NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes


1
Program of Immunology, Department of Pharmacology-Physiology, Faculty of Medicine and Health Sciences, University of Sherbrooke, Sherbrooke, QC J1H 5N4, Canada
2
Program of Immunology, Department of Pediatrics, CR-CHUS, Faculty of Medicine and Health Sciences, University of Sherbrooke, Sherbrooke, QC J1H 5N4, Canada
*
Author to whom correspondence should be addressed.
Cells 2019, 8(5), 400; https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050400
Submission received: 10 April 2019
/
Revised: 25 April 2019
/
Accepted: 28 April 2019
/
Published: 30 April 2019
Abstract
:Uptake of glutamate from the extracellular space and glutamate release to neurons are two major processes conducted by astrocytes in the central nervous system (CNS) that protect against glutamate excitotoxicity and strengthen neuronal firing, respectively. During inflammatory conditions in the CNS, astrocytes may lose one or both of these functions, resulting in accumulation of the extracellular glutamate, which eventually leads to excitotoxic neuronal death, which in turn worsens the CNS inflammation. NLRX1 is an innate immune NOD-like receptor that inhibits the major inflammatory pathways. It is localized in the mitochondria and was shown to inhibit cell death, enhance ATP production, and dampen oxidative stress. In the current work, using primary murine astrocyte cultures from WT and Nlrx1-/- mice, we demonstrate that NLRX1 potentiates astrocytic glutamate uptake by enhancing mitochondrial functions and the functional activity of glutamate transporters. Also, we report that NLRX1 inhibits glutamate release from astrocytes by repressing Ca2+-mediated glutamate exocytosis. Our study, for the first time, identified NLRX1 as a potential regulator of glutamate homeostasis in the CNS.
1. Introduction
Astrocytes are the most numerous glial cell type in the central nervous system (CNS). They perform many pivotal functions associated with neuronal support and maintenance of the CNS homeostasis [1]. One of these crucial functions is to uptake excess synaptically-released glutamate (the major excitatory neurotransmitter in the CNS) [2,3] from the extracellular space, metabolize it, and send it back to neurons [4,5]. This glutamate uptake is mediated primarily by the excitatory amino acid transporters 1 and 2 (EAAT1 and EAAT2), expressed by astrocytes (known in mice as glutamate-aspartate transporter (GLAST) and glutamate transporter-1 (GLT-1), respectively) [6,7,8,9,10,11]. In astrocytes, under physiological conditions, the uptake of glutamate against its concentration gradient relies on glutamate transporters and Na+/K+ ATPase that consumes high levels of intracellular adenosine triphosphate (ATP) [12,13]. Many factors in the CNS influence the expression, trafficking, and functional activity of glutamate transporters in astrocytes, including hormones, growth factors, inflammatory mediators, and oxidative stress [14,15,16].
Recent studies suggested that, along with glutamate uptake, astrocytes release glutamate, which helps synchronize and intensify firing of the surrounding neurons [4,17,18]. Astrocytic glutamate release is mediated mostly by Ca2+-dependent exocytosis [19]. In astrocytes, part of the sequestrated glutamate is transferred into small intracellular vesicles [20,21] by activation of the vesicular glutamate transporters 1 and 2 (VGLUT1 and VGLUT2), derived by the proton gradient generated by the vacuolar (H+) ATPase (V-ATPase) [22,23,24]. Astrocytes also express a Ca2+ sensor (synaptotagmin 4, 7, or 11) [25,26,27] and vesicular fusion proteins (vesicle-associated membrane protein 2 and 3 (VAMP2 and VAMP3)) [20,28]. In response to intracellular Ca2+ elevation, caused by Ca2+ release from the endoplasmic reticulum (ER), vesicular fusion proteins (VAMP2 and VAMP3) fuse with the astrocyte cell membrane fusion proteins (syntaxin and soluble N-ethylmaleimide-sensitive factor attachment protein 23 (SNAP23)) [29,30,31], resulting in glutamate release from the vesicles into the extracellular space.
In the context of CNS pathologies, astrocytes respond to inflammation by losing some of their vital functions or acquiring some deleterious effects that aggravate inflammatory conditions in the CNS and delay the processes of recovery [1]. Previous studies showed that neuropathological conditions in the CNS, such as brain trauma, infection by human immunodeficiency virus (HIV), multiples sclerosis (MS), amyotrophic lateral sclerosis (ALS), Alzheimer’s disease (AD), and Parkinson’s disease (PD) are associated with accumulation of glutamate in the extracellular space, caused by reduced glutamate uptake and/or increased glutamate release by astrocytes [32,33,34,35,36,37]. Excess extracellular glutamate induces hyperstimulation of glutamate receptors in neurons and eventually leads to neuronal death, in a process known as “glutamate excitotoxicity” [38]. The excess of extracellular glutamate and subsequent neuronal death, in turn, enhance the inflammatory response and worsen the pathological conditions in the CNS [38].
NOD-like receptors (NLRs) are cytosolic innate immune molecules that can exert either positive or negative effects on inflammation in the CNS [39]. NLRX1 is a recently discovered anti-inflammatory NLR that inhibits nuclear factor-κB (NF-κB) and RIG-1-MAVs signaling pathways, thus inhibiting antimicrobial immune responses [40,41,42,43,44] and sterile inflammations [45]. Since the introduction of Nlrx1-/- mice, many studies implicated NLRX1 in the development of various pathologies. For example, Nlrx1-/- mice show excessive inflammatory response following Influenza virus infection and LPS treatment [41]. Also, Nlrx1-/- mice show exacerbated severity of inflammatory bowel disease (IBD) [46] and increased incidence of colitis-associated colonic cancer [45]. In the CNS, lack of Nlrx1 in mice is associated with an excessive inflammation following CNS trauma [47], earlier onset, and a more aggressive course of the experimental autoimmune encephalomyelitis (EAE), a mouse model of MS [48]. Moreover, using a neuroblastoma cell line, NLRX1 was shown to inhibit neuronal death and redirect rotenone-treated neurons towards apoptosis instead of necrosis [49]. Unlike other NLRs, NLRX1 is located at the mitochondria. It enhances various mitochondrial functions and activities such as ATP production and respiration while inhibiting oxidative stress and apoptosis [44,49,50,51,52,53,54].
In the current study, we investigated the role of NLRX1 in glutamate uptake and release by primary murine astrocytes, and the potential mechanisms by which NLRX1 mediates its effects.
2. Materials and Methods
2.1. Mice
All mice handling and manipulations were approved by the Institutional Animal Care and Use Committee at the University of Sherbrooke (Protocols #280-15, 4 April 2017) according to the Canadian Council on Animal Care. All mice were bred on C57/BL6J background. Wild-type (WT) mice were bred in-house in the same conditions as Nlrx1-/- mice that were kindly provided by Dr. Jenny P. Y. Ting (Chapel Hill, NC, USA).
2.2. Primary Mouse Astrocyte Cultures
Glial cultures were prepared from 1-day-old pups, as previously described [55]. Pups were sacrificed by decapitation, and brains were harvested and placed in 100 mm culture plates. Brain tissue was dissociated by a commercial razor blade, followed by triturating in 10 mL DMEM/F12 medium (Wisent Inc., Montreal, QC, Canada) containing 10% deactivated fetal bovine serum (dFBS), 2 mM l-glutamine, 1% MEM amino acid, 1% sodium pyruvate, and 1% penicillin-streptomycin and amphotericin B (all from Wisent Inc., Montreal, QC, Canada). Dissociated tissue was passed through 70 µm cell strainer to remove tissue debris. Cells were plated in 100 mm cell culture plates (Corning Inc., Brooklyn, NY, USA) with DMEM/F12 complete medium and incubated in 37 °C incubator with 5% CO2. The medium was changed every 2–3 days to wash out cells other than glial cells. After 21 days, glial cultures were resuspended in 10% dimethyl sulfoxide (DMSO) in dFBS (freezing medium) and were frozen at −80 °C. One week before the experiments, cells were thawed and reseeded in 100 mm culture plates, in complete DMEM/F12 medium. Cells were stained with CD11b (eBioscience/Thermofisher scientific, Waltham, Massachusetts, USA # 12-0112-81) as a marker for microglia and the percentage of CD11b-expressing cells was measured by flow cytometry. In our experiments, we used cultures containing less than 10% CD11b+ cells (astrocytes ≥90%) since additional purification of astrocytes did not affect the glutamate uptake or release.
2.3. Glutamate Uptake and Release Assay
The assay was modified from Piao et al. 2015 [56]. 100,000 astrocytes were seeded in each well of a 96-well plate, and washed 2 times with Hank’s Balanced Salt Solution (HBSS) containing Ca2+ (Wisent Inc., Montreal, QC, Canada): 1.26 mM CaCl2 (anhydrous), 5.36 mM KCl, 0.44 mM KH2PO4, 0.811 mM MgSO4 (anhydrous), 137 mM NaCl, 0.336 mM Na2HPO4 (anhydrous), 4.166 mM NaHCO3, and 5.55 mM d-glucose, pH 7.25 ± 0.15 or Ca2+-free Lock’s solution: 140 mM NaCl, 4.7 mM KCl, 1.2 mM KH2PO4, 1.2 MgSO4, 11 mM glucose, and 15 mM HEPES-NaOH. For glutamate uptake, cells were incubated with 100 or 200 µM glutamate in the Ca2+-containing HBSS for 4 h, while for glutamate release, astrocytes were incubated in Ca2+-containing HBSS or Ca2+-free Lock’s solution for 1 h, in the 37 °C with 5% CO2 incubator. Then, culture supernatant was collected, and glutamate concentration in the medium was measured using a glutamate colorimetric assay kit (Sigma-Aldrich, Oakville, ON, Canada # MAK004 or Abcam, Toronto, ON, Canada #ab83389) according to the manufacturer’s instructions. Glutamate uptake by astrocytes was measured by subtracting the amount of glutamate measured in the medium from the amount initially added to the cells. Both glutamate uptake and release by astrocytes were normalized to the amount of protein in the corresponding cells, measured by Bradford assay (Bio-Rad, Montreal, QC, Canada).
2.4. Quantitative Real-Time PCR (qPCR)
RNA was extracted from astrocytes using TRIzol reagent (Life Technologies Inc./Thermofisher Scientific, Waltham, MA, USA # 15596-018) according to the manufacturer’s instructions. cDNA was synthesized from RNA, using Oligo(dT) primer (IDT, Coralville, IA, USA), dNTP Mix, M-MuLV Reverse Transcriptase, M-MuLV Reverse Transcriptase Buffer, and RNase inhibitor (all from New England Biolabs, Whitby, ON, Canada), as previously described [55]. qPCR was performed using KiCqStart® SYBR® Green qPCR ReadyMix (Sigma-Aldrich, Oakville, ON, Canada # KCQS00). The ΔΔCT method was used to calculate the relative gene expression to 18S as a housekeeping gene [57]. Primer sequences used (IDT, Coralville, IA, USA) are shown in Table 1.
2.5. Flow Cytometry Staining
To measure the total protein expression of glutamate transporters, intracellular staining was performed (protocol modified from Gharagozloo et al. 2018 and Schwarz et al. 2013) [58,59]. WT and Nlrx1-/- astrocytes were washed with phosphate-buffered saline (PBS), fixed, permeabilized, and blocked with 5% dFBS in washing buffer. Cells were stained with the anti-GLT-1 antibody (Novus Biologicals, Centennial, CO, USA # NBP1-20136) diluted 1:100 or anti-GLAST antibody (Novus Biologicals, Centennial, CO, USA # NB100-1869) diluted 1:200 and incubated for 30–40 min. Astrocytes were washed twice and incubated with the secondary anti-rabbit IgG antibody, Alexa Fluor® 555 Conjugated (New England Biolabs, Whitby, ON, Canada #4413) diluted 1:1000, for 20 min. Cells were washed twice after the secondary antibody and resuspended in PBS.
To measure the cell surface expression of the transporters, astrocytes were washed and stained using the same previous procedure, but with no cell fixation or permeabilization.
To detect the activity of reactive oxygen species (ROS), dihydrorhodamine 123 (DHR) was added to the cells to a final concentration of 0.5 µg/mL and incubated for 15 min at 37 °C, then cells were resuspended in PBS (protocol modified from Gris et al. 2008 and Farrell et al. 2011) [60,61].
Sample acquisition was realized using Beckman Coulter CytoFlex (Beckman Coulter, Brea, CA, USA). Data analysis was performed, and histograms produced using CytExpert 2.3 software (Beckman Coulter, Brea, CA, USA).
2.6. Measurement of Intracellular ATP
The assay was performed using the ATP bioluminescent assay kit (Sigma-Aldrich, Oakville, ON, Canada # FLAA) according to the manufacturer’s instructions, modified from Marcaida et al. 1997 [62]. 100,000 astrocytes from WT and Nlrx1-/- mice were lysed with 400 µL of somatic cell ATP-releasing reagent (Sigma-Aldrich, Oakville, ON, Canada # FLSAR). In a white opaque 96-well plate, 100 µL of the ATP reaction mix was added to each well and incubated for 3 min. Samples (100 µL) from the ATP standard or the cell lysate were added to the reaction mix and vigorously mixed. Immediately, using a luminometer, the amount of light produced from the reaction was measured, which reflected the amount of ATP in each well.
2.7. Measurement of Mitochondrial DNA (mtDNA)
DNA was extracted from astrocytes using TRIzol reagent (Life Technologies Inc./Thermofisher Scientific, Waltham, MA, USA # 15596-018) according to the manufacturer’s instructions. qPCR was performed, as previously described, to compare the relative amount of mtDNA between WT and Nlrx1-/- astrocytes [63], using 100 ng of the extracted DNA and KiCqStart® SYBR® Green qPCR ReadyMix (Sigma-Aldrich, Oakville, ON, Canada # KCQS00). To estimate the amount of mtDNA, the mitochondrial DNA region (D-loop) was amplified, using the two primer sequences: D1 (5′-CCC AAG CAT ATA AGC TAG TAC-3′) and D2 (5′-ATA TAA GTC ATA TTT TGG GAA CTA C-3′), with the thermal cycling protocol 95 °C for 20 s, 55 °C for 20 s, 72 °C for 80 s for 30 cycles after an initial denaturation. To estimate the amount of nuclear DNA as a reference, the (apo-B) region was amplified, using the two primer sequences: 5′-CGT GGG CTC CAG CAT TCT A-3′ and 5′-TCA CCA GTC ATT TCT GCC TTT G-3′, with the two-step thermal cycling protocol 95 °C for 10 s and 60 °C for 30 s for 40 cycles after an initial denaturation at 95 °C for 1 min. The relative amount of mtDNA to the nuclear DNA was calculated by the ΔΔCT method.
2.8. Statistical Analysis
All statistical analysis was conducted using GraphPad Prism 8 software (GraphPad, San Diego, CA, USA). Results were expressed as the mean ± standard error of the mean (SEM). Statistical differences between WT and Nlrx1-/- astrocytes were assessed by Mann–Whitney test. Glutamate uptake and glutamate release using different treatments were assessed by two-way ANOVA followed by Tukey’s test. The significance level was set at p < 0.05.
3. Results
3.1. NLRX1 Inhibits Excess Glutamate Release and Enhances Glutamate Uptake by Astrocytes
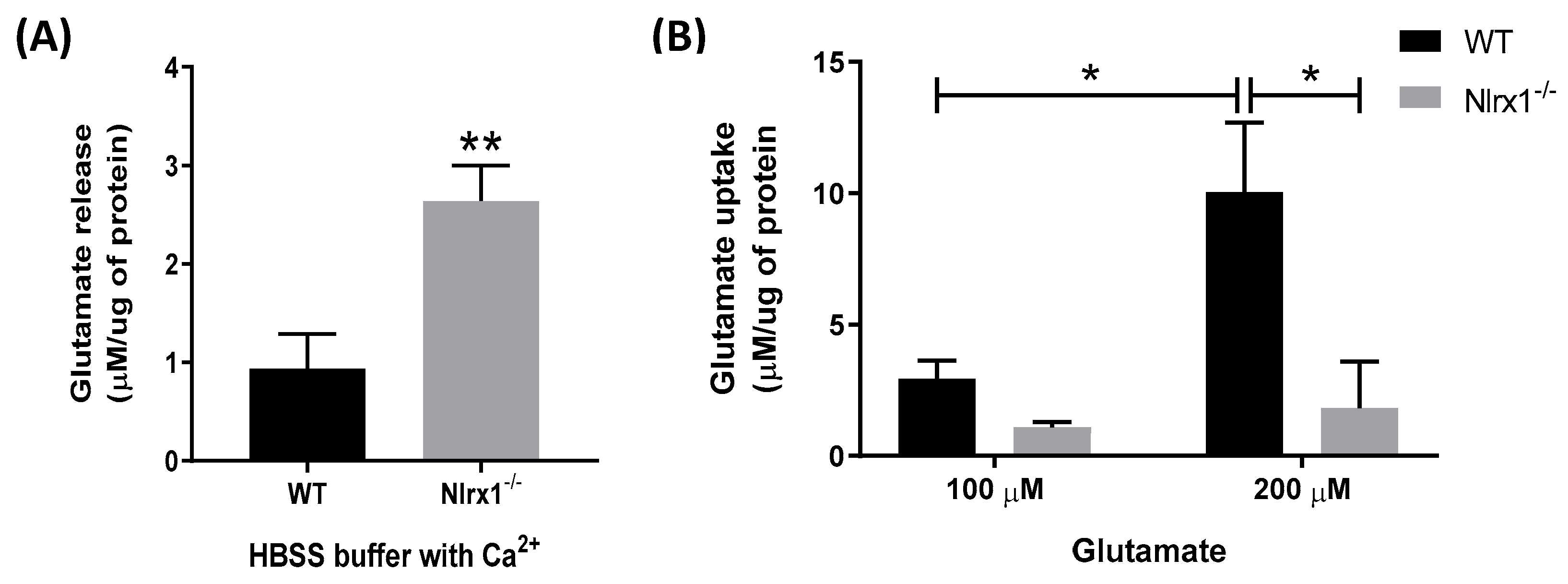
To determine the role of NLRX1 in the glutamate release and uptake by astrocytes, we incubated primary astrocyte cultures from WT and Nlrx1-/- mice with or without glutamate in a Ca2+-containing medium. Our data shows that in the cultures incubated with the glutamate-free medium for 4 h (Figure 1A) or 1 h (Figure 2A), Nlrx1-/- astrocytes released significantly higher levels of glutamate compared to WT astrocytes. In the cultures incubated with 100 µM glutamate, there was no significant difference in the glutamate uptake between WT and Nlrx1-/- astrocytes (Figure 1B). However, when we challenged both cultures with a higher concentration of glutamate (200 µM), WT astrocytes significantly enhanced their glutamate uptake, while there was no significant change in the Nlrx1-/- astrocytes’ glutamate uptake (Figure 1B). Therefore, after incubation with 200 µM glutamate, WT astrocytes had a significantly higher glutamate uptake (fivefold) than Nlrx1-/- astrocytes (Figure 1B).
3.2. Nlrx1-/- Astrocytes’ Excess Glutamate Release Is Ca2+-Dependent
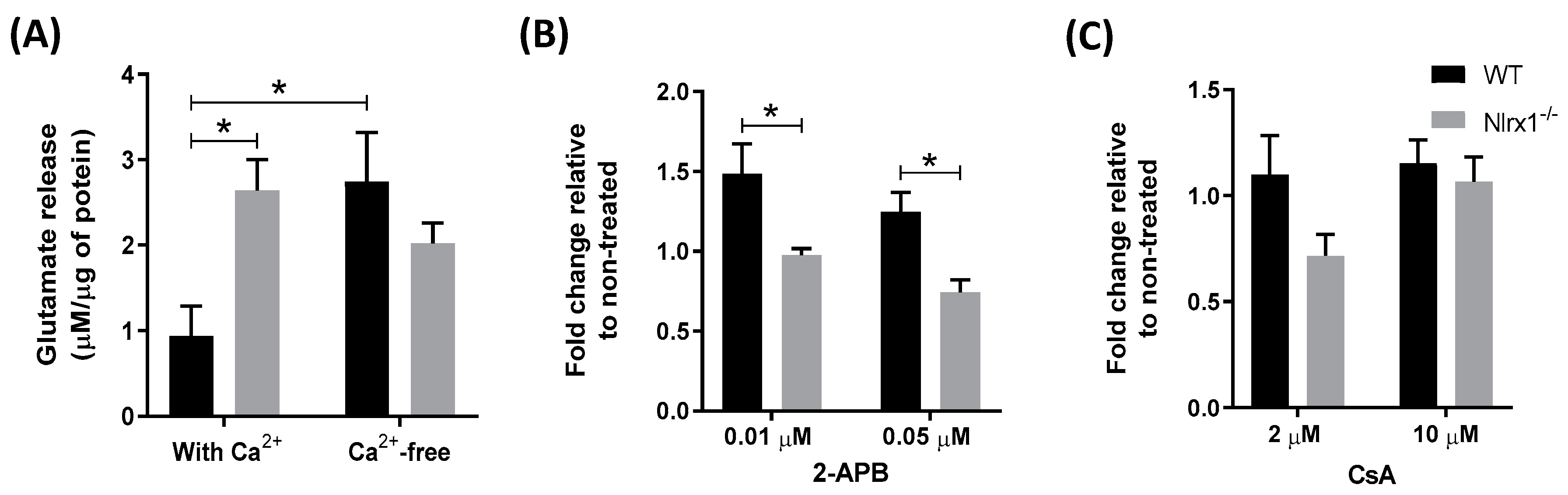
Given that glutamate release by astrocytes is mediated primarily by the elevation of intracellular Ca2+ levels [18], first, we examined whether the presence of extracellular Ca2+ plays a role in glutamate release from Nlrx1-/- astrocytes. We incubated WT and Nlrx1-/- astrocyte cultures in a Ca2+-containing or Ca2+-free medium for 1 h, followed by the measurement of glutamate in the medium. We observed that removal of Ca2+ from the medium resulted in a significant increase in glutamate release from WT astrocytes, while no significant changes in glutamate release were observed in the Nlrx1-/- astrocytes (Figure 2A). This suggests that the presence of extracellular Ca2+ does not have a significant effect on glutamate release in Nlrx1-/- cultures.
We further assessed whether this glutamate release is mediated by Ca2+ release from the intracellular Ca2+ stores, including ER and mitochondria. We incubated WT and Nlrx1-/- astrocyte cultures with different concentrations of 2-Aminoethyl diphenylborinate (2-APB, an inhibitor of inositol-1,4,5-trisphosphate (IP3) receptors that inhibits Ca2+ release from the ER) [64] or Cyclosporin A (CsA, an inhibitor of mitochondrial Ca2+ release) [65] in the Ca2+-free medium. A significant reduction was detected in the glutamate release from 2-APB-treated Nlrx1-/- astrocytes at both concentrations, compared to WT (Figure 2B). By contrast, in cultures treated with CsA, no significant change was detected in both genotypes (Figure 2C).
3.3. Glutamate Release by Nlrx1-/- Astrocytes Is Mediated By Exocytosis
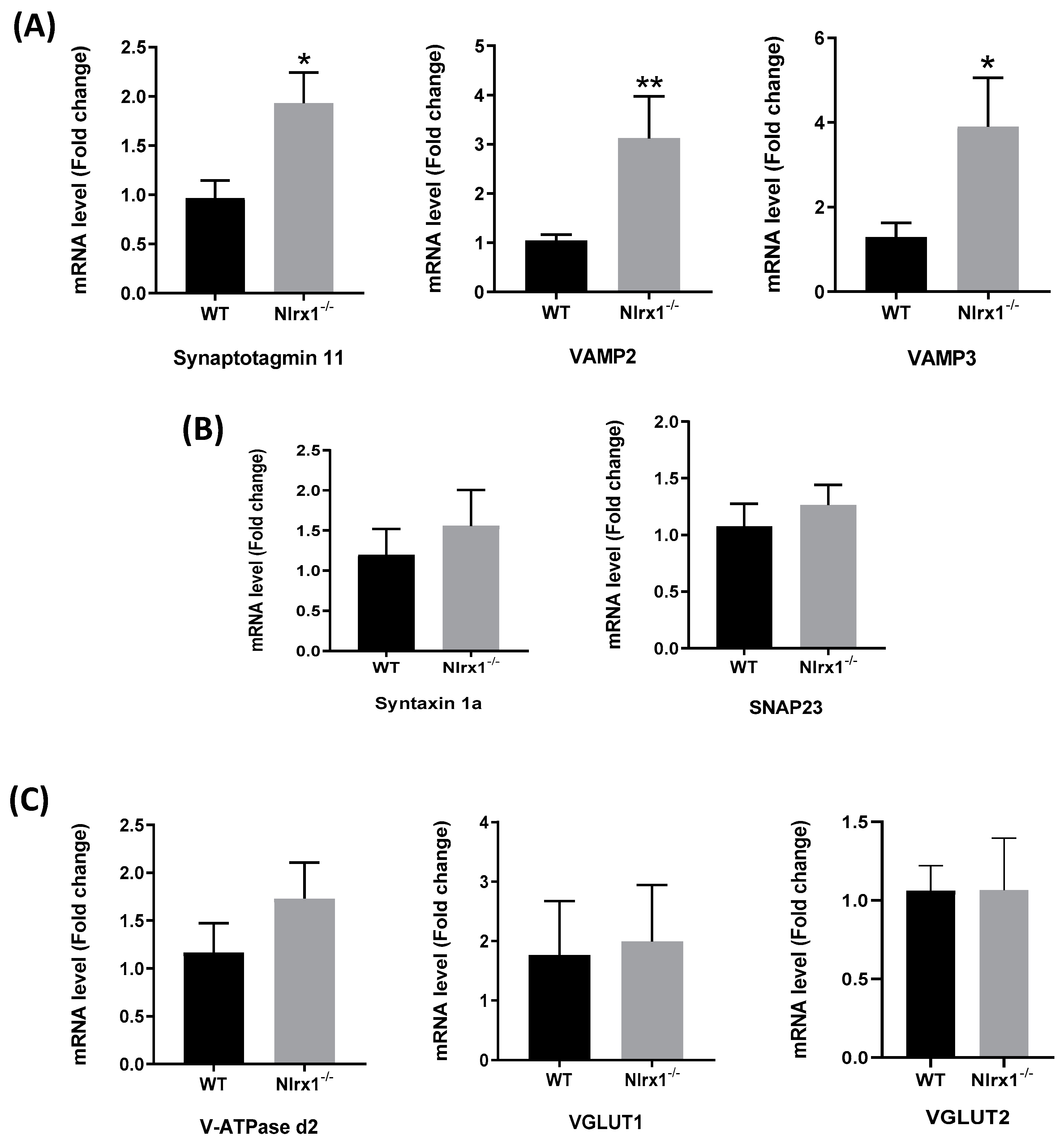
Since we found that the NLRX1-mediated glutamate release is Ca2+-dependent, we further evaluated whether it is mediated by exocytosis. We measured gene expression of the proteins involved in exocytosis, upstream and downstream of the Ca2+ release from the ER, in WT and Nlrx1-/- astrocyte cultures. The results demonstrated that the mRNA expression of the astrocytic Ca2+ sensor, synaptotagmin 11, and the vesicular fusion proteins (VAMP2 and VAMP3) was significantly upregulated in Nlrx1-/- astrocytes relative to WT (Figure 3A), while no significant change was observed in the mRNA expression of the cell membrane fusion proteins (Syntaxin 1a and SNAP23) (Figure 3B) or the proteins upstream of the Ca2+ release (V-ATPase d2, VGLUT1, and VGLUT2) (Figure 3C).
3.4. mRNA and Protein Expression of Glutamate Transporters in Astrocytes
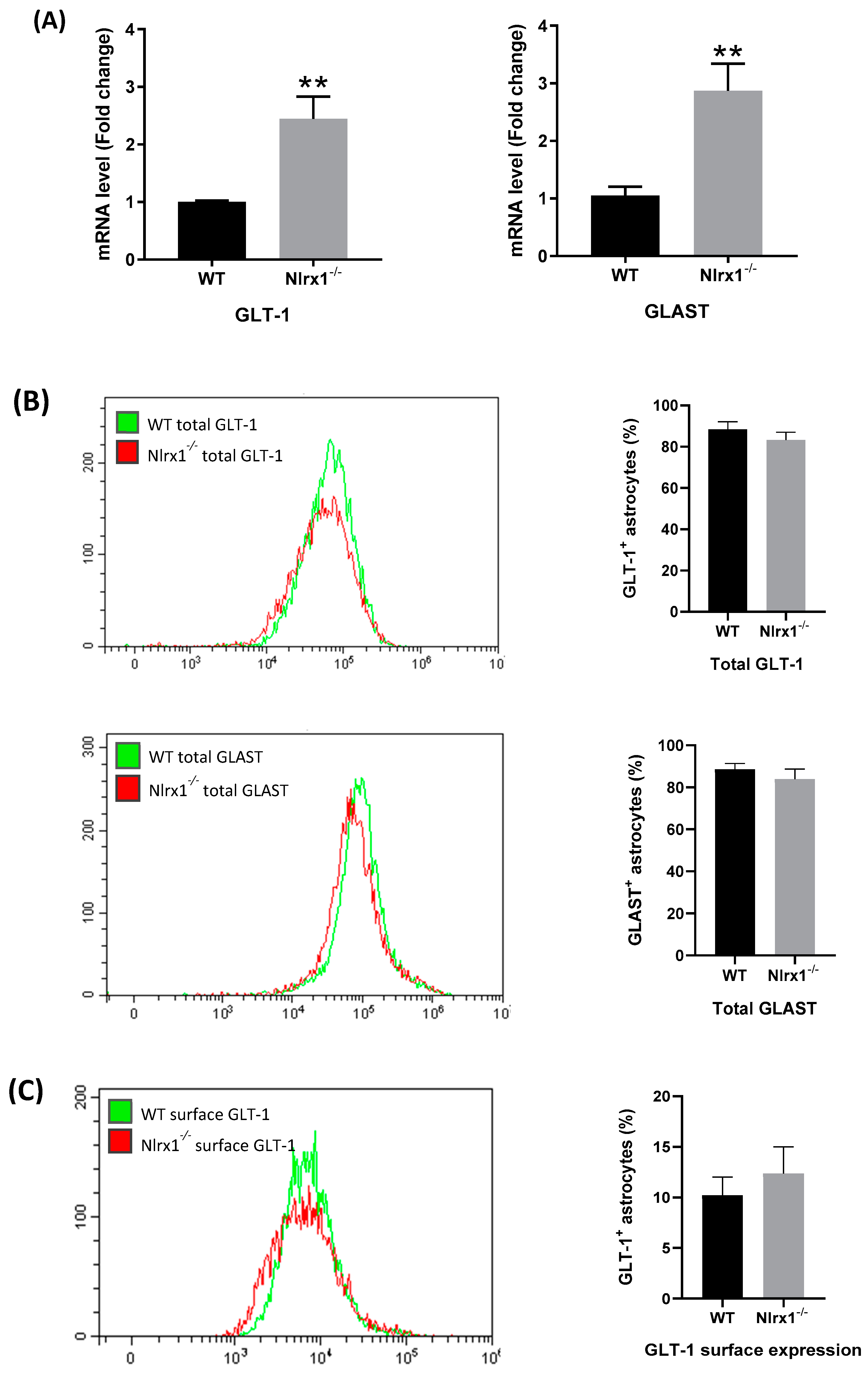
To further investigate the mechanism by which NLRX1 enhances glutamate uptake, we measured the relative gene expression of the astrocytes’ glutamate transporters, GLT-1 and GLAST, in WT and Nlrx1-/- astrocyte cultures, using qPCR. The mRNA expression of both transporters was significantly higher in Nlrx1-/- astrocyte cultures relative to WT (Figure 4A). In parallel, we stained WT and Nlrx1-/- astrocytes with anti-GLT-1 or anti-GLAST antibodies and quantified the total protein expression and the cell surface expression of both transporters by flow cytometry. As shown in Figure 4B,C, no significant change was detected in either GLT-1 or GLAST total protein expression (Figure 4B) or cell surface expression (Figure 4C) between astrocytes of both genotypes.
3.5. NLRX1 Enhances Mitochondrial Functions in Astrocytes
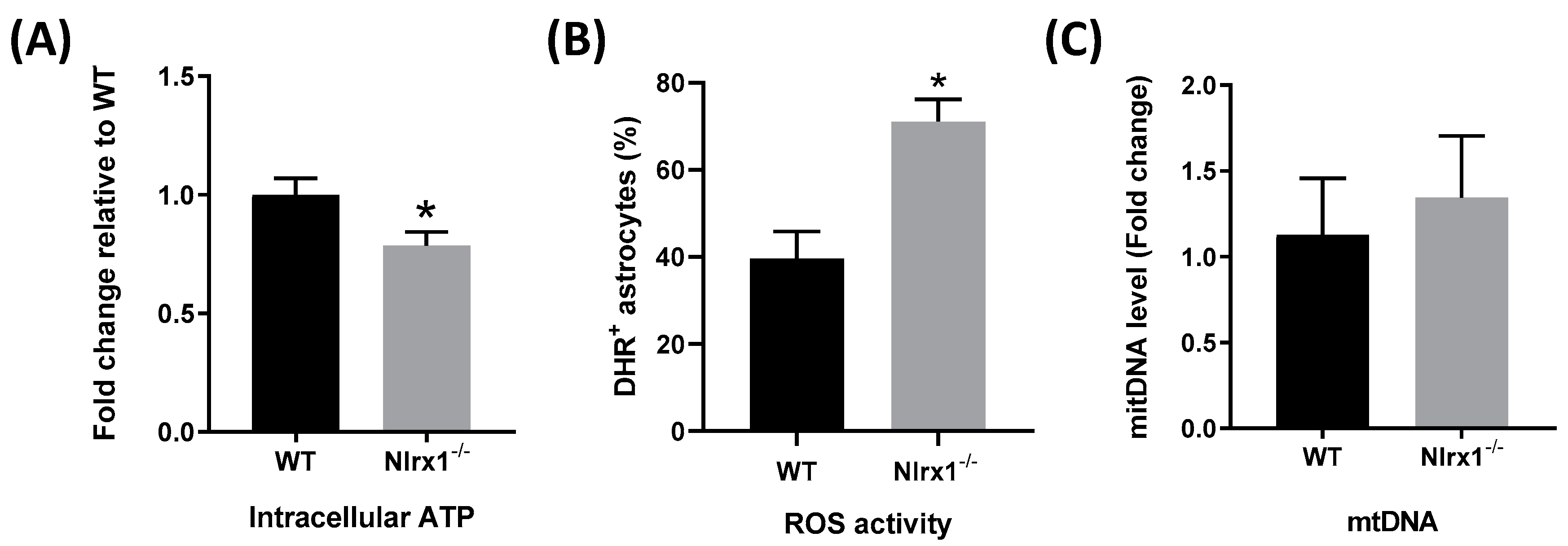
The optimal function of glutamate transporters requires an enormous amount of energy [12,13]. Accordingly, we measured levels of intracellular ATP in both WT and Nlrx1-/- astrocytes. We found that Nlrx1-/- astrocytes have significantly less (20%) intracellular ATP compared to WT (Figure 5A). Since oxidative stress exerts a negative effect on the functional activity of the transporters [66,67], we measured ROS activity in both WT and Nlrx1-/- astrocytes by flow cytometry. Our results demonstrated that Nlrx1-/- astrocytes have significantly higher oxidative activity than WT (Figure 5B). Since mitochondria are the major source of intracellular ATP and ROS in the cells, we evaluated the number of mitochondria in WT and Nlrx1-/- astrocytes. No significant difference was detected in the amount of mtDNA between WT and Nlrx1-/- astrocytes (Figure 5C).
4. Discussion
In this study, we report that the anti-inflammatory [40,41,42,43,44,45,47,48] and prosurvival molecule [49], NLRX1, helps maintain glutamate homeostasis in the CNS. Our findings suggest that NLRX1 enhances astroglial glutamate uptake by promoting the functional activity of glutamate transporters, and inhibits glutamate release from astrocytes by suppressing Ca2+-mediated glutamate exocytosis. To our knowledge, this is the first time that one protein has been shown to be implicated in both processes that regulate glutamate homeostasis.
Given that GLT-1 and GLAST are responsible for the uptake of more than 90% of the extracellular glutamate in the CNS [68,69], we measured the mRNA and protein expression of these glutamate transporters in astrocytes from WT and Nlrx1-/- mice. Unexpectedly, the mRNA expression of both transporters was significantly higher in Nlrx1-/- astrocytes than WT. However, when we measured the total protein expression, as well as the cell surface expression of both transporters, we did not see any significant difference between astrocytes of both genotypes. These findings suggest that the enhanced glutamate uptake in WT astrocytes cannot be attributed to changes in the transcription or translation of the glutamate transporters. In line with our findings, Conrad and Stoffel reported that the direct phosphorylation of GLAST protein by protein kinase C (PKC) reduces its glutamate uptake activity, while immunofluorescence does not show any effect on its protein expression [70]. Another study revealed that arachidonic acid (AA) downregulates glutamate uptake by EAAT-1 by decreasing its affinity to glutamate and the maximal transport rate approximately 30% with no effect on the expression of its protein [71]. In a third study, Trotti et al. reported that oxidative stress by H2O2 induces direct oxidation of the sulfhydryl (SH) group of both transporters, which decreases their glutamate uptake with no protein degradation or reduction in their surface expression [72].
Since NLRX1 is localized in the mitochondria, we hypothesized that NLRX1 enhances the glutamate uptake activity of both GLT-1 and GLAST by improving mitochondrial functions in astrocytes. The level of intracellular ATP in astrocytes is one of the crucial factors that determine the functional activity of glutamate transporters [12,13]. Many previous studies reported that ATP depletion, as in cases of brain ischemia, induces glutamate uptake failure caused by loss or reversal of the transporters’ function [73,74]. To pay for its own energy consumption, a large portion of glutamate metabolites in astrocytes is consumed in the tricarboxylic acid (TCA) cycle to produce more ATP [75]. Interestingly, we found that Nlrx1-/- astrocytes contain 20% less ATP that WT astrocytes, which could either be a cause and/or a result of the reduced glutamate uptake by these cells. Nevertheless, these data, in agreement with the previous studies [50,51], suggest that NLRX1 enhances mitochondrial ATP production.
Moreover, it was reported that the intracellular oxidative stress induced by H2O2 reduces the functional activity of glutamate transporters [66,67]. In this regard, we measured the level of ROS activity in both WT and Nlrx1-/- astrocytes. Our results demonstrated that Nlrx1-/- astrocytes have 50% more oxidative activity than WT, which could be responsible for the significant deficiency of their glutamate uptake. Furthermore, since there was no difference in the level of mtDNA between WT and Nlrx1-/- astrocytes, we excluded the possibility that NLRX1 increases the number of mitochondria.
Taken together, our results suggest that NLRX1 enhances mitochondrial functions in astrocytes, and thus boosts the functional activity of both GLT-1 and GLAST, rather than their protein expression. This effect of NLRX1 on the transporters is achieved by suppressing oxidative stress and, partially, by maintaining sufficient ATP production.
Recent studies suggest that astrocytes express components necessary for the Ca2+-mediated exocytosis, which is the principal mechanism of astroglial glutamate release under physiological conditions [4,17,18,19]. We evaluated whether the excess glutamate release from Nlrx1-/- astrocytes is Ca2+-dependent. First, to exclude the role of extracellular Ca2+, we incubated astrocytes in a Ca2+-free instead of the Ca2+-containing medium. We observed that WT astrocytes significantly upregulated their glutamate release after Ca2+ removal, which agrees with the previously published report by Kostic et al. (2017) [76]. The removal of Ca2+ from the medium stimulates Ca2+ release from the intracellular Ca2+ stores and results in augmentation of glutamate release from astrocytes [76]. However, this does not explain the phenotype in Nlrx1-/- astrocytes, as there was no change between their glutamate release in the Ca2+-containing and the Ca2+-free media, probably because they had already reached their maximum capacity of glutamate release, and Ca2+ removal does not result in any additional effect. In the second step, we evaluated whether this glutamate release occurs in response to Ca2+ release from the intracellular Ca2+ stores. We found that inhibiting Ca2+ release from the mitochondria does not exhibit any significant effect while inhibiting Ca2+ release from the ER reduces the excess glutamate release from Nlrx1-/- astrocytes. These observations corroborate previous findings that ER Ca2+ plays an essential role in glutamate release [4,17].
Consequently, Nlrx1-/- astrocytes show higher mRNA expression of exocytosis proteins downstream of Ca2+ release from the ER, including the Ca2+ sensor, synaptotagmin 11, and the vesicular fusion proteins (VAMP2 and VAMP3). In contrast, there is no significant change in the expression of the cell membrane fusion proteins (Syntaxin 1 a and SNAP23), or the proteins preceding Ca2+ release from the ER (V-ATPase d2, VGLUT1, and VGLUT2). Therefore, our results suggest that the excess glutamate release from Nlrx1-/- astrocytes is mediated by excess Ca2+ release from the ER, followed by an augmentation in the expression of the molecules of exocytosis as a result of the excess Ca2+ release. Collectively, these data provide evidence that NLRX1 mediates its inhibitory effect on glutamate release from astrocytes mainly by suppressing Ca2+ release from the ER, which consequently suppresses glutamate exocytosis.
The connection between the two mechanisms by which NLRX1 mediates its effects on astroglial glutamate uptake and release is still unclear. Being situated in the mitochondria, it is plausible that NLRX1 modifies mitochondrial functions [44,49,50,51]. However, the mechanism by which NLRX1 modifies the function of the ER requires more in-depth investigations.
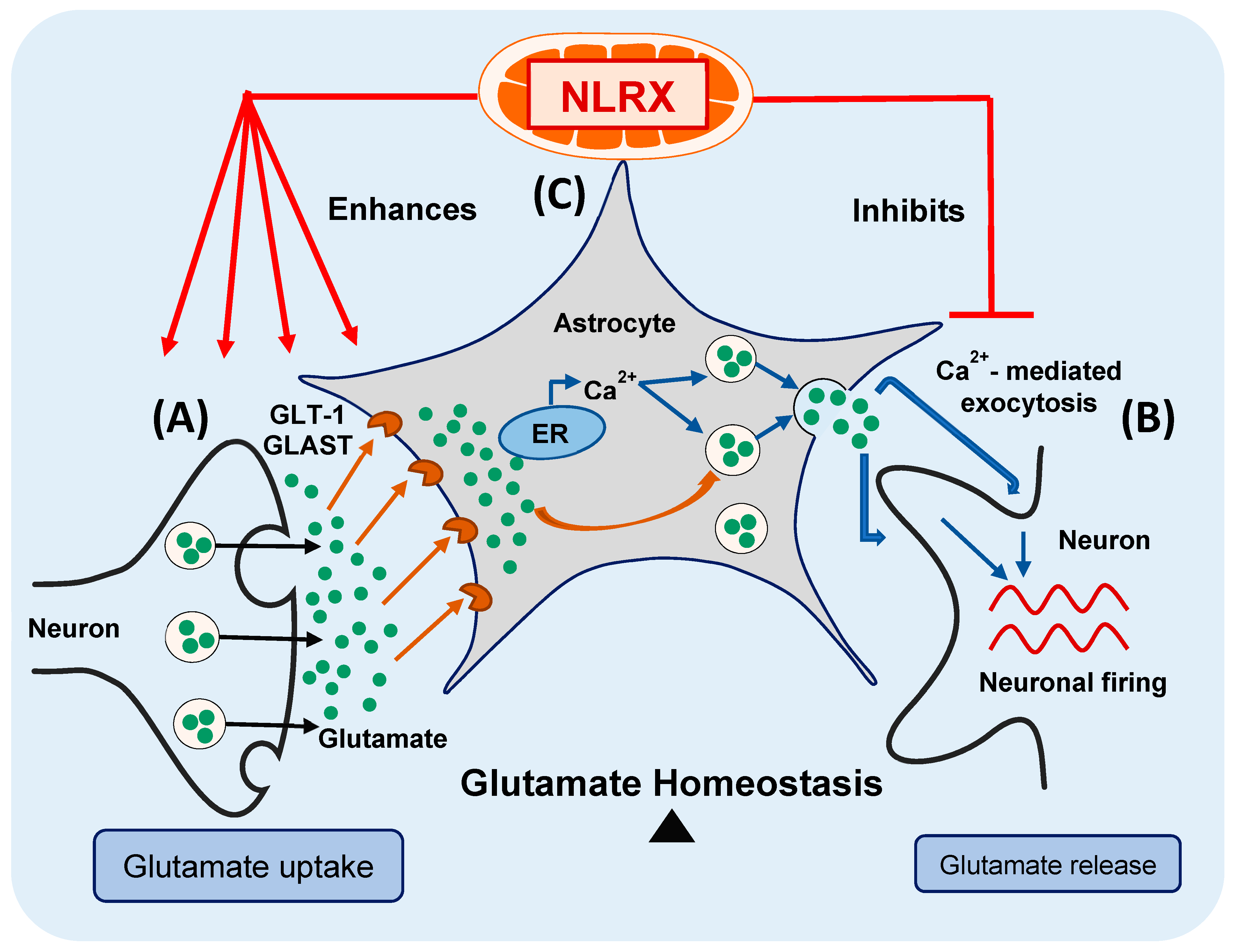
In conclusion, in the current study, we provide evidence that NLRX1 enhances astroglial glutamate uptake and inhibits excess glutamate release from astrocytes, thus maintaining glutamate homeostasis in the CNS (Figure 6—modified from the graphical abstract of our recent publication) [4]. Consequently, NLRX1 represents a potential therapeutic target for the inflammatory and neurodegenerative diseases associated with glutamate excitotoxicity in the CNS.
Author Contributions
S.M., A.A., and D.G. designed the study and the experiments. S.M. performed the experiments and the statistical analysis and wrote the manuscript. M.G. and C.S. as second coauthors helped with animal care, genotyping, and culture preparation. M.G. participated in the flow cytometry experiments and their analysis. C.S. performed the ROS experiment by flow cytometry and its analysis. C.S., A.A., and D.G. contributed to the conceptual reading and critical editing of the manuscript. All authors read and approved the manuscript.
Funding
This study was funded by grants from Association de la sclérose en plaques de l’Estrie (ASPE) and Fonds de la recherche en santé du Québec (FRQS) to S.M.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CNS | Central nervous system |
| EAAT1 and EAAT2 | Excitatory amino acid transporters 1 and 2 |
| GLAST | Glutamate–aspartate transporter |
| GLT-1 | Glutamate transporter-1 |
| ATP | Adenosine triphosphate |
| VGLUT1 and VGLUT2 | Vesicular glutamate transporters 1 and 2 |
| V-ATPase | Vacuolar (H+) ATPase |
| VAMP2 and VAMP3 | Vesicle-associated membrane protein 2 and 3 |
| ER | Endoplasmic reticulum |
| SNAP23 | Soluble N-ethylmaleimide-sensitive factor attachment protein 23 |
| HIV | Human immunodeficiency virus |
| MS | Multiple sclerosis |
| ALS | Amyotrophic lateral sclerosis |
| AD | Alzheimer’s disease |
| PD | Parkinson’s disease |
| NLRs | NOD-like receptors |
| NF-κB | Nuclear factor-κB |
| NLRX1 | NOD-like receptor X1 |
| IBD | Inflammatory bowel disease |
| EAE | Experimental autoimmune encephalomyelitis |
| WT | Wild-type |
| Nlrx1-/- | Nlrx1 knockout |
| dFBS | Deactivated fetal bovine serum |
| DMSO | Dimethyl sulfoxide |
| HBSS | Hank’s Balanced Salt Solution |
| qPCR | Quantitative real-time PCR |
| PBS | Phosphate-buffered saline |
| ROS | Reactive oxygen species |
| DHR | Dihydrorhodamine 123 |
| mtDNA | Mitochondrial DNA |
| SEM | Standard error of the mean |
| 2-APB | 2-Aminoethyl diphenylborinate |
| IP3 | Inositol-1,4,5-trisphosphate |
| CsA | Cyclosporin A |
| PKC | Protein kinase C |
| AA | Arachidonic acid |
| SH group | Sulfhydryl group |
| TCA | Tricarboxylic acid |
References
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Curtis, D.; Johnston, G. Amino acid transmitters in the mammalian central nervous system. In Ergebnisse der Physiologie Reviews of Physiology; Springer: Berlin/Heidelberg, Germany, 1974; Volume 69, pp. 97–188. [Google Scholar] [CrossRef]
- Fonnum, F. Glutamate: A neurotransmitter in mammalian brain. J. Neurochem. 1984, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D.; Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Swanson, R.A. Astrocyte glutamate transport: Review of properties, regulation, and physiological functions. Glia 2000, 32, 1–14. [Google Scholar] [CrossRef]
- Shashidharan, P.; Plaitakis, A. Cloning and characterization of a glutamate transporter cDNA from human cerebellum. Biochim. Biophys. Acta 1993, 1216, 161–164. [Google Scholar] [CrossRef]
- Shashidharan, P.; Wittenberg, I.; Plaitakis, A. Molecular cloning of human brain glutamate/aspartate transporter II. Biochim. Biophys. Acta-Biomembr. 1994, 1191, 393–396. [Google Scholar] [CrossRef]
- Storck, T.; Schulte, S.; Hofmann, K.; Stoffel, W. Structure, expression, and functional analysis of a Na(+)-dependent glutamate/aspartate transporter from rat brain. Proc. Natl. Acad. Sci. USA 1992, 89, 10955–10959. [Google Scholar] [CrossRef]
- Pines, G.; Danbolt, N.C.; Bjørås, M.; Zhang, Y.; Bendahan, A.; Eide, L.; Koepsell, H.; Storm-Mathisen, J.; Seeberg, E.; Kanner, B.I. Cloning and expression of a rat brain l-glutamate transporter. Nature 1992, 360, 464–467. [Google Scholar] [CrossRef]
- Kojima, S.; Nakamura, T.; Nidaira, T.; Nakamura, K.; Ooashi, N.; Ito, E.; Watase, K.; Tanaka, K.; Wada, K.; Kudo, Y.; et al. Optical detection of synaptically induced glutamate transport in hippocampal slices. J. Neurosci. 1999, 19, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Bergles, D.E.; Jahr, C.E. Glial contribution to glutamate uptake at Schaffer collateral-commissural synapses in the hippocampus. J. Neurosci. 1998, 18, 7709–7716. [Google Scholar] [CrossRef]
- Sibson, N.R.; Dhankhar, A.; Mason, G.F.; Rothman, D.L.; Behar, K.L.; Shulman, R.G. Stoichiometric coupling of brain glucose metabolism and glutamatergic neuronal activity. Proc. Natl. Acad. Sci. USA 1998, 95, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake stimulates Na+,K+-ATPase activity in astrocytes via activation of a distinct subunit highly sensitive to ouabain. J. Neurochem. 1997, 69, 2132–2137. [Google Scholar] [CrossRef] [PubMed]
- Gegelashvili, G.; Schousboe, A. High affinity glutamate transporters: Regulation of expression and activity. Mol. Pharmacol. 1997, 52, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Chi-Castañeda, D.; Suárez-Pozos, E.; Ortega, A. Regulation of Glutamate Transporter Expression in Glial Cells. Adv. Neurobiol. 2017, 16, 199–224. [Google Scholar]
- Sattler, R.; Rothstein, J.D. Regulation and dysregulation of glutamate transporters. Handb. Exp. Pharmacol. 2006, 277–303. [Google Scholar]
- Hamilton, N.B.; Attwell, D. Do astrocytes really exocytose neurotransmitters? Nat. Rev. Neurosci. 2010, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Parpura, V.; Haydon, P.G. Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 8629–8634. [Google Scholar] [CrossRef] [Green Version]
- Malarkey, E.B.; Parpura, V. Mechanisms of glutamate release from astrocytes. Neurochem. Int. 2008, 52, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Bezzi, P.; Gundersen, V.; Galbete, J.L.; Seifert, G.; Steinhäuser, C.; Pilati, E.; Volterra, A. Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate. Nat. Neurosci. 2004, 7, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, L.; Zhou, Y.; Zheng, L.-H.; Zhou, Z. “Kiss-and-Run” Glutamate Secretion in Cultured and Freshly Isolated Rat Hippocampal Astrocytes. J. Neurosci. 2005, 25, 9236–9243. [Google Scholar] [CrossRef]
- Araque, A.; Li, N.; Doyle, R.T.; Haydon, P.G. SNARE protein-dependent glutamate release from astrocytes. J. Neurosci. 2000, 20, 666–673. [Google Scholar] [CrossRef]
- Parpura, V.; Zorec, R. Gliotransmission: Exocytotic release from astrocytes. Brain Res. Rev. 2010, 63, 83–92. [Google Scholar] [CrossRef]
- Pasti, L.; Zonta, M.; Pozzan, T.; Vicini, S.; Carmignoto, G. Cytosolic calcium oscillations in astrocytes may regulate exocytotic release of glutamate. J. Neurosci. 2001, 21, 477–484. [Google Scholar] [CrossRef]
- Zhang, Q.; Fukuda, M.; Van Bockstaele, E.; Pascual, O.; Haydon, P.G. Synaptotagmin IV regulates glial glutamate release. Proc. Natl. Acad. Sci. USA 2004, 101, 9441–9446. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-T.; Lu, J.-C.; Bai, J.; Chang, P.Y.; Martin, T.F.J.; Chapman, E.R.; Jackson, M.B. Different domains of synaptotagmin control the choice between kiss-and-run and full fusion. Nature 2003, 424, 943–947. [Google Scholar] [CrossRef]
- Sugita, S.; Han, W.; Butz, S.; Liu, X.; Fernández-Chacón, R.; Lao, Y.; Südhof, T.C. Synaptotagmin VII as a plasma membrane Ca(2+) sensor in exocytosis. Neuron 2001, 30, 459–473. [Google Scholar] [CrossRef]
- Montana, V.; Ni, Y.; Sunjara, V.; Hua, X.; Parpura, V. Vesicular Glutamate Transporter-Dependent Glutamate Release from Astrocytes. J. Neurosci. 2004, 24, 2633–2642. [Google Scholar] [CrossRef] [Green Version]
- Parpura, V.; Liu, F.; Brethorst, S.; Jeftinija, K.; Jeftinija, S.; Haydon, P.G. Alpha-latrotoxin stimulates glutamate release from cortical astrocytes in cell culture. FEBS Lett. 1995, 360, 266–270. [Google Scholar]
- Hepp, R.; Perraut, M.; Chasserot-Golaz, S.; Galli, T.; Aunis, D.; Langley, K.; Grant, N.J. Cultured glial cells express the SNAP-25 analogue SNAP-23. Glia 1999, 27, 181–187. [Google Scholar] [CrossRef]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Van Landeghem, F.K.H.; Weiss, T.; Oehmichen, M.; Deimling, A. Von Decreased Expression of Glutamate Transporters in Astrocytes after Human Traumatic Brain Injury. J. Neurotrauma 2006, 23, 1518–1528. [Google Scholar] [CrossRef]
- Vesce, S.; Bezzi, P.; Rossi, D.; Meldolesi, J.; Volterra, A. HIV-1 gp120 glycoprotein affects the astrocyte control of extracellular glutamate by both inhibiting the uptake and stimulating the release of the amino acid. FEBS Lett. 1997, 411, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, A.L.; Robinson, M.B. The role of glutamate transporters in neurodegenerative diseases and potential opportunities for intervention. Neurochem. Int. 2007, 51, 333–355. [Google Scholar] [CrossRef] [Green Version]
- Pitt, D.; Werner, P.; Raine, C.S. Glutamate excitotoxicity in a model of multiple sclerosis. Nat. Med. 2000, 6, 67–70. [Google Scholar] [CrossRef]
- Bezzi, P.; Domercq, M.; Brambilla, L.; Galli, R.; Schols, D.; De Clercq, E.; Vescovi, A.; Bagetta, G.; Kollias, G.; Meldolesi, J.; et al. CXCR4-activated astrocyte glutamate release via TNFα: Amplification by microglia triggers neurotoxicity. Nat. Neurosci. 2001, 4, 702–710. [Google Scholar] [CrossRef]
- Bezzi, P.; Carmignoto, G.; Pasti, L.; Vesce, S.; Rossi, D.; Rizzini, B.L.; Pozzan, T.; Volterra, A. Prostaglandins stimulate calcium-dependent glutamate release in astrocytes. Nature 1998, 391, 281–285. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Z. Molecular and cellular mechanisms of excitotoxic neuronal death. Apoptosis 2010, 15, 1382–1402. [Google Scholar] [CrossRef]
- Gharagozloo, M.; Gris, K.V.; Mahvelati, T.; Amrani, A.; Lukens, J.R.; Gris, D. NLR-Dependent Regulation of Inflammation in Multiple Sclerosis. Front. Immunol. 2018, 8, 2012. [Google Scholar] [CrossRef]
- Moore, C.B.; Bergstralh, D.T.; Duncan, J.A.; Lei, Y.; Morrison, T.E.; Zimmermann, A.G.; Accavitti-Loper, M.A.; Madden, V.J.; Sun, L.; Ye, Z.; et al. NLRX1 is a regulator of mitochondrial antiviral immunity. Nature 2008, 451, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Allen, I.C.; Moore, C.B.; Schneider, M.; Lei, Y.; Davis, B.K.; Scull, M.A.; Gris, D.; Roney, K.E.; Zimmermann, A.G.; Bowzard, J.B.; et al. NLRX1 protein attenuates inflammatory responses to infection by interfering with the RIG-I-MAVS and TRAF6-NF-κB signaling pathways. Immunity 2011, 34, 854–865. [Google Scholar] [CrossRef]
- Huang, J.-H.; Liu, C.-Y.; Wu, S.-Y.; Chen, W.-Y.; Chang, T.-H.; Kan, H.-W.; Hsieh, S.-T.; Ting, J.P.-Y.; Wu-Hsieh, B.A. NLRX1 Facilitates Histoplasma capsulatum-Induced LC3-Associated Phagocytosis for Cytokine Production in Macrophages. Front. Immunol. 2018, 9, 2761. [Google Scholar] [CrossRef]
- Aikawa, C.; Nakajima, S.; Karimine, M.; Nozawa, T.; Minowa-Nozawa, A.; Toh, H.; Yamada, S.; Nakagawa, I. NLRX1 Negatively Regulates Group A Streptococcus Invasion and Autophagy Induction by Interacting With the Beclin 1–UVRAG Complex. Front. Cell. Infect. Microbiol. 2018, 8, 403. [Google Scholar] [CrossRef]
- Ma, D.; Zhao, Y.; She, J.; Zhu, Y.; Zhao, Y.; Liu, L.; Zhang, Y. NLRX1 alleviates lipopolysaccharide-induced apoptosis and inflammation in chondrocytes by suppressing the activation of NF-κB signaling. Int. Immunopharmacol. 2019, 71, 7–13. [Google Scholar] [CrossRef]
- Tattoli, I.; Killackey, S.A.; Foerster, E.G.; Molinaro, R.; Maisonneuve, C.; Rahman, M.A.; Winer, S.; Winer, D.A.; Streutker, C.J.; Philpott, D.J.; et al. NLRX1 Acts as an Epithelial-Intrinsic Tumor Suppressor through the Modulation of TNF-Mediated Proliferation. Cell Rep. 2016, 14, 2576–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leber, A.; Hontecillas, R.; Tubau-Juni, N.; Zoccoli-Rodriguez, V.; Abedi, V.; Bassaganya-Riera, J. NLRX1 Modulates Immunometabolic Mechanisms Controlling the Host-Gut Microbiota Interactions during Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Theus, M.H.; Brickler, T.; Meza, A.L.; Coutermarsh-Ott, S.; Hazy, A.; Gris, D.; Allen, I.C. Loss of NLRX1 Exacerbates Neural Tissue Damage and NF-κB Signaling following Brain Injury. J. Immunol. 2017, 199, 3547–3558. [Google Scholar] [CrossRef]
- Eitas, T.K.; Chou, W.-C.; Wen, H.; Gris, D.; Robbins, G.R.; Brickey, J.; Oyama, Y.; Ting, J.P.-Y. The Nucleotide-binding Leucine-rich Repeat (NLR) Family Member NLRX1 Mediates Protection against Experimental Autoimmune Encephalomyelitis and Represses Macrophage/Microglia-induced Inflammation. J. Biol. Chem. 2014, 289, 4173–4179. [Google Scholar] [CrossRef]
- Imbeault, E.; Mahvelati, T.M.; Braun, R.; Gris, P.; Gris, D. Nlrx1 regulates neuronal cell death. Mol. Brain 2014, 7, 90. [Google Scholar] [CrossRef]
- Stokman, G.; Kors, L.; Bakker, P.J.; Rampanelli, E.; Claessen, N.; Teske, G.J.D.; Butter, L.; van Andel, H.; van den Bergh Weerman, M.A.; Larsen, P.W.B.; et al. NLRX1 dampens oxidative stress and apoptosis in tissue injury via control of mitochondrial activity. J. Exp. Med. 2017, 214, 2405–2420. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Roy, M.; Prajapati, P.; Lipatova, A.; Sripada, L.; Gohel, D.; Singh, A.; Mane, M.; Godbole, M.M.; Chumakov, P.M.; et al. NLRX1 regulates TNF-α-induced mitochondria-lysosomal crosstalk to maintain the invasive and metastatic potential of breast cancer cells. Biochim. Biophys. Acta-Mol. Basis Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Killackey, S.A.; Rahman, M.A.; Soares, F.; Zhang, A.B.; Abdel-Nour, M.; Philpott, D.J.; Girardin, S.E. The mitochondrial Nod-like receptor NLRX1 modifies apoptosis through SARM1. Mol. Cell. Biochem. 2019, 453, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Soares, F.; Tattoli, I.; Rahman, M.A.; Robertson, S.J.; Belcheva, A.; Liu, D.; Streutker, C.; Winer, S.; Winer, D.A.; Martin, A.; et al. The mitochondrial protein NLRX1 controls the balance between extrinsic and intrinsic apoptosis. J. Biol. Chem. 2014, 289, 19317–19330. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, J.; Coulombe, F.; Downey, J.; Tzelepis, F.; Shalaby, K.; Tattoli, I.; Berube, J.; Rousseau, S.; Martin, J.G.; Girardin, S.E.; et al. NLRX1 prevents mitochondrial induced apoptosis and enhances macrophage antiviral immunity by interacting with influenza virus PB1-F2 protein. Proc. Natl. Acad. Sci. USA 2014, 111, E2110–E2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharagozloo, M.; Mahvelati, T.M.; Imbeault, E.; Gris, P.; Zerif, E.; Bobbala, D.; Ilangumaran, S.; Amrani, A.; Gris, D. The nod-like receptor, Nlrp12, plays an anti-inflammatory role in experimental autoimmune encephalomyelitis. J. Neuroinflammation 2015, 12, 198. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.; Ranaivo, H.R.; Rusie, A.; Wadhwani, N.; Koh, S.; Wainwright, M.S. Thrombin decreases expression of the glutamate transporter GLAST and inhibits glutamate uptake in primary cortical astrocytes via the Rho kinase pathway. Exp. Neurol. 2015, 273, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, J.M.; Smith, S.H.; Bilbo, S.D. FACS analysis of neuronal-glial interactions in the nucleus accumbens following morphine administration. Psychopharmacology 2013, 230, 525–535. [Google Scholar] [CrossRef]
- Gharagozloo, M.; Mahmoud, S.; Simard, C.; Mahvelati, T.M.; Amrani, A.; Gris, D. The Dual Immunoregulatory function of Nlrp12 in T Cell-Mediated Immune Response: Lessons from Experimental Autoimmune Encephalomyelitis. Cells 2018, 7, 119. [Google Scholar] [CrossRef]
- Gris, D.; Hamilton, E.F.; Weaver, L.C. The systemic inflammatory response after spinal cord injury damages lungs and kidneys. Exp. Neurol. 2008, 211, 259–270. [Google Scholar] [CrossRef]
- Farrell, S.M.J.; Groeger, G.; Bhatt, L.; Finnegan, S.; O’Brien, C.J.; Cotter, T.G. bFGF-mediated redox activation of the PI3K/Akt pathway in retinal photoreceptor cells. Eur. J. Neurosci. 2011, 33, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Marcaida, G.; Miñana, M.D.; Grisolía, S.; Felipo, V. Determination of intracellular ATP in primary cultures of neurons. Brain Res. Brain Res. Protoc. 1997, 1, 75–78. [Google Scholar] [CrossRef]
- Fuke, S.; Kubota-Sakashita, M.; Kasahara, T.; Shigeyoshi, Y.; Kato, T. Regional variation in mitochondrial DNA copy number in mouse brain. Biochim. Biophys. Acta-Bioenerg. 2011, 1807, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Kanaji, T.; Nakade, S.; Kanno, T.; Mikoshiba, K. 2APB, 2-aminoethoxydiphenyl borate, a membrane-penetrable modulator of Ins(1,4,5)P3-induced Ca2+ release. J. Biochem. 1997, 122, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Reyes, R.C.; Parpura, V. Mitochondria modulate Ca2+-dependent glutamate release from rat cortical astrocytes. J. Neurosci. 2008, 28, 9682–9691. [Google Scholar] [CrossRef]
- Volterra, A.; Trotti, D.; Floridi, S.; Racagni, G. Reactive oxygen species inhibit high-affinity glutamate uptake: Molecular mechanism and neuropathological implications. Ann. N. Y. Acad. Sci. 1994, 738, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Sorg, O.; Horn, T.F.; Yu, N.; Gruol, D.L.; Bloom, F.E. Inhibition of astrocyte glutamate uptake by reactive oxygen species: Role of antioxidant enzymes. Mol. Med. 1997, 3, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Lehre, K.P.; Danbolt, N.C. The number of glutamate transporter subtype molecules at glutamatergic synapses: Chemical and stereological quantification in young adult rat brain. J. Neurosci. 1998, 18, 8751–8757. [Google Scholar] [CrossRef] [PubMed]
- Eulenburg, V.; Gomeza, J. Neurotransmitter transporters expressed in glial cells as regulators of synapse function. Brain Res. Rev. 2010, 63, 103–112. [Google Scholar] [CrossRef]
- Conradt, M.; Stoffel, W. Inhibition of the high-affinity brain glutamate transporter GLAST-1 via direct phosphorylation. J. Neurochem. 1997, 68, 1244–1251. [Google Scholar] [CrossRef]
- Zerangue, N.; Arriza, J.L.; Amara, S.G.; Kavanaugh, M.P. Differential modulation of human glutamate transporter subtypes by arachidonic acid. J. Biol. Chem. 1995, 270, 6433–6435. [Google Scholar] [CrossRef]
- Trotti, D.; Rizzini, B.L.; Rossi, D.; Haugeto, O.; Racagni, G.; Danbolt, N.C.; Volterra, A. Neuronal and glial glutamate transporters possess an SH-based redox regulatory mechanism. Eur. J. Neurosci. 1997, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Grewer, C.; Gameiro, A.; Zhang, Z.; Tao, Z.; Braams, S.; Rauen, T. Glutamate forward and reverse transport: From molecular mechanism to transporter-mediated release after ischemia. IUBMB Life 2008, 60, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, D.J.; Oshima, T.; Attwell, D. Glutamate release in severe brain ischaemia is mainly by reversed uptake. Nature 2000, 403, 316–321. [Google Scholar] [CrossRef]
- McKenna, M.C. Glutamate Pays Its Own Way in Astrocytes. Front. Endocrinol. 2013, 4, 191. [Google Scholar] [CrossRef]
- Kostic, M.; Zivkovic, N.; Cvetanovic, A.; Stojanovic, I.; Colic, M. IL-17 signalling in astrocytes promotes glutamate excitotoxicity: Indications for the link between inflammatory and neurodegenerative events in multiple sclerosis. Mult. Scler. Relat. Disord. 2017, 11, 12–17. [Google Scholar] [CrossRef]
Figure 1.
NLRX1 inhibits glutamate release and enhances glutamate uptake by astrocytes. (A) WT and Nlrx1-/- astrocyte cultures were incubated in glutamate-free, Ca2+-containing HBSS medium. After 4 h, the culture supernatant was collected, and glutamate release in the medium was measured (n = 7), ** p < 0.01 as determined by Mann–Whitney test. (B) For evaluation of glutamate uptake, astrocyte cultures were incubated with 100 or 200 µM glutamate in the Ca2+-containing HBSS medium (n = 5), * p < 0.05 as determined by Tukey’s test, results are presented as mean ± SEM.
Figure 1.
NLRX1 inhibits glutamate release and enhances glutamate uptake by astrocytes. (A) WT and Nlrx1-/- astrocyte cultures were incubated in glutamate-free, Ca2+-containing HBSS medium. After 4 h, the culture supernatant was collected, and glutamate release in the medium was measured (n = 7), ** p < 0.01 as determined by Mann–Whitney test. (B) For evaluation of glutamate uptake, astrocyte cultures were incubated with 100 or 200 µM glutamate in the Ca2+-containing HBSS medium (n = 5), * p < 0.05 as determined by Tukey’s test, results are presented as mean ± SEM.

Figure 2.
Excess Ca2+ release from the ER mediates glutamate release from Nlrx1-/- astrocytes. (A) WT and Nlrx1-/- astrocytes were incubated in a Ca2+-containing HBSS buffer or Ca2+-free Lock’s solution for 1 h (n = 7); (B) astrocyte cultures were incubated with 0.01 or 0.05 µM of 2-APB (n = 5), or (C) with 2 or 10 µM of CsA (n = 6) in the Ca2+-free Lock’s solution for 1 h. The supernatant was collected from all cultures and glutamate in the medium was measured by the glutamate assay kit. * p < 0.05 as determined by Tukey’s test, results are presented as mean ± SEM.
Figure 2.
Excess Ca2+ release from the ER mediates glutamate release from Nlrx1-/- astrocytes. (A) WT and Nlrx1-/- astrocytes were incubated in a Ca2+-containing HBSS buffer or Ca2+-free Lock’s solution for 1 h (n = 7); (B) astrocyte cultures were incubated with 0.01 or 0.05 µM of 2-APB (n = 5), or (C) with 2 or 10 µM of CsA (n = 6) in the Ca2+-free Lock’s solution for 1 h. The supernatant was collected from all cultures and glutamate in the medium was measured by the glutamate assay kit. * p < 0.05 as determined by Tukey’s test, results are presented as mean ± SEM.

Figure 3.
Increased mRNA expression of the proteins of exocytosis in Nlrx1-/- astrocytes. mRNA expression of the proteins of exocytosis in WT and Nlrx1-/- astrocytes shows (A) significant upregulation of mRNA expression of the Ca2+ sensor, synaptotagmin 11, and vesicular fusion proteins, VAMP2 and VAMP3, in Nlrx1-/- astrocytes compared to WT; (B) no significant change in the cell membrane fusion proteins’ mRNA expression; and (C) no significant change in the expression of the proteins upstream of Ca2+ release from the ER. ** p < 0.01 and * p < 0.05 as determined by Mann–Whitney test (n ≥ 5), results are presented as mean ± SEM.
Figure 3.
Increased mRNA expression of the proteins of exocytosis in Nlrx1-/- astrocytes. mRNA expression of the proteins of exocytosis in WT and Nlrx1-/- astrocytes shows (A) significant upregulation of mRNA expression of the Ca2+ sensor, synaptotagmin 11, and vesicular fusion proteins, VAMP2 and VAMP3, in Nlrx1-/- astrocytes compared to WT; (B) no significant change in the cell membrane fusion proteins’ mRNA expression; and (C) no significant change in the expression of the proteins upstream of Ca2+ release from the ER. ** p < 0.01 and * p < 0.05 as determined by Mann–Whitney test (n ≥ 5), results are presented as mean ± SEM.

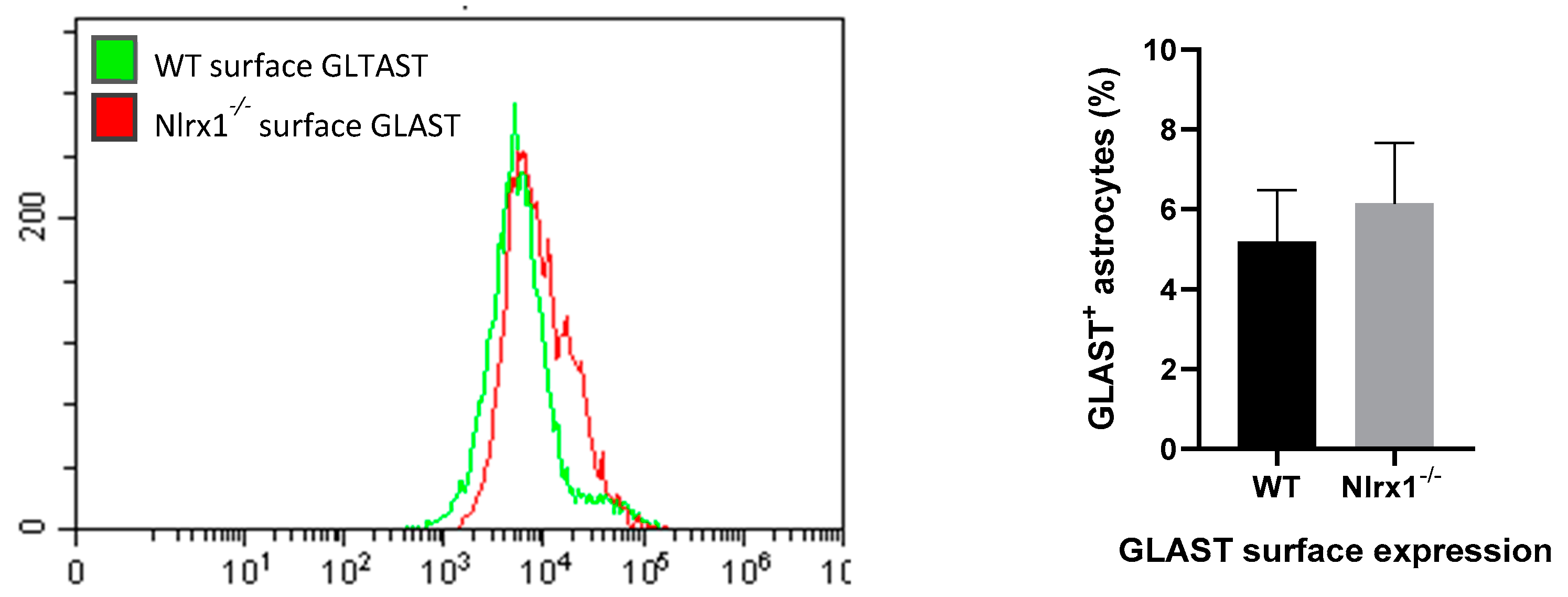
Figure 4.
mRNA and protein expression of GLT-1 and GLAST in astrocytes. (A) mRNA expression of GLT-1 and GLAST is significantly upregulated in Nlrx1-/- astrocytes compared to WT (n = 5). ** p < 0.01 as determined by Mann–Whitney test; (B) the total protein expression of GLT-1 and GLAST proteins in WT and Nlrx1-/- astrocytes was measured by flow cytometry (n = 5); (C) the cell surface expression of both transporters on astrocytes was measured by flow cytometry (n = 7). Representative flow cytometric histograms presented on the left side, p > 0.05 as determined by Mann–Whitney test, results are presented as mean ± SEM.
Figure 4.
mRNA and protein expression of GLT-1 and GLAST in astrocytes. (A) mRNA expression of GLT-1 and GLAST is significantly upregulated in Nlrx1-/- astrocytes compared to WT (n = 5). ** p < 0.01 as determined by Mann–Whitney test; (B) the total protein expression of GLT-1 and GLAST proteins in WT and Nlrx1-/- astrocytes was measured by flow cytometry (n = 5); (C) the cell surface expression of both transporters on astrocytes was measured by flow cytometry (n = 7). Representative flow cytometric histograms presented on the left side, p > 0.05 as determined by Mann–Whitney test, results are presented as mean ± SEM.


Figure 5.
NLRX1 enhances mitochondrial functions in astrocytes. (A) The level of intracellular ATP was measured in WT and Nlrx1-/- astrocytes using an ATP bioluminescent assay kit (n = 5); (B) the level of oxidative activity was measured in WT and Nlrx1-/- astrocytes by flow cytometry (n = 4); (C) the difference between the amount of mtDNA in WT and Nlrx1-/- astrocytes was measured by qPCR (n = 3). * p < 0.05 as determined by Mann–Whitney test, results are presented as mean ± SEM.
Figure 5.
NLRX1 enhances mitochondrial functions in astrocytes. (A) The level of intracellular ATP was measured in WT and Nlrx1-/- astrocytes using an ATP bioluminescent assay kit (n = 5); (B) the level of oxidative activity was measured in WT and Nlrx1-/- astrocytes by flow cytometry (n = 4); (C) the difference between the amount of mtDNA in WT and Nlrx1-/- astrocytes was measured by qPCR (n = 3). * p < 0.05 as determined by Mann–Whitney test, results are presented as mean ± SEM.

Figure 6.
NLRX1 maintains glutamate homeostasis in the CNS. (A) Uptake of the extracellular glutamate by astrocytes is mediated by the glutamate uptake transporters (GLT-1 and GLAST); (B) Ca2+-mediated exocytosis mediates glutamate release from astrocytes, in response to Ca2+ release from the ER. The released glutamate helps to synchronize and reinforce the firing of the surrounding neurons; (C) NLRX1 enhances astroglial glutamate uptake and inhibits its Ca2+-mediated glutamate exocytosis, hence maintaining glutamate homeostasis in the CNS.
Figure 6.
NLRX1 maintains glutamate homeostasis in the CNS. (A) Uptake of the extracellular glutamate by astrocytes is mediated by the glutamate uptake transporters (GLT-1 and GLAST); (B) Ca2+-mediated exocytosis mediates glutamate release from astrocytes, in response to Ca2+ release from the ER. The released glutamate helps to synchronize and reinforce the firing of the surrounding neurons; (C) NLRX1 enhances astroglial glutamate uptake and inhibits its Ca2+-mediated glutamate exocytosis, hence maintaining glutamate homeostasis in the CNS.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer sequences used for qPCR.
| 18S | F: 5′ CGG CTA CCA CAT CCA AGG AA ′3 R: 5′ GCT GGA ATT ACC GCG GCT ′3 |
| Exocytosis | |
| V-ATPase d2 | F: 5′ TTC AGT TGC TAT CCA GGA CTC GGA ′3 R: 5′ GCA TGT CAT GTA GGT GAG AAA TGT GCT CA ′3 |
| VGLUT1 | F: 5′ GGT GGA GGG GGT CAC ATA C ′3 R: 5′ AGA TCC CGA AGC TGC CAT AGA ′3 |
| VGLUT2 | F: 5′ CCC TGG AGG TGC CTG AGA A ′3 R: 5′ GCG GTG GAT AGT GCT GTT GTT ′3 |
| Synaptotagmin 11 | F: 5′ GAC ACT TGC CGA AGA TGG ATA TC ′3 R: 5′ TGC GTT TTC TGC CGT AGT AGA ′3 |
| VAMP2 | F: 5′ CAC AAT CTG GTT CTT TGA GGA G ′3 R: 5′ AGA GAC TTC AGG CAG GAA TTA G ′3 |
| VAMP3 | F: 5′ CTC ACC AAG GCA TCA GTC TG ′3 R: 5′ ATT CTA AGA GCA CCA GGC ATC ′3 |
| Syntaxin 1a | F: 5′ TCC AAG CTA AAG AGC ATT GAG C ′3 R: 5′ GGC GTT GTA CTC GGA CAT GA ′3 |
| SNAP23 | F: 5′ AAT CCT GGG TTT AGC CAT TGA GTC ′3 R: 5′ TTG GTC CAT GCC TTC TTC TAT GC ′3 |
| Glutamate transporters | |
| GLT-1 | F: 5′ CGA TGA GCC AAA GCA CCG AA ′3 R: 5′ CTG GAG ATG ATA AGA GGG AGG ATG ′3 |
| GLAST | F: 5′ TCA AGT TCT GCC ACC CTA CC ′3 R: 5′ TCT GTC CAA AGT TCA GGT CAA ′3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mahmoud, S.; Gharagozloo, M.; Simard, C.; Amrani, A.; Gris, D. NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes. Cells 2019, 8, 400. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050400
AMA Style
Mahmoud S, Gharagozloo M, Simard C, Amrani A, Gris D. NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes. Cells. 2019; 8(5):400. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050400
Chicago/Turabian StyleMahmoud, Shaimaa, Marjan Gharagozloo, Camille Simard, Abdelaziz Amrani, and Denis Gris. 2019. "NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes" Cells 8, no. 5: 400. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050400
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.