Perspective: Mitochondria-ER Contacts in Metabolic Cellular Stress Assessed by Microscopy

 ,
,  , , and
, , and
Abstract
:
1. “Contactology” a New Branch of Cytology
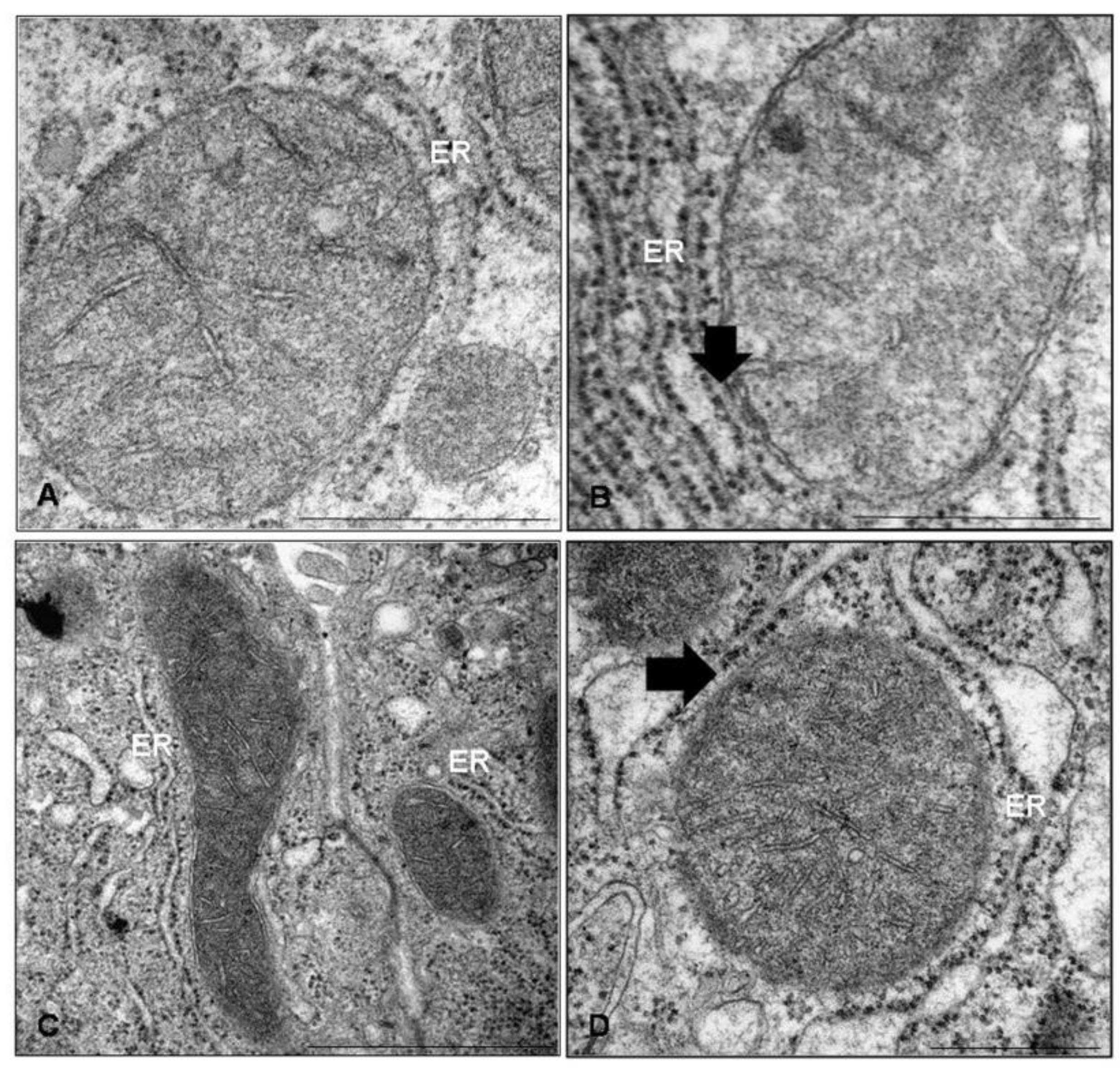
2. Imaging of Mitochondria-ER Interface by Electron Microscopy
3. Imaging of Mitochondria-ER Interface by Dynamic Fluorescent Light Microscopy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Csordas, G.; Weaver, D.; Hajnoczky, G. Endoplasmic reticulum-mitochondrial contactology: Structure and signaling functions. Trends Cell Biol. 2018, 28, 523–540. [Google Scholar] [CrossRef]
- Simmens, T.; Herrera-Cruz, M. Plastic mitochondria-endoplasmic reticulum (ER) contacts use chaperones and tethers to mould their structure and signalling. Curr. Opin. Cell Biol. 2018, 53, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Area-Gomez, E.; Schon, E. On the pathogenesis of Alzheimer’s disease: The MAM hypothesis. FASEB J. 2017, 31, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S.; Patergnani, S.; Missiroli, S.; Morciano, G.; Rimessi, A.; Wiecknowski, M.; Giorgi, C.; Pinton, P. Mitochondrial and endoplasmic reticulum calcium homeostasis and cell death. Cell Calcium 2018, 69, 62–72. [Google Scholar] [CrossRef]
- Rieusset, J. The role of endoplasmic reticulum-mitochondria contact sites in the control of glucose homeostasie: An update. Cell Death Dis. 2018, 9, 388. [Google Scholar] [CrossRef]
- Cho, I.; Adelman, G.; Lim, Y.; Marto, J.; Cho, G.; Golden, J. Ascorbate peroxidase proximity labelling coupled with biochemical fractionation identifies promoters of endoplasmic reticulum-mitochondrial contacts. J. Biol. Chem. 2017, 292, 16382–16392. [Google Scholar] [CrossRef] [PubMed]
- Hung, V.; Lam, S.; Udeshi, N.; Svinkina, T.; Guzman, G.; Mootha, V.; Carr, S.; Ting, A. Proteomic mapping of cytosol-facing outer mitochondrial and ER membranes in living human cells by proximity biotinylation. Elife 2017, 6, e24463. [Google Scholar] [CrossRef]
- Booth, D.; Enyedi, B.; Geiszt, M.; Varnai, P.; Hajnoczky, G. Redox nanodomains are induced by and control calcium signaling at the ER-mitochondrial interface. Mol. Cell 2016, 63, 240–248. [Google Scholar] [CrossRef]
- Rasmussen, N. Mitochondrial structure and the practice of cell biology in the 1950s. J. Hist. Biol. 1995, 28, 381–429. [Google Scholar] [CrossRef]
- Pagliarini, D.; Rutter, J. Hallmarks of a new era in mitochondrial biochemistry. Genes Dev. 2013, 27, 2615–2627. [Google Scholar] [CrossRef] [Green Version]
- Sjostrand, F.S. Electron microscopy of mitochondria and cytoplasmic double membranes. Nature 1953, 171, 30–32. [Google Scholar] [CrossRef] [PubMed]
- Palade, GE. An electron microscope study of the mitochondrial structure. J. Histochem. Cytochem. 1953, 1, 188–211. [Google Scholar] [CrossRef] [PubMed]
- Mannella, C. Structure and dynamics of the mitochondrial inner membrane cristae. Biochim. Biophys. Acta 2006, 1763, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Zick, M.; Rabl, R.; Reichert, A. Cristae formation-linking ultrastructure and function of mitochondria. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 5–19. [Google Scholar] [CrossRef]
- Cogliati, S.; Enriquez, J.; Scorrano, L. Mitochondrial cristae: Where beauty meets functionality. Trends Biochem. Sci. 2016, 41, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Baloyannis, S. Mitochondrial alterations in Alzheimer’s disease. J. Alzheimers Dis. 2006, 9, 119–126. [Google Scholar] [CrossRef]
- Chung, M.; Shu, Y. Ultrastructural changes of mitochondria in the skeletal muscle of patients with amyotrophic lateral sclerosis. Ultrastruct. Pathol. 2002, 26, 3–7. [Google Scholar] [CrossRef]
- Beal, M. Mitochondria and neurodegeneration. Novartis Found. Symp. 2007, 287, 183–192. [Google Scholar]
- Nielsen, J.; Gejl, K.; Hey-Mogensen, M.; Holmberg, H.; Suetta, C.; Krustrup, P.; Elemans, C.; Ortenblad, N. Plasticity in mitochondrial cristae density allows metabolic capacity modulation in human skeletal muscle. J. Physiol. 2017, 595, 2839–2847. [Google Scholar] [CrossRef]
- Jimenez, L.; Laporte, D.; Duvezin-Caubet, S.; Courtout, F.; Sagot, I. Mitochondrial ATP synthase cluster as discrete domains that reorganize with the cellular demand for oxidative phosphorylation. J. Cell. Sci. 2014, 127, 719–726. [Google Scholar] [CrossRef]
- Scorrano, L.; Ashiya, M.; Buttle, K.; Weiler, S.; Oakes, S.; Mannella, C.; Korsmeyer, S. A distinct pathway remodels mitochondrial cristae and mobilizes cytochrome c during apoptosis. Dev. Cell 2002, 2, 55–67. [Google Scholar] [CrossRef]
- Stacchiotti, A.; Favero, G.; Giugno, L.; Golic, I.; Korac, A.; Rezzani, R. Melatonin efficacy in obese leptin-deficient mice heart. Nutrients 2017, 9, 1323. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Favero, G.; Giugno, L.; Lavazza, A.; Reiter, R.; Rodella, L.; Rezzani, R. Mitochondrial and metabolic dysfunction in renal convoluted tubules of obese mice: Protective role of melatonin. PLoS ONE 2014, 9, e111141. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Berridge, M.; Lipp, P.; Bootman, M. Mitochondria are morphologically and functionally heterogeneous within cells. EMBO J. 2002, 21, 1616–1627. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.; Galloway, C.; Jhen, B.; Yu, T. Mitochondrial dynamics in diabetes. Antioxid. Redox Signal. 2011, 14, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Galloway, C.; Lee, H.; Yoon, Y. Mitochondrial morphology-emerging role in bioenergetics. Free Radic. Biol. Med. 2012, 53, 2218–2228. [Google Scholar] [CrossRef]
- Youle, R.; Van der Bliek, A. Mitochondrial fission, fusion, and stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef]
- Varanita, T.; Soriano, M.; Romanello, V.; Zaglia, T.; Quintana-Cabrera, R.; Semenzato, M.; Menabò, R.; Costa, V.; Civiletto, G.; Pesce, P.; et al. The OPA1-dependent mitochondrial cristae remodeling pathway controls atrophic, apoptotic, and ischemic tissue damage. Cell Metab. 2015, 21, 834–844. [Google Scholar] [CrossRef]
- Wells, W. Ribosomes, or the particles of Palade. J. Cell. Biol. 2005, 168, 12. [Google Scholar] [CrossRef]
- Palade, G.; Porter, K. Studies on the endoplasmic reticulum. I. Its identification in cells in situ. J. Exp. Med. 1954, 100, 641–656. [Google Scholar]
- Federovitch, C.; Rou, D.; Hampton, R. The dynamic ER: Experimental approaches and current questions. Curr. Opin. Cell Biol. 2005, 17, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Borgese, N.; Francolini, M.; Snapp, E. Endoplasmic reticulum architecture: Structure in flux. Curr. Opin. Cell Biol. 2006, 18, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Fossati, M.; Borgese, N.; Colombo, S.; Francolini, M. Visualization of endoplasmic reticulum subdomains in cultured cells. J. Vis. Exp. 2014, 84, 50985. [Google Scholar] [CrossRef] [PubMed]
- Fasana, E.; Fossati, M.; Ruggiano, A.; Brambillasca, S.; Hoogenraad, C.; Navone, F.; Francolini, M.; Borgese, N. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 2010, 24, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Blower, M. The endoplasmic reticulum: Structure, function and response to cellular signalling. Cell. Mol. Life Sci. 2016, 73, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Valdez, R.; Flock, D.; Martin, L.; Northington, F. Endoplasmic reticulum pathology and stress response in neurons precede programmed necrosis after neonatal hypoxia-ischemia. Int. J. Dev. Neurosci. 2016, 48, 58–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, A.; Voeltz, G. Endoplasmic reticulum-mitochondria contacts: Function of the junction. Nat. Rev. Mol. Cell. Biol. 2012, 13, 607–625. [Google Scholar] [CrossRef]
- English, A.; Voeltz, G. Endoplasmic reticulum structure and interconnections with other organelles. Cold Spring Harb. Perspect. Biol. 2013, 5, a013227. [Google Scholar] [CrossRef]
- Cohen, S.; Valen, A.; Lippincott-Schwartz, J. Interacting organelles. Curr. Opin. Cell Biol. 2018, 53, 84–91. [Google Scholar] [CrossRef]
- Csordas, G.; Renken, C.; Varnai, P.; Walter, L.; Weaver, D.; Buttle, K.; Balla, T.; Mannella, C.; Hajnoczky, G. Structural and functional features and significance of the physical linkage between ER and mitochondria. J. Cell Biol. 2006, 174, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Csordas, G.; Hainoczky, G. SR/ER-mitochondrial local communication: Calcium and ROS. Biochim. Biophys. Acta 2009, 1787, 1352–1362. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.; Lackner, L.; West, M.; Di Benedetto, J.; Nunnari, J.; Voeltz, G. ER tubules mark sites of mitochondrial division. Science 2011, 334, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pellegrini, L. The coming of age of the mitochondria-ER contact: A matter of thickness. Cell Death Differ. 2016, 23, 1417–1427. [Google Scholar] [CrossRef]
- Wu, H.; Carvalho, P.; Voeltz, G. Here, there, and everywhere: The importance of ER membrane contact sites. Science 2018, 361. [Google Scholar] [CrossRef] [PubMed]
- Raturi, A.; Simmen, T. Where the endoplasmic reticulum and the mitochondrion tie the knot: The mitochondria-associated membrane (MAM). Biochim. Biophys. Acta 2013, 1833, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Vance, J. MAM (mitochondria-associated membranes) in mammalian cells: Lipids and beyond. Biochim. Biophys. Acta 2014, 1841, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Missiroli, S.; Patergnani, S.; Duszynski, J.; Wieckowski, M.; Pinton, P. Mitochondria-associated membranes: Composition, molecular mechanisms, and pathophysiological implications. Antioxid. Redox Signal. 2015, 22, 995–1019. [Google Scholar] [CrossRef]
- Kornmann, B.; Walter, P. ERMES-mediated ER-mitochondria contacts: Molecular hubs for the regulation of mitochondrial biology. J. Cell Sci. 2010, 123, 1389–1393. [Google Scholar] [CrossRef]
- Kornmann, B.; Currie, E.; Collins, S.; Schuldiner, M.; Nunnari, J.; Weissman, J.; Walter, P. An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 2009, 325, 477–481. [Google Scholar] [CrossRef]
- Scharwey, M.; Tatsuta, T.; Langer, T. Mitochondrial lipid transport at a glance. J. Cell Sci. 2013, 126, 5317–5323. [Google Scholar] [CrossRef] [Green Version]
- Flis, V.; Daum, G. Lipid transport between the endoplasmic reticulum and mitochondria. Cold Spring Harb. Perspect. Biol. 2013, 5, a013235. [Google Scholar] [CrossRef] [PubMed]
- Tasseva, G.; Bai, H.; Davidescu, M.; Haromy, A.; Michelakis, E.; Vance, J. Phosphatidylethanolamine deficiency in mammalian mitochondria impairs oxidative phosphorylation and alters mitochondrial morphology. J. Biol. Chem. 2013, 288, 4158–4173. [Google Scholar] [CrossRef] [PubMed]
- Rockenfeller, P.; Koska, M.; Pietrocola, F.; Minois, N.; Knittelfelder, O.; Sica, V.; Franz, J.; Carmona-Gutierrez, D.; Kroemer, G.; Madeo, F. Phosphatidylethanolamine positively regulates autophagy and longevity. Cell Death Differ. 2015, 22, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamasaki, M.; Furuta, N.; Matsuda, A.; Nezu, A.; Yamamoto, A.; Fujita, N.; Ooomori, I.; Noda, T.; Haraguchi, T.; Hiraoka, Y. Autophagosomes form at ER-mitochondria contacts site. Nature 2013, 495, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Garcin, P.; Fu, M.; Masoudi, M.; St-Pierre, P.; Panté, N.; Nabi, I. Distinct mechanisms controlling rough and smooth endoplasmic reticulum contacts with mitochondria. J. Cell Sci. 2015, 128, 2759–2765. [Google Scholar] [CrossRef]
- Sood, A.; Jeyaraju, V.; Prudent, J.; Caron, A.; Lemieux, P.; McBride, H.; Laplante, M.; Toth, K.; Pellegrini, L. A mitofusin-2-dependent inactivating cleavage of Opa 1 links changes in mitochondria cristae and ER contacts in the postprandial liver. Proc. Natl. Acad. Sci. USA 2014, 111, 16017–16022. [Google Scholar] [CrossRef] [PubMed]
- Rizzuto, R.; Pinton, P.; Carrington, W.; Fay, F.; Fogarty, K.; Lifshitz, L.; Tuft, R.; Pozzan, T. Close contacts with the endoplasmic reticulum as determinants of mitochondrial Ca2+ responses. Science 1998, 280, 1763–1766. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Vicencio, J.; Parra, V.; Troncoso, F.; Munoz, J.; Bui, M.; Quiroga, C.; Rodriguez, A.; Verdejo, H.; Ferreira, J.; et al. Increased ER-mitochondrial coupling promotes mitochondrial respiration and bioenergetics during early phases of ER stress. J. Cell Sci. 2011, 124, 2143–2152. [Google Scholar] [CrossRef] [Green Version]
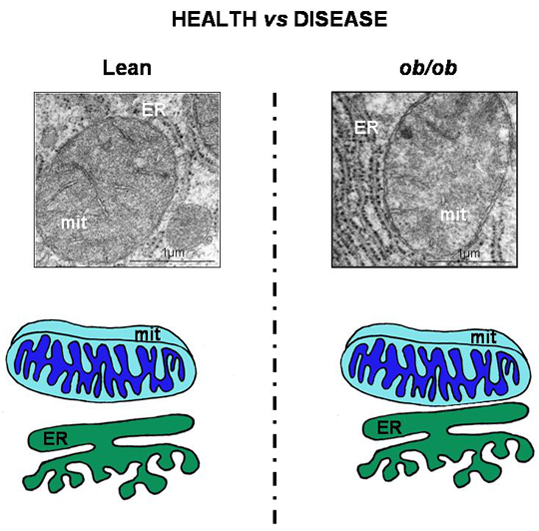
- Arruda, P.; Pers, B.; Parlakgül, G.; Güney, E.; Inouye, K.; Hotamisligil, G. Chronic enrichment of hepatic endoplasmic reticulum-mitochondria contact leads to mitochondrial dysfunction in obesity. Nat. Med. 2014, 20, 1427–1435. [Google Scholar] [CrossRef]
- Stacchiotti, A.; Favero, G.; Lavazza, A.; Golic, I.; Aleksic, M.; Korac, A.; Rodella, L.; Rezzani, R. Hepatic macrosteatosis is partially converted to microsteatosis by melatonin supplementation in ob/ob mice non-alcoholic fatty liver disease. PLoS ONE 2016, 11, e0148115. [Google Scholar] [CrossRef]
- Tang, B. Sirt1 and the mitochondria. Mol. Cells 2016, 39, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Tubbs, E.; Theurey, P.; Vial, G.; Bendridi, N.; Bravard, A.; Chavin, M.; Ti-Cao, J.; Zoulim, F.; Bartosch, B.; Ovize, M.; et al. Mitochondria-associated endoplasmic reticulum membrane (MAM) integrity is required for insulin signaling and is implicated in hepatic insulin resistance. Diabetes 2014, 63, 3279–3294. [Google Scholar] [CrossRef] [PubMed]
- Paillusson, S.; Stoica, R.; Gomez-Suaga, P.; Lau, D.; Mueller, S.; Miller, T.; Miller, C. There’s something wrong in my MAM; the ER-mitochondria axis and neurodegenerative diseases. Trends Neuros. 2016, 39, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Whiteus, C.; Xu, S.; Hayworth, K.; Weinberg, R.; Hess, H.; De Camilli, P. Contacts between the endoplasmic reticulum and other membrane in neurons. PNAS 2017, 114, E4859–E4867. [Google Scholar] [CrossRef] [PubMed]
- Csordas, G.; Várnai, P.; Golenár, T.; Roy, S.; Purkins, G.; Schneider, T.; Balla, T.; Hajnóczky, G. Imaging interorganelle contacts and local calcium dynamics at the ER-mitochondrial interface. Mol. Cell 2010, 39, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Elgass, K.; Smith, E.; Le Gros, M.; Larabell, C.; Ryan, M. Analysis of ER-mitochondria contacts using correlative fluorescent microscopy and soft X ray tomography of mammalian cells. J. Cell Sci. 2015, 128, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Harmon, M.; Larkman, P.; Hardingham, G.; Jackson, M.; Skehel, P. A Bi-fluorescence complementation system to detect associations between the endoplasmic reticulum and mitochondria. Sci. Rep. 2017, 7, 174–167. [Google Scholar] [CrossRef]
- Naon, D.; Zaninello, M.; Giacomello, M.; Varanita, T.; Grespi, F.; Lakshminaranayan, S.; Serafini, A.; Semenzato, M.; Herkenne, S.; Hernandez-Alvarez, M.; et al. Critical reappraisal confirms that mitofusin 2 is an endoplasmic reticulum-mitochondria tether. PNAS 2016, 113, 11249–11254. [Google Scholar] [CrossRef]
- Galmes, R.; Houcine, A.; Van Vlieta, A.; Agostinis, P.; Jackson, C.; Giordano, F. ORP5/ORP8 localize to endoplasmic reticulum-mitochondria contacts and are involved in mitochondrial function. EMBO Rep. 2016, 17, 800–810. [Google Scholar] [CrossRef]
- Cieri, D.; Vicario, M.; Giacomello, M.; Vallese, F.; Filadi, R.; Wagner, T.; Pozzan, T.; Pizzo, P.; Scorrano, L.; Brini, M.; et al. SPLICS: A split green fluorescent protein-based contact site sensor for narrow and wide heterotypic organelle juxtaposition. Cell Death Differ. 2018, 25, 1131–1145. [Google Scholar] [CrossRef]
- Theurey, P.; Tubbs, E.; Vial, G.; Jacquemetton, J.; Bendridi, N.; Chauvin, M.; Alam, M.; Le Romancer, M.; Vidal, H.; Rieusset, J. Mitochondria-associated endoplasmic reticulum membranes allow adaptation of mitochondrial metabolism to glucose availability in the liver. J. Mol. Cell Biol. 2016, 8, 129–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieusset, J.; Fauconnier, J.; Paillard, M.; Belaidi, E.; Tubbs, E.; Chauvin, A.; Durand, A.; Bravard, A.; Teixeira, G.; Bartosch, B.; et al. Disruption of calcium transfer from ER to mitochondria links alterations of mitochondria-associated ER membrane integrity to hepatic insulin resistance. Diabetologia 2016, 59, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Holcman, D.; Parutto, P.; Chambers, J.; Fantham, M.; Young, L.; Marciniak, S.; Kaminski, C.; Ron, D.; Avezov, E. Single particle trajectories reveal active endoplasmic reticulum luminal flow. Nature Cell Biol. 2018, 20, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Erpazoglou, Z.; Mouton-Liger, F.; Corti, O. From dysfunctional endoplasmic reticulum mitochondria coupling to neurodegeneration. Neurochem. Int. 2017, 109, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Angebault, C.; Fauconnier, J.; Patergnani, S.; Rieusset, J.; Danese, A.; Affortit, C.; Jagodzinska, J.; Megy, C.; Quiles, M.; Cazevielle, C.; et al. ER-mitochondria cross talk is regulated by the Ca2+ sensor NCS1 and is impaired in Wolfram syndrome. Sci. Signal. 2018, 11, 1380. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Microscopy/Methods [References] | Advantages | Disadvantages |
|---|---|---|
| Transmission Electron Microscopy-TEM [40,41,42,59,60] | Elective for nanoscale resolution | Not suitable for living cells Technically hard Expensive |
| Cryo-TEM plus tomography [43] | 3D images in small volumes | Not suitable for living cells |
| Scanning Electron Microscopy-SEM FIB [64] | Good resolution 3D images in large volumes | Not suitable for living cells |
| Confocal Laser Fluorescence Microscopy [41,48,49,52,56,62,66,70,71,72] | Suitable for living cells and dynamic events Quantification contacts | Toxic for cells after long time Lower Resolution Unstable Specific probes required |
| Total Internal Reflection Fluorescence Microscopy [69] | High Brightness Suitable for dynamic events | Sensitive to refraction index Thermogenic |
| Super-resolution Microscopy [67,73,74] | High Brightness 3D images Quantification contacts | Specific probes required Expensive |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stacchiotti, A.; Favero, G.; Lavazza, A.; Garcia-Gomez, R.; Monsalve, M.; Rezzani, R. Perspective: Mitochondria-ER Contacts in Metabolic Cellular Stress Assessed by Microscopy. Cells 2019, 8, 5. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8010005
Stacchiotti A, Favero G, Lavazza A, Garcia-Gomez R, Monsalve M, Rezzani R. Perspective: Mitochondria-ER Contacts in Metabolic Cellular Stress Assessed by Microscopy. Cells. 2019; 8(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8010005
Chicago/Turabian StyleStacchiotti, Alessandra, Gaia Favero, Antonio Lavazza, Raquel Garcia-Gomez, Maria Monsalve, and Rita Rezzani. 2019. "Perspective: Mitochondria-ER Contacts in Metabolic Cellular Stress Assessed by Microscopy" Cells 8, no. 1: 5. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8010005