
Patterned Arteriole-Scale Vessels Enhance Engraftment, Perfusion, and Vessel Branching Hierarchy of Engineered Human Myocardium for Heart Regeneration
 , , , , and
, , , , and 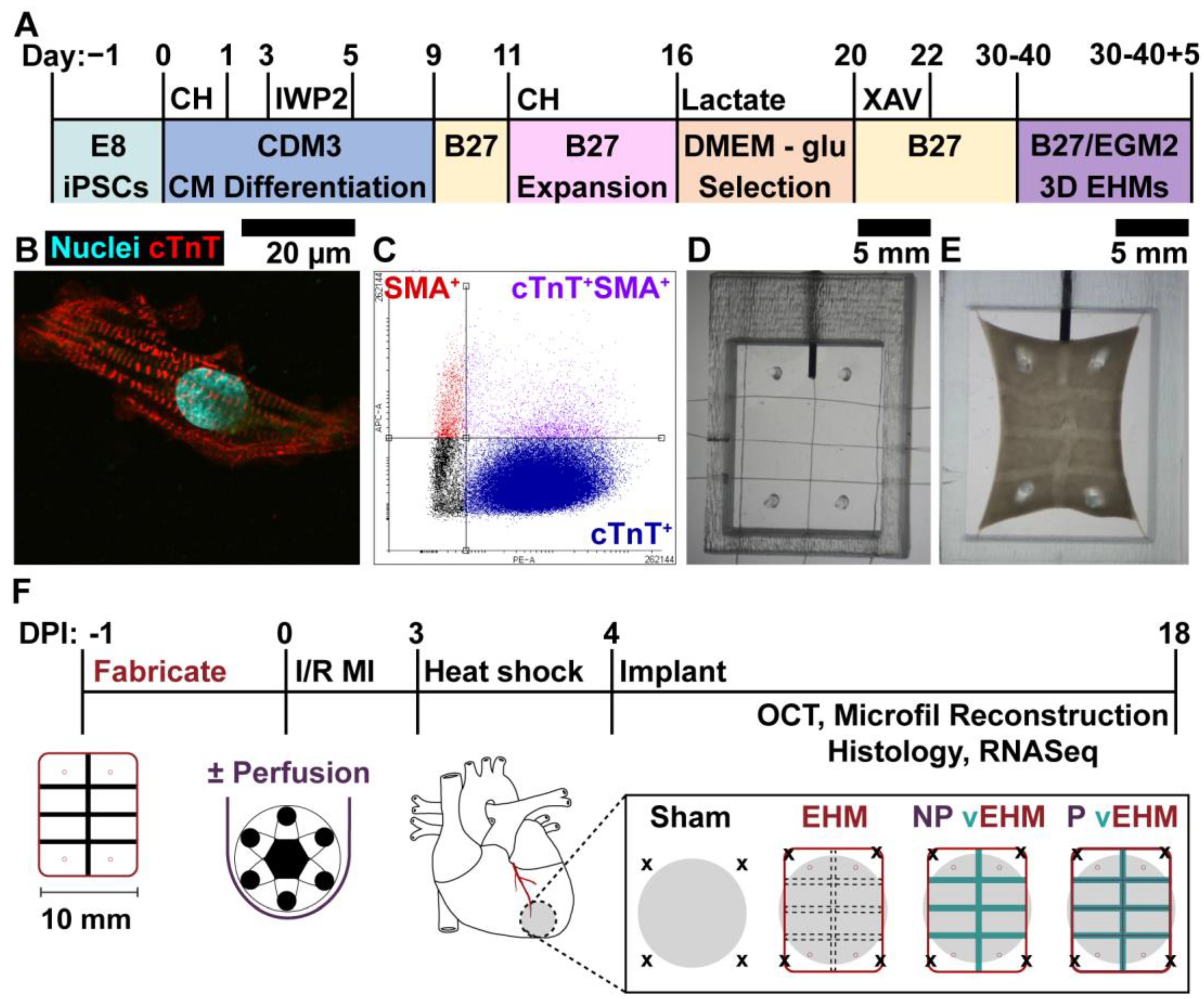{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. hiPSC Maintenance
2.2. hiPSC-CM Differentiation, Expansion, and Lactate Selection
2.3. Cardiac Fibroblast Mainenance
2.4. Endothelial Cell Mainenance
2.5. Cellular Viability, Proliferation, and Morphological Assays
2.6. Mechanical Characterization of EHMs
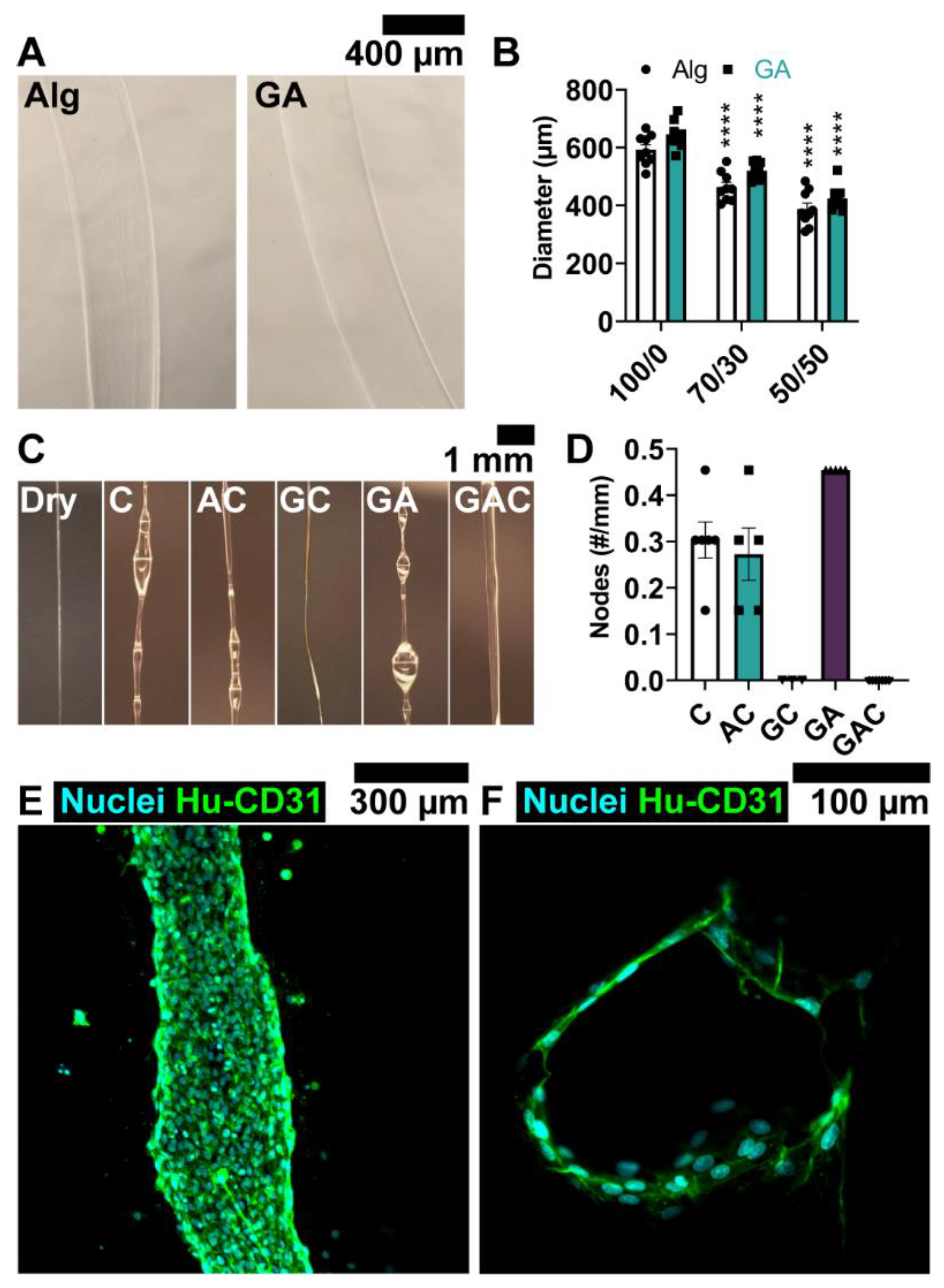
2.7. Gelatin-Alginate Fiber Wet Spinning
2.8. Collagen Microfiber Wet Spinning
2.9. EHM Fabrication for In Vivo Studies
2.10. GA Fiber Un-Crosslinking and Perfusion Culture
2.11. Implantation of EHMs on Infarcted Hearts and Explant
2.12. Optical Coherence Tomography Angiography
2.13. Vascular Reconstructive Analysis and Immunohistochemistry
2.14. Exploratory Bulk RNA Sequencing
2.15. Data and Statistical Analysis
3. Results
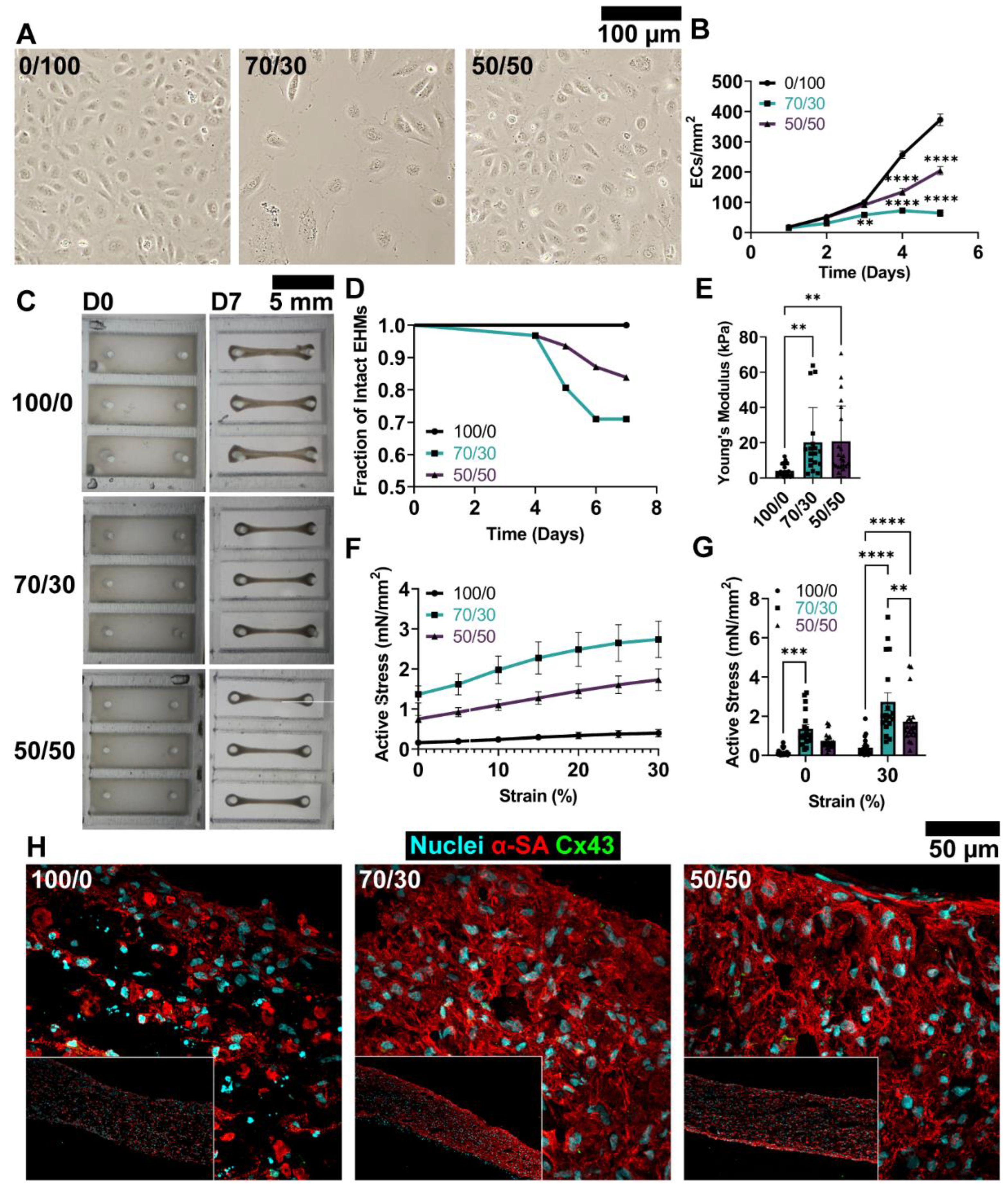
3.1. Medium Composition Influences hiPSC-CM and EC Structure and Function
3.2. Coating Solution Enables Local Delivery of ECs to Channel Wall
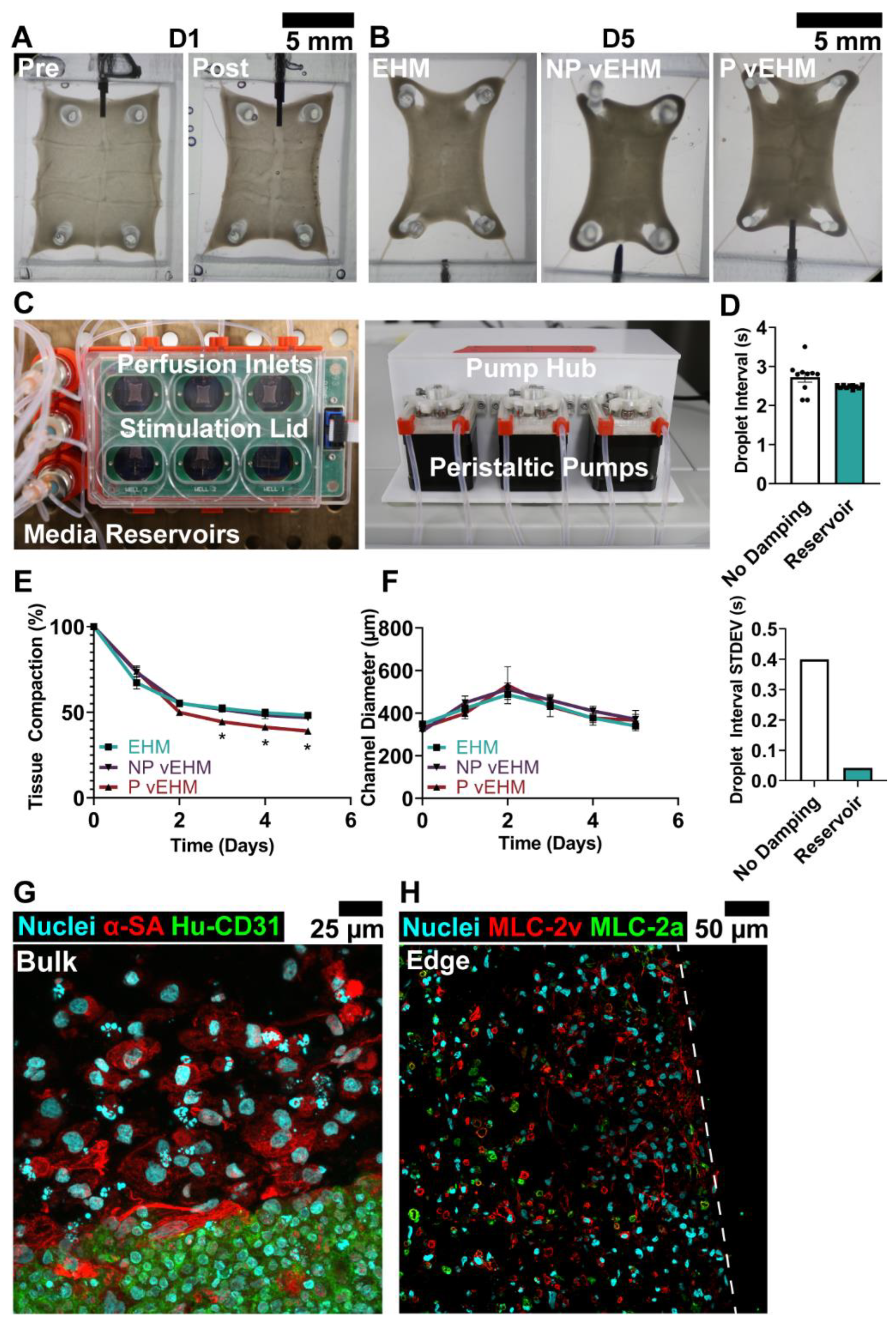
3.3. Dynamic Perfusion Culture of vEHMs
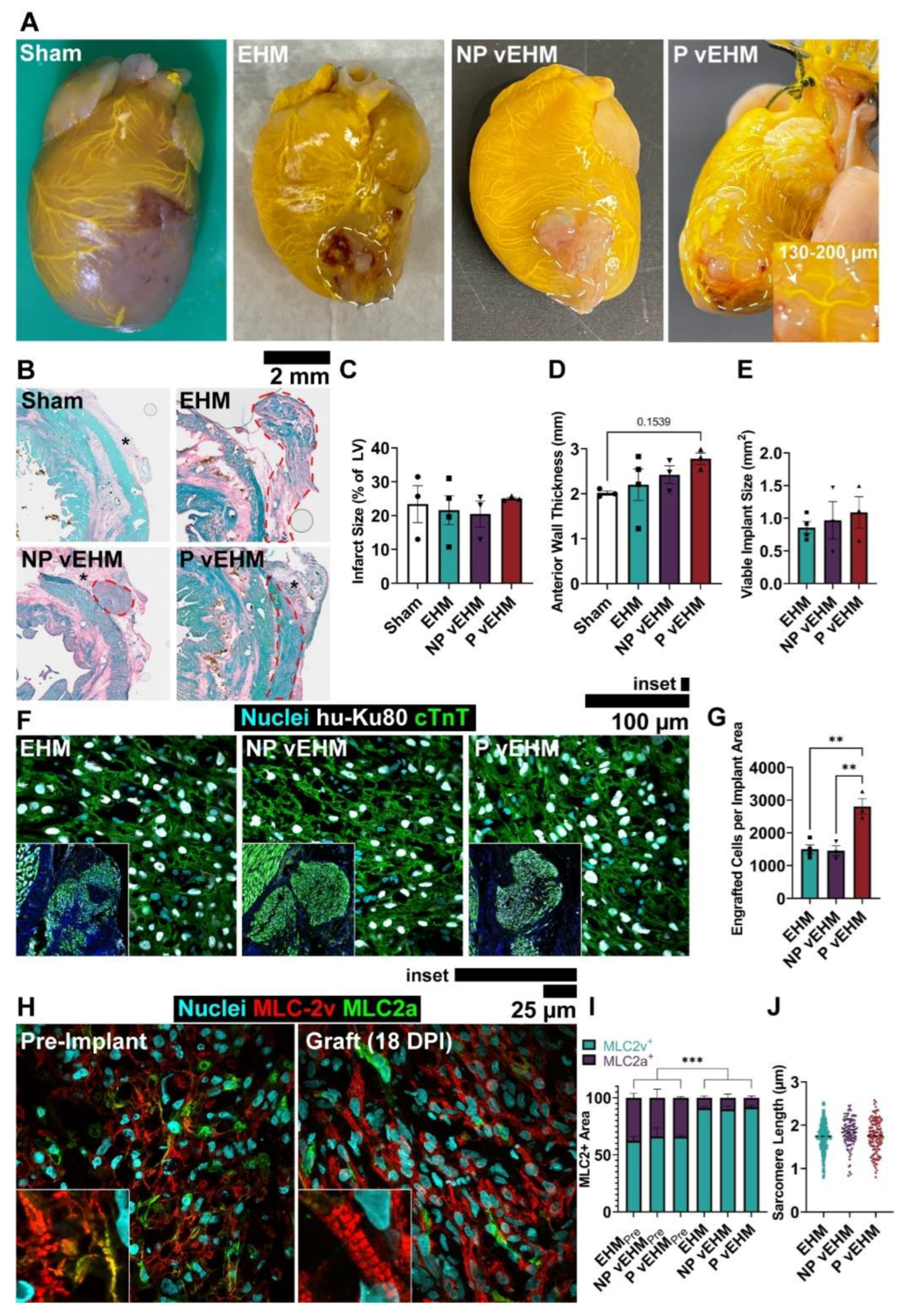
3.4. P vEHMs Exhibit Semi-Conserved Pattern Morphology and Improved Engraftment
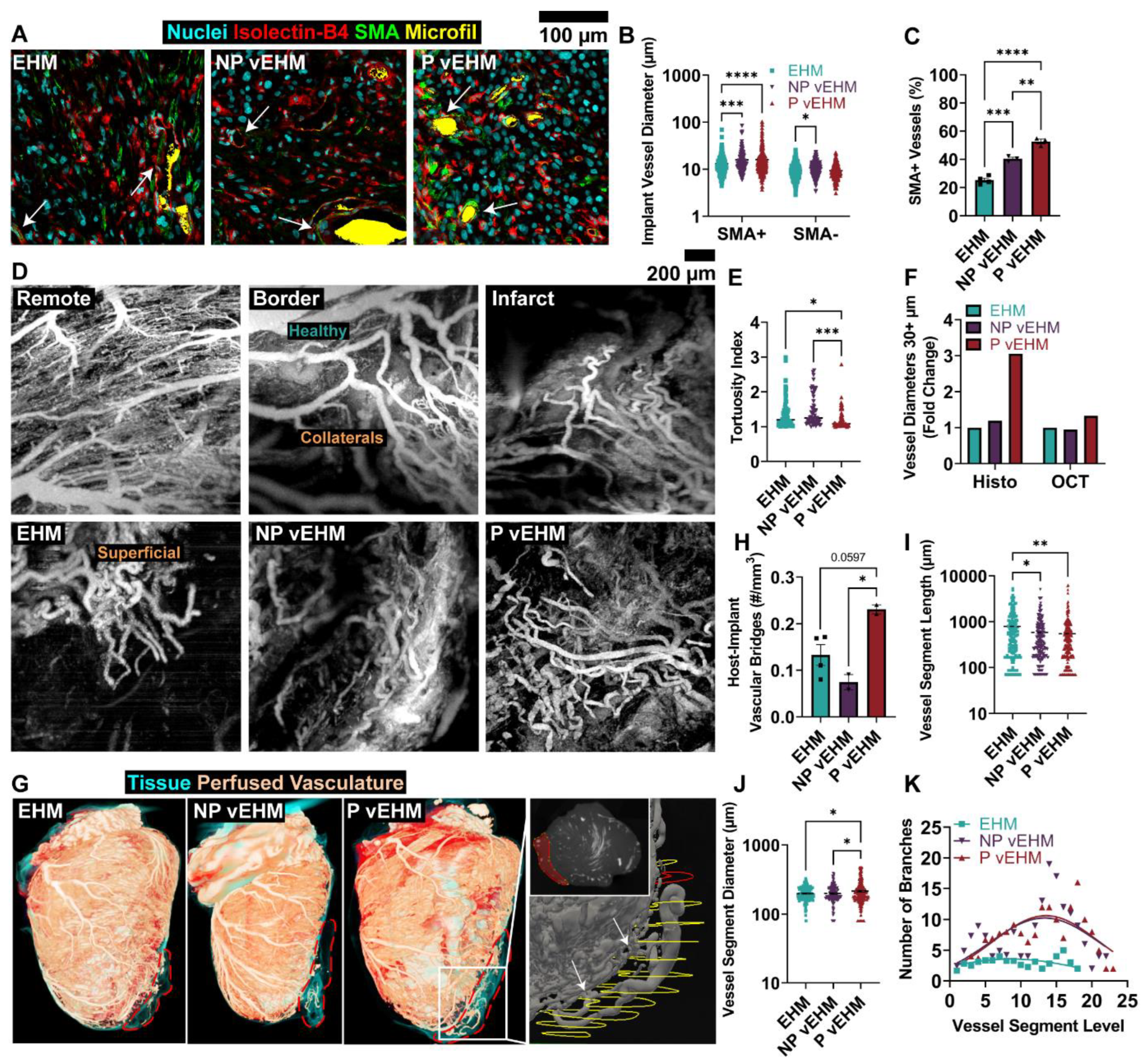
3.5. P vEHMs Develop Larger and More Hierarchically Organized Vasculature
3.6. Exploratory Bulk RNA Sequencing of Explanted vEHMs Suggests Perfusion Influences hiPSC-CM Development and Contractility
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laflamme, M.A.; Murry, C.E. Heart regeneration. Nature 2011, 473, 326–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munarin, F.; Kant, R.J.; Rupert, C.E.; Khoo, A.; Coulombe, K.L. Engineered human myocardium with local release of angiogenic proteins improves vascularization and cardiac function in injured rat hearts. Biomaterials 2020, 251, 120033. [Google Scholar] [CrossRef] [PubMed]
- Ogle, B.M.; Bursac, N.; Domian, I.; Huang, N.F.; Menasché, P.; Murry, C.E.; Pruitt, B.; Radisic, M.; Wu, J.C.; Wu, S.M.; et al. Distilling complexity to advance cardiac tissue engineering. Sci. Transl. Med. 2016, 8, 342ps13. [Google Scholar] [CrossRef] [Green Version]
- Kant, R.J.; Coulombe, K.L. Integrated approaches to spatiotemporally directing angiogenesis in host and engineered tissues. Acta Biomater. 2018, 69, 42–62. [Google Scholar] [CrossRef]
- Dvir, T.; Kedem, A.; Ruvinov, E.; Levy, O.; Freeman, I.; Landa, N.; Holbova, R.; Feinberg, M.S.; Dror, S.; Etzion, Y.; et al. Prevascularization of cardiac patch on the omentum improves its therapeutic outcome. Proc. Natl. Acad. Sci. USA 2009, 106, 14990–14995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koffler, J.; Kaufman-Francis, K.; Shandalov, Y.; Egozi, D.; Amiad Pavlov, D.; Landesberg, A.; Levenberg, S. Improved vascular organization enhances functional integration of engineered skeletal muscle grafts. Proc. Natl. Acad. Sci. USA 2012, 109, 14789–14794. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, J.A.; Guzman, P.A.; Riemenschneider, S.B.; Kamp, T.J.; Tranquillo, R.T. A cardiac patch from aligned microvessel and cardiomyocyte patches. J. Tissue Eng. Regen. Med. 2018, 12, 546–556. [Google Scholar] [CrossRef]
- Sun, X.; Wu, J.; Qiang, B.; Romagnuolo, R.; Gagliardi, M.; Keller, G.; Laflamme, M.A.; Li, R.-K.; Nunes, S.S. Transplanted microvessels improve pluripotent stem cell–derived cardiomyocyte engraftment and cardiac function after infarction in rats. Sci. Transl. Med. 2020, 12, 2992. [Google Scholar] [CrossRef]
- Mirabella, T.; MacArthur, J.W.; Cheng, D.; Ozaki, C.K.; Woo, Y.J.; Yang, M.T.; Chen, C.S. 3D-printed vascular networks direct therapeutic angiogenesis in ischaemia. Nat. Biomed. Eng. 2017, 1, 0083. [Google Scholar] [CrossRef] [Green Version]
- Mirdamadi, E.; Tashman, J.W.; Shiwarski, D.J.; Palchesko, R.N.; Feinberg, A.W. FRESH 3D Bioprinting a Full-Size Model of the Human Heart. ACS Biomater. Sci. Eng. 2020, 6, 6453–6459. [Google Scholar] [CrossRef]
- Kinstlinger, I.S.; Calderon, G.A.; Royse, M.K.; Means, A.K.; Grigoryan, B.; Miller, J.S. Perfusion and endothelialization of engineered tissues with patterned vascular networks. Nat. Protoc. 2021, 16, 3089–3113. [Google Scholar] [CrossRef] [PubMed]
- Brady, E.L.; Kirby, M.A.; Olszewski, E.; Grosjean, P.; Johansson, F.; Davis, J.; Wang, R.K.; Stevens, K.R. Guided vascularization in the rat heart leads to transient vessel patterning. APL Bioeng. 2020, 4, 016105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redd, M.A.; Zeinstra, N.; Qin, W.; Wei, W.; Martinson, A.; Wang, Y.; Wang, R.K.; Murry, C.E.; Zheng, Y. Patterned human microvascular grafts enable rapid vascularization and increase perfusion in infarcted rat hearts. Nat. Commun. 2019, 10, 584. [Google Scholar] [CrossRef] [Green Version]
- Vollert, I.; Seiffert, M.; Bachmair, J.; Sander, M.; Eder, A.; Conradi, L.; Vogelsang, A.; Schulze, T.; Uebeler, J.; Holnthoner, W.; et al. In-vitro perfusion of engineered heart tissue through endothelialized channels. Tissue Eng. Part A 2014, 20, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Vollmar, B.; Menger, M.D. Inosculation: Connecting the Life-Sustaining Pipelines. Tissue Eng. Part B Rev. 2009, 15, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Weyers, J.J.; Schwartz, S.M.; Minami, E.; Carlson, D.D.; Dupras, S.K.; Weitz, K.; Simons, M.; Cox, T.C.; Murry, C.E.; Mahoney, W.M. Effects of Cell Grafting on Coronary Remodeling After Myocardial Infarction. J. Am. Heart Assoc. 2013, 2, e000202. [Google Scholar] [CrossRef] [Green Version]
- Weyers, J.J.; Gunaje, J.J.; Van Biber, B.; Martinson, A.; Reinecke, H.; Mahoney, W.M.; Schwartz, S.M.; Cox, T.C.; Murry, C.E. Sonic Hedgehog upregulation does not enhance the survival and engraftment of stem cell-derived cardiomyocytes in infarcted hearts. PLoS ONE 2020, 15, e0227780. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Roberts, M.A.; Qi, X.; Murry, C.E.; Zheng, Y.; Wang, R.K. Depth-resolved 3D visualization of coronary microvasculature with optical microangiography. Phys. Med. Biol. 2016, 61, 7536–7550. [Google Scholar] [CrossRef] [Green Version]
- Vinegoni, C.; Aguirre, A.D.; Lee, S.; Weissleder, R. Imaging the beating heart in the mouse using intravital microscopy techniques. Nat. Protoc. 2015, 10, 1802–1819. [Google Scholar] [CrossRef] [Green Version]
- Merz, S.F.; Korste, S.; Bornemann, L.; Michel, L.; Stock, P.; Squire, A.; Soun, C.; Engel, D.R.; Detzer, J.; Lörchner, H.; et al. Contemporaneous 3D characterization of acute and chronic myocardial I/R injury and response. Nat. Commun. 2019, 10, 2312. [Google Scholar] [CrossRef] [Green Version]
- Anbazhakan, S.; Coronado, P.E.R.; Sy-Quia, A.N.L.; Seow, L.W.; Hands, A.M.; Zhao, M.; Dong, M.L.; Pfaller, M.R.; Amir, Z.A.; Raftrey, B.C.; et al. Blood flow modeling reveals improved collateral artery performance during the regenerative period in mammalian hearts. Nat. Cardiovasc. Res. 2022, 1, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Garcia, M.J.; Quesnel, E.; Al-Attar, R.; Laskary, A.R.; Laflamme, M.A. Maturation of human pluripotent stem cell derived cardiomyocytes in vitro and in vivo. Semin. Cell Dev. Biol. 2021, 118, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pu, W.T. Cardiomyocyte Maturation. Circ. Res. 2020, 126, 1086–1106. [Google Scholar] [CrossRef] [PubMed]
- Dhahri, W.; Valdman, T.S.; Wilkinson, D.; Pereira, E.; Ceylan, E.; Andharia, N.; Qiang, B.; Masoudpour, H.; Wulkan, F.; Quesnel, E.; et al. In Vitro Matured Human Pluripotent Stem Cell–Derived Cardiomyocytes Form Grafts with Enhanced Structure and Function in Injured Hearts. Circulation 2022, 145, 1412–1426. [Google Scholar] [CrossRef]
- Abilez, O.J.; Tzatzalos, E.; Yang, H.; Zhao, M.-T.; Jung, G.; Zöllner, A.M.; Tiburcy, M.; Riegler, J.; Matsa, E.; Shukla, P.; et al. Passive Stretch Induces Structural and Functional Maturation of Engineered Heart Muscle as Predicted by Computational Modeling. Stem Cells 2018, 36, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querdel, E.; Reinsch, M.; Castro, L.; Köse, D.; Bähr, A.; Reich, S.; Geertz, B.; Ulmer, B.; Schulze, M.; Lemoine, M.D.; et al. Human Engineered Heart Tissue Patches Remuscularize the Injured Heart in a Dose-Dependent Manner. Circulation 2021, 143, 1991–2006. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Nunes, S.S. Maturation of human stem cell-derived cardiomyocytes in biowires using electrical stimulation. J. Vis. Exp. 2017, 2017, e55373. [Google Scholar]
- Tohyama, S.; Hattori, F.; Sano, M.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Hashimoto, H.; Suzuki, T.; Yamashita, H.; Satoh, Y.; et al. Distinct Metabolic Flow Enables Large-Scale Purification of Mouse and Human Pluripotent Stem Cell-Derived Cardiomyocytes. Cell Stem Cell 2013, 12, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Feyen, D.A.M.; McKeithan, W.L.; Bruyneel, A.A.N.; Spiering, S.; Hörmann, L.; Ulmer, B.; Zhang, H.; Briganti, F.; Schweizer, M.; Hegyi, B.; et al. Metabolic Maturation Media Improve Physiological Function of Human iPSC-Derived Cardiomyocytes. Cell Rep. 2020, 32, 107925. [Google Scholar] [CrossRef]
- Miki, K.; Deguchi, K.; Nakanishi-Koakutsu, M.; Lucena-Cacace, A.; Kondo, S.; Fujiwara, Y.; Hatani, T.; Sasaki, M.; Naka, Y.; Okubo, C.; et al. ERRγ enhances cardiac maturation with T-tubule formation in human iPSC-derived cardiomyocytes. Nat. Commun. 2021, 12, 3596. [Google Scholar] [CrossRef]
- Dwyer, K.D.; Kant, R.J.; Soepriatna, A.H.; Roser, S.M.; Daley, M.C.; Sabe, S.A.; Xu, C.M.; Choi, B.-R.; Sellke, F.W.; Coulombe, K.L.K. One Billion hiPSC-Cardiomyocytes: Upscaling Engineered Cardiac Tissues to Create High Cell Density Therapies for Clinical Translation in Heart Regeneration. Bioengineering 2023, 10, 587. [Google Scholar] [CrossRef] [PubMed]
- Von Bibra, C.; Shibamiya, A.; Bähr, A.; Geertz, B.; Köhne, M.; Stuedemann, T.; Starbatty, J.; der Sluis, V.H.-V.; Klostermeier, U.C.; Hornaschewitz, N.; et al. Immature human engineered heart tissues engraft in a guinea pig chronic injury model. Dis. Model. Mech. 2023, 16, dmm049834. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.C.; Maas, R.G.C.; Van Adrichem, I.; Doevendans, P.A.M.; Mercola, M.; Šarić, T.; Buikema, J.W.; Van Mil, A.; Chamuleau, S.A.J.; Sluijter, J.P.G.; et al. Metabolic Maturation Increases Susceptibility to Hypoxia-induced Damage in Human iPSC-derived Cardiomyocytes. Stem Cells Transl. Med. 2022, 11, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, S.; Fernandes, I.; Mastikhina, O.; Wilkinson, D.; Tran, T.; Dhahri, W.; Mazine, A.; Yang, D.; Burnett, B.; Lee, J.; et al. Generation of mature compact ventricular cardiomyocytes from human pluripotent stem cells. Nat. Commun. 2021, 12, 3115. [Google Scholar] [CrossRef]
- Biagi, D.; Fantozzi, E.T.; Campos-Oliveira, J.C.; Naghetini, M.V.; Ribeiro, A.F.; Rodrigues, S.; Ogusuku, I.; Vanderlinde, R.; Christie, M.L.A.; Mello, D.B.; et al. In Situ Maturated Early-Stage Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes Improve Cardiac Function by Enhancing Segmental Contraction in Infarcted Rats. J. Pers. Med. 2021, 11, 374. [Google Scholar] [CrossRef]
- Riegler, J.; Tiburcy, M.; Ebert, A.D.; Tzatzalos, E.; Raaz, U.; Abilez, O.J.; Shen, Q.; Kooreman, N.G.; Neofytou, E.; Chen, V.C.; et al. Human Engineered Heart Muscles Engraft and Survive Long Term in a Rodent Myocardial Infarction Model. Circ. Res. 2015, 117, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Pedde, R.D.; Mirani, B.; Navaei, A.; Styan, T.; Wong, S.; Mehrali, M.; Thakur, A.; Mohtaram, N.K.; Bayati, A.; Dolatshahi-Pirouz, A.; et al. Emerging Biofabrication Strategies for Engineering Complex Tissue Constructs. Adv. Mater. 2017, 29, 1606061. [Google Scholar] [CrossRef] [PubMed]
- Eschenhagen, T.; Ridders, K.; Weinberger, F. How to repair a broken heart with pluripotent stem cell-derived cardiomyocytes. J. Mol. Cell. Cardiol. 2022, 163, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Shadrin, I.Y.; Allen, B.W.; Qian, Y.; Jackman, C.P.; Carlson, A.L.; Juhas, M.E.; Bursac, N. Cardiopatch platform enables maturation and scale-up of human pluripotent stem cell-derived engineered heart tissues. Nat. Commun. 2017, 8, 1825. [Google Scholar] [CrossRef] [Green Version]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat. Protoc. 2012, 8, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupert, C.E.; Irofuala, C.; Coulombe KL, K. Practical adoption of state-of-the-art hiPSC-cardiomyocyte differentiation techniques. PLoS ONE 2020, 15, e0230001. [Google Scholar] [CrossRef] [PubMed]
- Buikema, J.W.; Lee, S.; Goodyer, W.R.; Maas, R.G.; Chirikian, O.; Li, G.; Miao, Y.; Paige, S.L.; Lee, D.; Wu, H.; et al. Wnt Activation and Reduced Cell-Cell Contact Synergistically Induce Massive Expansion of Functional Human iPSC-Derived Cardiomyocytes. Cell Stem Cell 2020, 27, 50–63.e5. [Google Scholar] [CrossRef]
- Munarin, F.; Kaiser, N.J.; Kim, T.Y.; Choi, B.-R.; Coulombe, K.L. Laser-Etched Designs for Molding Hydrogel-Based Engineered Tissues. Tissue Eng. Part C Methods 2017, 23, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, N.J.; Munarin, F.; Coulombe KL, K. Custom Engineered Tissue Culture Molds from Laser-etched Masters. J. Vis. Exp. 2018, 2018, e57239. [Google Scholar]
- Rupert, C.E.; Kim, T.Y.; Choi, B.R.; Coulombe KL, K. Human Cardiac Fibroblast Number and Activation State Modulate Electromechanical Function of hiPSC-Cardiomyocytes in Engineered Myocardium. Stem Cells Int. 2020, 2020, 9363809. [Google Scholar] [CrossRef]
- Kant, R.J.; Bare, M.C.F.; Coulombe, K.L. Tissues with Patterned Vessels or Protein Release Induce Vascular Chemotaxis in an In Vitro Platform. Tissue Eng. Part A 2021, 27, 1290–1304. [Google Scholar] [CrossRef]
- Kaiser, N.J.; Bellows, J.A.; Kant, R.J.; Coulombe, K.L. Digital Design and Automated Fabrication of Bespoke Collagen Microfiber Scaffolds. Tissue Eng. Part C Methods 2019, 25, 687–700. [Google Scholar] [CrossRef] [Green Version]
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef]
- Cezar, C.A.; Arany, P.; Vermillion, S.A.; Seo, B.R.; Vandenburgh, H.H.; Mooney, D.J. Timed Delivery of Therapy Enhances Functional Muscle Regeneration. Adv. Healthc. Mater. 2017, 6, 1700202. [Google Scholar] [CrossRef]
- Krzemiński, T.F.; Nożyński, J.K.; Grzyb, J.; Porc, M. Wide-spread myocardial remodeling after acute myocardial infarction in rat. Features for heart failure progression. Vasc. Pharmacol. 2008, 48, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Updegrove, A.; Wilson, N.M.; Merkow, J.; Lan, H.; Marsden, A.L.; Shadden, S.C. SimVascular: An Open Source Pipeline for Cardiovascular Simulation. Ann. Biomed. Eng. 2017, 45, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhuang, Z.W.; Sinusas, A.J.; Staib, L.H.; Papademetris, X. Vessel Connectivity Using Murray’s Hypothesis. J. Investig. Dermatol. 2011, 135, 528–536. [Google Scholar]
- Afgan, E.; Baker, D.; Batut, B.; Van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballermann, B.J.; Dardik, A.; Eng, E.; Liu, A. Shear stress and the endothelium. Kidney Int. 1998, 54, S100–S108. [Google Scholar] [CrossRef] [Green Version]
- Mughal, M.M.; Khan, M.K.; DeMarco, J.K.; Majid, A.; Shamoun, F.; Abela, G.S. Shear stress and plaque development. Expert Rev. Cardiovasc. Ther. 2010, 8, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Debbi, L.; Zohar, B.; Shuhmaher, M.; Shandalov, Y.; Goldfracht, I.; Levenberg, S. Integrating engineered macro vessels with self-assembled capillaries in 3D implantable tissue for promoting vascular integration in-vivo. Biomaterials 2022, 280, 121286. [Google Scholar] [CrossRef]
- Fang, Y.; Ouyang, L.; Zhang, T.; Wang, C.; Lu, B.; Sun, W. Optimizing Bifurcated Channels within an Anisotropic Scaffold for Engineering Vascularized Oriented Tissues. Adv. Healthc. Mater. 2020, 9, 2000782. [Google Scholar] [CrossRef]
- Kurokawa, Y.K.; Yin, R.; Shang, M.R.; Shirure, V.; Moya, M.L.; George, S.C. Human Induced Pluripotent Stem Cell-Derived Endothelial Cells for Three-Dimensional Microphysiological Systems. Tissue Eng. Part C Methods 2017, 23, 474–484. [Google Scholar] [CrossRef]
- Minor, A.J.; Coulombe, K.L.K. Stimulating Calcium Handling in hiPSC-Derived Engineered Cardiac Tissues Enhances Force Production. Stem Cells Transl. Med. 2022, 11, 97–106. [Google Scholar] [CrossRef]
- Shi, L.; Ko, S.; Ko, M.L.; Kim, A.J.; Ko, G.Y.-P. Peptide Lv augments L-type voltage-gated calcium channels through vascular endothelial growth factor receptor 2 (VEGFR2) signaling. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 1154–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottbauer, W.; Just, S.; Wessels, G.; Trano, N.; Most, P.; Katus, H.A.; Fishman, M.C. VEGF-PLCγ1 pathway controls cardiac contractility in the embryonic heart. Genes Dev. 2005, 19, 1624–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupert, C.E.; Coulombe, K.L.K. IGF1 and NRG1 Enhance Proliferation, Metabolic Maturity, and the Force-Frequency Response in hESC-Derived Engineered Cardiac Tissues. Stem Cells Int. 2017, 2017, 7648409. [Google Scholar] [CrossRef] [Green Version]
- Gerbin, K.A.; Yang, X.; Murry, C.E.; Coulombe, K.L.K. Enhanced Electrical Integration of Engineered Human Myocardium via Intramyocardial versus Epicardial Delivery in Infarcted Rat Hearts. PLoS ONE 2015, 10, e0131446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberger, F.; Breckwoldt, K.; Pecha, S.; Kelly, A.; Geertz, B.; Starbatty, J.; Yorgan, T.; Cheng, K.-H.; Lessmann, K.; Stolen, T.; et al. Cardiac repair in guinea pigs with human engineered heart tissue from induced pluripotent stem cells. Sci. Transl. Med. 2016, 8, 363148. [Google Scholar] [CrossRef]
- Brady, E.L.; Prado, O.; Johansson, F.; Mitchell, S.N.; Martinson, A.M.; Karbassi, E.; Reinecke, H.; Murry, C.E.; Davis, J.; Stevens, K.R. Engineered tissue vascularization and engraftment depends on host model. Sci. Rep. 2023, 13, 1973. [Google Scholar] [CrossRef]
- Helle, E.; Ampuja, M.; Antola, L.; Kivelä, R. Flow-Induced Transcriptomic Remodeling of Endothelial Cells Derived From Human Induced Pluripotent Stem Cells. Front. Physiol. 2020, 11, 591450. [Google Scholar] [CrossRef]
- White, S.M.; Pittman, C.R.; Hingorani, R.; Arora, R.; Esipova, T.V.; Vinogradov, S.A.; Hughes, C.C.; Choi, B.; George, S.C. Implanted Cell-Dense Prevascularized Tissues Develop Functional Vasculature That Supports Reoxygenation After Thrombosis. Tissue Eng. Part A 2014, 20, 2316–2328. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shaul, S.; Landau, S.; Merdler, U.; Levenberg, S. Mature vessel networks in engineered tissue promote graft–host anastomosis and prevent graft thrombosis. Proc. Natl. Acad. Sci. USA 2019, 116, 2955–2960. [Google Scholar] [CrossRef] [Green Version]
- Szklanny, A.A.; Machour, M.; Redenski, I.; Chochola, V.; Goldfracht, I.; Kaplan, B.; Epshtein, M.; Yameen, H.S.; Merdler, U.; Feinberg, A.; et al. 3D Bioprinting of Engineered Tissue Flaps with Hierarchical Vessel Networks (VesselNet) for Direct Host-To-Implant Perfusion. Adv. Mater. 2021, 33, 2661. [Google Scholar] [CrossRef] [PubMed]
- Baranski, J.D.; Chaturvedi, R.R.; Stevens, K.R.; Eyckmans, J.; Carvalho, B.; Solorzano, R.D.; Yang, M.T.; Miller, J.S.; Bhatia, S.N.; Chen, C.S. Geometric control of vascular networks to enhance engineered tissue integration and function. Proc. Natl. Acad. Sci. USA 2013, 110, 7586–7591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Manz, X.D.; Bongers, A.; Odell, R.A.; Joukhdar, H.; Whitelock, J.M.; Lord, M.S.; Rnjak-Kovacina, J. Microchannels Are an Architectural Cue That Promotes Integration and Vascularization of Silk Biomaterials in Vivo. ACS Biomater. Sci. Eng. 2020, 6, 1476–1486. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, K.; Du, L.; Cheng, Z.; Zhang, T.; Ding, J.; Li, W.; Xu, B.; Zhu, M. Construction of chitosan scaffolds with controllable microchannel for tissue engineering and regenerative medicine. Mater. Sci. Eng. C 2021, 126, 112178. [Google Scholar] [CrossRef] [PubMed]
- Parkhideh, S.; Calderon, G.A.; Janson, K.D.; Mukherjee, S.; Mai, A.K.; Doerfert, M.D.; Yao, Z.; Sazer, D.W.; Veiseh, O. Perfusable cell-laden matrices to guide patterning of vascularization in vivo. Biomater. Sci. 2022, 11, 461–471. [Google Scholar] [CrossRef]
- Lee, J.; Wu, W.; Jiang, J.Y.; Zhu, B.; Boas, D.A. Dynamic light scattering optical coherence tomography. Opt. Express 2012, 20, 22262–22277. [Google Scholar] [CrossRef]
- Redenski, I.; Guo, S.; Machour, M.; Szklanny, A.; Landau, S.; Egozi, D.; Gabet, Y.; Levenberg, S. Microcomputed Tomography-Based Analysis of Neovascularization within Bioengineered Vascularized Tissues. ACS Biomater. Sci. Eng. 2022, 8, 232–241. [Google Scholar] [CrossRef]
- Cruz-Moreira, D.; Visone, R.; Vasques-Nóvoa, F.; Barros, A.S.; Leite-Moreira, A.; Redaelli, A.; Moretti, M.; Rasponi, M. Assessing the influence of perfusion on cardiac microtissue maturation: A heart-on-chip platform embedding peristaltic pump capabilities. Biotechnol. Bioeng. 2021, 118, 3128–3137. [Google Scholar] [CrossRef]
- Karra, R.; Foglia, M.J.; Choi, W.-Y.; Belliveau, C.; DeBenedittis, P.; Poss, K.D. Vegfaa instructs cardiac muscle hyperplasia in adult zebrafish. Proc. Natl. Acad. Sci. USA 2018, 115, 8805–8810. [Google Scholar] [CrossRef] [Green Version]
- DeBenedittis, P.; Karpurapu, A.; Henry, A.; Thomas, M.C.; McCord, T.J.; Brezitski, K.; Prasad, A.; Baker, C.E.; Kobayashi, Y.; Shah, S.H.; et al. Coupled myovascular expansion directs cardiac growth and regeneration. Development 2022, 149, dev200654. [Google Scholar] [CrossRef]
- Tiburcy, M.; Hudson, J.E.; Balfanz, P.; Schlick, S.; Meyer, T.; Liao, M.-L.C.; Levent, E.; Raad, F.; Zeidler, S.; Wingender, E.; et al. Defined Engineered Human Myocardium with Advanced Maturation for Applications in Heart Failure Modeling and Repair. Circulation 2017, 135, 1832–1847. [Google Scholar] [CrossRef] [PubMed]
- Quaife-Ryan, G.A.; Sim, C.B.; Ziemann, M.; Kaspi, A.; Rafehi, H.; Ramialison, M.; El-Osta, A.; Hudson, J.E.; Porrello, E.R. Multicellular Transcriptional Analysis of Mammalian Heart Regeneration. Circulation 2017, 136, 1123–1139. [Google Scholar] [CrossRef] [PubMed]
- McLellan, M.A.; Skelly, D.A.; Dona, M.S.; Squiers, G.T.; Farrugia, G.E.; Gaynor, T.L.; Cohen, C.D.; Pandey, R.; Diep, H.; Vinh, A.; et al. High-Resolution Transcriptomic Profiling of the Heart During Chronic Stress Reveals Cellular Drivers of Cardiac Fibrosis and Hypertrophy. Circulation 2020, 142, 1448–1463. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yao, F.; Wang, L.; Li, Z.; Ren, Z.; Li, D.; Zhang, M.; Han, L.; Wang, S.-Q.; Zhou, B.; et al. Single-cell analysis of murine fibroblasts identifies neonatal to adult switching that regulates cardiomyocyte maturation. Nat. Commun. 2020, 11, 2585. [Google Scholar] [CrossRef]
- Marchiano, S.; Nakamura, K.; Reinecke, H.; Neidig, L.; Lai, M.; Kadota, S.; Perbellini, F.; Yang, X.; Klaiman, J.M.; Blakely, L.P.; et al. Gene editing to prevent ventricular arrhythmias associated with cardiomyocyte cell therapy. Cell Stem Cell 2023, 30, 741. [Google Scholar] [CrossRef] [PubMed]
- Kluin, R.J.C.; Kemper, K.; Kuilman, T.; De Ruiter, J.R.; Iyer, V.; Forment, J.V.; Cornelissen-Steijger, P.; De Rink, I.; Ter Brugge, P.; Song, J.-Y.; et al. XenofilteR: Computational deconvolution of mouse and human reads in tumor xenograft sequence data. BMC Bioinform. 2018, 19, 366. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kant, R.J.; Dwyer, K.D.; Lee, J.-H.; Polucha, C.; Kobayashi, M.; Pyon, S.; Soepriatna, A.H.; Lee, J.; Coulombe, K.L.K. Patterned Arteriole-Scale Vessels Enhance Engraftment, Perfusion, and Vessel Branching Hierarchy of Engineered Human Myocardium for Heart Regeneration. Cells 2023, 12, 1698. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12131698
Kant RJ, Dwyer KD, Lee J-H, Polucha C, Kobayashi M, Pyon S, Soepriatna AH, Lee J, Coulombe KLK. Patterned Arteriole-Scale Vessels Enhance Engraftment, Perfusion, and Vessel Branching Hierarchy of Engineered Human Myocardium for Heart Regeneration. Cells. 2023; 12(13):1698. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12131698
Chicago/Turabian StyleKant, Rajeev J., Kiera D. Dwyer, Jang-Hoon Lee, Collin Polucha, Momoka Kobayashi, Stephen Pyon, Arvin H. Soepriatna, Jonghwan Lee, and Kareen L. K. Coulombe. 2023. "Patterned Arteriole-Scale Vessels Enhance Engraftment, Perfusion, and Vessel Branching Hierarchy of Engineered Human Myocardium for Heart Regeneration" Cells 12, no. 13: 1698. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12131698