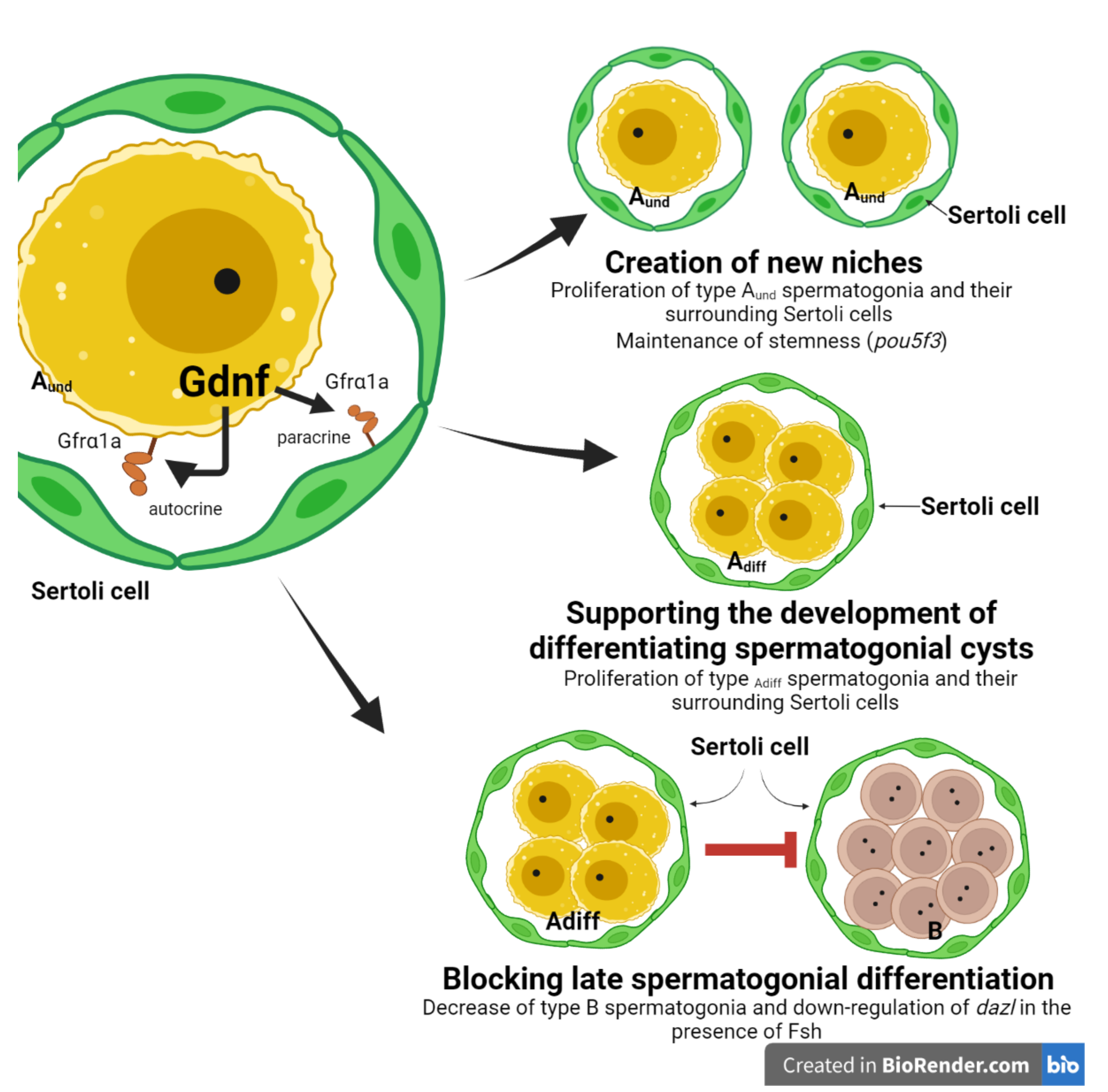
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zihlmann, K.B.; Ducray, A.D.; Schaller, B.; Huber, A.W.; Krebs, S.H.; Andres, R.H.; Seiler, R.W.; Meyer, M.; Widmer, H.R. The GDNF family members neurturin, artemin and persephin promote the morphological differentiation of cultured ventral mesencephalic dopaminergic neurons. Brain Res. Bull. 2005, 68, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Meng, X. Regulation of Cell Fate Decision of Undifferentiated Spermatogonia by GDNF. Science 2000, 287, 1489–1493. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16489–16494. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Braun, R.E. Cyclical expression of GDNF is required for spermatogonial stem cell homeostasis. Development 2018, 145, dev151555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakiuchi, K.; Taniguchi, K.; Kubota, H. Conserved and non-conserved characteristics of porcine glial cell line-derived neurotrophic factor expressed in the testis. Sci. Rep. 2018, 8, 7656. [Google Scholar] [CrossRef]
- Kubota, H.; Wu, X.; Avarbock, M.R.; Brinster, R.L. Glial cell line-derived neurotrophic factor and endothelial cells promote self-renewal of rabbit germ cells with spermatogonial stem cell properties. FASEB J. 2011, 25, 2604–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parekh, P.; Garcia, T.X.; Hofmann, M.C. Regulation of GDNF expression in Sertoli cells. Reproduction 2019, 157, R95–R107. [Google Scholar] [CrossRef]
- Mäkelä, J.A.; Hobbs, R.M. Molecular regulation of spermatogonial stem cell renewal and differentiation. Reproduction 2019, 158, R169–R187. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, E.; Tumbar, T.; Guasch, G. Socializing with the neighbors: Stem cells and their niche. Cell 2004, 116, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, M.-C. Gdnf signaling pathways within the mammalian spermatogonial stem cell niche. Mol. Cell. Endocrinol. 2008, 288, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Willis, W.D.; Eddy, E.M. Targeting the Gdnf Gene in peritubular myoid cells disrupts undifferentiated spermatogonial cell development. Proc. Natl. Acad. Sci. USA 2016, 113, 1829–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.Y.; Brown, P.R.; Willis, W.B.; Eddy, E.M. Peritubular myoid cells participate in male mouse spermatogonial stem cell maintenance. Endocrinology 2014, 155, 4964–4974. [Google Scholar] [CrossRef] [Green Version]
- Bhang, D.H.; Kim, B.J.; Kim, B.G.; Schadler, K.; Baek, K.H.; Kim, Y.H.; Hsiao, W.; Ding, B.S.; Rafii, S.; Weiss, M.J.; et al. Testicular endothelial cells are a critical population in the germline stem cell niche. Nat. Commun. 2018, 9, 4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helsel, A.R.; Yang, Q.E.; Oatley, M.J.; Lord, T.; Sablitzky, F.; Oatley, J.M. ID4 levels dictate the stem cell state in mouse spermatogonia. Development 2017, 144, 624–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, T.; Oatley, J.M. Functional assessment of spermatogonial stem cell purity in experimental cell populations. Stem Cell Res. 2018, 29, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Viglietto, G.; Dolci, S.; Bruni, P.; Baldassarre, G.; Chiariotti, L.; Melillo, R.M.; Salvatore, G.; Chiappetta, G.; Sferratore, F.; Fusco, A.; et al. Glial cell line-derived neutrotrophic factor and neurturin can act as paracrine growth factors stimulating DNA synthesis of Ret-expressing spermatogonia. Int. J. Oncol. 2000, 16, 783. [Google Scholar] [CrossRef] [PubMed]
- Dettin, L.; Rubinstein, N.; Aoki, A.; Rabinovich, G.A.; Maldonado, C.A. Regulated expression and ultrastructural localization of galectin-1, a proapoptotic β-galactoside-binding lectin, during spermatogenesis in rat testis. Biol. Reprod. 2003, 68, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, M.C.; Braydich-Stolle, L.; Dym, M. Isolation of male germ-line stem cells; Influence of GDNF. Dev. Biol. 2005, 279, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Ebata, K.T.; Zhang, X.; Nagano, M.C. Expression patterns of cell-surface molecules on male germ line stem cells during postnatal mouse development. Mol. Reprod. Dev. 2005, 72, 171–181. [Google Scholar] [CrossRef]
- Grasso, M.; Fuso, A.; Dovere, L.; De Rooij, D.G.; Stefanini, M.; Boitani, C.; Vicini, E. Distribution of GFRA1-expressing spermatogonia in adult mouse testis. Reproduction 2012, 143, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Aiyama, Y.; Ishii-Inagaki, M.; Hara, K.; Tsunekawa, N.; Harikae, K.; Kanai, Y. Cyclical and patch-like GDNF distribution along the basal surface of Sertoli cells in mouse and hamster testes. PLoS ONE 2011, 6, e28367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuijk, E.W.; Colenbrander, B.; Roelen, B.A. The effects of growth factors on in vitro-cultured porcine testicular cells. Reproduction 2009, 138, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Campos-Junior, P.H.A.; Costa, G.M.J.; Lacerda, S.M.S.N.; Rezende-Neto, J.V.; de Paula, A.M.; Hofmann, M.-C.; de França, L.R. The Spermatogonial Stem Cell Niche in the Collared Peccary (Tayassu tajacu). Biol. Reprod. 2012, 86, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lara, N.L.M.; Costa, G.M.J.; Avelar, G.F.; Guimarães, D.A.; França, L.R. Postnatal testis development in the collared peccary (Tayassu tajacu), with emphasis on spermatogonial stem cells markers and niche. Gen. Comp. Endocrinol. 2019, 273, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Mahla, R.S.; Reddy, N.; Goel, S. Spermatogonial stem cells (SSCs) in buffalo (Bubalus bubalis) testis. PLoS ONE 2012, 7, e36020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, G.M.J.; Avelar, G.F.; Rezende-Neto, J.V.; Campos-Junior, P.H.A.; Lacerda, S.M.S.N.; Andrade, B.S.C.; Thomé, R.G.; Hofmann, M.-C.; Franca, L.R. Spermatogonial Stem Cell Markers and Niche in Equids. PLoS ONE 2012, 7, e44091. [Google Scholar] [CrossRef] [Green Version]
- Gassei, K.; Ehmcke, J.; Dhir, R.; Schlatt, S. Magnetic activated cell sorting allows isolation of spermatogonia from adult primate testes and reveals distinct GFRa1-positive subpopulations in men. J. Med. Primatol. 2010, 39, 83–91. [Google Scholar] [CrossRef]
- He, Z.; Kokkinaki, M.; Jiang, J.; Dobrinski, I.; Dym, M. Isolation, characterization, and culture of human spermatogonia. Biol. Reprod. 2010, 82, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Bosseboeuf, A.; Gautier, A.; Auvray, P.; Mazan, S.; Sourdaine, P. Characterization of spermatogonial markers in the mature testis of the dogfish (Scyliorhinus canicula L.). Reproduction 2014, 147, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, S.; Hayashi, M.; Kouguchi, T.; Yamaguchi, K.; Miwa, M.; Yoshizaki, G. Expression patterns of gdnf and gfrα1 in rainbow trout testis. Gene Expr. Patterns 2014, 14, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Bellaiche, J.; Lareyre, J.-J.; Cauty, C.; Yano, A.; Allemand, I.; Le Gac, F. Spermatogonial Stem Cell Quest: Nanos2, Marker of a Subpopulation of Undifferentiated A Spermatogonia in Trout Testis. Biol. Reprod. 2014, 90, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Maouche, A.; Curran, E.; Goupil, A.S.; Sambroni, E.; Bellaiche, J.; Le Gac, F.; Lareyre, J.J. New insights into the evolution, hormonal regulation, and spatiotemporal expression profiles of genes involved in the Gfra1/Gdnf and Kit/Kitlg regulatory pathways in rainbow trout testis. Fish Physiol. Biochem. 2018, 44, 1599–1616. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, Z.; Wang, Y.; Luo, Y.; Da, F.; Tao, W.; Zhou, L.; Wang, D.; Wei, J. Both Gfrα1a and Gfrα1b Are Involved in the Self-Renewal and Maintenance of Spermatogonial Stem Cells in Medaka. Stem Cells Dev. 2018, 27, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, L.; Fan, Z.; Hong, Y.; Zhao, Y.; Zhou, L.; Wang, D. Identification, prokaryote expression of medaka gdnfa/b and their biological activity in a spermatogonial cell line. Stem Cells Dev. 2017, 26, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, D.J.; Eisen, J.S. Headwaters of the zebrafish—emergence of a new model vertebrate. Nat. Rev. Genet. 2002, 3, 717–724. [Google Scholar] [CrossRef]
- Spence, R.; Fatema, M.K.; Reichard, M.; Huq, K.A.; Wahab, M.A.; Ahmed, Z.F.; Smith, C. The distribution and habitat preferences of the zebrafish in Bangladesh. J. Fish Biol. 2006, 69, 1435–1448. [Google Scholar] [CrossRef]
- Engeszer, R.E.; Da Barbiano, L.A.; Ryan, M.J.; Parichy, D.M. Timing and plasticity of shoaling behaviour in the zebrafish, Danio rerio. Anim. Behav. 2007, 74, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Hajam, Y.A.; Rani, R.; Sharma, P.; Kumar, R.; Verma, S.K. Zebrafish (Danio rerio): A Versatile Model for Reproductive Biology. In Recent Updates in Molecular Endocrinology and Reproductive Physiology of Fish; Springer: New York, NY, USA, 2021; pp. 105–120. [Google Scholar]
- Phillips, J.B.; Westerfield, M. Zebrafish models in translational research: Tipping the scales toward advancements in human health. Dis. Model Mech. 2014, 7, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ashton, B.; Cheung, M.; Heled, J.; Kearse, M.; Moir, R.; Wilson, A. Geneious v4.8.5. 2009. Available online: http://www.geneious.com (accessed on 24 December 2021).
- Flicek, P.; Amode, M.R.; Barrell, D.; Beal, K.; Brent, S.; Chen, Y.; Searle, S.M. Ensembl 2011. Nucleic Acids Res. 2010, 39 (Suppl. 1), D800–D806. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraldo, M.T.; Valente, G.T.; Braz, A.S.; Martins, C. The discovery of Foxl2 paralogs in chondrichthyan, coelacanth and tetrapod genomes reveals an ancient duplication in vertebrates. Heredity 2013, 111, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Nóbrega, R.H.; Greebe, C.D.; van de Kant, H.; Bogerd, J.; de França, L.R.; Schulz, R.W. Spermatogonial stem cell niche and spermatogonial stem cell transplantation in zebrafish. PLoS ONE 2010, 5, e12808. [Google Scholar] [CrossRef] [Green Version]
- Morais, R.D.V.S.; Crespo, D.; Nóbrega, R.H.; Lemos, M.S.; van de Kant, H.J.G.; de França, L.R.; Male, R.; Bogerd, J.; Schulz, R.W. Antagonistic regulation of spermatogonial differentiation in zebrafish (Danio rerio) by Igf3 and Amh. Mol. Cell. Endocrinol. 2017, 454, 112–124. [Google Scholar] [CrossRef] [Green Version]
- Tovo-Neto, A.; Martinez, E.R.; Melo, A.G.; Doretto, L.B.; Butzge, A.J.; Rodrigues, M.S.; Nóbrega, R.H. Cortisol Directly Stimulates Spermatogonial Differentiation, Meiosis, and Spermiogenesis in Zebrafish (Danio rerio) Testicular Explants. Biomolecules 2020, 10, 429. [Google Scholar] [CrossRef] [Green Version]
- García-López, Á.; De Jonge, H.; Nóbrega, R.H.; De Waal, P.P.; Van Dijk, W.; Hemrika, W.; Taranger, G.L.; Bogerd, J.; Schulz, R.W. Studies in zebrafish reveal unusual cellular expression patterns of gonadotropin receptor messenger ribonucleic acids in the testis and unexpected functional differentiation of the gonadotropins. Endocrinology 2010, 151, 2349–2360. [Google Scholar] [CrossRef] [Green Version]
- Hinfray, N.; Nóbrega, R.H.; Caulier, M.; Baudiffier, D.; Maillot-Marechal, E.; Chadili, E.; Brion, F. Cyp17a1 and Cyp19a1 in the zebrafish testis are differentially affected by oestradiol. J. Endocrinol. 2013, 216, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Leal, M.C.; de Waal, P.P.; García-López, Á.; Chen, S.X.; Bogerd, J.; Schulz, R.W. Zebrafish primary testis tissue culture: An approach to study testis function ex vivo. Gen. Comp. Endocrinol. 2009, 162, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Gautier, A.; Bosseboeuf, A.; Auvray, P.; Sourdaine, P. Maintenance of potential spermatogonial stem cells in vitro by GDNF treatment in a chondrichthyan model (Scyliorhinus canicula L.). Biol. Reprod. 2014, 91, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nóbrega, R.H.; De Souza Morais, R.D.V.; Crespo, D.; De Waal, P.P.; De França, L.R.; Schulz, R.W.; Bogerd, J. Fsh stimulates spermatogonial proliferation and differentiation in zebrafish via Igf3. Endocrinology 2015, 156, 3804–3817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, M.C.; Cardoso, E.R.; Nóbrega, R.H.; Batlouni, S.R.; Bogerd, J.; França, L.R.; Schulz, R.W. Histological and Stereological Evaluation of Zebrafish (Danio rerio) Spermatogenesis with an Emphasis on Spermatogonial Generations. Biol. Reprod. 2009, 81, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovell, S.C.; Davis, I.W.; Arendall III, W.B.; De Bakker, P.I.W.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Calpha geometry: phi, psi and Cbeta deviation. Proteins Struct. Funct. Genet. 2002, 50, 437–450. [Google Scholar] [CrossRef]
- Bucher, P. Weight matrix descriptions of four eukaryotic RNA polymerase II promoter elements derived from 502 unrelated promoter sequences. J. Mol. Biol. 1990, 212, 563–578. [Google Scholar] [CrossRef]
- Wang, Q.; Li, W.; Liu, X.S.; Carroll, J.S.; Jänne, O.A.; Keeton, E.K.; Chinnaiyan, A.M.; Pienta, K.J.; Brown, M. A Hierarchical Network of Transcription Factors Governs Androgen Receptor-Dependent Prostate Cancer Growth. Mol. Cell 2007, 27, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Lamberti, D.; Vicini, E. Promoter analysis of the gene encoding GDNF in murine Sertoli cells. Mol. Cell. Endocrinol. 2014, 394, 105–114. [Google Scholar] [CrossRef]
- Kuri, P.; Ellwanger, K.; Kufer, T.A.; Leptin, M.; Bajoghli, B. A high-sensitivity bi-directional reporter to monitor NF-κB activity in cell culture and zebrafish in real time. J. Cell Sci. 2017, 130, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.P.; Correard, S.; Gheorghe, M.; Baranašić, D.; et al. JASPAR 2020: Update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2020, 48, D87–D92. [Google Scholar] [CrossRef]
- França, L.R.; Nóbrega, R.H.; Morais, R.D.V.S.; De Castro Assis, L.H.; Schulz, R.W. Sertoli cell structure and function in anam niote vertebrates. In Sertoli Cell Biology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 385–407. [Google Scholar]
- Crespo, D.; Assis, L.H.; Furmanek, T.; Bogerd, J.; Schulz, R.W. Expression profiling identifies Sertoli and Leydig cell genes as Fsh targets in adult zebrafish testis. Mol. Cell. Endocrinol. 2016, 437, 237–251. [Google Scholar] [CrossRef]
- Skaar, K.S.; Nóbrega, R.H.; Magaraki, A.; Olsen, L.C.; Schulz, R.W.; Male, R. Proteolytically activated, recombinant anti-Müllerian hormone inhibits androgen secretion, proliferation, and differentiation of spermatogonia in adult zebrafish testis organ cultures. Endocrinology 2011, 152, 3527–3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.M.; Zhang, Q.; Zhang, Q.; Zhu, W.; He, C.; Lu, C.L.; Chen, Z.Y. Identification of the key amino acids of glial cell line-derived neurotrophic factor family receptor α1 involved in its biological function. J. Biol. Chem. 2004, 279, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airaksinen, M.S.; Holm, L.; Hätinen, T. Evolution of the GDNF family ligands and receptors. Brain Behav. Evol. 2006, 68, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. Structural studies of GDNF family ligands with their receptors—Insights into ligand recognition and activation of receptor tyrosine kinase RET. Biochim. Biophys. Acta Proteins Proteom. 2013, 1834, 2205–2212. [Google Scholar] [CrossRef] [PubMed]
- Maouche, A. Caractérisation de Voies de Régulation Paracrine Potentiellement Impliquées Dans le Devenir des Cellules Souches Spermatogoniales Chez la Truite arc-en-ciel (Oncorhynchus mykiss). Ph.D. Thesis, Université Bretagne Loire (COMUE), Rennes, France, 2018. [Google Scholar]
- Hoegg, S.; Brinkmann, H.; Taylor, J.S.; Meyer, A. Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J. Mol. Evol. 2004, 59, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Van de Peer, Y. From 2R to 3R: Evidence for a fish-specific genome duplication (FSGD). Int. J. Bioassays 2005, 27, 937–945. [Google Scholar] [CrossRef] [Green Version]
- Buaas, F.W.; Kirsh, A.L.; Sharma, M.; McLean, D.J.; Morris, J.L.; Griswold, M.D.; Braun, R.E. Plzf is required in adult male germ cells for stem cell self-renewal. Nat. Genet. 2004, 36, 647–652. [Google Scholar] [CrossRef]
- Costoya, J.A.; Hobbs, R.M.; Barna, M.; Cattoretti, G.; Manova, K.; Sukhwani, M.; Pandolfi, P.P. Essential role of Plzf in maintenance of spermatogonial stem cells. Nat. Genet. 2004, 36, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Lacerda, S.M.S.N.; Costa, G.M.J.; da Silva, M.D.A.; Campos-Junior, P.H.A.; Segatelli, T.M.; Peixoto, M.T.D.; de França, L.R. Phenotypic characterization and in vitro propagation and transplantation of the Nile tilapia (Oreochromis niloticus) spermatogonial stem cells. Gen. Com. Endocrinol. 2013, 192, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Schulz, R.W.; Menting, S.; Bogerd, J.; França, L.R.; Vilela, D.A.; Godinho, H.P. Sertoli cell proliferation in the adult testis—evidence from two fish species belonging to different orders. Biol. Reprod. 2005, 73, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Inoue, K.; Takashima, S.; Takehashi, M.; Ogonuki, N.; Morimoto, H.; Shinohara, T. Reconstitution of mouse spermatogonial stem cell niches in culture. Cell Stem Cell 2012, 11, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Templeman, J.L.; Smith, R.A.; Mackay, S. Effects of glial cell line-derived neurotrophic factor on isolated developing mouse Sertoli cells in vitro. J. Anat. 2005, 206, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, H.; Shi, Y.; Pan, Z.; Wang, J. Activation of the GFRa1/NCAM pathway stimulates Sertoli cell proliferation in vitro. Belg. J. Zool. 2008, 138, 177–183. [Google Scholar]
- Safian, D.; Ryane, N.; Bogerd, J.; Schulz, R.W. Fsh stimulates Leydig cell Wnt5a production, enriching zebrafish type A spermatogonia. J. Endocrinol. 2018, 239, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Safian, D.; Bogerd, J.; Schulz, R.W. Regulation of spermatogonial development by Fsh: The complementary roles of locally produced Igf and Wnt signaling molecules in adult zebrafish testis. Gen. Comp. Endocrinol. 2019, 284, 113244. [Google Scholar] [CrossRef]
- Simon, L.; Ekman, G.C.; Tyagi, G.; Hess, R.A.; Murphy, K.M.; Cooke, P.S. Common and distinct factors regulate expression of mRNA for ETV5 and GDNF, Sertoli cell proteins essential for spermatogonial stem cell maintenance. Exp. Cell Res. 2007, 313, 3090–3099. [Google Scholar] [CrossRef]
- Walker, W.H.; Fucci, L.I.N.D.A.; Habener, J.F. Expression of the gene encoding transcription factor cyclic adenosine 3′, 5′-monophosphate (cAMP) response element-binding protein (CREB): Regulation by follicle-stimulating hormone-induced cAMP signaling in primary rat Sertoli cells. Endocrinology 1995, 136, 3534–3545. [Google Scholar] [CrossRef]
- Garcia, T.X.; Parekh, P.; Gandhi, P.; Sinha, K.; Hofmann, M.C. The NOTCH ligand JAG1 regulates GDNF expression in Sertoli cells. Stem Cells Dev. 2017, 26, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Tanigaki, K.; Honjo, T. Two opposing roles of RBP-J in Notch signaling. Curr. Top. Dev. Biol. 2010, 92, 231–252. [Google Scholar]
- Iso, T.; Kedes, L.; Hamamori, Y. HES and HERP families: Multiple effectors of the Notch signaling pathway. J. Cell Physiol. 2003, 194, 237–255. [Google Scholar] [CrossRef]
- Kageyama, R.; Ohtsuka, T.; Kobayashi, T. The Hes gene family: Repressors and oscillators that orchestrate embryogenesis. Development 2007, 134, 1243–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, S.; Bernard, F. Notch targets and their regulation. Curr. Top. Dev. Biol. 2010, 92, 253–275. [Google Scholar] [PubMed]
- Lin, Q.; Mei, J.; Li, Z.; Zhang, X.; Zhou, L.; Gui, J.F. Distinct and cooperative roles of amh and dmrt1 in self-renewal and differentiation of male germ cells in zebrafish. Genetics 2017, 207, 1007–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adolfi, M.C.; Nakajima, R.T.; Nóbrega, R.H.; Schartl, M. Intersex, hermaphroditism, and gonadal plasticity in vertebrates: Evolution of the Müllerian duct and Amh/Amh5r2 signaling. Ann. Rev. Anim. Biosci. 2019, 7, 149–172. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Primer Sequences (5′–3′) | References |
|---|---|---|
| ef1α | GCCGTCCCACCGACAAG (Fw) | Morais et al. [48] |
| CCACACGACCCACAGGTACAG (Rv) | ||
| b-actin | AGACATCAGGGAGTGATGGT (Fw) | Tovo-Neto et al. [49] |
| CAATACCGTGCTCAATGGGG (Rv) | ||
| gdnfa | GAAGCTCCGGTCTGTATGGA (Fw) | This paper |
| GGAGCTCAGGAGCAACAAAC (Rv) | ||
| gdnfb | AGGAGTAAATCAGTGGGCCAAA (Fw) | This paper |
| AGTAGCTGAATATGAGCTCCTCC (Rv) | ||
| gfrα1a | TCGACTGGCTCCCATCTATTC (Fw) | This paper |
| AGGTGTCATTCAGGTTGCAGG (Rv) | ||
| gfrα1b | CCTGTGCTTGATTTAGTGCA (Fw) | This paper |
| GCATCCGTACTTTCCCAAAC (Rv) | ||
| igf3 | TGTGCGGAGACAGAGGCTTT (Fw) | Morais et al. [48] |
| CGCCGCACTTTCTTGGATT (Rv) | ||
| amh | CTCTGACCTTGATGAGCCTCATTT (Fw) | García-Lopez et al. [50] |
| GGATGTCCCTTAAGAACTTTTGCA (Rv) | ||
| fshr | GAGGATTCCCAGTAATGCTTTCCT (Fw) | García-Lopez et al. [50] |
| TCTATCTCACGAATCCCGTTCTTC (Rv) | ||
| pou5f3 | GAGAGATGTAGTGCGTGTAT (Fw) | Tovo-Neto et al. [49] |
| GCTCGTAATACTGTGCTTCA (Rv) | ||
| dazl | AGTGCAGACTTTGCTAACCCTTATGTA (Fw) | Morais et al. [49] |
| GTCCACTGCTCCAAGTTGCTCT (Rv) | ||
| sycp3l | AGAAGCTGACCCAAGATCATTCC (Fw) | García-Lopez et al. [50] |
| AGCTTCAGTTGCTGGCGAAA (Rv) | ||
| gdnfa-ish | T7Rpps-CCGCAGTGAGAGCCCCG (Fw) | This paper |
| T3Rpps-TCCCGTTAGGTCATATTGTTCCTC (Rv) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doretto, L.B.; Butzge, A.J.; Nakajima, R.T.; Martinez, E.R.M.; de Souza, B.M.; Rodrigues, M.d.S.; Rosa, I.F.; Ricci, J.M.B.; Tovo-Neto, A.; Costa, D.F.; et al. Gdnf Acts as a Germ Cell-Derived Growth Factor and Regulates the Zebrafish Germ Stem Cell Niche in Autocrine- and Paracrine-Dependent Manners. Cells 2022, 11, 1295. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11081295
Doretto LB, Butzge AJ, Nakajima RT, Martinez ERM, de Souza BM, Rodrigues MdS, Rosa IF, Ricci JMB, Tovo-Neto A, Costa DF, et al. Gdnf Acts as a Germ Cell-Derived Growth Factor and Regulates the Zebrafish Germ Stem Cell Niche in Autocrine- and Paracrine-Dependent Manners. Cells. 2022; 11(8):1295. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11081295
Chicago/Turabian StyleDoretto, Lucas B., Arno J. Butzge, Rafael T. Nakajima, Emanuel R. M. Martinez, Beatriz Marques de Souza, Maira da Silva Rodrigues, Ivana F. Rosa, Juliana M. B. Ricci, Aldo Tovo-Neto, Daniel F. Costa, and et al. 2022. "Gdnf Acts as a Germ Cell-Derived Growth Factor and Regulates the Zebrafish Germ Stem Cell Niche in Autocrine- and Paracrine-Dependent Manners" Cells 11, no. 8: 1295. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11081295