NDRG2 Sensitizes Myeloid Leukemia to Arsenic Trioxide via GSK3β–NDRG2–PP2A Complex Formation


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Reagents, and Short Hairpin RNAs (shRNAs)
2.2. Western Blotting
2.3. Co-Immunoprecipitation and Glutathion S-Tranferase Tag(GST) Pull-Down Assay
2.4. PP2Ac Activity Assay
2.5. Lentivirus Production and Infection
2.6. Flow Cytometry
2.7. Statistics
3. Results
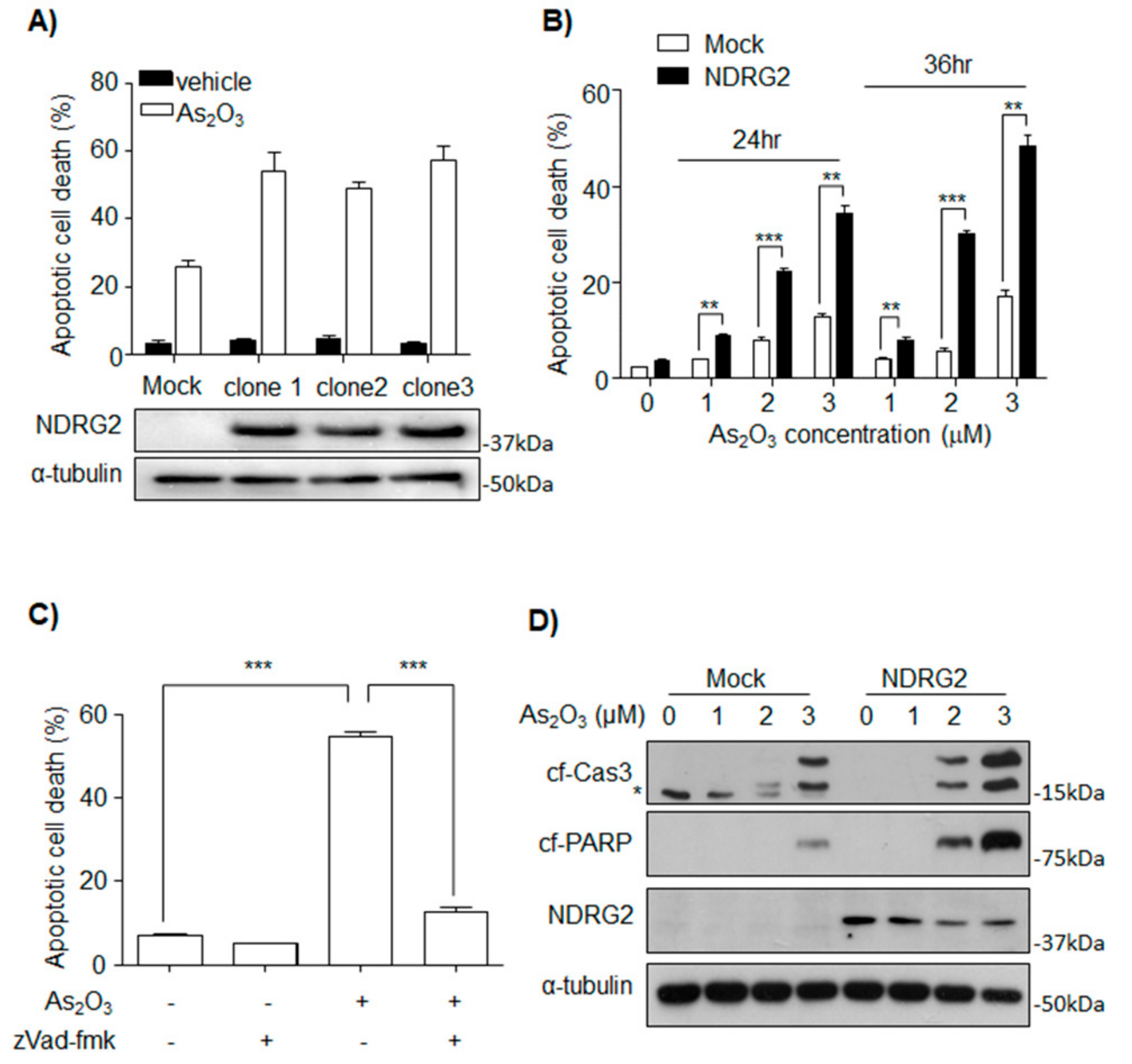
3.1. NDRG2 Expression Sensitizes U937 Cells to As2O3
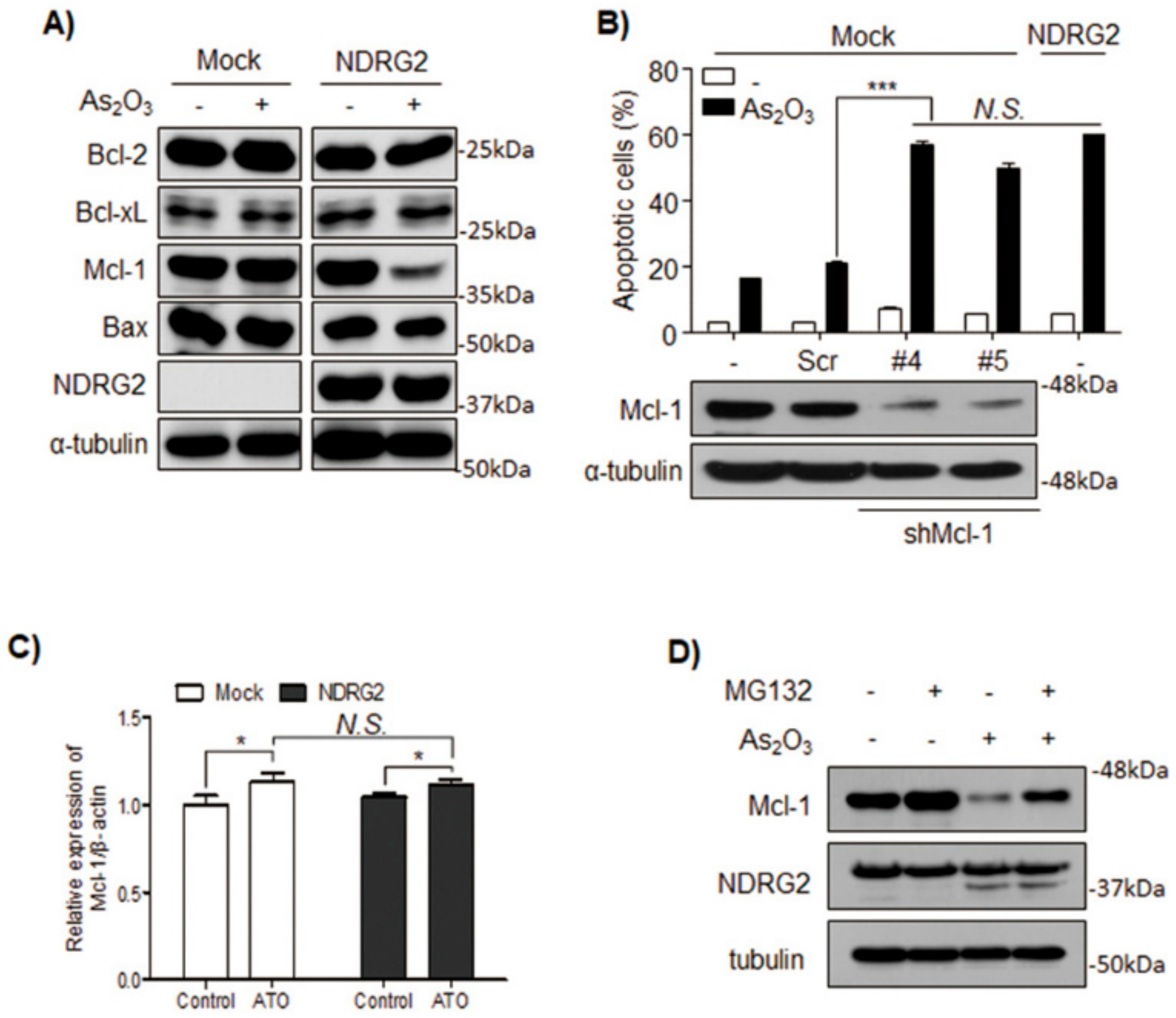
3.2. The Sensitivity of U937-NDRG2 to As2O3 Is Determined by Mcl-1 Degradation
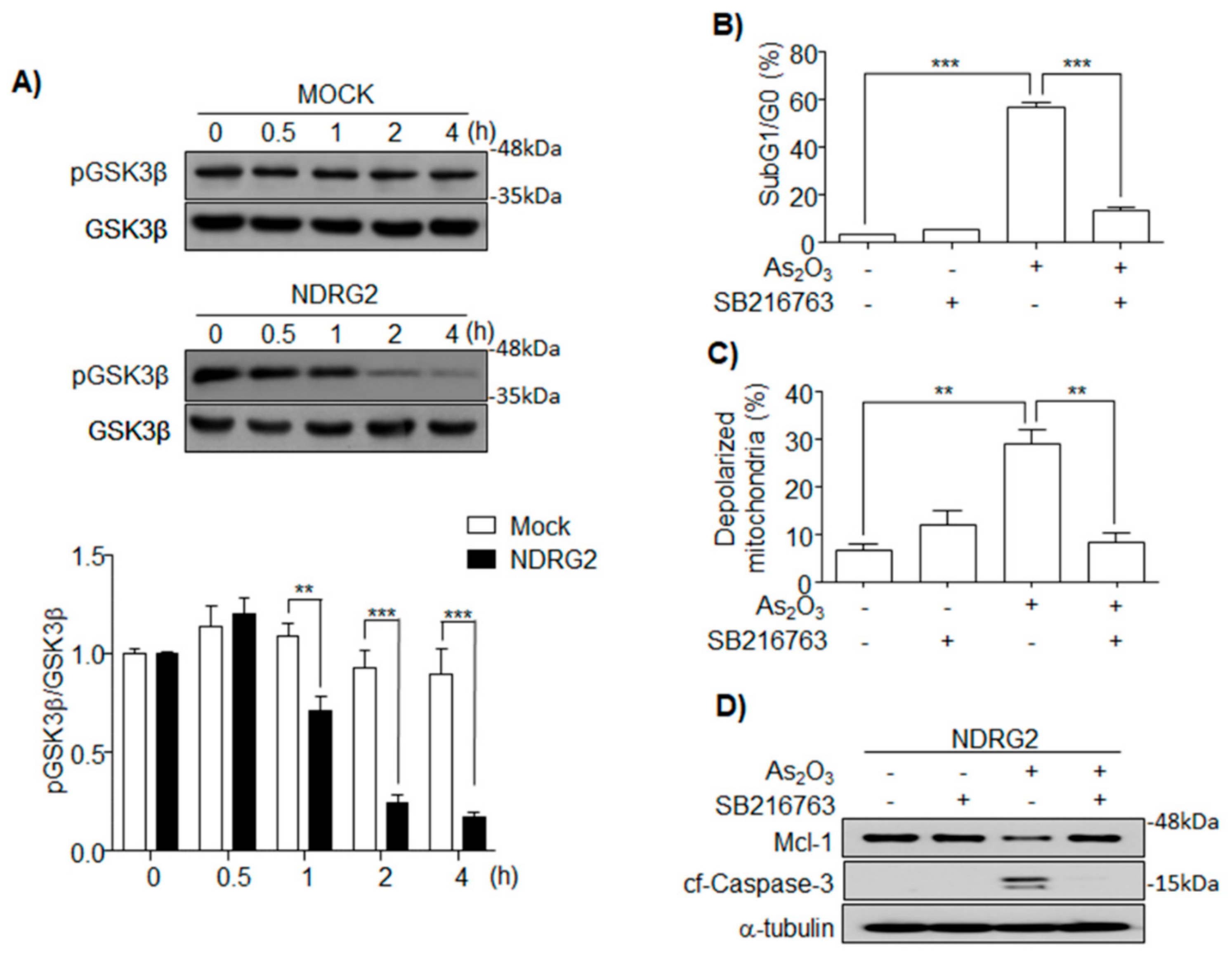
3.3. As2O3 Induces Mcl-1 Degradation through GSK3β Activation in U937-NDRG2
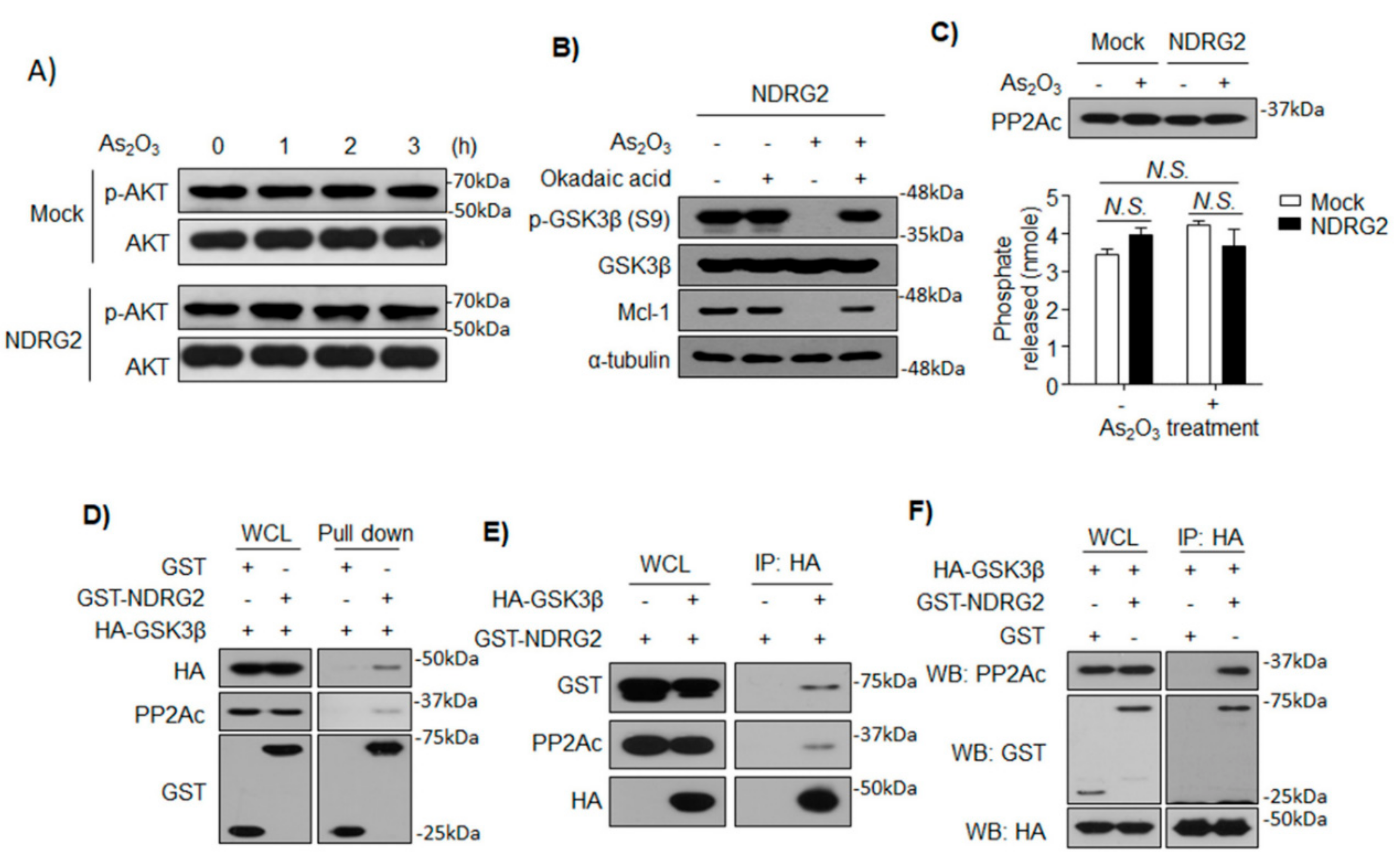
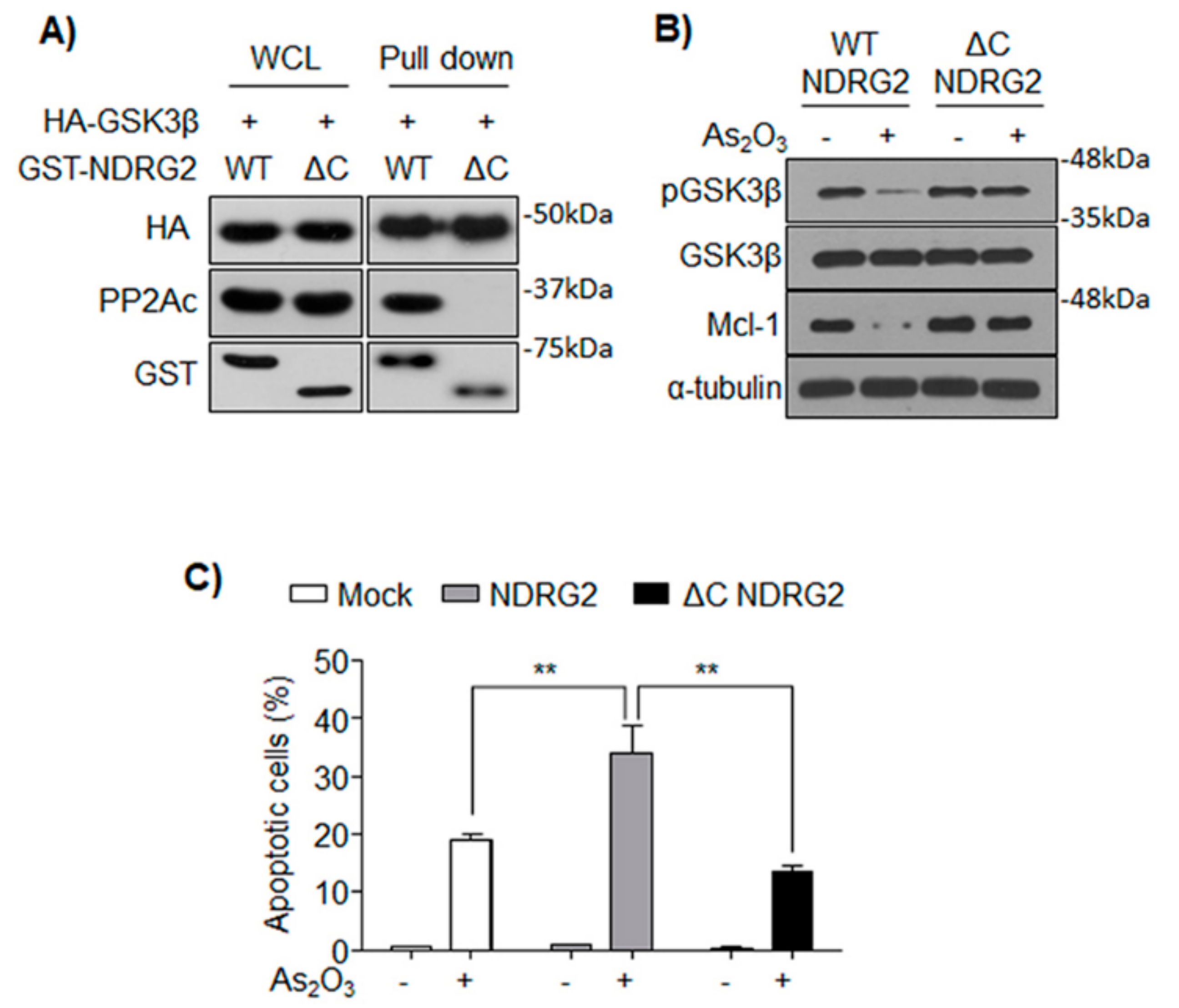
3.4. NDRG2 Mediates the Interaction between GSK3β and PP2A
3.5. The Formation of GSK3β/NDRG2/PP2A Complex Determines the Sensitivity of U937 to As2O3
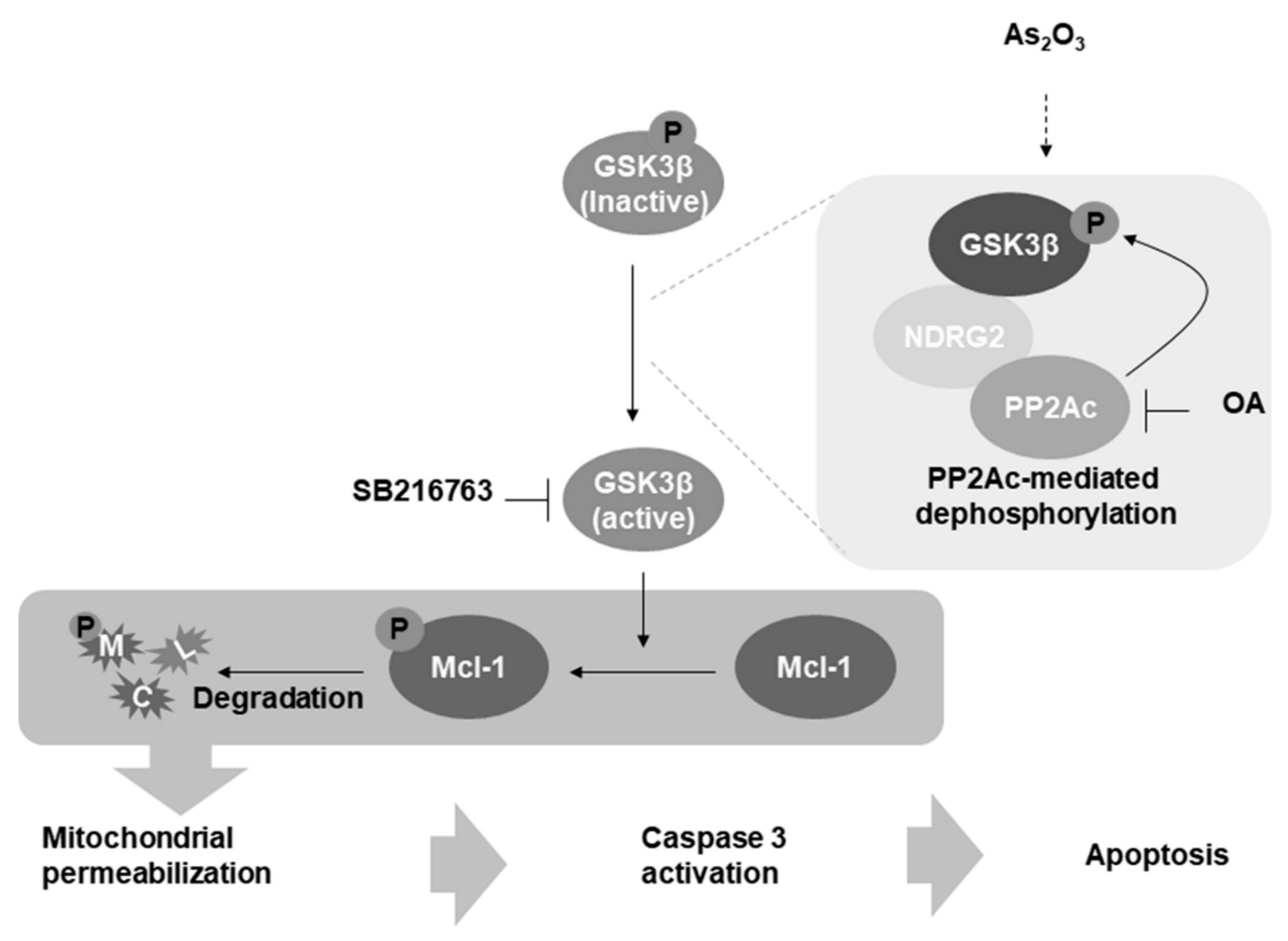
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, W.; Yang, Y.; Fan, C.; Ma, Z.; Deng, C.; Li, T.; Lv, J.; Yao, W.; Gao, J. Clinical and pathological significance of N-Myc downstream-regulated gene 2 (NDRG2) in diverse human cancers. Apoptosis 2016, 21, 675–682. [Google Scholar] [CrossRef]
- Hu, W.; Fan, C.; Jiang, P.; Ma, Z.; Yan, X.; Di, S.; Jiang, S.; Li, T.; Cheng, Y.; Yang, Y. Emerging role of N-myc downstream-regulated gene 2 (NDRG2) in cancer. Oncotarget 2016, 7, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.C.; Kang, Y.K.; Kim, W.H.; Jang, Y.J.; Kim, D.J.; Park, I.Y.; Sohn, B.H.; Sohn, H.A.; Lee, H.G.; Lim, J.S.; et al. Functional and clinical evidence for NDRG2 as a candidate suppressor of liver cancer metastasis. Cancer Res. 2008, 68, 4210–4220. [Google Scholar] [CrossRef]
- Choi, S.C.; Yoon, S.R.; Park, Y.P.; Song, E.Y.; Kim, J.W.; Kim, W.H.; Yang, Y.; Lim, J.S.; Lee, H.G. Expression of NDRG2 is related to tumor progression and survival of gastric cancer patients through Fas-mediated cell death. Exp. Mol. Med. 2007, 39, 705–714. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yoon, S.Y.; Kim, J.T.; Song, E.Y.; Lee, H.G.; Son, H.J.; Kim, S.Y.; Cho, D.; Choi, I.; Kim, J.H.C. NDRG2 expression decreases with tumor stages and regulates TCF/beta-catenin signaling in human colon carcinoma. Carcinogenesis 2009, 30, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.S.; Kim, D.; Kim, D.H.; Chang, H.H.; Sohn, K.C.; Kim, K.H.; Jung, S.H.; Lee, B.K.; Kim, J.H.; Kim, K.D. NDRG2 correlated with favorable recurrence-free survival inhibits metastasis of mouse breast cancer cells via attenuation of active TGF-beta production. Carcinogenesis 2012, 33, 1882–1888. [Google Scholar] [CrossRef]
- Manoukian, A.S.; Woodgett, J.R. Role of glycogen synthase kinase-3 in cancer: Regulation by Wnts and other signaling pathways. Adv. Cancer Res. 2002, 84, 203–229. [Google Scholar] [PubMed]
- Hardt, S.E.; Sadoshima, J. Glycogen synthase kinase-3beta: A novel regulator of cardiac hypertrophy and development. Cir. Res. 2002, 90, 1055–1063. [Google Scholar] [CrossRef]
- Kim, J.T.; Kim, J.W.; Kang, Y.H.; Kim, K.D.; Lee, S.J.; Choi, S.C.; Kim, K.S.; Chae, S.K.; Lim, J.S.; Lee, H.G. NDRG2 and PRA1 interact and synergistically inhibit T-cell factor/beta-catenin signaling. FEBS Lett. 2012, 586, 3962–3968. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kang, H.B.; Yim, H.S.; Kim, J.H.; Kim, J.W. NDRG2 positively regulates E-cadherin expression and prolongs overall survival in colon cancer patients. Oncol. Rep. 2013, 30, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Nakahata, S.; Ichikawa, T.; Maneesaay, P.; Saito, Y.; Nagai, K.; Tamura, T.; Manachai, N.; Yamakawa, N.; Hamasaki, M.; Kitabayashi, I.; et al. Loss of NDRG2 expression activates PI3K-AKT signalling via PTEN phosphorylation in ATLL and other cancers. Nat. Commun. 2014, 5, 3393. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, T.; Nakahata, S.; Tamura, T.; Manachai, N.; Morishita, K. The loss of NDRG2 expression improves depressive behavior through increased phosphorylation of GSK3beta. Cell Signal. 2015, 27, 2087–2098. [Google Scholar] [CrossRef]
- Soignet, S.L.; Maslak, P.; Wang, Z.G.; Jhanwar, S.; Calleja, E.; Dardashti, L.J.; Corso, D.; DeBlasio, A.; Gabrilove, J.; Scheinberg, D.A.; et al. Complete remission after treatment of acute promyelocytic leukemia with arsenic trioxide. New Engl. J. Med. 1998, 339, 1341–1348. [Google Scholar] [CrossRef]
- Niu, C.; Yan, H.; Yu, T.; Sun, H.P.; Liu, J.X.; Li, X.S.; Wu, W.; Zhang, F.Q.; Chen, Y.; Zhou, L.; et al. Studies on treatment of acute promyelocytic leukemia with arsenic trioxide: Remission induction, follow-up, and molecular monitoring in 11 newly diagnosed and 47 relapsed acute promyelocytic leukemia patients. Blood 1999, 94, 3315–3324. [Google Scholar]
- Mathews, V.; George, B.; Lakshmi, K.M.; Viswabandya, A.; Bajel, A.; Balasubramanian, P.; Shaji, R.V.; Srivastava, V.M.; Srivastava, A.; Chandy, M. Single-agent arsenic trioxide in the treatment of newly diagnosed acute promyelocytic leukemia: Durable remissions with minimal toxicity. Blood 2006, 107, 2627–2632. [Google Scholar] [CrossRef]
- Ravandi, F.; van Besien, K. Clinical activity of arsenic trioxide in Burkitt-like lymphoma. Leukemia 2003, 17, 271–272. [Google Scholar] [CrossRef] [Green Version]
- Akao, Y.; Yamada, H.; Nakagawa, Y. Arsenic-induced apoptosis in malignant cells in vitro. Leuk. Lymphoma 2000, 37, 53–63. [Google Scholar] [CrossRef]
- Rust, D.M.; Soignet, S.L. Risk/benefit profile of arsenic trioxide. Oncologist 2001, 6 Suppl 2, 29–32. [Google Scholar] [CrossRef]
- Mills, J.R.; Hippo, Y.; Robert, F.; Chen, S.M.; Malina, A.; Lin, C.J.; Trojahn, U.; Wendel, H.G.; Charest, A.; Bronson, R.T.; et al. mTORC1 promotes survival through translational control of Mcl-1. Proc. Natl. Acad. Sci. USA 2008, 105, 10853–10858. [Google Scholar] [CrossRef] [Green Version]
- Martelli, A.M.; Evangelisti, C.; Chiarini, F.; Grimaldi, C.; Manzoli, L.; McCubrey, J.A. Targeting the PI3K/AKT/mTOR signaling network in acute myelogenous leukemia. Exp. Opin. Investig. Drugs 2009, 18, 1333–1349. [Google Scholar] [CrossRef]
- Thomas, L.W.; Lam, C.; Edwards, S.W. Mcl-1; the molecular regulation of protein function. FEBS Lett. 2010, 584, 2981–2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Xia, L.; Gabrilove, J.; Waxman, S.; Jing, Y. Downregulation of Mcl-1 through GSK-3beta activation contributes to arsenic trioxide-induced apoptosis in acute myeloid leukemia cells. Leukemia 2013, 27, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Dai, J.; Chalmers-Redman, R.M.; Tatton, W.G.; Waxman, S. Arsenic trioxide selectively induces acute promyelocytic leukemia cell apoptosis via a hydrogen peroxide-dependent pathway. Blood 1999, 94, 2102–2111. [Google Scholar]
- Iwama, K.; Nakajo, S.; Aiuchi, T.; Nakaya, K. Apoptosis induced by arsenic trioxide in leukemia U937 cells is dependent on activation of p38, inactivation of ERK and the Ca2+-dependent production of superoxide. Int. J. Cancer 2001, 92, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Dahia, P.L.; Aguiar, R.C.; Alberta, J.; Kum, J.B.; Caron, S.; Sill, H.; Marsh, D.J.; Ritz, J.; Freedman, A.; Stiles, C.; et al. PTEN is inversely correlated with the cell survival factor Akt/PKB and is inactivated via multiple mechanismsin haematological malignancies. Hum. Mol. Genet. 1999, 8, 185–193. [Google Scholar] [CrossRef]
- Choi, S.C.; Kim, K.D.; Kim, J.T.; Oh, S.S.; Yoon, S.Y.; Song, E.Y.; Lee, H.G.; Choe, Y.K.; Choi, I.; Lim, J.S.; et al. NDRG2 is one of novel intrinsic factors for regulation of IL-10 production in human myeloid cell. Biochem. Biophys. Res. Commun. 2010, 396, 684–690. [Google Scholar] [CrossRef]
- Lengfelder, E.; Hofmann, W.K.; Nowak, D. Impact of arsenic trioxide in the treatment of acute promyelocytic leukemia. Leukemia 2012, 26, 433–442. [Google Scholar] [CrossRef]
- Kchour, G.; Tarhini, M.; Kooshyar, M.M.; El Hajj, H.; Wattel, E.; Mahmoudi, M.; Hatoum, H.; Rahimi, H.; Maleki, M.; Rafatpanah, H.; et al. Phase 2 study of the efficacy and safety of the combination of arsenic trioxide, interferon alpha, and zidovudine in newly diagnosed chronic adult T-cell leukemia/lymphoma (ATL). Blood 2009, 113, 6528–6532. [Google Scholar] [CrossRef]
- Uslu, R.; Sanli, U.A.; Sezgin, C.; Karabulut, B.; Terzioglu, E.; Omay, S.B.; Goker, E. Arsenic trioxide-mediated cytotoxicity and apoptosis in prostate and ovarian carcinoma cell lines. Clin. Cancer Res. 2000, 6, 4957–4964. [Google Scholar]
- Rousselot, P.; Larghero, J.; Arnulf, B.; Poupon, J.; Royer, B.; Tibi, A.; Madelaine-Chambrin, I.; Cimerman, P.; Chevret, S.; Hermine, O.; et al. A clinical and pharmacological study of arsenic trioxide in advanced multiple myeloma patients. Leukemia 2004, 18, 1518–1521. [Google Scholar] [CrossRef] [Green Version]
- Maurer, U.; Charvet, C.; Wagman, A.S.; Dejardin, E.; Green, D.R. Glycogen synthase kinase-3 regulates mitochondrial outer membrane permeabilization and apoptosis by destabilization of MCL-1. Mol. Cell 2006, 21, 749–760. [Google Scholar] [CrossRef]
- Ding, Q.; He, X.; Hsu, J.M.; Xia, W.; Chen, C.T.; Li, L.Y.; Lee, D.F.; Liu, J.C.; Zhong, Q.; Wang, X.; et al. Degradation of Mcl-1 by beta-TrCP mediates glycogen synthase kinase 3-induced tumor suppression and chemosensitization. Mol. Cell Biol. 2007, 27, 4006–4017. [Google Scholar] [CrossRef]
- Zhong, Q.; Gao, W.; Du, F.; Wang, X. Mule/ARF-BP1, a BH3-only E3 ubiquitin ligase, catalyzes the polyubiquitination of Mcl-1 and regulates apoptosis. Cell 2005, 121, 1085–1095. [Google Scholar] [CrossRef]
- Inuzuka, H.; Shaik, S.; Onoyama, I.; Gao, D.; Tseng, A.; Maser, R.S.; Zhai, B.; Wan, L.; Gutierrez, A.; Lau, A.W.; et al. SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for ubiquitylation and destruction. Nature 2011, 471, 104–109. [Google Scholar] [CrossRef]
- Chen, P.; Zhan, W.; Wang, B.; You, P.; Jin, Q.; Hou, D.; Wang, X.; You, R.; Zou, H.; Chen, Y.; et al. Homoharringtonine potentiates the antileukemic activity of arsenic trioxide against acute myeloid leukemia cells. Exp. Cell Res. 2019, 376, 114–123. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Bertrand, F.E.; Davis, N.M.; Sokolosky, M.; Abrams, S.L.; Montalto, G.; D’Assoro, A.B.; Libra, M.; Nicoletti, F.; et al. GSK-3 as potential target for therapeutic intervention in cancer. Oncotarget 2014, 5, 2881–2911. [Google Scholar] [CrossRef] [Green Version]
- Aggerholm, A.; Gronbaek, K.; Guldberg, P.; Hokland, P. Mutational analysis of the tumour suppressor gene MMAC1/PTEN in malignant myeloid disorders. Eur. J. Haematol. 2000, 65, 109–113. [Google Scholar] [CrossRef]
- Mitra, A.; Menezes, M.E.; Pannell, L.K.; Mulekar, M.S.; Honkanen, R.E.; Shevde, L.A.; Samant, R.S. DNAJB6 chaperones PP2A mediated dephosphorylation of GSK3beta to downregulate beta-catenin transcription target, osteopontin. Oncogene 2012, 31, 4472–4483. [Google Scholar] [CrossRef]
- Chang, Y.W.; Chen, M.W.; Chiu, C.F.; Hong, C.C.; Cheng, C.C.; Hsiao, M.; Chen, C.A.; Wei, L.H.; Su, J.L. Arsenic trioxide inhibits CXCR4-mediated metastasis by interfering miR-520h/PP2A/NF-kappaB signaling in cervical cancer. Ann. Surg. Oncol. 2014, 21, S687–S695. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Han, H.-T.; Oh, S.-S.; Kim, D.H.; Jeong, J.-W.; Lee, K.W.; Kim, M.; Lim, J.S.; Cho, Y.Y.; Hwangbo, C.; et al. NDRG2 Sensitizes Myeloid Leukemia to Arsenic Trioxide via GSK3β–NDRG2–PP2A Complex Formation. Cells 2019, 8, 495. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050495
Park S, Han H-T, Oh S-S, Kim DH, Jeong J-W, Lee KW, Kim M, Lim JS, Cho YY, Hwangbo C, et al. NDRG2 Sensitizes Myeloid Leukemia to Arsenic Trioxide via GSK3β–NDRG2–PP2A Complex Formation. Cells. 2019; 8(5):495. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050495
Chicago/Turabian StylePark, Soojong, Hyun-Tak Han, Sang-Seok Oh, Dong Hyeok Kim, Jin-Woo Jeong, Ki Won Lee, Minju Kim, Jong Seok Lim, Yong Yeon Cho, Cheol Hwangbo, and et al. 2019. "NDRG2 Sensitizes Myeloid Leukemia to Arsenic Trioxide via GSK3β–NDRG2–PP2A Complex Formation" Cells 8, no. 5: 495. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050495