
Functional Role of STIM-1 and Orai1 in Human Microvascular Aging
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Tissues
2.2. Functional Evaluation of Human Mesenteric Arteries (MA)
2.3. Functional Evaluation of Human Aorta
2.4. Immunofluorescence Assay
2.5. Western Blot Analysis
3. Results
3.1. Orai Channel Inhibition Reverses Age-Related Adrenergic Hypercontractility of Mesenteric Arteries but Has No Functional Effect on the Aorta from Older Human Subjects
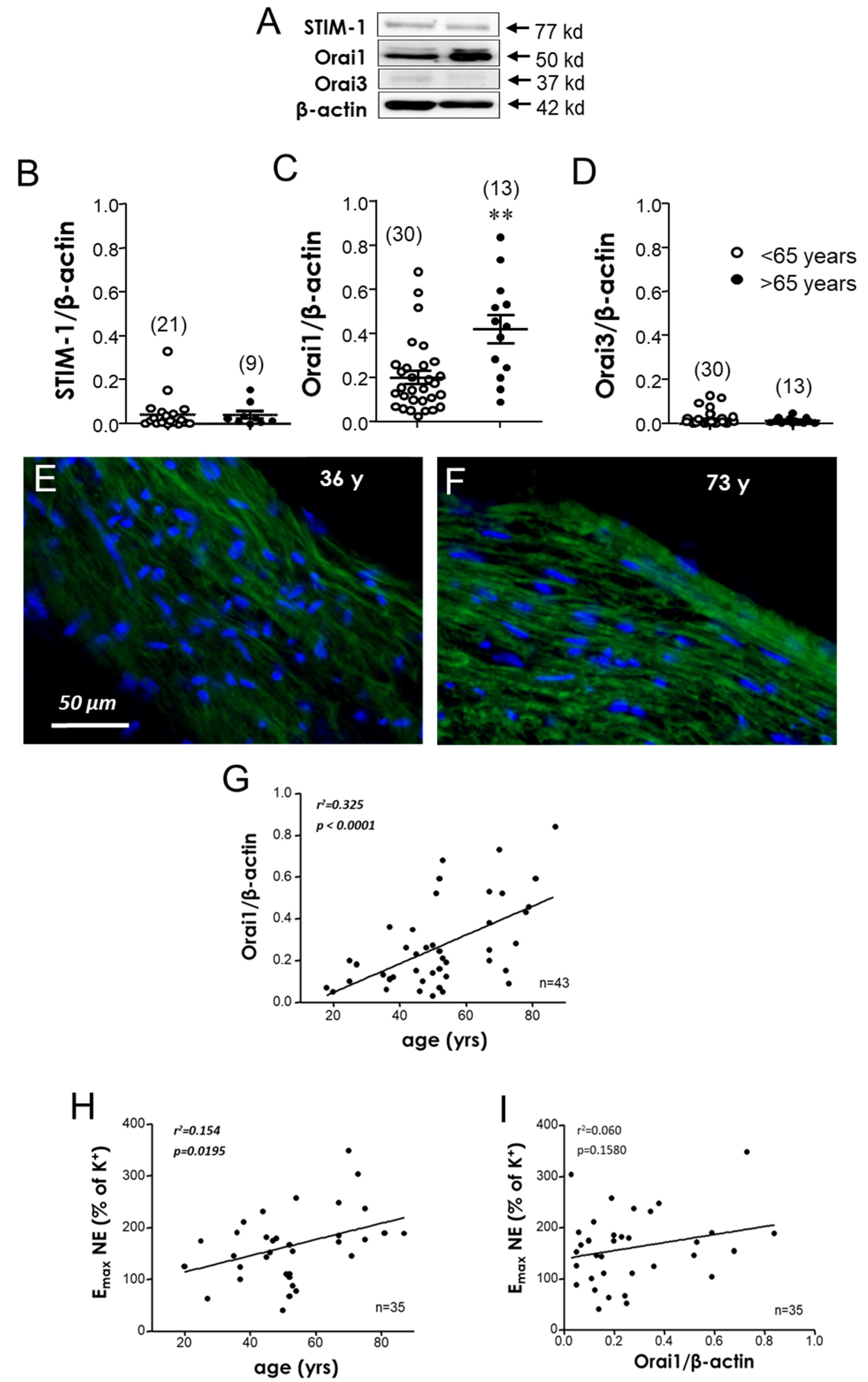
3.2. Orai1 Expression Is Increased in Aorta Derived from Aged Subjects
3.3. Human Aging Is Associated with an Increased Expression of STIM-1 and Orai1 in MA
3.4. Plasma Orai1 Concentrations Are Associated with Age-Related Circulating Markers of Endothelial Dysfunction and Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossman, M.J.; Kaplon, R.E.; Hill, S.D.; McNamara, M.N.; Santos-Parker, J.R.; Pierce, G.L.; Seals, D.R.; Donato, A.J. Endothelial Cell Senescence with Aging in Healthy Humans: Prevention by Habitual Exercise and Relation to Vascular Endothelial Function. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H890–H895. [Google Scholar] [CrossRef]
- Angulo, J.; El Assar, M.; Álvarez-Bustos, A.; Rodríguez-Mañas, L. Physical Activity and Exercise: Strategies to Manage Frailty. Redox Biol. 2020, 35, 101513. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Donato, A.J.; Galvan, V.; Csiszar, A. Mechanisms of Vascular Aging. Circ. Res. 2018, 123, 849–867. [Google Scholar] [CrossRef]
- Walker, A.E.; Kaplon, R.E.; Pierce, G.L.; Nowlan, M.J.; Seals, D.R. Prevention of Age-Related Endothelial Dysfunction by Habitual Aerobic Exercise in Healthy Humans: Possible Role of Nuclear Factor ΚB. Clin. Sci. 2014, 127, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Mañas, L.; El-Assar, M.; Vallejo, S.; López-Dóriga, P.; Solís, J.; Petidier, R.; Montes, M.; Nevado, J.; Castro, M.; Gómez-Guerrero, C.; et al. Endothelial Dysfunction in Aged Humans Is Related with Oxidative Stress and Vascular Inflammation. Aging Cell 2009, 8, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Angulo, J.; El Assar, M.; Sevilleja-Ortiz, A.; Fernández, A.; Sánchez-Ferrer, A.; Romero-Otero, J.; Martínez-Salamanca, J.I.; La Fuente, J.M.; Rodríguez-Mañas, L. Short-Term Pharmacological Activation of Nrf2 Ameliorates Vascular Dysfunction in Aged Rats and in Pathological Human Vasculature. A Potential Target for Therapeutic Intervention. Redox Biol. 2019, 26, 101271. [Google Scholar] [CrossRef]
- El Assar, M.; Angulo, J.; Rodríguez-Mañas, L. Oxidative Stress and Vascular Inflammation in Aging. Free Radic. Biol. Med. 2013, 65, 380–401. [Google Scholar] [CrossRef]
- Dinh, Q.N.; Drummond, G.R.; Kemp-Harper, B.K.; Diep, H.; de Silva, T.M.; Kim, H.A.; Vinh, A.; Robertson, A.A.B.; Cooper, M.A.; Mansell, A.; et al. Pressor Response to Angiotensin II Is Enhanced in Aged Mice and Associated with Inflammation, Vasoconstriction and Oxidative Stress. Aging 2017, 9, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Assar, M.; Angulo, J.; García-Rojo, E.; Sevilleja-Ortiz, A.; García-Gómez, B.; Fernández, A.; Sánchez-Ferrer, A.; la Fuente, J.M.; Romero-Otero, J.; Rodríguez-Mañas, L. Early Manifestation of Aging-Related Vascular Dysfunction in Human Penile Vasculature—A Potential Explanation for the Role of Erectile Dysfunction as a Harbinger of Systemic Vascular Disease. Geroscience 2022, 44, 485–501. [Google Scholar] [CrossRef]
- El Assar, M.; Fernández, A.; Sánchez-Ferrer, A.; Angulo, J.; Rodríguez-Mañas, L. Multivessel Analysis of Progressive Vascular Aging in the Rat: Asynchronous Vulnerability among Vascular Territories. Mech. Ageing Dev. 2018, 173, 39–49. [Google Scholar] [CrossRef]
- Luttrell, M.; Kim, H.; Shin, S.Y.; Holly, D.; Massett, M.P.; Woodman, C.R. Heterogeneous Effect of Aging on Vasorelaxation Responses in Large and Small Arteries. Physiol. Rep. 2020, 8, e14341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harraz, O.F.; Jensen, L.J. Vascular Calcium Signalling and Ageing. J. Physiol. 2021, 599, 5361–5377. [Google Scholar] [CrossRef] [PubMed]
- Schach, C.; Wester, M.; Leibl, F.; Redel, A.; Gruber, M.; Maier, L.S.; Endemann, D.; Wagner, S. Reduced Store-Operated Ca2+ Entry Impairs Mesenteric Artery Function in Response to High External Glucose in Type 2 Diabetic ZDF Rats. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1145–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerman, E.M.; Everhart, J.E.; Segal, S.S. Advanced Age Decreases Local Calcium Signaling in Endothelium of Mouse Mesenteric Arteries in Vivo. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1091–H1096. [Google Scholar] [CrossRef] [Green Version]
- Boerman, E.M.; Segal, S.S. Aging Alters Spontaneous and Neurotransmitter-mediated Ca 2+ Signaling in Smooth Muscle Cells of Mouse Mesenteric Arteries. Microcirculation 2020, 27, e12607. [Google Scholar] [CrossRef]
- Pan, Z.; Brotto, M.; Ma, J. Store-Operated Ca2+ Entry in Muscle Physiology and Diseases. BMB Rep. 2014, 47, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Collins, H.E.; Zhang, D.; Chatham, J.C. STIM and Orai Mediated Regulation of Calcium Signaling in Age-Related Diseases. Front. Aging 2022, 3, 876785. [Google Scholar] [CrossRef]
- Tanwar, J.; Trebak, M.; Motiani, R.K. Cardiovascular and Hemostatic Disorders: Role of STIM and Orai Proteins in Vascular Disorders. Adv. Exp. Med. Biol. 2017, 993, 425–452. [Google Scholar] [CrossRef]
- Ruhle, B.; Trebak, M. Emerging Roles for Native Orai Ca2+ Channels in Cardiovascular Disease. Curr. Top. Membr. 2013, 71, 209–235. [Google Scholar]
- Giachini, F.R.C.; Chiao, C.-W.; Carneiro, F.S.; Lima, V.V.; Carneiro, Z.N.; Dorrance, A.M.; Tostes, R.C.; Webb, R.C. Increased Activation of Stromal Interaction Molecule-1/Orai-1 in Aorta from Hypertensive Rats. Hypertension 2009, 53, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Bhullar, S.K.; Shah, A.K.; Dhalla, N.S. Store-Operated Calcium Channels: Potential Target for the Therapy of Hypertension. Rev. Cardiovasc. Med. 2019, 20, 139. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, J.; Wang, X.; Xue, N.; Du, J.; Meng, X.; Shen, B. Contrasting Patterns of Agonist-Induced Store-Operated Ca2+ Entry and Vasoconstriction in Mesenteric Arteries and Aorta with Aging. J. Cardiovasc. Pharmacol. 2015, 65, 571–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevilleja-Ortiz, A.; El Assar, M.; García-Rojo, E.; García-Gómez, B.; Fernández, A.; Sánchez-Ferrer, A.; la Fuente, J.M.; Romero-Otero, J.; Rodríguez-Mañas, L.; Angulo, J. Ageing-Induced Hypercontractility Is Related to Functional Enhancement of STIM/Orai and Upregulation of Orai 3 in Rat and Human Penile Tissue. Mech. Ageing Dev. 2021, 200, 111590. [Google Scholar] [CrossRef] [PubMed]
- El Assar, M.; Angulo, J.; Santos-Ruiz, M.; Ruiz de Adana, J.C.; Pindado, M.L.; Sánchez-Ferrer, A.; Hernández, A.; Rodríguez-Mañas, L. Asymmetric Dimethylarginine (ADMA) Elevation and Arginase up-Regulation Contribute to Endothelial Dysfunction Related to Insulin Resistance in Rats and Morbidly Obese Humans. J. Physiol. 2016, 594, 3045–3060. [Google Scholar] [CrossRef]
- Angulo, J.; Vallejo, S.; El Assar, M.; García-Septiem, J.; Sánchez-Ferrer, C.F.; Rodríguez-Mañas, L. Age-Related Differences in the Effects of α and γ Peroxisome Proliferator-Activated Receptor Subtype Agonists on Endothelial Vasodilation in Human Microvessels. Exp. Gerontol. 2012, 47, 734–740. [Google Scholar] [CrossRef]
- Sevilleja-Ortiz, A.; El Assar, M.; García-Rojo, E.; Romero-Otero, J.; García-Gómez, B.; Fernández, A.; Medina-Polo, J.; la Fuente, J.M.; Rodríguez-Mañas, L.; Angulo, J. Enhanced Contribution of Orai Channels to Contractility of Human Penile Smooth Muscle in Erectile Dysfunction. J. Sex. Med. 2020, 17, 881–891. [Google Scholar] [CrossRef]
- Ungvari, Z.; Tarantini, S.; Sorond, F.; Merkely, B.; Csiszar, A. Mechanisms of Vascular Aging, A Geroscience Perspective: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 931–941. [Google Scholar] [CrossRef]
- Donato, A.J.; Machin, D.R.; Lesniewski, L.A. Mechanisms of Dysfunction in the Aging Vasculature and Role in Age-Related Disease. Circ. Res. 2018, 123, 825–848. [Google Scholar] [CrossRef]
- Casso, A.G.; VanDongen, N.S.; Gioscia-Ryan, R.A.; Clayton, Z.S.; Greenberg, N.T.; Ziemba, B.P.; Hutton, D.A.; Neilson, A.P.; Davy, K.P.; Seals, D.R.; et al. Initiation of 3,3-Dimethyl-1-Butanol at Midlife Prevents Endothelial Dysfunction and Attenuates in Vivo Aortic Stiffening with Ageing in Mice. J. Physiol. 2022, 600, 4633–4651. [Google Scholar] [CrossRef]
- Wei, X.; Lan, T.; Zhou, Y.; Cheng, J.; Li, P.; Zeng, X.; Yang, Y. Mechanism of A1-Adrenergic Receptor-Induced Increased Contraction of Rat Mesenteric Artery in Aging Hypertension Rats. Gerontology 2021, 67, 323–337. [Google Scholar] [CrossRef]
- Spinelli, A.M.; Trebak, M. Orai Channel-Mediated Ca2+ Signals in Vascular and Airway Smooth Muscle. Am. J. Physiol. Cell Physiol. 2016, 310, C402–C413. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Trebak, M. ORAI Channels in Cellular Remodeling of Cardiorespiratory Disease. Cell Calcium 2019, 79, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zöphel, D.; Hof, C.; Lis, A. Altered Ca2+ Homeostasis in Immune Cells during Aging: Role of Ion Channels. Int. J. Mol. Sci. 2020, 22, 110. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.; Moreno, A.; Briones, A.; Ivorra, M.D.; D’Ocon, P.; Vila, E. Alterations by Age of Calcium Handling in Rat Resistance Arteries. J. Cardiovasc. Pharmacol. 2002, 40, 832–840. [Google Scholar] [CrossRef]
- Guéguinou, M.; ChantÔme, A.; Fromont, G.; Bougnoux, P.; Vandier, C.; Potier-Cartereau, M. KCa and Ca2+ Channels: The Complex Thought. Biochim. Biophys. Acta 2014, 1843, 2322–2333. [Google Scholar] [CrossRef] [Green Version]
- Félétou, M. Endothelium-Dependent Hyperpolarization and Endothelial Dysfunction. J. Cardiovasc. Pharmacol. 2016, 67, 373–387. [Google Scholar] [CrossRef]
- Morikawa, K.; Fujiki, T.; Matoba, T.; Kubota, H.; Hatanaka, M.; Takahashi, S.; Shimokawa, H. Important Role of Superoxide Dismutase in EDHF-Mediated Responses of Human Mesenteric Arteries. J. Cardiovasc. Pharmacol. 2004, 44, 552–556. [Google Scholar] [CrossRef]
- Rivilla, F.; Vallejo, S.; Peiró, C.; Sánchez-Ferrer, C.F. Characterization of Endothelium-Dependent Relaxations in the Mesenteric Vasculature: A Comparative Study with Potential Pathophysiological Relevance. J. Pediatr. Surg. 2012, 47, 2044–2049. [Google Scholar] [CrossRef]
- Lompre, A.M.; Benard, L.; Saliba, Y.; Aubart, F.; Fauconnier, J.; Hulot, J.S. STIM1 and Orai in Cardiac Hypertrophy and Vascular Proliferative Diseases. Front. Biosci. 2013, 5, 766–773. [Google Scholar] [CrossRef] [Green Version]
- El Assar, M.; Angulo, J.; Vallejo, S.; Peiró, C.; Sánchez-Ferrer, C.F.; Rodríguez-Mañas, L. Mechanisms Involved in the Aging-Induced Vascular Dysfunction. Front. Physiol. 2012, 3, 132. [Google Scholar] [CrossRef] [Green Version]
- Murdaca, G.; Spanò, F.; Cagnati, P.; Puppo, F. Free Radicals and Endothelial Dysfunction: Potential Positive Effects of TNF-α Inhibitors. Redox Rep. 2013, 18, 95–99. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| <65-Years Old (n = 30) | >65-Years Old (n = 15) | p-Value | |
|---|---|---|---|
| Age (years) | 43.0 ± 10.8 | 73.7 ± 5.9 | <0.001 |
| Female (%) | 7 (23.3) | 7 (46.6) | 0.172 |
| Diabetes Mellitus (%) | 1 (3.3) | 4 (26.7) | 0.042 |
| Hypertension (%) | 3 (10.0) | 9 (60.0) | 0.001 |
| Dyslipidemia (%) | 3 (10.0) | 5 (33.3) | 0.099 |
| Cardiovascular Disease (%) | 3 (10.0) | 4 (26.7) | 0.212 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Assar, M.; García-Rojo, E.; Sevilleja-Ortiz, A.; Sánchez-Ferrer, A.; Fernández, A.; García-Gómez, B.; Romero-Otero, J.; Rodríguez-Mañas, L.; Angulo, J. Functional Role of STIM-1 and Orai1 in Human Microvascular Aging. Cells 2022, 11, 3675. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11223675
El Assar M, García-Rojo E, Sevilleja-Ortiz A, Sánchez-Ferrer A, Fernández A, García-Gómez B, Romero-Otero J, Rodríguez-Mañas L, Angulo J. Functional Role of STIM-1 and Orai1 in Human Microvascular Aging. Cells. 2022; 11(22):3675. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11223675
Chicago/Turabian StyleEl Assar, Mariam, Esther García-Rojo, Alejandro Sevilleja-Ortiz, Alberto Sánchez-Ferrer, Argentina Fernández, Borja García-Gómez, Javier Romero-Otero, Leocadio Rodríguez-Mañas, and Javier Angulo. 2022. "Functional Role of STIM-1 and Orai1 in Human Microvascular Aging" Cells 11, no. 22: 3675. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11223675