Establishment of Highly Transplantable Cholangiocarcinoma Cell Lines from a Patient-Derived Xenograft Mouse Model

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. CCA Tissue Collection and Storage
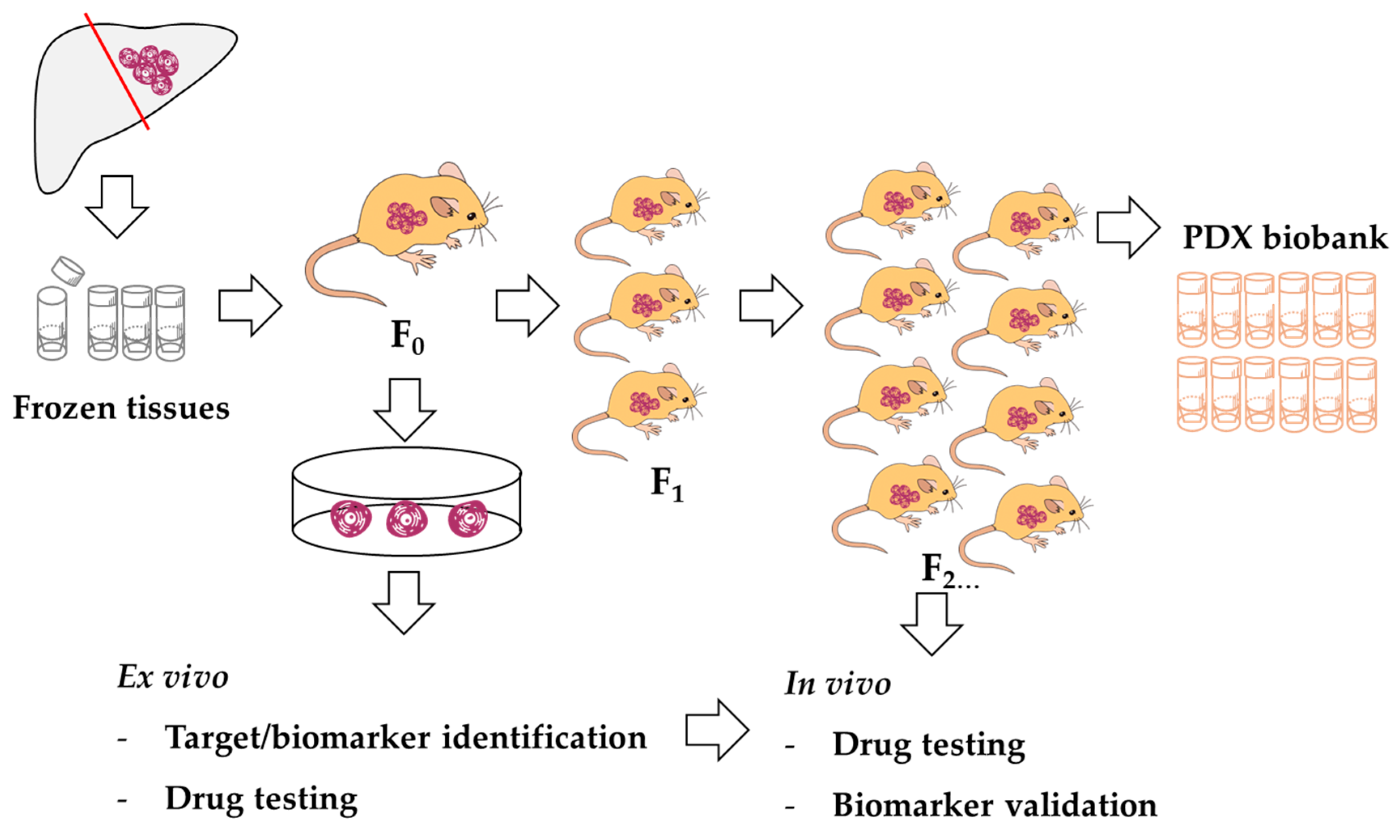
2.3. Cell Line Establishment
2.4. The Expressions of Bile Duct and Hepatocyte-Related Genes
2.5. Cell Line Authentication and TP53 Mutation Analysis
2.6. Xenograft Transplantation of Cell Lines
2.7. Histological Characterization and Evaluation
2.8. Statistical Analysis
3. Results
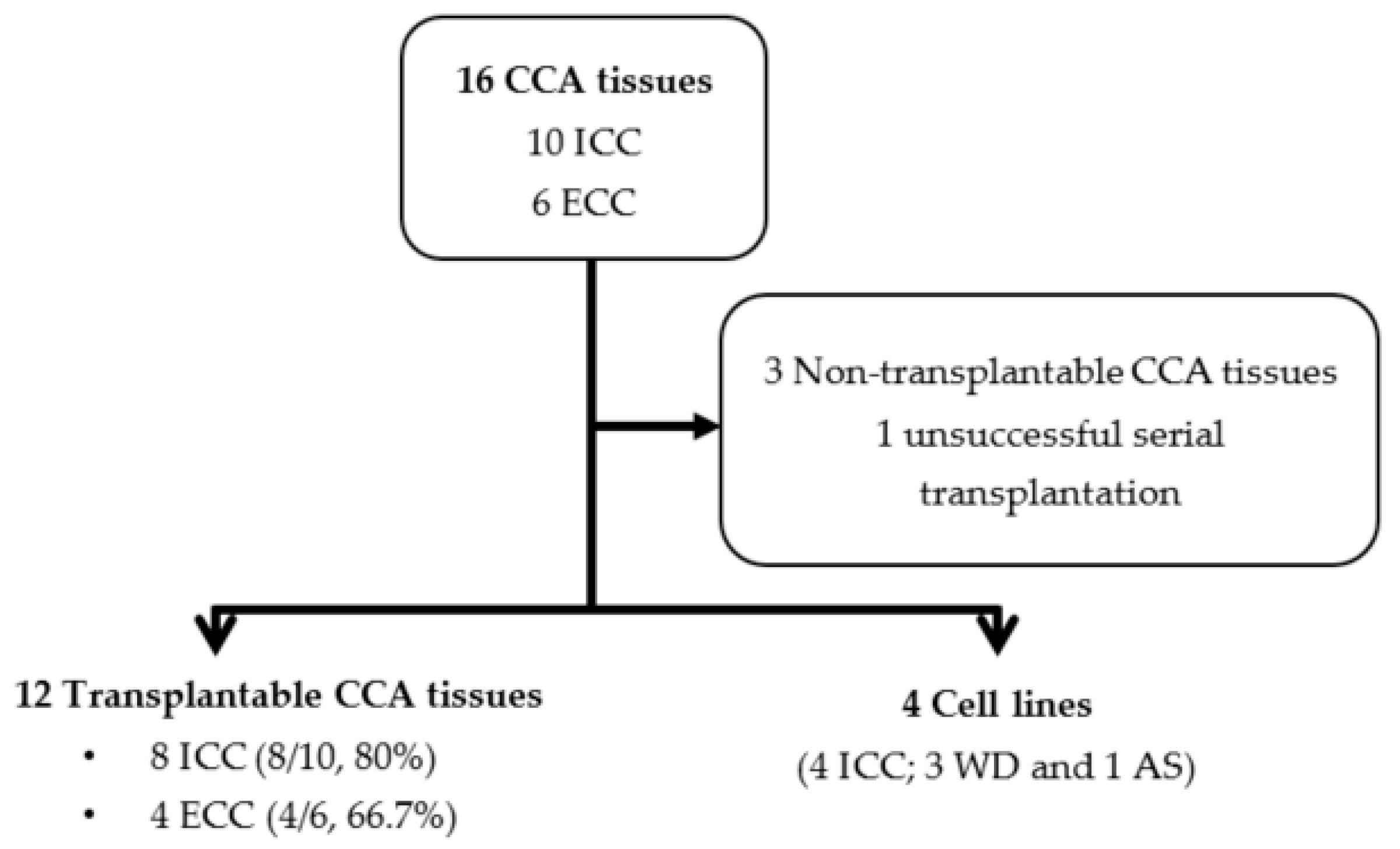
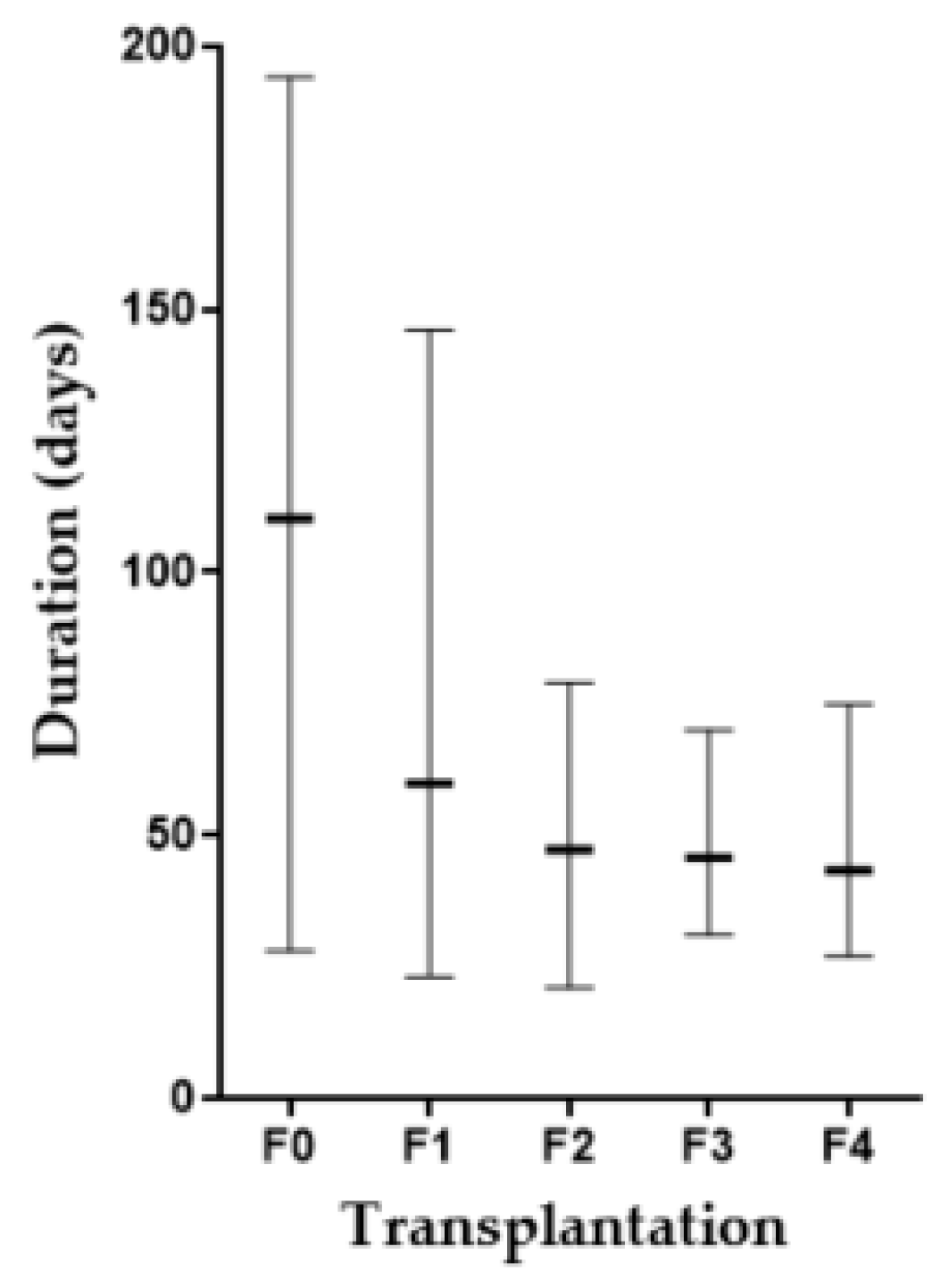
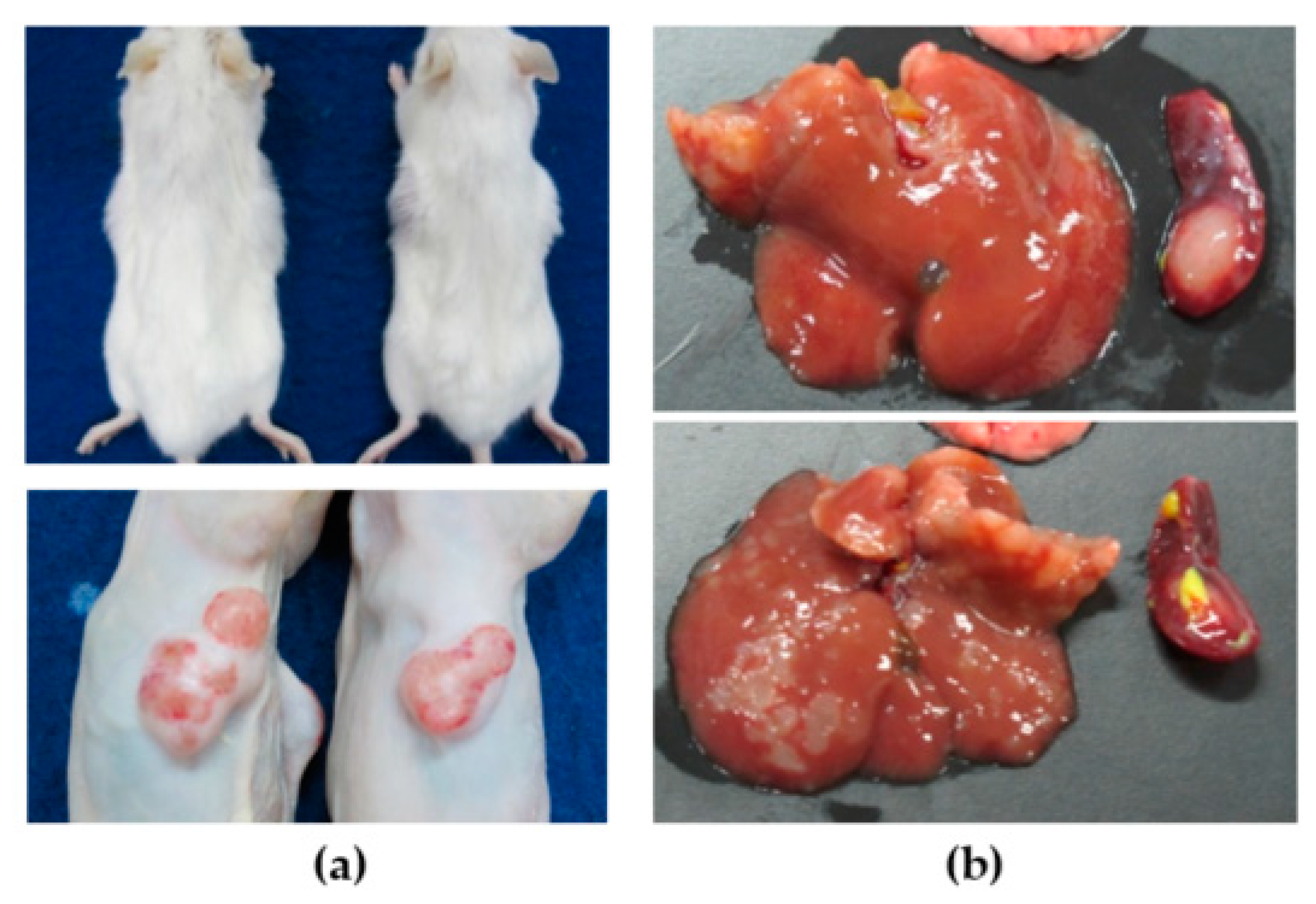
3.1. CCA Patient Tissue Transplantation
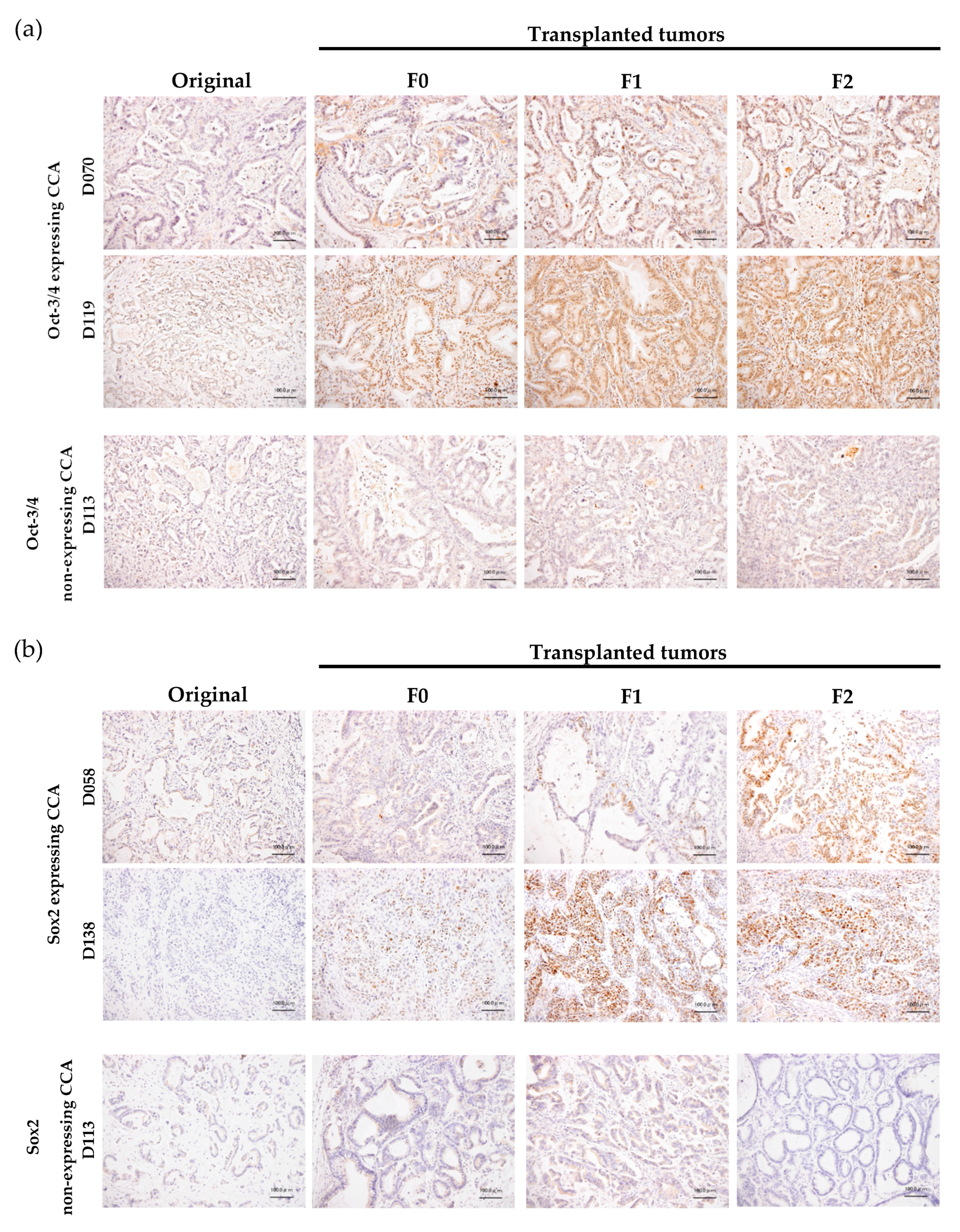
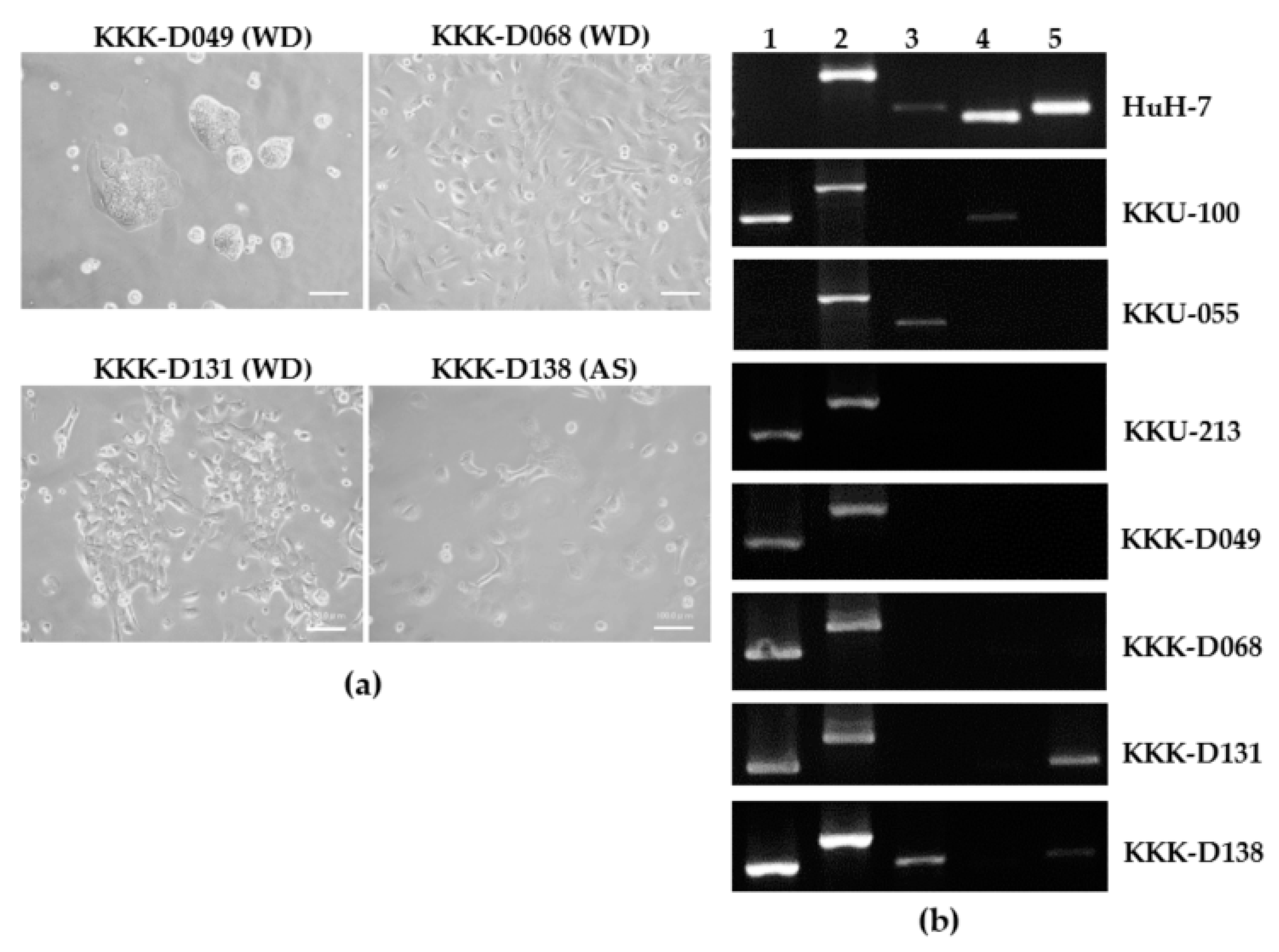
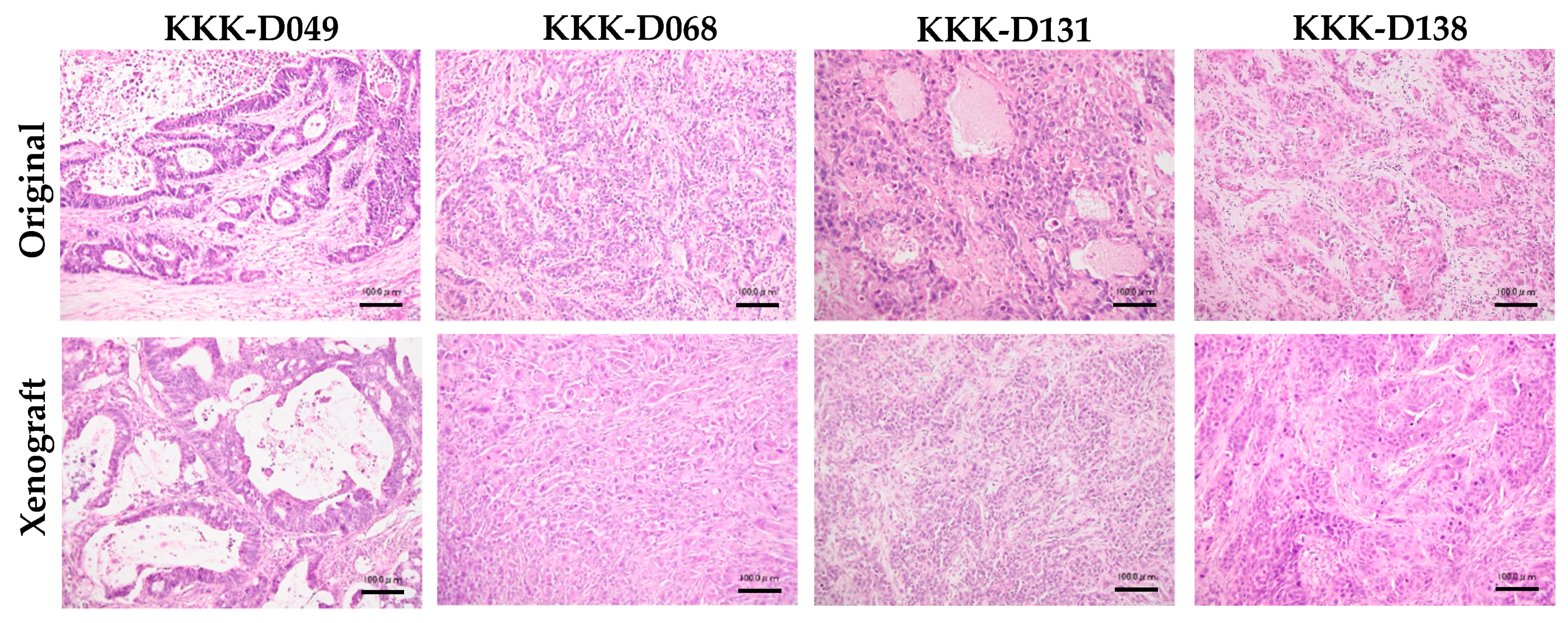
3.2. Cell Line Establishment and Characterization
3.3. Cell Line Transplantation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sripa, B.; Pairojkul, C. Cholangiocarcinoma: Lessons from Thailand. Curr. Opin. Gastroenterol. 2008, 24, 349–356. [Google Scholar] [CrossRef]
- Bridgewater, J.; Galle, P.R.; Khan, S.A.; Llovet, J.M.; Park, J.-W.; Patel, T.; Pawlik, T.M.; Gores, G.J. Guidelines for the diagnosis and management of intrahepatic cholangiocarcinoma. J. Hepatol. 2014, 60, 1268–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Expert consensus document: Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef] [PubMed]
- Doherty, B.; Nambudiri, V.E.; Palmer, W.C. Update on the Diagnosis and Treatment of Cholangiocarcinoma. Gastroenterol. Rep. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.C.; Coburn, N.G.; Baxter, N.N.; Kiss, A.; Law, C.H. Surgical management of intrahepatic cholangiocarcinoma—A population-based study. Ann. Surg. Oncol. 2008, 15, 600–608. [Google Scholar] [CrossRef]
- Vogel, A.; Wege, H.; Caca, K.; Nashan, B.; Neumann, U. The Diagnosis and Treatment of Cholangiocarcinoma. Dtsch. Aerzteblatt Int. 2014, 111, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Horgan, A.M.; Amir, E.; Walter, T.; Knox, J.J. Adjuvant Therapy in the Treatment of Biliary Tract Cancer: A Systematic Review and Meta-Analysis. J. Clin. Oncol. 2012, 30, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Patel, T. New insights into the molecular pathogenesis of intrahepatic cholangiocarcinoma. J. Gastroenterol. 2014, 49, 165–172. [Google Scholar] [CrossRef]
- Tyson, G.L.; El-Serag, H.B. Risk factors for cholangiocarcinoma. Hepatology 2011, 54, 173–184. [Google Scholar] [CrossRef]
- Sripa, B.; Brindley, P.J.; Mulvenna, J.; Laha, T.; Smout, M.J.; Mairiang, E.; Bethony, J.M.; Loukas, A. The tumorigenic liver fluke Opisthorchis viverrini –multiple pathways to cancer. Trends Parasitol. 2012, 28, 395–407. [Google Scholar] [CrossRef]
- Jinawath, N.; Chamgramol, Y.; Furukawa, Y.; Obama, K.; Tsunoda, T.; Sripa, B.; Pairojkul, C.; Nakamura, Y. Comparison of gene expression profiles between Opisthorchis viverrini and non-Opisthorchis viverrini associated human intrahepatic cholangiocarcinoma. Hepatology 2006, 44, 1025–1038. [Google Scholar] [CrossRef]
- Ong, C.K.; Subimerb, C.; Pairojkul, C.; Wongkham, S.; Cutcutache, I.; Yu, W.; McPherson, J.R.; E Allen, G.; Ng, C.C.Y.; Wong, B.H.; et al. Exome sequencing of liver fluke–associated cholangiocarcinoma. Nat. Genet. 2012, 44, 690–693. [Google Scholar] [CrossRef]
- Chan-On, W.; Nairismägi, M.-L.; Ong, C.K.; Lim, W.K.; Dima, S.; Pairojkul, C.; Lim, K.H.; McPherson, J.R.; Cutcutache, I.; Heng, H.L.; et al. Exome sequencing identifies distinct mutational patterns in liver fluke–related and non-infection-related bile duct cancers. Nat. Genet. 2013, 45, 1474–1478. [Google Scholar] [CrossRef]
- Wardell, C.P.; Fujita, M.; Yamada, T.; Simbolo, M.; Fassan, M.; Karlić, R.; Polak, P.; Kim, J.; Hatanaka, Y.; Maejima, K.; et al. Genomic characterization of biliary tract cancers identifies driver genes and predisposing mutations. J. Hepatol. 2018, 68, 959–969. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.B.; Spee, B.; Blechacz, B.R.; Avital, I.; Komuta, M.; Barbour, A.; Conner, E.A.; Gillen, M.C.; Roskams, T.; Roberts, L.R.; et al. Genomic and genetic characterization of cholangiocarcinoma identifies therapeutic targets for tyrosine kinase inhibitors. Gastroenterology 2012, 142, 1021–1031.e15. [Google Scholar] [CrossRef]
- Saha, S.K.; Gordan, J.D.; Kleinstiver, B.P.; Vu, P.; Najem, M.S.; Yeo, J.-C.; Shi, L.; Kato, Y.; Levin, R.S.; Webber, J.T.; et al. Isocitrate dehydrogenase mutations confer dasatinib hypersensitivity and SRC-dependence in intrahepatic cholangiocarcinoma. Cancer Discov. 2016, 6, 727–739. [Google Scholar] [CrossRef]
- Sia, D.; Hoshida, Y.; Villanueva, A.; Roayaie, S.; Ferrer, J.; Tabak, B.; Peix, J.; Sole, M.; Tovar, V.; Alsinet, C.; et al. Integrative Molecular Analysis of Intrahepatic Cholangiocarcinoma Reveals 2 Classes That Have Different Outcomes. Gastroenterology 2013, 144, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Jusakul, A.; Cutcutache, I.; Yong, C.H.; Lim, J.Q.; Ni Huang, M.; Padmanabhan, N.; Nellore, V.; Kongpetch, S.; Ng, A.W.T.; Ng, L.M.; et al. Whole-Genome and Epigenomic Landscapes of Etiologically Distinct Subtypes of Cholangiocarcinoma. Cancer Discov. 2017, 7, 1116–1135. [Google Scholar] [CrossRef] [PubMed]
- Chaisaingmongkol, J.; Budhu, A.; Dang, H.; Rabibhadana, S.; Pupacdi, B.; Kwon, S.M.; Forgues, M.; Pomyen, Y.; Bhudhisawasdi, V.; Lertprasertsuke, N.; et al. Common Molecular Subtypes among Asian Hepatocellular Carcinoma and Cholangiocarcinoma. Cancer Cell 2017, 32, 57–70.e3. [Google Scholar] [CrossRef] [PubMed]
- Kopetz, S.; Lemos, R.; Powis, G. The Promise of Patient-Derived Xenografts: The best laid plans of mice and men. Clin. Cancer Res. 2012, 18, 5160–5162. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Seol, H.S.; Chang, S. The Generation and Application of Patient-Derived Xenograft Model for Cancer Research. Cancer Res. Treat. Off. J. Korean Cancer Assoc. 2018, 50, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Kim, M.-S.; Kim, S.J.; An, S.; Park, J.; Park, H.; Lee, J.H.; Song, K.-B.; Hwang, D.W.; Chang, S.; et al. Establishment and characterization of 6 novel patient-derived primary pancreatic ductal adenocarcinoma cell lines from Korean pancreatic cancer patients. Cancer Cell Int. 2017, 17, 47. [Google Scholar] [CrossRef]
- Ku, J.-L.; Yoon, K.-A.; Kim, I.-J.; Kim, W.-H.; Jang, J.-Y.; Suh, K.-S.; Kim, S.-W.; Park, Y.-H.; Hwang, J.-H.; Yoon, Y.-B.; et al. Establishment and characterisation of six human biliary tract cancer cell lines. Br. J. Cancer 2002, 87, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Okada, S.; Vaeteewoottacharn, K.; Kariya, R. Establishment of a Patient-Derived Tumor Xenograft Model and Application for Precision Cancer Medicine. Chem. Pharm. 2018, 66, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Damhofer, H.; A Ebbing, E.; Steins, A.; Welling, L.; A Tol, J.; Krishnadath, K.K.; Van Leusden, T.; Van De Vijver, M.J.; Besselink, M.G.; Busch, O.R.; et al. Establishment of patient-derived xenograft models and cell lines for malignancies of the upper gastrointestinal tract. J. Transl. Med. 2015, 13, 115. [Google Scholar] [CrossRef]
- Pham, K.; Delitto, D.; Knowlton, A.E.; Hartlage, E.R.; Madhavan, R.; Gonzalo, D.H.; Thomas, R.M.; Behrns, K.E.; George, T.J.; Hughes, S.J.; et al. Isolation of Pancreatic Cancer Cells from a Patient-Derived Xenograft Model Allows for Practical Expansion and Preserved Heterogeneity in Culture. Am. J. Pathol. 2016, 186, 1537–1546. [Google Scholar] [CrossRef] [Green Version]
- Ojima, H.; Yoshikawa, D.; Ino, Y.; Shimizu, H.; Miyamoto, M.; Kokubu, A.; Hiraoka, N.; Morofuji, N.; Kondo, T.; Onaya, H.; et al. Establishment of six new human biliary tract carcinoma cell lines and identification of MAGEH1 as a candidate biomarker for predicting the efficacy of gemcitabine treatment. Cancer Sci. 2010, 101, 882–888. [Google Scholar] [CrossRef]
- Cavalloni, G.; Peraldo-Neia, C.; Sassi, F.; Chiorino, G.; Sarotto, I.; Aglietta, M.; Leone, F. Establishment of a patient-derived intrahepatic cholangiocarcinoma xenograft model with KRAS mutation. BMC Cancer 2016, 16, 288. [Google Scholar] [CrossRef]
- Sripa, B.; Leungwattanawanit, S.; Nitta, T.; Wongkham, C.; Bhudhisawasdi, V.; Puapairoj, A.; Sripa, C.; Miwa, M.; Sripa, S.L.B. Establishment and characterization of an opisthorchiasis-associated cholangiocarcinoma cell line (KKU-100). World J. Gastroenterol. 2005, 11, 3392–3397. [Google Scholar] [CrossRef]
- Nakabayashi, H.; Taketa, K.; Miyano, K.; Yamane, T.; Sato, J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res 1982, 42, 3858–3863. [Google Scholar]
- Ono, A.; Hattori, S.; Kariya, R.; Iwanaga, S.; Taura, M.; Harada, H.; Suzu, S.; Okada, S. Comparative Study of Human Hematopoietic Cell Engraftment into Balb/c and C57BL/6 Strain of Rag-2/Jak3 Double-Deficient Mice. J. Biomed. Biotechnol. 2011, 2011, 1–6. [Google Scholar] [CrossRef]
- Kariya, R.; Matsuda, K.; Gotoh, K.; Vaeteewoottacharn, K.; Hattori, S.; Okada, S. Establishment of nude mice with complete loss of lymphocytes and NK cells and application for in vivo bio-imaging. In Vivo 2014, 28, 779–784. [Google Scholar] [PubMed]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th Edition of the AJCC Cancer Staging Manual and the Future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Puthdee, N.; Vaeteewoottacharn, K.; Seubwai, W.; Wonkchalee, O.; Kaewkong, W.; Juasook, A.; Pinlaor, S.; Pairojkul, C.; Wongkham, C.; Okada, S.; et al. Establishment of an Allo-Transplantable Hamster Cholangiocarcinoma Cell Line and Its Application for In Vivo Screening of Anti-Cancer Drugs. Korean J. Parasitol. 2013, 51, 711–717. [Google Scholar] [CrossRef]
- Cai, J.; Zhao, Y.; Liu, Y.; Ye, F.; Song, Z.; Qin, H.; Meng, S.; Chen, Y.; Zhou, R.; Song, X.; et al. Directed differentiation of human embryonic stem cells into functional hepatic cells. Hepatology 2007, 45, 1229–1239. [Google Scholar] [CrossRef]
- Vaeteewoottacharn, K.; Kariya, R.; Dana, P.; Fujikawa, S.; Matsuda, K.; Ohkuma, K.; Kudo, E.; Kraiklang, R.; Wongkham, C.; Okada, S. Inhibition of carbonic anhydrase potentiates bevacizumab treatment in cholangiocarcinoma. Tumor Boil. 2016, 37, 9023–9035. [Google Scholar] [CrossRef]
- Detection of TP53 Mutations by Direct Sequencing. Available online: http://p53.iarc.fr/protocolsandtools.aspx (accessed on 20 April 2019).
- Vaeteewoottacharn, K.; Kariya, R.; Pothipan, P.; Fujikawa, S.; Pairojkul, C.; Waraasawapati, S.; Kuwahara, K.; Wongkham, C.; Wongkham, S.; Okada, S. Attenuation of CD47-SIRPalpha Signal in Cholangiocarcinoma Potentiates Tumor-Associated Macrophage-Mediated Phagocytosis and Suppresses Intrahepatic Metastasis. Transl. Oncol. 2019, 12, 217–225. [Google Scholar] [CrossRef]
- Hanna, N. Role of natural killer cells in control of cancer metastasis. Cancer Metastasis Rev. 1982, 1, 45–64. [Google Scholar] [CrossRef]
- Golan, H.; Shukrun, R.; Caspi, R.; Vax, E.; Pode-Shakked, N.; Goldberg, S.; Pleniceanu, O.; Bar-Lev, D.D.; Mark-Danieli, M.; Pri-Chen, S.; et al. In Vivo Expansion of Cancer Stemness Affords Novel Cancer Stem Cell Targets: Malignant Rhabdoid Tumor as an Example. Stem Cell Rep. 2018, 11, 795–810. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-J.; Jeong, S.-Y.; Kim, H.J. Y chromosome loss and other genomic alterations in hepatocellular carcinoma cell lines analyzed by CGH and CGH array. Cancer Genet. Cytogenet. 2006, 166, 56–64. [Google Scholar] [CrossRef]
- Wallrapp, C.; Hähnel, S.; Boeck, W.; Soder, A.; Mincheva, A.; Lichter, P.; Leder, G.; Gansauge, F.; Sorio, C.; Scarpa, A.; et al. Loss of the Y chromosome is a frequent chromosomal imbalance in pancreatic cancer and allows differentiation to chronic pancreatitis. Int. J. Cancer 2001, 91, 340–344. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Gender | Age | Subtype | TMN ** | Stage ** | Ov# | PDX ## | Histological Classification |
|---|---|---|---|---|---|---|---|---|
| D039 | F | 66 | ICC | T3N0M0 | III | No | − | WD, papillo-tubular adenocarcinoma |
| D042 | M | 56 | ECC | T2bN0M0 | II | No | − | Invasive, intraductal papillary carcinoma |
| D049 * | M | 55 | ICC | T2bN0M0 | II | Ov | + | WD, tubular adenocarcinoma |
| D058 | F | 64 | ICC | T3N1M0 | IVA | Ov | + | WD, tubular adenocarcinoma |
| D068 * | M | 61 | ICC | T2aN1M0 | IVA | No | + | WD, tubular adenocarcinoma with micropapillary foci |
| D070 | M | 65 | ICC | T3N1M0 | IVA | No | + | WD, tubular adenocarcinoma |
| D078 | F | 44 | ECC | T4N1M0 | IVA | No | + | WD, tubular adenocarcinoma |
| D088 | F | 68 | ICC | T3N0M0 | III | No | + | MD, tubular adenocarcinoma |
| D090 | F | 65 | ECC | T2bN0M0 | II | No | + | Invasive, intraductal papillary carcinoma |
| D096 | M | 45 | ECC | T3N1M0 | IIIB | No | + | WD, tubular adenocarcinoma |
| D106 | M | 54 | ECC | T2bN1M0 | IIIB | No | +/− | Invasive, intraductal papillary carcinoma |
| D113 | M | 70 | ICC | T3N0M0 | III | No | + | Invasive, intraductal papillary carcinoma |
| D117 | M | 58 | ICC | T3N1M0 | IVA | No | − | WD, tubular adenocarcinoma with micropapillary foci |
| D119 | M | 71 | ECC | T3N1M0 | IIIB | No | + | WD, tubular adenocarcinoma |
| D131 * | M | 66 | ICC | T3N1M0 | IVA | No | + | WD, tubular adenocarcinoma |
| D138 * | F | 60 | ICC | T3N0M0 | III | No | + | Adenosquamous carcinoma |
| Loci | D049 | D068 | D131 | D138 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WBC | Tissue | Cell | WBC | Tissue | Cell | WBC | Tissue | Cell | WBC | Tissue | Cell | |
| D8S1179 | 12, 17 | 12, 17 | 12, 17 | 12, 16 | 12, 16 | 12, 16 | 12, 13 | 12, 13 | 12, 13 | ND ** | 10, 14 | 10, 14 |
| D21S11 | 29, 30 | 29, 30 | 29, 30 | 30, 33.2 | 30, 33.2 | 30, 33.2 | 29 | 29 | 29 | 29, 31.2 | 29 | |
| D7S820 * | 8, 10 | 8, 10 | 8, 10 | 8, 10 | 8, 10 | 8, 10 | 8, 11 | 8, 11 | 8, 11 | 10, 11 | 10, 11 | |
| CSF1PO * | 11, 12 | 11, 12 | 11, 12 | 11 | 11 | 11 | 12, 13 | 12, 13, 14 | 14 | 10, 11 | 10, 11 | |
| D3S1358 | 15, 16 | 15, 16 | 15, 16 | 15 | 15 | 15 | 14, 15 | 14, 15 | 14, 15 | 16, 18 | 16, 18 | |
| TH01 * | 9 | 9 | 9 | 7 | 7 | 7 | 8, 9.3 | 8, 9.3 | 9.3 | 8, 9.3 | 8, 9.3 | |
| D13S317 * | 8, 9 | 8, 9 | 8, 9 | 8, 12 | 8, 12 | 8, 12 | 10, 11 | 10, 11 | 10, 11 | 8, 11 | 11 | |
| D16S539 * | 13, 14 | 13, 14 | 13, 14 | 9, 11 | 9, 11 | 9 | 9, 11 | 9, 11 | 11 | 9, 11 | 9, 11 | |
| D2S1338 | 19, 25 | 19, 25 | 19, 25 | 19 | 19 | 19 | 20, 23 | 20, 23 | 20, 23 | 24, 25 | 24, 25 | |
| D19S433 | 13, 15.2 | 13, 15.2 | 13, 15.2 | 14, 14.2 | 14, 14.2 | 14, 14.2 | 13.2, 14 | 13.2, 14 | 13.2 | 13.2, 14.2 | 13.2, 14.2 | |
| vWA * | 14, 17 | 14, 17 | 14, 17 | 14, 16 | 14, 16 | 14, 16 | 14, 16 | 14, 16 | 14, 16 | 14, 18 | 14, 18 | |
| TPOX * | 8, 9 | 8, 9 | 8, 9 | 8, 11 | 8, 11 | 8, 11 | 8, 11 | 8, 11 | 8, 11 | 11 | 11 | |
| D18S51 | 11, 16 | 11 | 11 | 12 | 12 | 12 | 17 | 17 | 17 | 15 | 15 | |
| D5S818 * | 10, 12 | 10, 12 | 10, 12 | 11, 12 | 11, 12 | 11, 12 | 10, 12 | 10, 12, 13 | 13 | 9, 10 | 9, 10 | |
| FGA | 23, 24.2 | 23, 24.2 | 23, 24.2 | 23, 25 | 23, 25 | 23, 25 | 19, 21 | 19, 21 | 21 | 18, 24.2 | 18, 24.2 | |
| Amelogenin | X, Y | X, Y | X, Y | X, Y | X, Y | X | X, Y | X, Y | X | X, X | X, X | |
| Cell Lines | Route | Transplantation Rate (%) |
|---|---|---|
| KKK-D049 | SC | 4/4 (100%) |
| IS | 1/2 (50%) | |
| KKK-D068 | SC | 4/4 (100%) |
| IS | 2/2 (100%) | |
| KKK-D131 | SC | 4/4 (100%) |
| IS | 2/2 (100%) | |
| KKK-D138 | SC | 4/4 (100%) |
| IS | 2/2 (100%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaeteewoottacharn, K.; Pairojkul, C.; Kariya, R.; Muisuk, K.; Imtawil, K.; Chamgramol, Y.; Bhudhisawasdi, V.; Khuntikeo, N.; Pugkhem, A.; Saeseow, O.-T.; et al. Establishment of Highly Transplantable Cholangiocarcinoma Cell Lines from a Patient-Derived Xenograft Mouse Model. Cells 2019, 8, 496. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050496
Vaeteewoottacharn K, Pairojkul C, Kariya R, Muisuk K, Imtawil K, Chamgramol Y, Bhudhisawasdi V, Khuntikeo N, Pugkhem A, Saeseow O-T, et al. Establishment of Highly Transplantable Cholangiocarcinoma Cell Lines from a Patient-Derived Xenograft Mouse Model. Cells. 2019; 8(5):496. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050496
Chicago/Turabian StyleVaeteewoottacharn, Kulthida, Chawalit Pairojkul, Ryusho Kariya, Kanha Muisuk, Kanokwan Imtawil, Yaovalux Chamgramol, Vajarabhongsa Bhudhisawasdi, Narong Khuntikeo, Ake Pugkhem, O-Tur Saeseow, and et al. 2019. "Establishment of Highly Transplantable Cholangiocarcinoma Cell Lines from a Patient-Derived Xenograft Mouse Model" Cells 8, no. 5: 496. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050496