
Role of Th17 Cytokines in the Liver’s Immune Response during Fatal Yellow Fever: Triggering Cell Damage Mechanisms

 ,
,  ,
,  , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Methods
2.1. Study Design
2.2. Patients, Samples, and Diagnosis of YFV Infection
2.3. Histology and Semi-Quantitative Analysis
2.4. Immunohistochemistry
2.5. Quantitative Analysis and Photo-Documentation
2.6. Statistical Analysis
2.7. Ethical Aspects
2.8. Availability of Data and Materials
3. Results
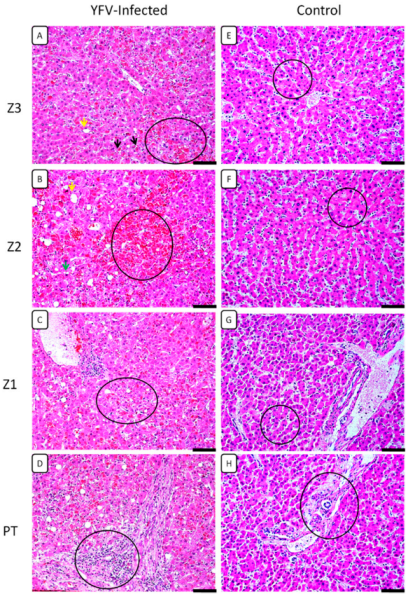
3.1. Histopathological Analysis
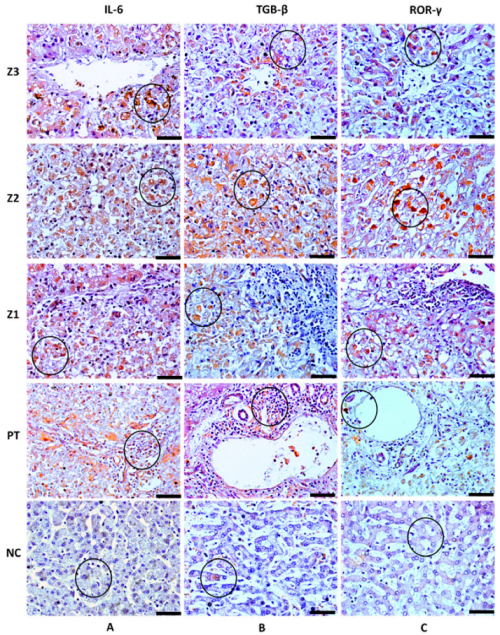
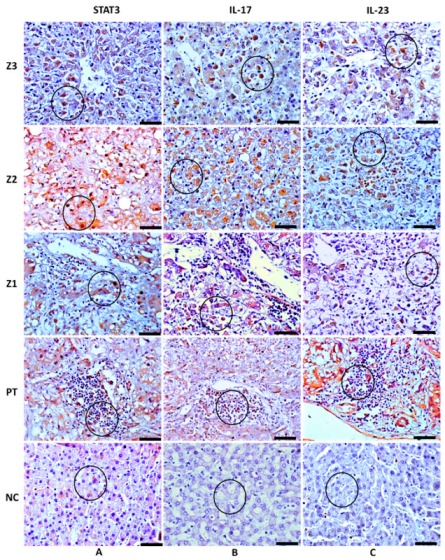
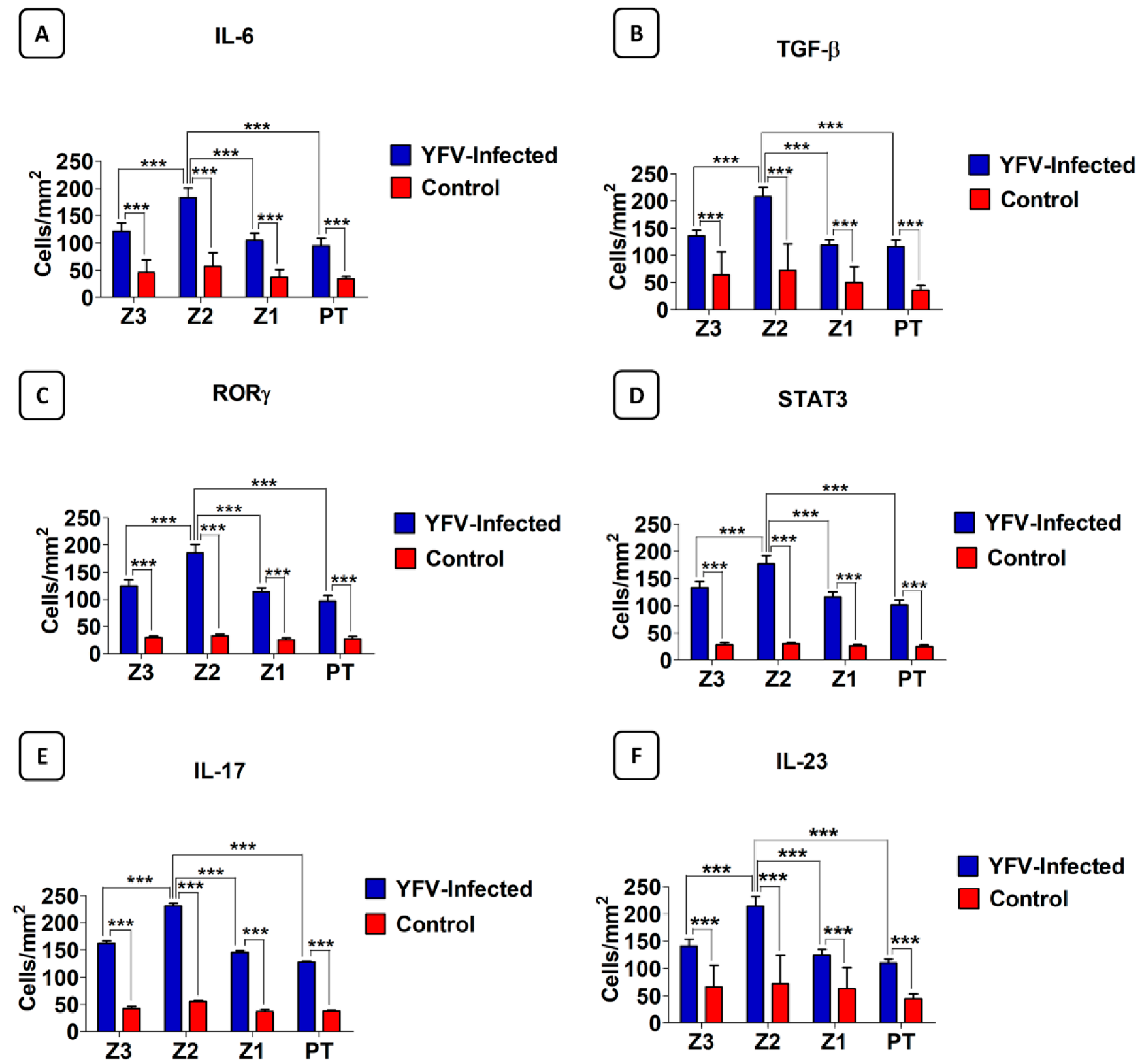
3.2. Analysis of the Response to the Th17 Profile in the Hepatic Parenchyma in Cases with Fatal YF
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McArthur, M.A.; Zhang, S.L.; Li, L.; Tesh, R.B.; Barrett, A.D. Molecular Characterization of Hamster-Adapted Yellow Fever Virus. Vector Borne Zoonotic Dis. 2020, 20, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Gardner, C.L.; Ryman, K.D. Yellow fever: A reemerging threat. (Special Issue: Emerging pathogens.). Clin. Lab. Med. 2010, 30, 237–260. [Google Scholar] [CrossRef] [Green Version]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide Sequence of Yellow Fever Virus: Implications for Flavivirus Gene Expression and Evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, T.J.; McCourt, D.W.; Rice, C.M. Production of yellow fever virus proteins in infected cells: Identification of discrete polyprotein species and analysis of cleavage kinetics using region-specific polyclonal antisera. Virology 1990, 177, 159–174. [Google Scholar] [CrossRef]
- Ferreira, M.S.; Júnior, P.S.B.; Cerqueira, V.D.; Rivero, G.R.C.; Júnior, C.A.O.; Castro, P.H.G.; Da Silva, G.A.; Da Silva, W.B.; Imbeloni, A.A.; Sousa, J.R.; et al. Experimental yellow fever virus infection in the squirrel monkey (Saimiri spp.) I: Gross anatomical and histopathological findings in organs at necropsy. Mem. Inst. Oswaldo Cruz. 2020, 115, e190501. [Google Scholar] [CrossRef] [PubMed]
- Olímpio, F.A.; Falcão, L.F.M.; Carvalho, M.L.G.; Lopes, J.D.C.; Mendes, C.C.H.; Filho, A.J.M.; da Silva, C.A.M.; Miranda, V.D.S.C.; dos Santos, L.C.; Vilacoert, F.S.d.S.; et al. Endothelium Activation during Severe Yellow Fever Triggers an Intense Cytokine-Mediated Inflammatory Response in the Liver Parenchyma. Pathogens 2022, 11, 101. [Google Scholar] [CrossRef]
- Lopes, R.L.; Pinto, J.R.; Junior, G.B.D.S.; Santos, A.K.T.; Souza, M.T.O.; Daher, E.; Da Silva, G.B. Kidney involvement in yellow fever: A review. Rev. Inst. Med. Trop. São Paulo 2019, 61, e35. [Google Scholar] [CrossRef] [PubMed]
- Ministério da Saúde (MS). Secretaria de Vigilância em Saúde. Emergência epidemiológica de febre amarela no Brasil, no período de dezembro de 2016 a julho de 2017. Bol. Epidemiol. 2017, 48, 1–22. [Google Scholar]
- Faria, N.R.; Kraemer, M.U.G.; Hill, S.C.; De Jesus, J.G.; Aguiar, R.S.; Iani, F.C.M.; Xavier, J.; Quick, J.; Du Plessis, L.; Dellicour, S.; et al. Genomic and epidemiological monitoring of yellow fever virus transmission potential. Science 2018, 361, 894–899. [Google Scholar] [CrossRef] [Green Version]
- Giovanetti, M.; De Mendonça, M.C.L.; Fonseca, V.; Mares-Guia, M.A.; Fabri, A.; Xavier, J.; De Jesus, J.G.; Gräf, T.; Rodrigues, C.D.D.S.; dos Santos, C.C.; et al. Yellow Fever Virus Reemergence and Spread in Southeast Brazil, 2016–2019. J. Virol. 2019, 94, e01623-19. [Google Scholar] [CrossRef]
- Quaresma, J.A.S.; Barros, V.L.R.S.; Fernandes, E.R.; Pagliari, C.; Guedes, F.; Vasconcelos, P.F.D.C.; Junior, H.F.D.A.; Duarte, M.I.S. Immunohistochemical examination of the role of Fas ligand and lymphocytes in the pathogenesis of human liver yellow fever. Virus Res. 2006, 116, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Quaresma, J.A.; Barros, V.L.; Pagliari, C.; Fernandes, E.R.; Guedes, F.; Takakura, C.F.; Andrade, H.F., Jr.; Vasconcelos, P.F.; Duarte, M.I. Revisiting the liver in human yellow fever: Virus-induced apoptosis in hepatocytes associated with TGF-β, TNF-α and NK cells activity. Virology 2006, 345, 22–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaresma, J.A.S.; Barros, V.L.R.S.; Fernandes, E.R.; Pagliari, C.; Takakura, C.; Vasconcelos, P.F.D.C.; de Andrade, H.F.; Duarte, M.I.S. Reconsideration of histopathology and ultrastructural aspects of the human liver in yellow fever. Acta Trop. 2005, 94, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Quaresma, J.A.S.; Pagliari, C.; Medeiros, D.B.A.; Duarte, M.I.S.; Vasconcelos, P.F.C. Immunity and immune response, pathology and pathologic changes: Progress and challenges in the immunopathology of yellow fever. Rev. Med. Virol. 2013, 23, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P. Yellow fever: An update. Lancet Infect. Dis. 2001, 1, 11–20. [Google Scholar] [CrossRef]
- Patel, D.D.; Kuchroo, V.K. Th17 Cell Pathway in Human Immunity: Lessons from Genetics and Therapeutic Interventions. Immunity 2015, 43, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Miossec, P.; Kolls, J.K. Targeting IL-17 and TH17 cells in chronic inflammation. Nat. Rev. Drug Discov. 2012, 11, 763–776. [Google Scholar] [CrossRef]
- Bystrom, J.; Clanchy, F.I.L.; Taher, T.E.; Al-Bogami, M.; Ong, V.H.; Abraham, D.J.; Williams, R.O.; Mageed, R.A. Functional and phenotypic heterogeneity of Th17 cells in health and disease. Eur. J. Clin. Investig. 2018, 49, e13032. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wei, C.; Xu, H.; Jia, J.; Wei, Z.; Guo, R.; Jia, Y.; Wu, Y.; Li, Y.; Qi, X.; et al. The Immunoregulation of Th17 in Host against Intracellular Bacterial Infection. Mediat. Inflamm. 2018, 2018, 6587296. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, R.S.S.; De Sousa, J.R.; Araujo, M.T.F.; Filho, A.J.M.; De Alcantara, B.N.; Araujo, F.M.C.; Queiroz, M.G.L.; Cruz, A.C.R.; Vasconcelos, B.H.B.; Chiang, J.O.; et al. In situ immune response and mechanisms of cell damage in central nervous system of fatal cases microcephaly by Zika virus. Sci. Rep. 2018, 8, 1. [Google Scholar] [CrossRef]
- Hernández-Santos, N.; Gaffen, S.L. Th17 Cells in Immunity to Candida albicans. Cell Host Microbe 2012, 11, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Vargas, L.A.; Hernández-Flores, K.G.; Thomas-Dupont, P.; Izaguirre-Hernández, I.Y.; Sánchez-Marce, E.E.; Remes-Ruiz, R.; Fonseca-Coronado, S.; Hernández-Romano, P.A.; Flores-Collins, M.E.; Vivanco-Cid, H. Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections. Viruses 2020, 12, 1435. [Google Scholar] [CrossRef] [PubMed]
- Shearer, F.M.; Moyes, C.; Pigott, D.M.; Brady, O.J.; Marinho, F.; Deshpande, A.; Longbottom, J.; Browne, A.J.; Kraemer, M.U.G.; O'Reilly, K.; et al. Global yellow fever vaccination coverage from 1970 to 2016: An adjusted retrospective analysis. Lancet Infect. Dis. 2017, 17, 1209–1217. [Google Scholar] [CrossRef] [Green Version]
- Paules, C.I.; Fauci, A.S. Yellow Fever—Once Again on the Radar Screen in the Americas. N. Engl. J. Med. 2017, 376, 1397–1399. [Google Scholar] [CrossRef]
- Tranquilin, M.V.; Lehmkühl, R.C.; Maron, A.; Da Silva, L.R.; Ziliotto, L.; Seki, M.C.; Salomon, G.R.; Carrasco, A.D.O.T. First report of yellow fever virus in non-human primates in the State of Parana, Brazil. Rev. Soc. Bras. Med. Trop. 2013, 46, 522–524. [Google Scholar] [CrossRef] [Green Version]
- Monath, T.P.; Vasconcelos, P.F. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- Holanda, G.M.; Casseb, S.M.M.; Quaresma, J.A.S.; Vasconcelos, P.F.C.; Cruz, A.C.R. Yellow fever virus modulates cytokine mRNA expression and induces activation of caspase 3/7 in the human hepatocarcinoma cell line HepG2. Arch. Virol. 2019, 164, 1187–1192. [Google Scholar] [CrossRef]
- Douam, F.; Ploss, A. Yellow Fever Virus: Knowledge Gaps Impeding the Fight Against an Old Foe. Trends Microbiol. 2018, 26, 913–928. [Google Scholar] [CrossRef]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Liu, J.; Xu, Y.; Cao, H. Role of macrophages in experimental liver injury and repair in mice (Review). Exp. Ther. Med. 2019, 17, 3835–3847. [Google Scholar] [CrossRef] [Green Version]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Heide, D.; Weiskirchen, R.; Bansal, R. Therapeutic Targeting of Hepatic Macrophages for the Treatment of Liver Diseases. Front. Immunol. 2019, 10, 2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabregat, I.; Caballero-Díaz, D. Transforming Growth Factor-β-Induced Cell Plasticity in Liver Fibrosis and Hepatocarcinogenesis. Front. Oncol. 2018, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Mallat, A.; Lotersztajn, S. Cellular Mechanisms of Tissue Fibrosis. 5. Novel insights into liver fibrosis. Am. J. Physiol. Physiol. 2013, 305, C789–C799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Arras, D.; Rose-John, S. IL-6 pathway in the liver: From physiopathology to therapy. J. Hepatol. 2016, 64, 1403–1415. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Zhang, Z.; Wang, F.-S. Liver fibrosis: Mechanisms of immune-mediated liver injury. Cell. Mol. Immunol. 2011, 9, 296–301. [Google Scholar] [CrossRef]
- Póvoa, T.F.; Oliveira, E.R.; Basílio-De-Oliveira, C.A.; Nuovo, G.J.; Chagas, V.L.A.; Salomão, N.G.; Mota, E.M.; Paes, M. Peripheral Organs of Dengue Fatal Cases Present Strong Pro-Inflammatory Response with Participation of IFN-Gamma-, TNF-Alpha- and RANTES-Producing Cells. PLoS ONE 2016, 11, e0168973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, B.N.; Isaza, D.M.; Salazar, C.L.; Ramírez, R.; Ospina, M.; Alvarez, L.G. Serum levels of interleukin-6, tumor necrosis factor-alpha and interferon-gama in infants with and without dengue. Rev. Soc. Bras. Med. Trop. 2008, 41, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Iani, F.C.; Antonelli, L.R.; Gollob, K.J.; Costa, P.A.C.; Duarte, M.M.; Caldas, S.; Cecílio, A.B.; Cury, A.L.F. Dengue Patients with Early Hemorrhagic Manifestations Lose Coordinate Expression of the Anti-Inflammatory Cytokine IL-10 with the Inflammatory Cytokines IL-6 and IL-8. Am. J. Trop. Med. Hyg. 2016, 95, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Qi, Y.-F.; Yu, Y.-R. STAT3: A key regulator in liver fibrosis. Ann. Hepatol. 2020, 21, 100224. [Google Scholar] [CrossRef]
- Chua, J.J.-E.; Bhuvanakantham, R.; Chow, V.T.-K.; Ng, M.-L. Recombinant non-structural 1 (NS1) protein of dengue-2 virus interacts with human STAT3β protein. Virus Res. 2005, 112, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, J.D.; Fernandes-Siqueira, L.O.; Barbosa, G.M.; Da Poian, A.T. Non-Canonical Roles of Dengue Virus Non-Structural Proteins. Viruses 2017, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Pandey, N.; Garg, R.K.; Kumar, R. IL-17 Level in Patients with Dengue Virus Infection & its Association with Severity of Illness. J. Clin. Immunol. 2012, 33, 613–618. [Google Scholar] [CrossRef]
- Guabiraba, R.; Besnard, A.-G.; Marques, R.E.; Maillet, I.; Fagundes, C.T.; Conceição, T.M.; Rust, N.M.; Charreau, S.; Paris, I.; Lecron, J.-C.; et al. IL-22 modulates IL-17A production and controls inflammation and tissue damage in experimental dengue infection. Eur. J. Immunol. 2013, 43, 1529–1544. [Google Scholar] [CrossRef]
- McAleer, J.P.; Kolls, J.K. Mechanisms controlling Th17 cytokine expression and host defense. J. Leukoc. Biol. 2011, 90, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, M.; Li, Y.; He, H.; Ding, H.; Chen, K.; Du, J.; Chen, T.; Wu, Z.; Liu, H.; Wang, D.; et al. IL-23 production of liver inflammatory macrophages to damaged hepatocytes promotes hepatocellular carcinoma development after chronic hepatitis B virus infection. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2018, 1864, 3759–3770. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Sex | Age (Years) | Geographic Location |
|---|---|---|---|
| 1 | F | 03 | GO |
| 2 | F | 36 | GO |
| 3 | M | 56 | DF |
| 4 | M | 48 | MT |
| 5 | F | 50 | MG |
| 6 | M | 38 | MG |
| 7 | - | - | - |
| 8 | M | 39 | MG |
| 9 | M | 28 | MG |
| 10 | F | - | MG |
| 11 | M | 33 | MG |
| 12 | - | - | - |
| 13 | M | 23 | GO |
| 14 | M | 54 | GO |
| 15 | M | 32 | DF |
| 16 | M | 24 | GO |
| 17 | F | 63 | GO |
| 18 | M | 55 | DF |
| 19 | M | 46 | DF |
| 20 | M | 35 | DF |
| 21 | M | - | - |
| Morphological Changes | Z3 | Z2 | Z1 | PT |
|---|---|---|---|---|
| Cell swelling | 2++ | +++ | +++ | - |
| Macrovesicular steatosis | + | ++ | + | - |
| Microvesicular steatosis | ++ | +++ | ++ | - |
| Lytic necrosis | + | ++ | ++ | - |
| Coagulative necrosis | + | + | + | - |
| Councilman bodies | +++ | +++ | +++ | - |
| Kupffer cell hyperplasia | + | ++ | ++ | - |
| Kupffer cell hypertrophy | + | ++ | ++ | - |
| Oedema | - | - | - | ++ |
| Congestion | - | - | - | ++ |
| Inflammatory Infiltrate | ||||
| Lymphocytes | + | ++ | + | ++ |
| Neutrophils | + | + | + | + |
| Plasma cells | + | ++ | + | ++ |
| Eosinophils | + | + | + | + |
| Macrophages | - | - | - | ++ |
| Sinusoidal endothelial alteration | ++ | ++ | ++ | - |
| Alteration centrilobular vein | +++ | + | + | - |
| Sinusoidal congestion | + | ++ | + | - |
| Cholestasis | + | + | + | + |
| Sinusoidal dilatation | + | + | + | - |
| Portal vein alteration | - | - | - | ++ |
| Portal artery alteration | - | - | - | ++ |
| Bile canaliculus alteration | - | - | - | + |
| Limiting plaque injury | - | - | - | + |
| Markers | Z3 (Cells/mm2) | Z2 (Cells/mm2) | Z1 (Cells/mm2) | PT (Cells/mm2) | ANOVA (p ≤ 0.05) |
|---|---|---|---|---|---|
| IL-17A Control | 162.20 ± 18.81 42.56 ± 7.72 | 230.90 ± 22.63 55.36 ± 4.32 | 145.80 ± 13.36 36.80 ± 8.23 | 128.10 ± 7.20 37.76 ± 3.85 | *** |
| Tukey (p ≤ 0.05) | *** | *** | *** | *** | |
| IL-23 | 140.80 ± 12.77 | 213.80 ± 17.54 | 124.60 ± 10.05 | 109.80 ± 7.46 | *** |
| Control | 66.24 ± 39.15 | 71.36 ± 52.59 | 62.72 ± 38.77 | 44.16 ± 9.37 | |
| Tukey (p ≤ 0.05) | *** | *** | *** | *** | |
| IL-6 | 121.10 ± 15.78 | 182.60 ± 17.91 | 104.80 ± 12.64 | 94.32 ± 14.69 | *** |
| Control | 45.44 ± 23.61 | 56.96 ± 25.21 | 36.80 ± 14.49 | 33.92 ± 4.58 | |
| Tukey (p ≤ 0.05) | *** | *** | *** | *** | |
| STAT3 | 133.20 ± 11.15 | 176.90 ± 15.22 | 115.70 ± 8.95 | 101.40 ± 8.82 | *** |
| Control | 28.16 ± 3.50 | 29.76 ± 2.42 | 25.92 ± 2.62 | 24.96 ± 2.90 | |
| Tukey (p ≤ 0.05) | *** | *** | *** | *** | |
| RORγ | 124.30 ± 11.20 | 185.10 ± 15.42 | 113.40 ± 7.67 | 96.15 ± 10.67 | *** |
| Control | 29.44 ± 2.67 | 32.32 ± 3.46 | 25.28 ± 3.46 | 26.88 ± 4.71 | |
| Tukey (p ≤ 0.05) | *** | *** | *** | *** | |
| TGF-β | 135.90 ± 9.64 | 207.40 ± 17.86 | 119.20 ± 10.03 | 115.90 ± 12.10 | *** |
| Control | 63.68 ± 42.85 | 72.32 ± 48.31 | 49.60 ± 29.18 | 35.52 ± 9.28 | |
| Tukey (p ≤ 0.05) | *** | *** | *** | *** |
| Markers | Z2 vs. Z1 (Cells/mm2) | Tukey p | Z2 vs. Z3 (Cells/mm2) | Tukey p | Z2 vs. PT (Cells/mm2) | p Tukey |
|---|---|---|---|---|---|---|
| IL-17A | 230.90 ± 22.63 vs.145.80 ± 13.36 | *** | 230.90 ± 22.63 vs.162.20 ± 18.81 | *** | 230.90 ± 22.63 vs.128.10 ± 7.200 | *** |
| IL-23 | 213.80 ± 17.54 vs.124.60 ± 10.05 | *** | 213.80 ± 17.54 vs.140.80 ± 12.77 | *** | 213.80 ± 17.54 vs.109.80 ± 7.461 | *** |
| IL-6 | 182.60 ± 17.91 vs.104.80 ± 12.64 | *** | 182.60 ± 17.91 vs.121.10 ± 15.78 | *** | 182.60 ± 17.91 vs.94.32 ± 14.69 | *** |
| STAT3 | 176.90 ± 15.22 vs.115.70 ± 8.956 | *** | 176.90 ± 15.22 vs.133.20 ± 11.15 | *** | 176.90 ± 15.22 vs.101.40 ± 8.82 | *** |
| RORγ | 185.10 ± 15.42 vs.113.40 ± 7.67 | *** | 185.10 ± 15.42 vs.124.30 ± 11.20 | *** | 185.10 ± 15.42 vs.96.15 ± 10.67 | *** |
| TGF-β | 207.40 ± 17.86 vs.119.20 ± 10.03 | *** | 207.40 ± 17.86 vs.135.90 ± 9.646 | *** | 207.40 ± 17.86 vs.115.90± 12.10 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, M.L.G.; Falcão, L.F.M.; Lopes, J.d.C.; Mendes, C.C.H.; Olímpio, F.A.; Miranda, V.d.S.C.; Santos, L.C.d.; de Moraes, D.D.P.; Bertonsin Filho, M.V.; da Costa, L.D.; et al. Role of Th17 Cytokines in the Liver’s Immune Response during Fatal Yellow Fever: Triggering Cell Damage Mechanisms. Cells 2022, 11, 2053. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11132053
Carvalho MLG, Falcão LFM, Lopes JdC, Mendes CCH, Olímpio FA, Miranda VdSC, Santos LCd, de Moraes DDP, Bertonsin Filho MV, da Costa LD, et al. Role of Th17 Cytokines in the Liver’s Immune Response during Fatal Yellow Fever: Triggering Cell Damage Mechanisms. Cells. 2022; 11(13):2053. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11132053
Chicago/Turabian StyleCarvalho, Marcos Luiz Gaia, Luiz Fábio Magno Falcão, Jeferson da Costa Lopes, Caio Cesar Henriques Mendes, Fábio Alves Olímpio, Vanessa do Socorro Cabral Miranda, Lais Carneiro dos Santos, Daniel Dias Pinheiro de Moraes, Marcos Virgilio Bertonsin Filho, Luccas Delgado da Costa, and et al. 2022. "Role of Th17 Cytokines in the Liver’s Immune Response during Fatal Yellow Fever: Triggering Cell Damage Mechanisms" Cells 11, no. 13: 2053. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11132053