Heterologous Immunity between Adenoviruses and Hepatitis C Virus (HCV): Recombinant Adenovirus Vaccine Vectors Containing Antigens from Unrelated Pathogens Induce Cross-Reactive Immunity Against HCV Antigens

 , ,
, ,
Abstract
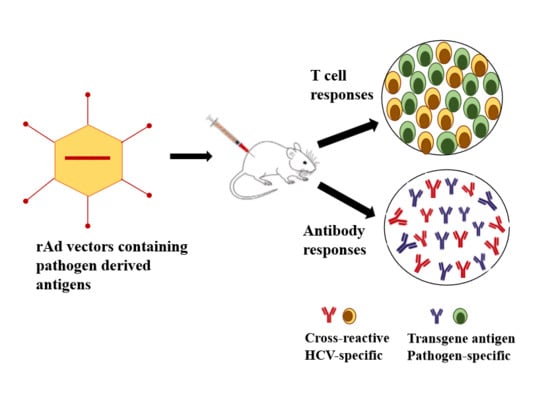
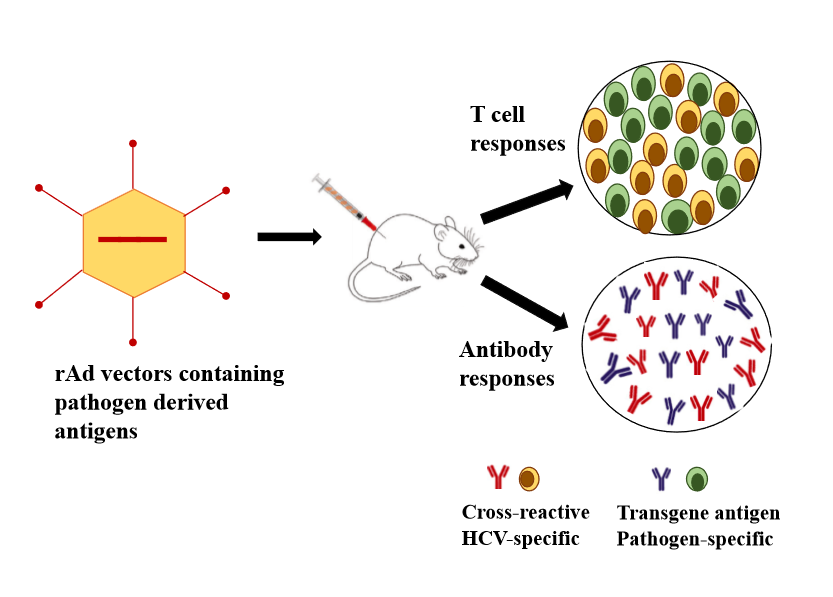
:
1. Introduction
2. Methods and Materials
2.1. Adenovirus Vector
2.2. Cloning of Mtb Ag85B, HIV-Gag and HIV-nef into Plasmids
2.3. Construction of Recombinant Adenovirus Vectors
2.4. Mice Immunizations
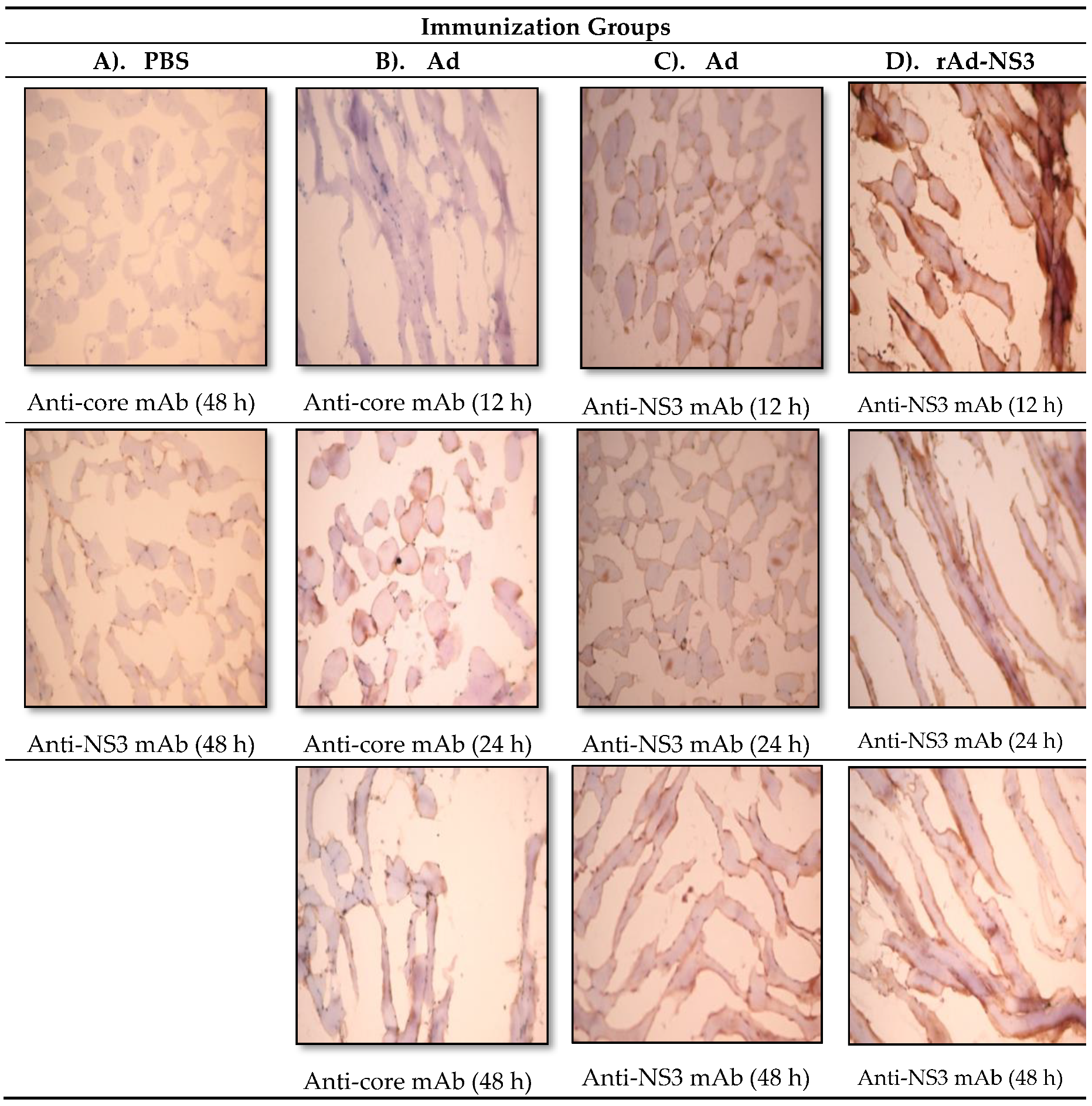
2.5. Immunohistochemistry
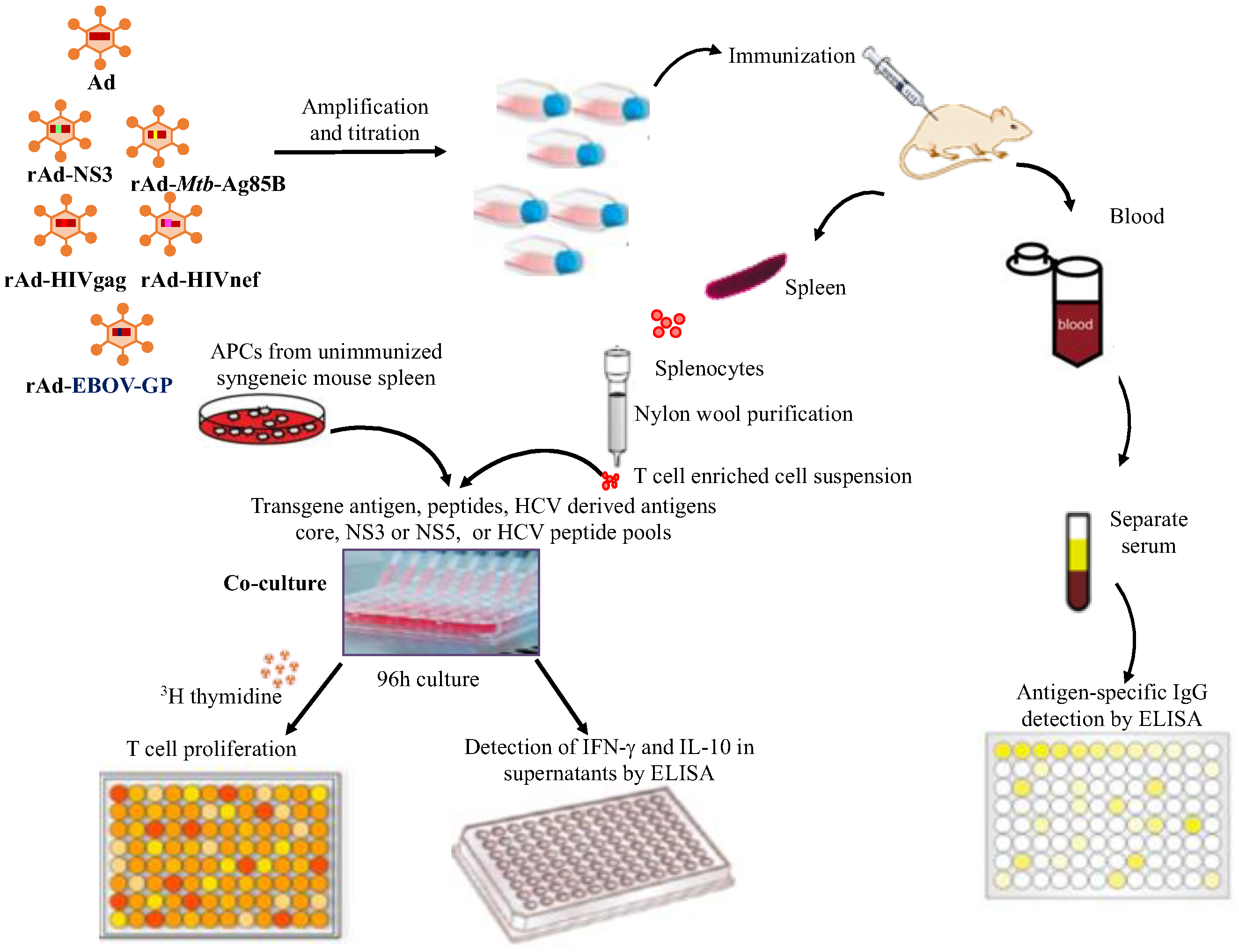
2.6. Experimental Design for High Throughput Screening of Cross-Reactive Humoral and Cellular Immune Responses Induced by Recombinant Ads Containing Antigens from Unrelated Pathogens
2.7. T Cell Proliferation Assay
2.8. Cytokine ELISA
2.9. Antibody ELISA
2.10. Statistical Analysis
3. Results
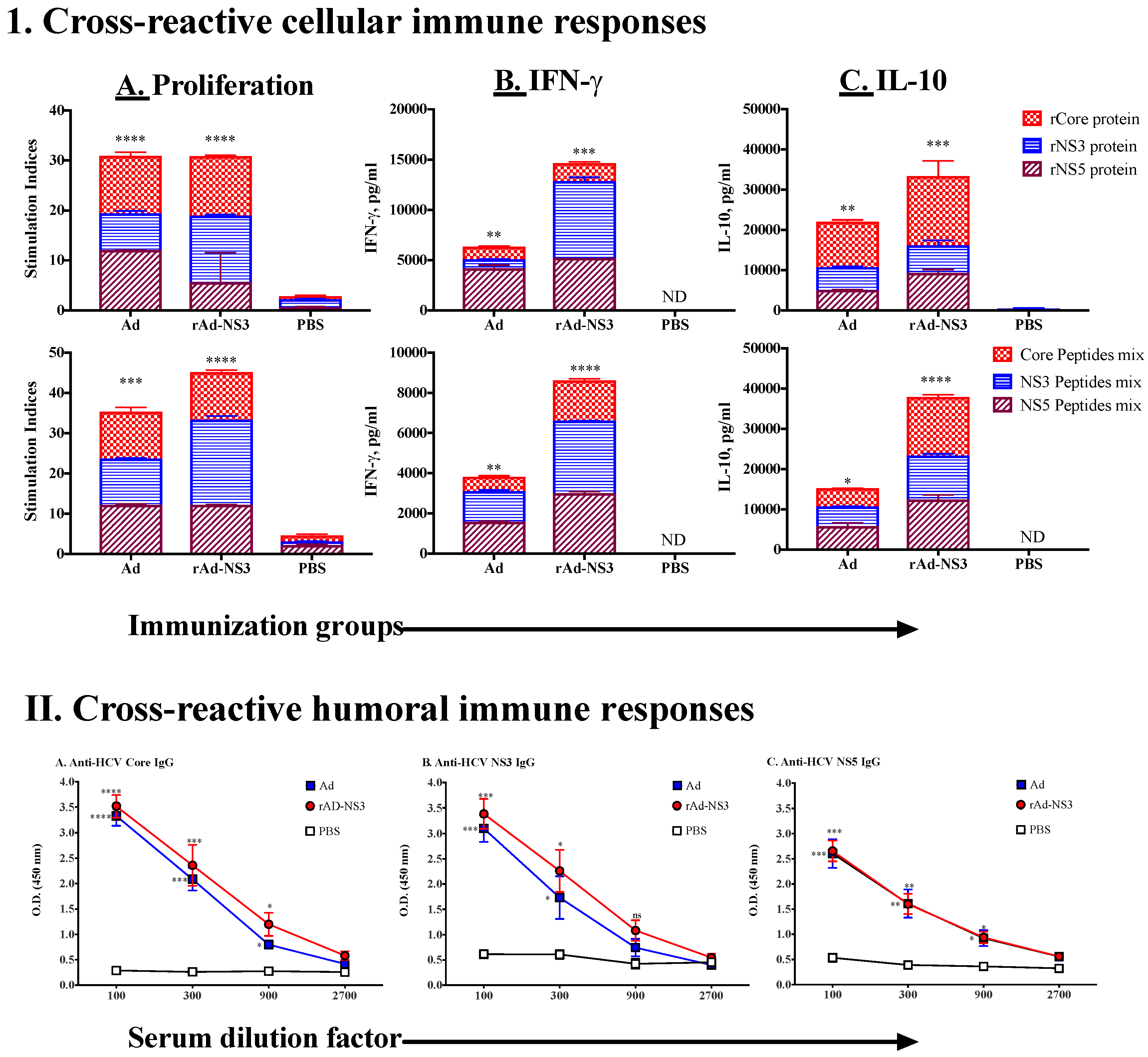
3.1. In Vivo Immune Cross-Reactivity between Ad and HCV
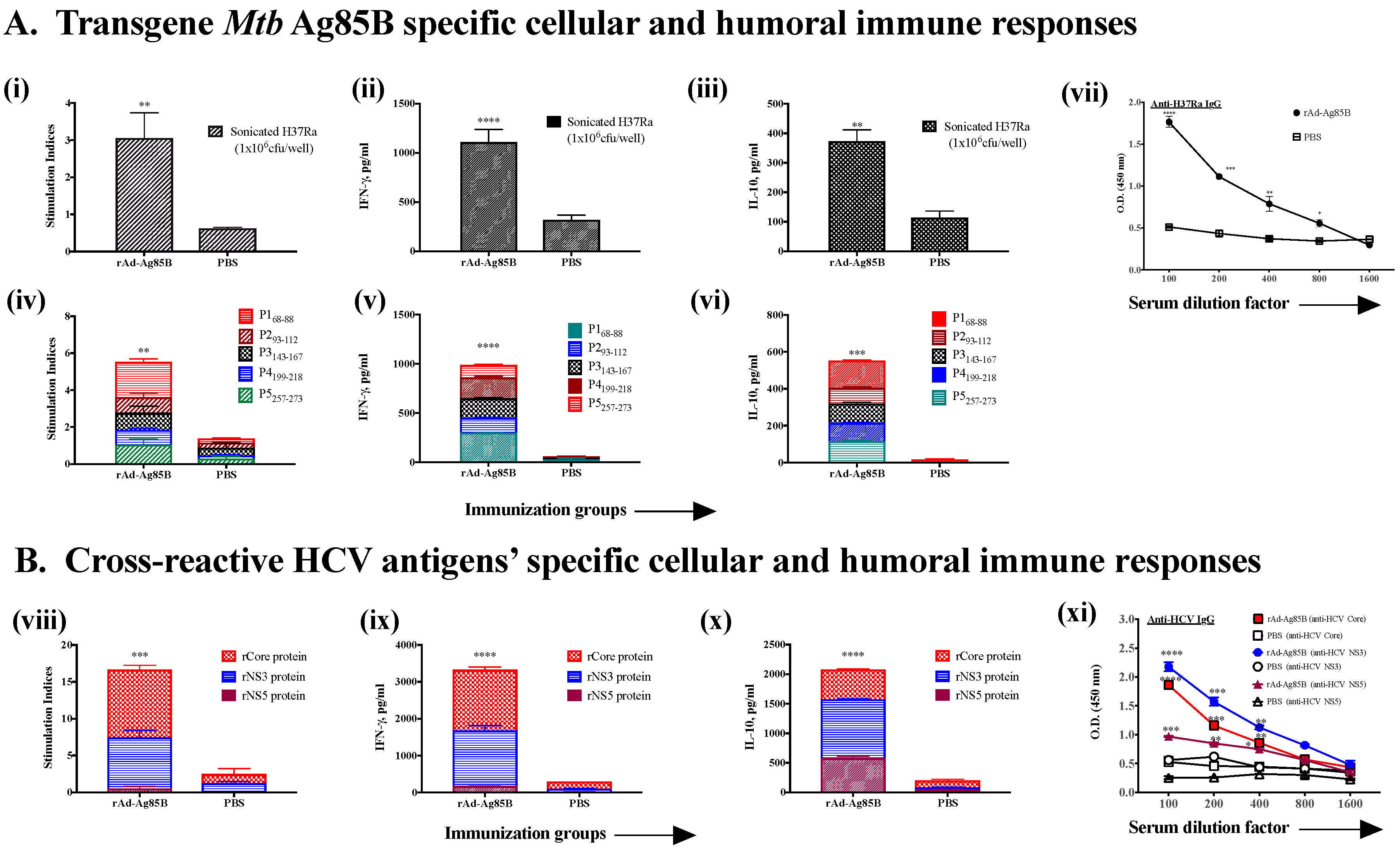
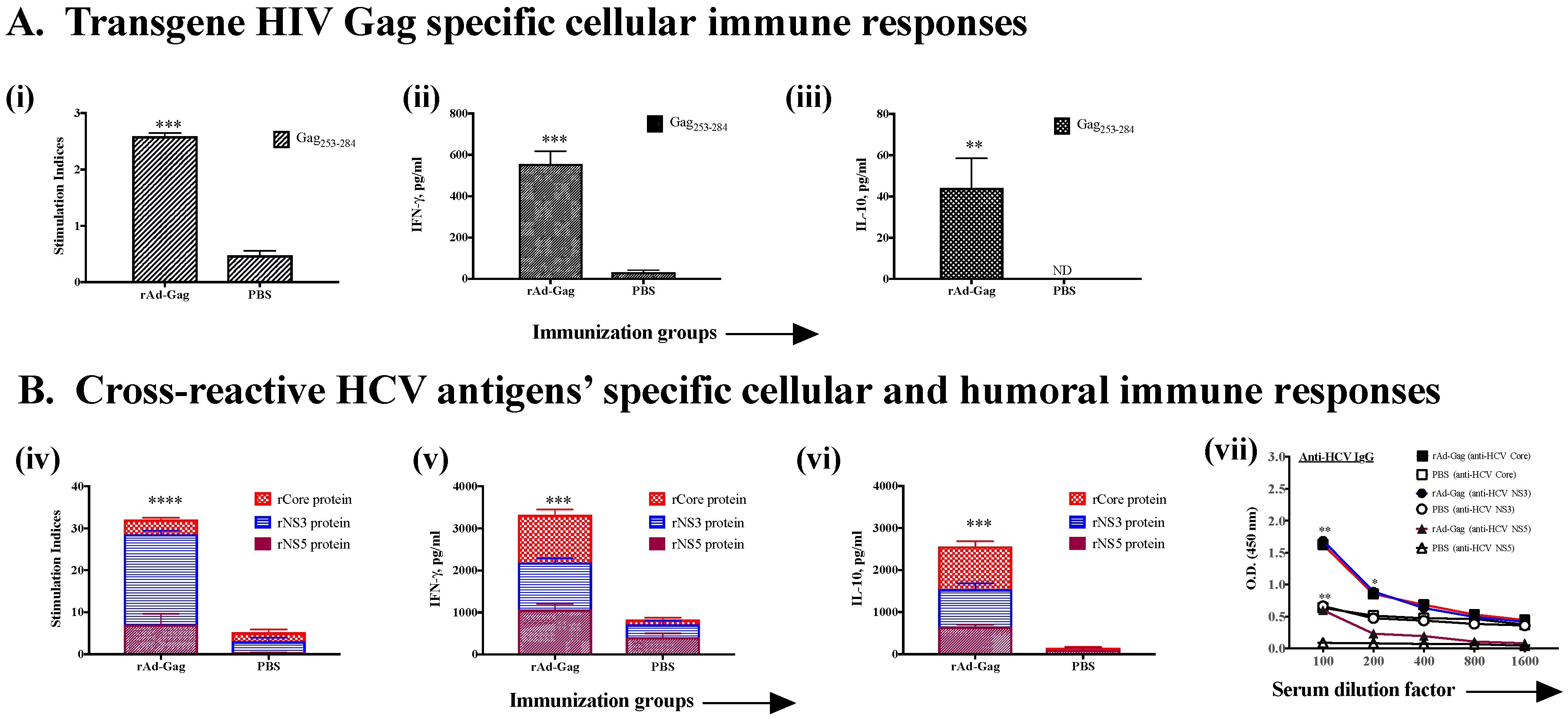
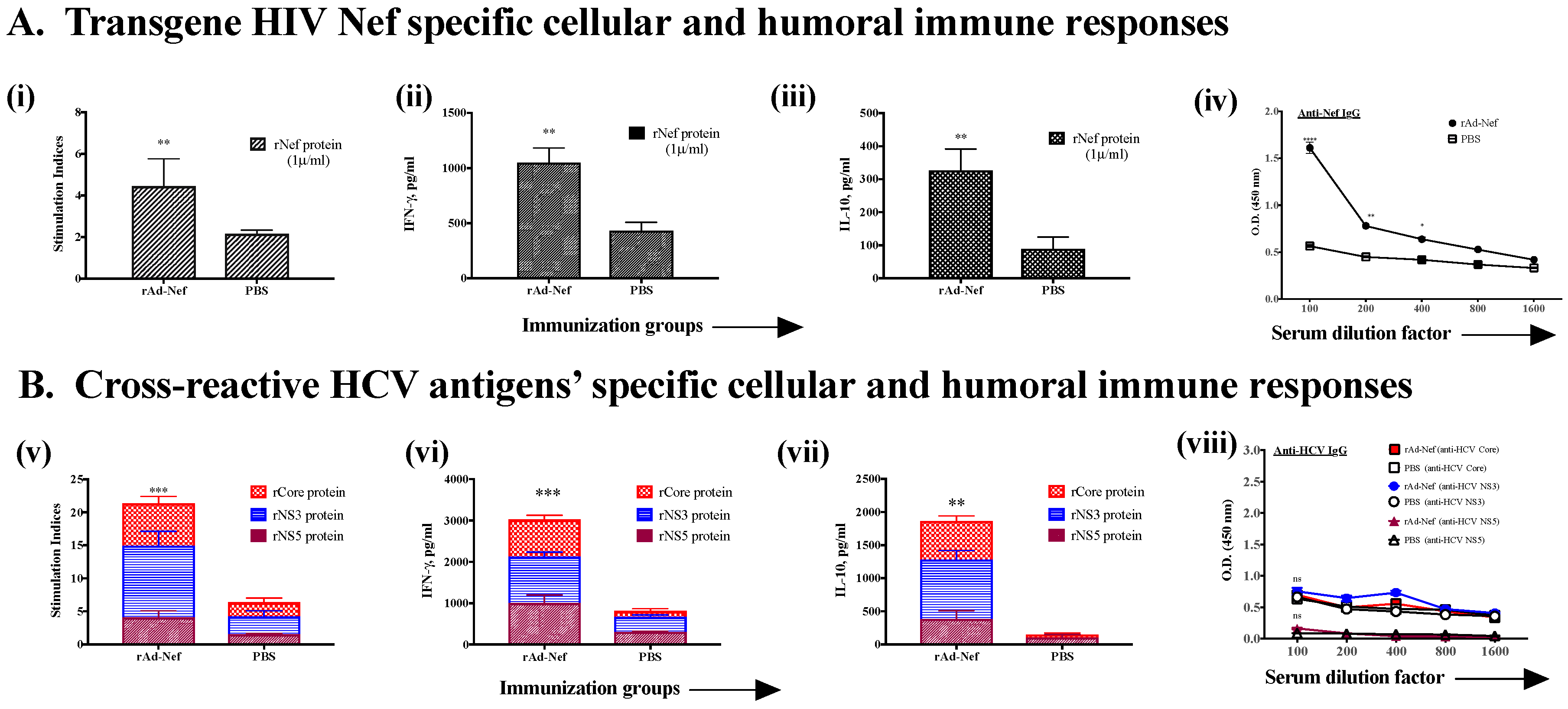
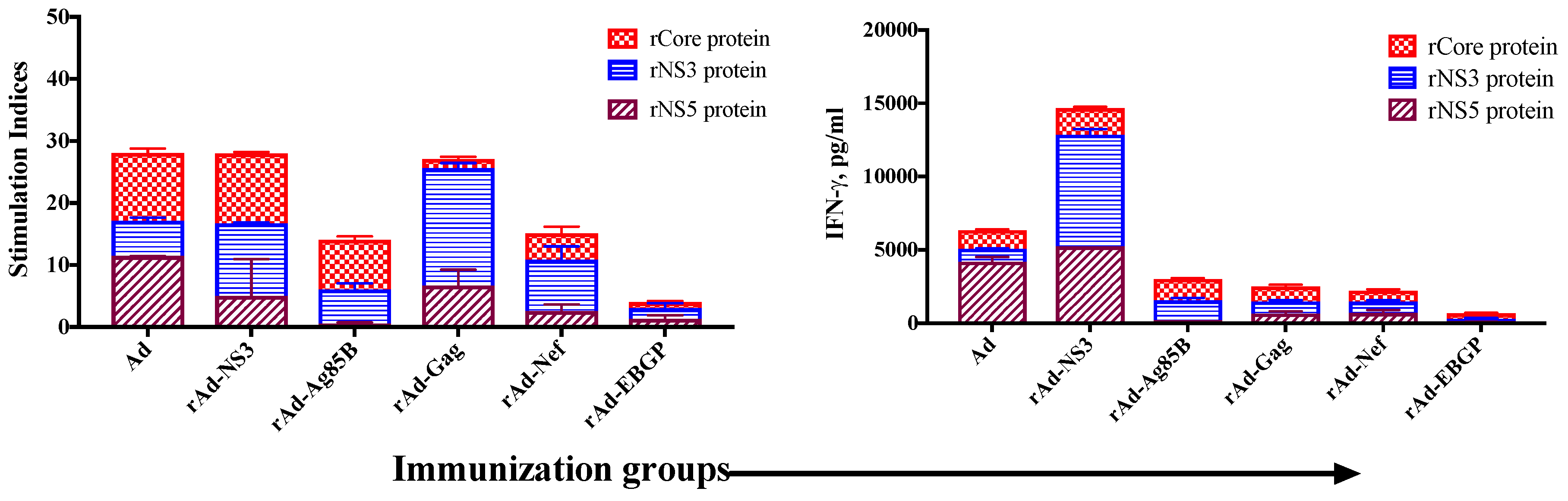
3.2. Induction of Cross-Reactive Humoral and Cellular Anti-HCV Immune Responses Induced upon Immunization with Recombinant Ad Vectors Individually Containing Antigens from HCV (NS3), Mtb (Ag85B), HIV (gag, Nef), and EBOV (GP)
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mohd, H.K.; Groeger, J.; Flaxman, A.D.; Wiersma, S.T. Global epidemiology of hepatitis C virus infection: New estimates of age-specific antibody to HCV seroprevalence. Hepatology 2013, 57, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.M.; Luciani, F.; Cameron, B.; Bull, R.A.; Beard, M.R.; Booth, D.; Lloyd, A.R. Resistance to hepatitis C virus: Potential genetic and immunological determinants. Lancet Infect. Dis. 2015, 15, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Hajarizadeh, B.; Grebely, J.; Dore, G.J. Epidemiology and natural history of HCV infection. Nat. Rev. Gastrpenterol. Hepatol. 2013, 10, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Bowen, D.G.; Walker, C.M. Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature 2005, 436, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Rehermann, B. Hepatitis C virus versus innate and adaptive immune responses: A tale of coevolution and co-existence. J. Clin. Investig. 2009, 119, 1745–1754. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Rehermann, B. Immune responses to HCV and other hepatitis viruses. Immunity 2014, 40, 13–24. [Google Scholar] [CrossRef]
- Ashfaq, U.A.; Javed, T.; Rehman, S.; Nawaz, Z.; Riazuddin, S. An overview of HCV molecular biology, replication and immune responses. Virol. J. 2011, 161. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Naglaa, H.S. Protective immunity against hepatitis C: Many shades of gray. Front. Immunol. 2014, 5, 274. [Google Scholar] [CrossRef]
- Mullis, C.E.; Laeyendecker, O.; Reynolds, S.J.; Ocama, P.; Quinn, J.; Boaz, I.; Gray, R.H.; Kirk, G.D.; Thomas, D.L.; Quinn, T.C.; Stabinski, L. High frequency of false-positive hepatitis C virus enzyme-linked immunosorbent assay in Rakai, Uganda. Clin. Infect. Dis. 2013, 57, 1747–1750. [Google Scholar] [CrossRef]
- Sakiani, S.; Koh, C.; Heller, T. Understanding the presence of false-positive antibodies in acute hepatitis. J. Infect. Dis. 2014, 210, 1886–1889. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.C.; Drobenuic, J.; Kamili, S. Prevalence of false-positive hepatitis C antibody results, National Health and Nutrition Examination Study (NHANES) 2007–2012. J. Clin. Virol. 2017, 89, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kesli, R. Evaluation of assay methods and false positive results in the laboratory diagnosis of hepatitis C virus infection. Arch. Clin. Microbiol. 2011, 2, 1–4. [Google Scholar]
- Srivastava, A.V.; Hrobowski, T.; Krese, L.; Huang, M.A.; Nemeh, H.; Tita, C.; Williams, C.; Brewer, R.; Lanfear, D.E. High rates of false-positive hepatitis c antibody tests can occur after left ventricular assist device implantation. ASAIO J. 2013, 59, 660–661. [Google Scholar] [CrossRef]
- Komitova, R.T.; Atanasova, M.V.; Pavlova, T.A.; Nyagolov, M.S.; Ivanova, A.V. Diagnostic Dilemmas in Hepatitis C Virus Infection for Hemodialysis Patients. Folia Med. (Plovdiv) 2017, 59, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Abdelwahab, S.F. Cellular immune response to hepatitis-C-virus in subjects without viremia or seroconversion: Is it important? Infect. Agent. Cancer 2016, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Schröter, M.; Feucht, H.H.; Schäfer, P.; Zöllner, B.; Polywka, S.; Laufs, R. Definition of false-positive reactions in screening for hepatitis C virus antibodies. J. Clin. Microbiol. 1999, 37, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Rivière, Y.; Montange, T.; Janvier, G.; Marnata, C.; Durrieu, L.; Chaix, M.L.; Isaguliants, M.; Launay, O.; Bresson, J.L.; Pol, S. Hepatitis C virus-specific cellular immune responses in individuals with no evidence of infection. Virol. J. 2012, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Bakshi, R.K.; Suneetha, P.V.; Fytili, P.; Antunes, D.A.; Vieira, G.F.; Jacobs, R.; Klade, C.S.; Manns, M.P.; Kraft, A.R.; et al. Frequency, Private Specificity, and Cross-Reactivity of Preexisting Hepatitis C Virus (HCV)-Specific CD8+ T Cells in HCV-Seronegative Individuals: Implications for Vaccine Responses. J. Virol. 2015, 89, 8304–8317. [Google Scholar] [CrossRef]
- Cornberg, M.; Wedemeyer, H. Hepatitis C virus infection from the perspective of heterologous immunity. Curr. Opin. Virol. 2016, 16, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Thomas, P.G. The two faces of heterologous immunity: Protection or immunopathology. J. Leukoc. Biol. 2014, 95, 405–416. [Google Scholar] [CrossRef]
- Welsh, R.M.; Fujinami, R.S. Pathogenic epitopes, heterologous immunity and vaccine design. Nat. Rev. Microbiol. 2007, 5, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Rehermann, B.; Shin, E.C. Private aspects of heterologous immunity. J. Exp. Med. 2005, 201, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Samrat, S.K.; Vedi, S.; Singh, S.; Li, W.; Kumar, R.; Agrawal, B. Immunization with Recombinant Adenoviral Vectors Expressing HCV Core or F Proteins Leads to T Cells with Reduced Effector Molecules Granzyme B and IFN-γ: A Potential New Strategy for Immune Evasion in HCV Infection. Viral. Immunol. 2015, 28, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Vedi, S.; Li, W.; Samrat, S.K.; Kumar, R.; Agrawal, B. Recombinant adenoviral vector expressing HCV NS4 induces protective immune responses in a mouse model of Vaccinia-HCV virus infection: A dose and route conundrum. Vaccine 2014, 32, 2712–2721. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, J.; Tyrrell, D.L.J.; Agrawal, B. Expression of hepatitic C virus-derived core or NS3 antigens in human dendritic cells leads to induction of pro-inflammatory cytokines and normal T-cell stimulation capabilities. J. Gen. Virol. 2006, 87, 61–72. [Google Scholar] [CrossRef]
- Krishnadas, D.K.; Li, W.; Kumar, R.; Tyrrell, D.L.; Agrawal, B. HCV-core and NS3 antigens play disparate role in inducing regulatory or effector T cells in vivo: Implications for viral persistence or clearance. Vaccine 2010, 28, 2104–2114. [Google Scholar] [CrossRef] [PubMed]
- Krishnadas, D.K.; Li, W.; Kumar, R.; Tyrrell, L.J.; Agrawal, B. In vitro activation and differentiation of naïve CD4+and CD8+T cells into HCV Core- and NS3-specific armed effector cells: A new role for CD4+T cells. Cell Immunol. 2009, 259, 141–149. [Google Scholar] [CrossRef]
- Singh, S.; Vedi, S.; Samrat, S.K.; Li, W.; Kumar, R.; Agrawal, B. Heterologous immunity between adenoviruses and hepatitis C virus: A new paradigm in HCV immunity and vaccines. PLoS ONE 2016, 11, 1–23. [Google Scholar] [CrossRef]
- Agrawal, B.; Singh, S.; Gupta, N.; Li, W.; Vedi, S.; Kumar, R. Unsolved puzzles surrounding HCV immunity: Heterologous immunity adds another dimension. Int. J. Mol. Sci. 2017, 18, 1626. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.; Richardson, J.S.; Cutts, T.; Qiu, X.; Kobinger, G.P. Intranasal immunization with an adenovirus vaccine protects guinea pigs from Ebola virus transmission by infected animals. Antiviral Res. 2015, 116, 17–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, R.M.; Che, J.W.; Brehm, M.A.; Selin, L.K. Heterologous immunity between viruses. Immunol. Rev. 2010, 235, 244–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Fries, R.; Mitsuhashi, M. Quantification of mitogen induced human lymphocyte proliferation: Comparison of alamarBlue assay to 3H-thymidine incorporation assay. J. Clin. Lab. Anal. 1995, 9, 89–95. [Google Scholar] [CrossRef]
- Waritani, T.; Chang, J.; McKinney, B.; Terato, K. An ELISA protocol to improve the accuracy and reliability of serological antibody assays. MethodsX 2017, 4, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.W.; Williams, A.; Okkels, L.M.; Hatch, G.; Andersen, P. Protective effect of a tuberculosis subunit vaccine based on a fusion of antigen 85B and ESAT-6 in the aerosol guinea pig model. Infect. Immun. 2004, 72, 6148–6150. [Google Scholar] [CrossRef] [PubMed]
- Wiker, H.G.; Harboe, M. The antigen 85 complex: A major secretion product of Mycobacterium tuberculosis. Microbiol. Mol. Biol. Rev. 1992, 56, 648–661. [Google Scholar] [CrossRef]
- Pardini, M.; Giannoni, F.; Palma, C.; Iona, E.; Cafaro, A.; Brunori, L.; Rinaldi, M.; Fazio, V.M.; Laguardia, M.E.; Carbonella, D.C.; et al. Immune response and protection by DNA vaccines expressing antigen 85B of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 2006, 262, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Meerak, J.; Wanichwecharungruang, S.P.; Palaga, T. Enhancement of immune response to a DNA vaccine against Mycobacterium tuberculosis Ag85B by incorporation of an autophagy inducing system. Vaccine 2013, 31, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Axelsson-Robertson, R.; Magalhaes, I.; Parida, S.K.; Zumla, A.; Maeurer, M. The immunological footprint of mycobacterium tuberculosis t-cell epitope recognition. J. Infect. Dis. 2012, 15, 205. [Google Scholar] [CrossRef]
- Castelli, F.A.; Szely, N.; Maillère, B. P17-18. ANRS lipo5 sequences induce in vitro cross-reactive CD4+ T cell response against clade B and C. Retrovirology 2009, 6, P300. [Google Scholar] [CrossRef]
- Barouch, D.H. Novel adenovirus vector-based vaccines for HIV-1. Curr. Opin. HIV AIDS 2010, 5, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Picker, L.J. Novel vaccine vectors for HIV.1. Nat. Rev. Microbiol. 2014, 12, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Jiang, Y.; Gong, T.; Zhang, Z.; Sun, X. Intranasal Vaccination against HIV-1 with Adenoviral Vector-Based Nanocomplex Using Synthetic TLR-4 Agonist Peptide as Adjuvant. Mol. Pharm. 2016, 13, 885–894. [Google Scholar] [CrossRef]
- Dixon, M.G.; Schafer, I.J. Ebola viral disease outbreak - West Africa, 2014. Ann. Emerg. Med. 2015, 65, 1101–1108. [Google Scholar] [CrossRef]
- Richardson, J.S.; Pillet, S.; Bello, A.J.; Kobinger, G.P. Airway Delivery of an Adenovirus-Based Ebola Virus Vaccine Bypasses Existing Immunity to Homologous Adenovirus in Nonhuman Primates. J. Virol. 2013, 87, 3668–3677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusch, E.; Renz, H.; Skevaki, C. Respiratory virus-induced heterologous immunity: Part of the problem or part of the solution? Allergo. J. 2018, 27, 28–45. [Google Scholar] [CrossRef]
- Mast, T.C.; Kierstead, L.; Gupta, S.B.; Nikas, A.A.; Kallas, E.G.; Novitsky, V.; Mbewe, B.; Pitisuttithum, P.; Schechter, M.; Vardas, E.; et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: Correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010, 28, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Benkö, M.; Harrach, B. Molecular evolution of adenoviruses. Curr. Top Microbiol. Immunol. 2003, 272, 3–35. [Google Scholar] [CrossRef]
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-existing immunity against Ad vectors: Humoral, cellular, and innate response, what’s important? Hum. Vaccin. Immunother. 2014, 10, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.P.; Eichholz, K.; Amelio, P.; Moyer, C.; Nemerow, G.R.; Perreau, M.; Mennechet, F.J.D.; Kremer, E.J. Humoral immune response to adenovirus induce tolerogenic bystander dendritic cells that promote generation of regulatory T cells. PLoS Pathog. 2018, 14, e1007127. [Google Scholar] [CrossRef]
- Sewell, A.K. Why must T cells be cross-reactive? Nat. Rev. Immunol. 2012, 12, 669. [Google Scholar] [CrossRef]
- Welsch, R.M.; Selin, L.K. No one is naïve: Significance of heterologous T cell immunity. Nat. Rev. Immunol. 2002, 2, 417–426. [Google Scholar] [CrossRef]
- Petrova, G.; Ferrante, A.; Gorski, J. Cross-reactivity of T cells and its role in the immune system. Crit. Rev. Immunol. 2012, 32, 349–372. [Google Scholar] [CrossRef]
- Yin, Y.; Mariuzza, R.A. The multiple mechanisms of T cell cross-reactivity. Immunity 2009, 31, 849–851. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.R.; Borbulevych, O.Y.; Piepenbrink, K.H.; Corcelli, S.A.; Baker, B.M. Disparate degrees of hypervariable loop flexibility control T cell receptor cross-reativity, specificity and binding mechanisms. J. Mol. Biol. 2011, 414, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Bulteel, N.; Partha Sarathy, P.; Forrest, E.; Stanley, A.J.; Innes, H.; Mills, P.R.; Valerio, H.; Gunson, R.N.; Aitken, C.; Morris, J.; et al. Factors associated with spontaneous clearance of chronic hepatitis C virus infection. J. Hepatol. 2016, 65, 266–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.D.; McMahon, B.J.; Bruden, D.; Sullivan, D.; Homan, C.; Christensen, C.; Gretch, D.R. High rate of spontaneous negativity for hepatitis C virus RNA after establishment of chronic infection in Alaska Natives. Clin. Infect. Dis. 2006, 42, 945–952. [Google Scholar] [CrossRef]
- Raghuraman, S.; Park, H.; Osburn, W.O.; Winkelstein, E.; Edlin, B.R.; Rehermann, B. Spontaneous clearance of chronic hepatitis C virus infection is associated with appearance of neutralizing antibodies and reversal of T-cell exhaustion. J. Infect. Dis. 2012, 205, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Krishnadas, D.K.; Kumar, R.; Tyrrell, D.L.; Agrawal, B. Priming and stimulation of hepatitis C virus-specific CD4+ and CD8+ T cells against HCV antigens NS4, NS5a or NS5b from HCV-naive individuals: Implications for prophylactic vaccine. Int. Immunol. 2008, 20, 89–104. [Google Scholar] [CrossRef]
- Kennedy, P.T.F.; Urbani, S.; Moses, R.A.; Amadei, B.; Fisicaro, P.; Lloyd, J.; Maini, M.K.; Dusheiko, G.; Ferrari, C.; Bertoletti, A. The influence of T cell cross-reactivity on HCV-peptide specific human T cell response. Hepatology 2006, 43, 602–611. [Google Scholar] [CrossRef]
- Castelli, F.A.; Leleu, M.; Pouvelle-Moratille, S.; Farci, S.; Zarour, H.M.; Andrieu, M.; Auriault, C.; Ménez, A.; Georges, B.; Maillere, B. Differential capacity of T cell priming in naive donors of promiscuous CD4+ T cell epitopes of HCV NS3 and Core proteins. Eur. J. Immunol. 2007, 37, 1513–1523. [Google Scholar] [CrossRef]
- Hutnick, N.A.; Carnathan, D.; Demers, K.; Makedonas, G.; Ertl, H.C.; Betts, M.R. Adenovirus-specific human T cells are pervasive, polyfunctional, and cross-reactive. Vaccine 2010, 28, 1932–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Protein | Peptide # | Location | Amino Acid Sequence |
|---|---|---|---|---|
| HCV | Core | 5 | 17–31 | RRPQDVKFPGGGQIV |
| 14 | 53–67 | SERSQPRGRRQPIPK | ||
| 16 | 61–75 | RRQPIPKARRPEGRT | ||
| 17 | 65–79 | IPKARRPEGRTWAQP | ||
| 27 | 105–119 | PSWGPTDPRRRSRNL | ||
| NS3 | 2 | 1367–1381 | LSTTGEIPFYGKAIP | |
| 5 | 1450–1464 | SVIDCNTCVTQTVDF | ||
| 6 | 1127–1142 | SSDLYLVTRHADVIP | ||
| 8 | 1467–1482 | RRGRTGRGKPGIYRF | ||
| 10 | 1607–1622 | MWKCLIRLKPTLHGP | ||
| NS5a | 6 | 2047–2066 | VGPRTCRNMWSGTFPINAYT | |
| 24 | 2317–2336 | PPPRSPPVPPPRKKRTVVLT | ||
| NS5b | 5 | 2481–2500 | DSHYQDVLKEVKAAASKVKA | |
| 19 | 2691–2710 | GENCGYRRCRASGVLTTSCG | ||
| 27 | 2811–2830 | PLARAAWETARHTPVNSWLG | ||
| Mtb | Ag85B | P1 | 68–88 | SGGNNSPAVYLLDGLRAQDDY |
| P2 | 93–112 | INTPAFEWYYQSGLSIVMPV | ||
| P3 | 143–167 | SELPQWLSANRAVKPTGSAAIGLSM | ||
| P4 | 199–218 | MGPSLIGLAMGDAGGYKAAD | ||
| P5 | 257–273 | NELGGANIPAEFLENFV | ||
| HIV | Gag | Gag253-284 | 253–284 | NPPIPVGEIYKRWIILGLNKIVRMYSPTSILD |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrawal, B.; Gupta, N.; Vedi, S.; Singh, S.; Li, W.; Garg, S.; Li, J.; Kumar, R. Heterologous Immunity between Adenoviruses and Hepatitis C Virus (HCV): Recombinant Adenovirus Vaccine Vectors Containing Antigens from Unrelated Pathogens Induce Cross-Reactive Immunity Against HCV Antigens. Cells 2019, 8, 507. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050507
Agrawal B, Gupta N, Vedi S, Singh S, Li W, Garg S, Li J, Kumar R. Heterologous Immunity between Adenoviruses and Hepatitis C Virus (HCV): Recombinant Adenovirus Vaccine Vectors Containing Antigens from Unrelated Pathogens Induce Cross-Reactive Immunity Against HCV Antigens. Cells. 2019; 8(5):507. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050507
Chicago/Turabian StyleAgrawal, Babita, Nancy Gupta, Satish Vedi, Shakti Singh, Wen Li, Saurabh Garg, Jie Li, and Rakesh Kumar. 2019. "Heterologous Immunity between Adenoviruses and Hepatitis C Virus (HCV): Recombinant Adenovirus Vaccine Vectors Containing Antigens from Unrelated Pathogens Induce Cross-Reactive Immunity Against HCV Antigens" Cells 8, no. 5: 507. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8050507