Protective Effects of Licochalcone A Improve Airway Hyper-Responsiveness and Oxidative Stress in a Mouse Model of Asthma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sensitization and Administration of Licochalcone A
2.3. Airway Hyper-Responsiveness (AHR)
2.4. Bronchoalveolar Lavage Fluid (BALF) and Cell Counting
2.5. Histological Analysis of Lung Tissue
2.6. Immunofluorescence Staining
2.7. Glutathione (GSH) Assay
2.8. Malondialdehyde (MDA) Activity
2.9. RNA Isolation and Quantitative Real-Time PCR Analysis
2.10. Western Immunoblot Analysis
2.11. Serum Collection
2.12. ELISA
2.13. Biochemical Analysis of Serum
2.14. BEAS-2B Cell Culture and Licochalcone A Treatment
2.15. Determination of ROS Production
2.16. Cell–Cell Adhesion Assay
2.17. Statistical Analysis
3. Results
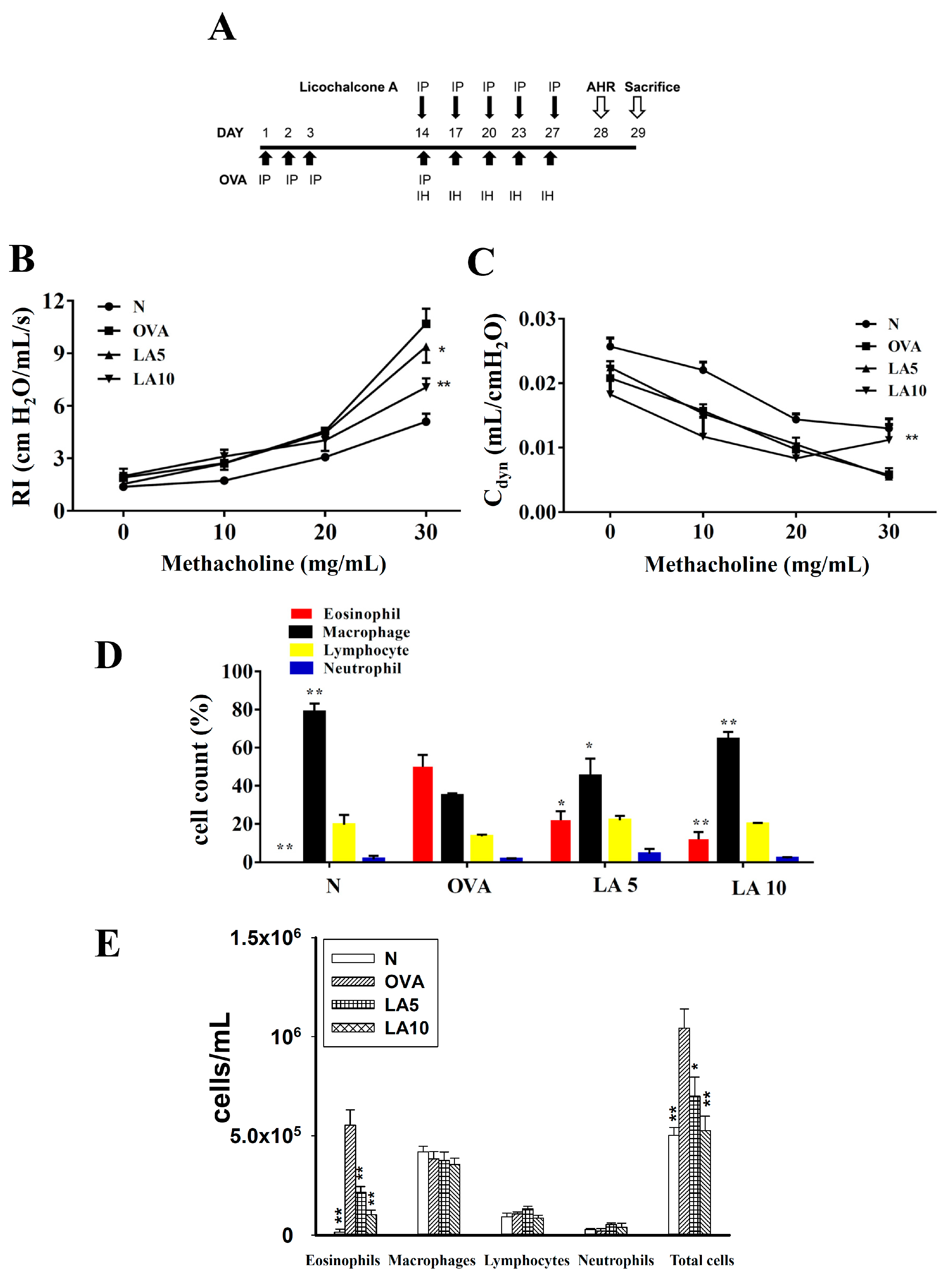
3.1. Licochalcone A Attenuated AHR in Asthmatic Mice
3.2. Licochalcone A Reduced Eosinophils in BALF
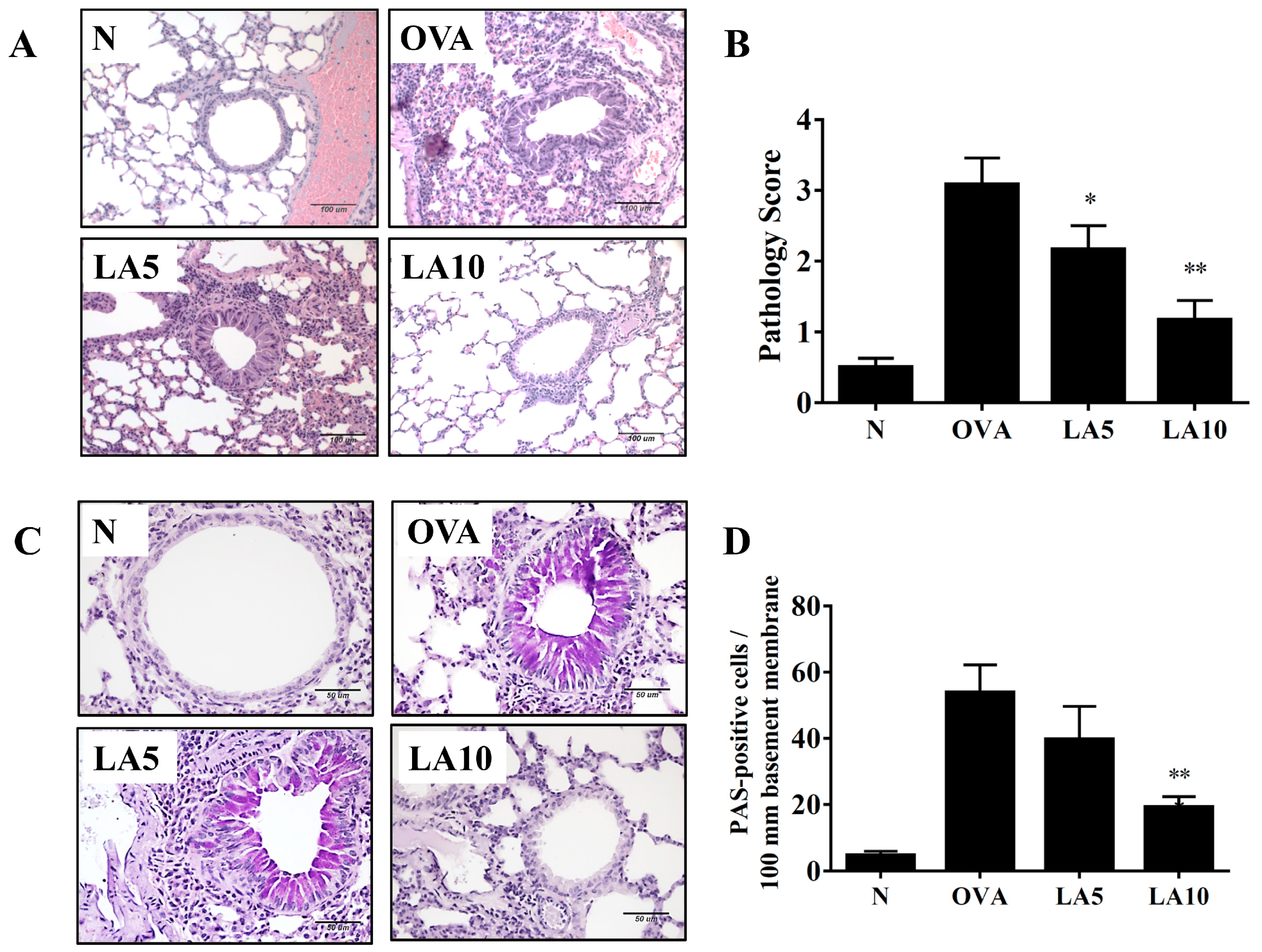
3.3. Licochalcone A Suppressed Eosinophil Infiltration and Goblet Cell Hyperplasia in the Lungs
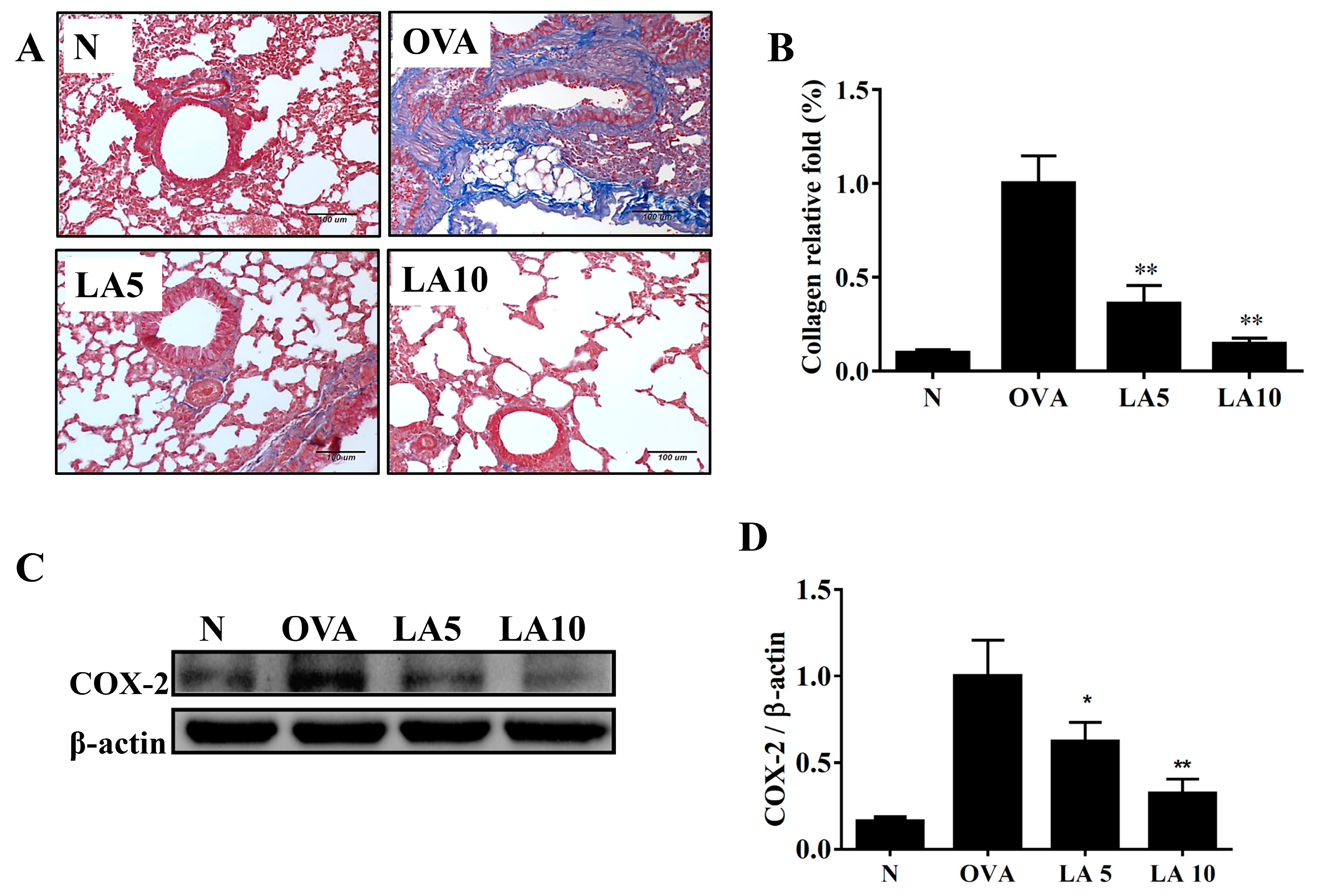
3.4. Licochalcone A Reduced Collagen and COX-2 Expression in the Lungs
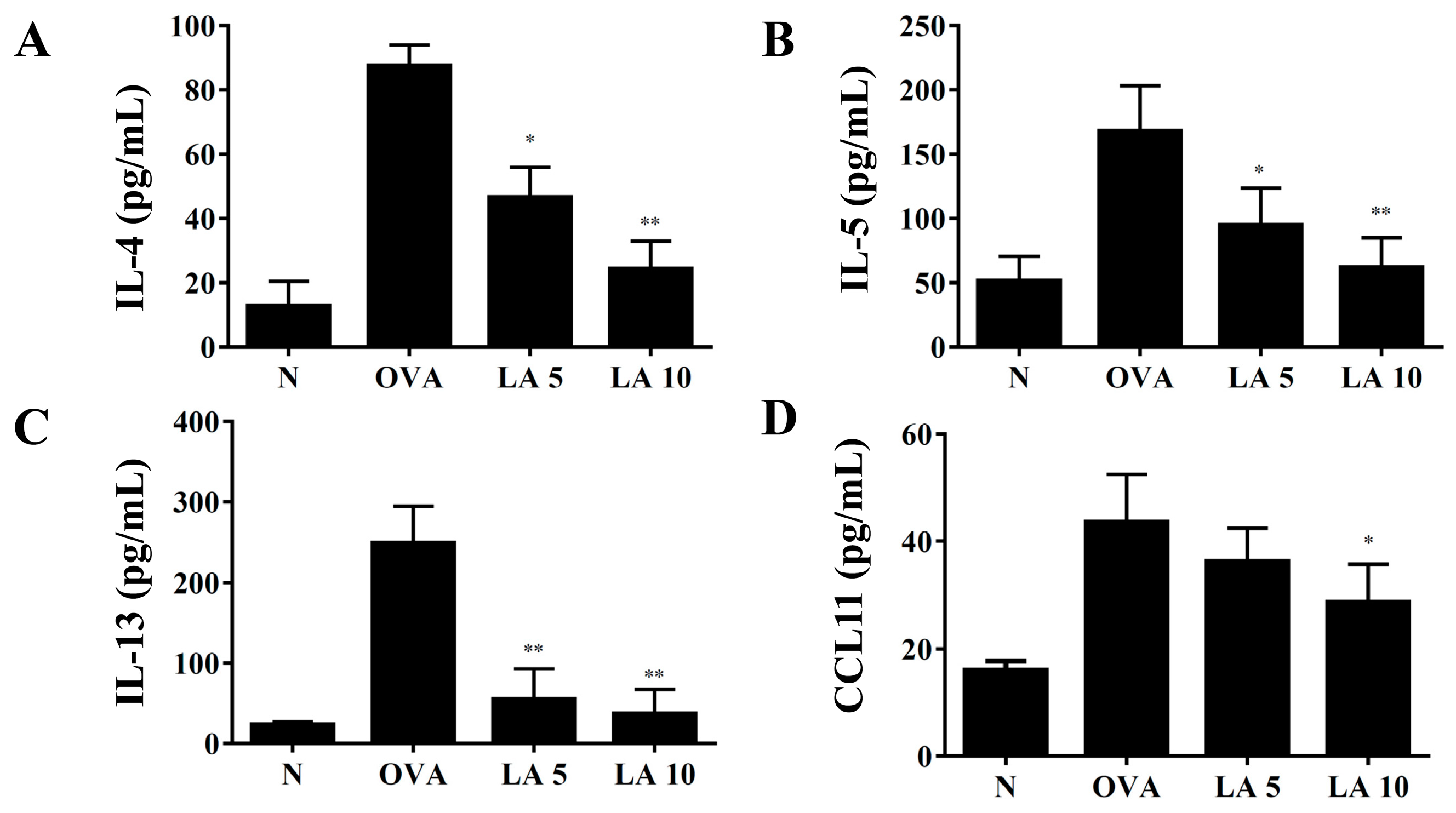
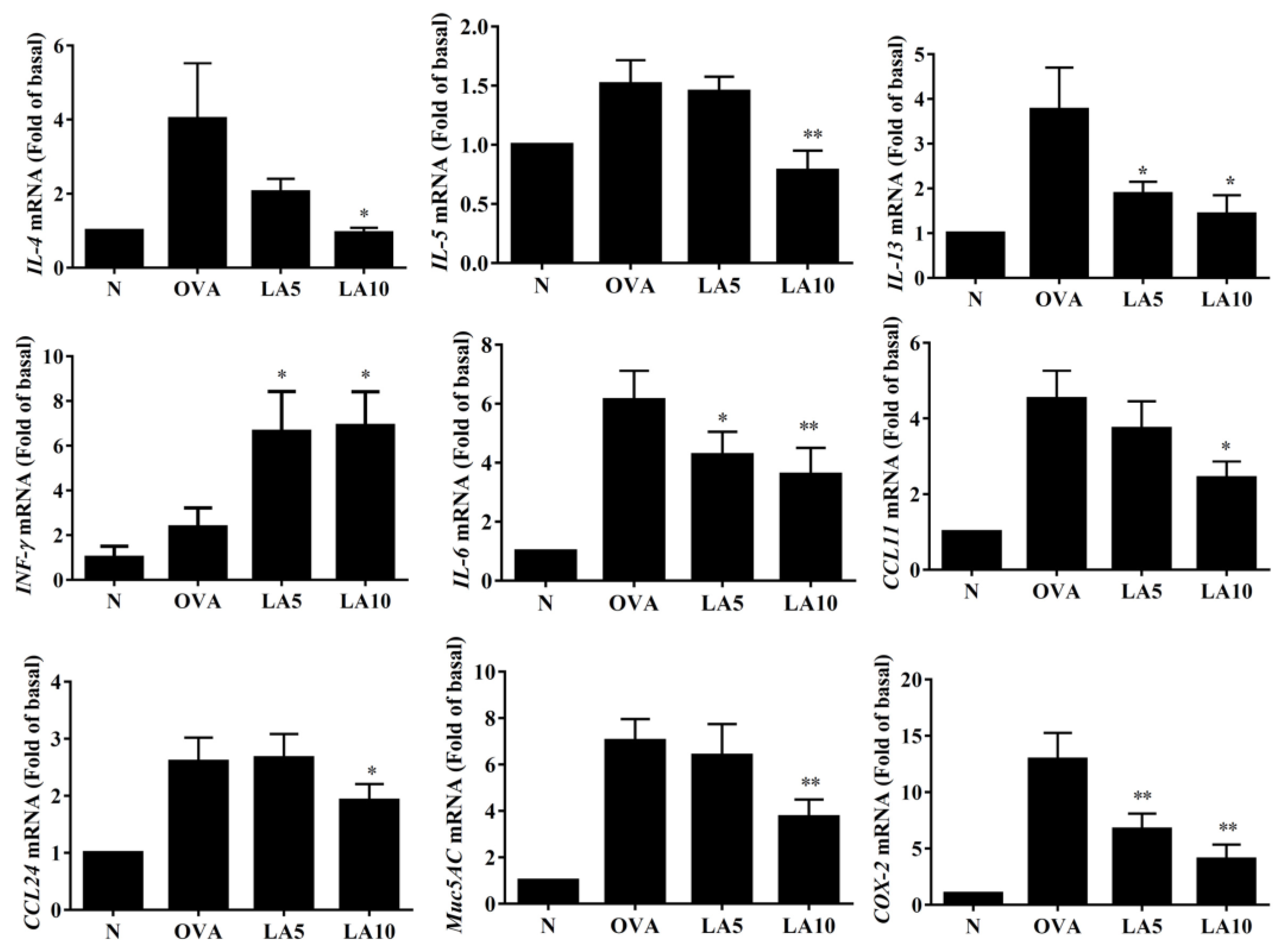
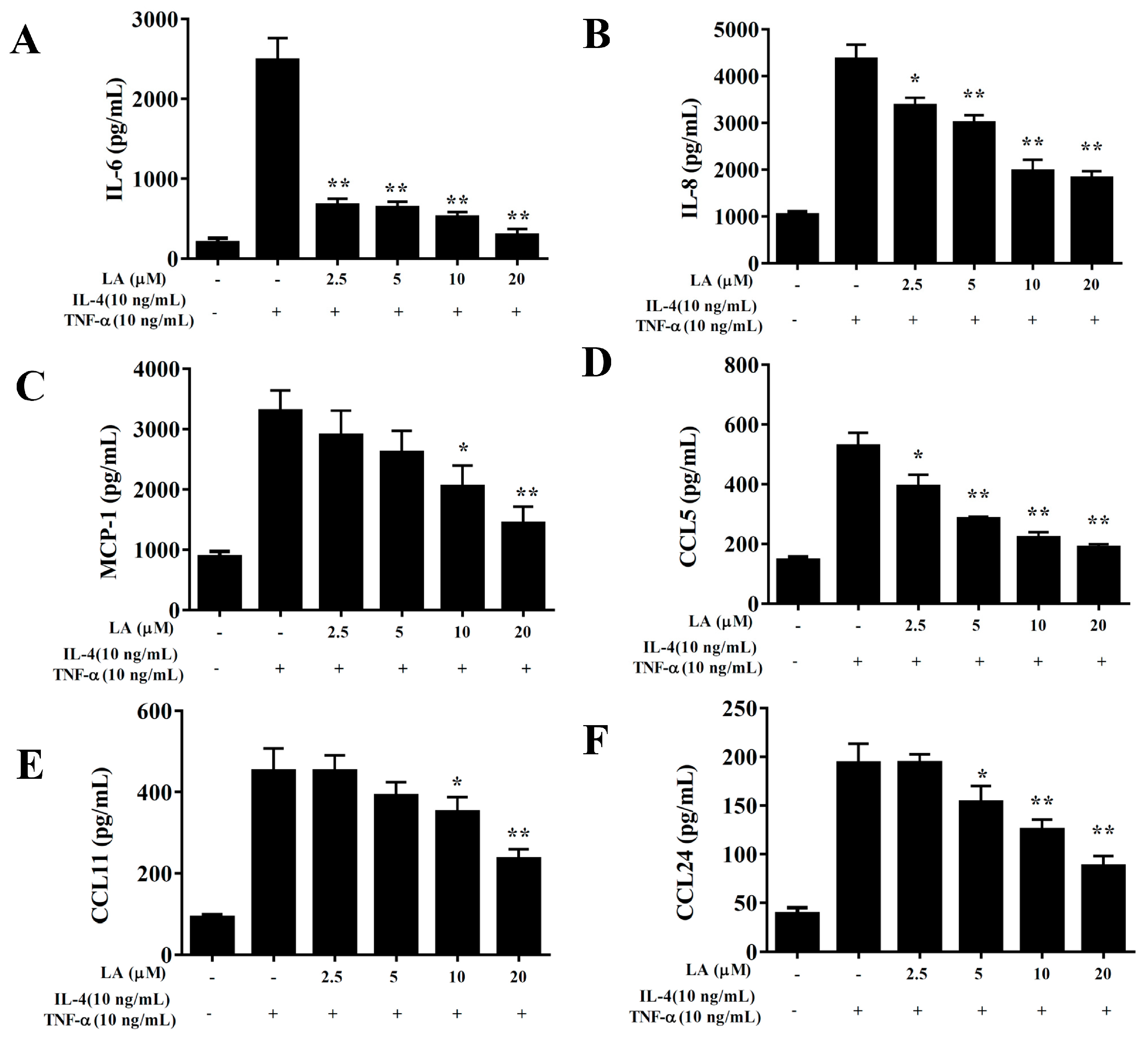
3.5. Licochalcone A Modulated Cytokine and Chemokine Levels in Lung Tissue and BALF
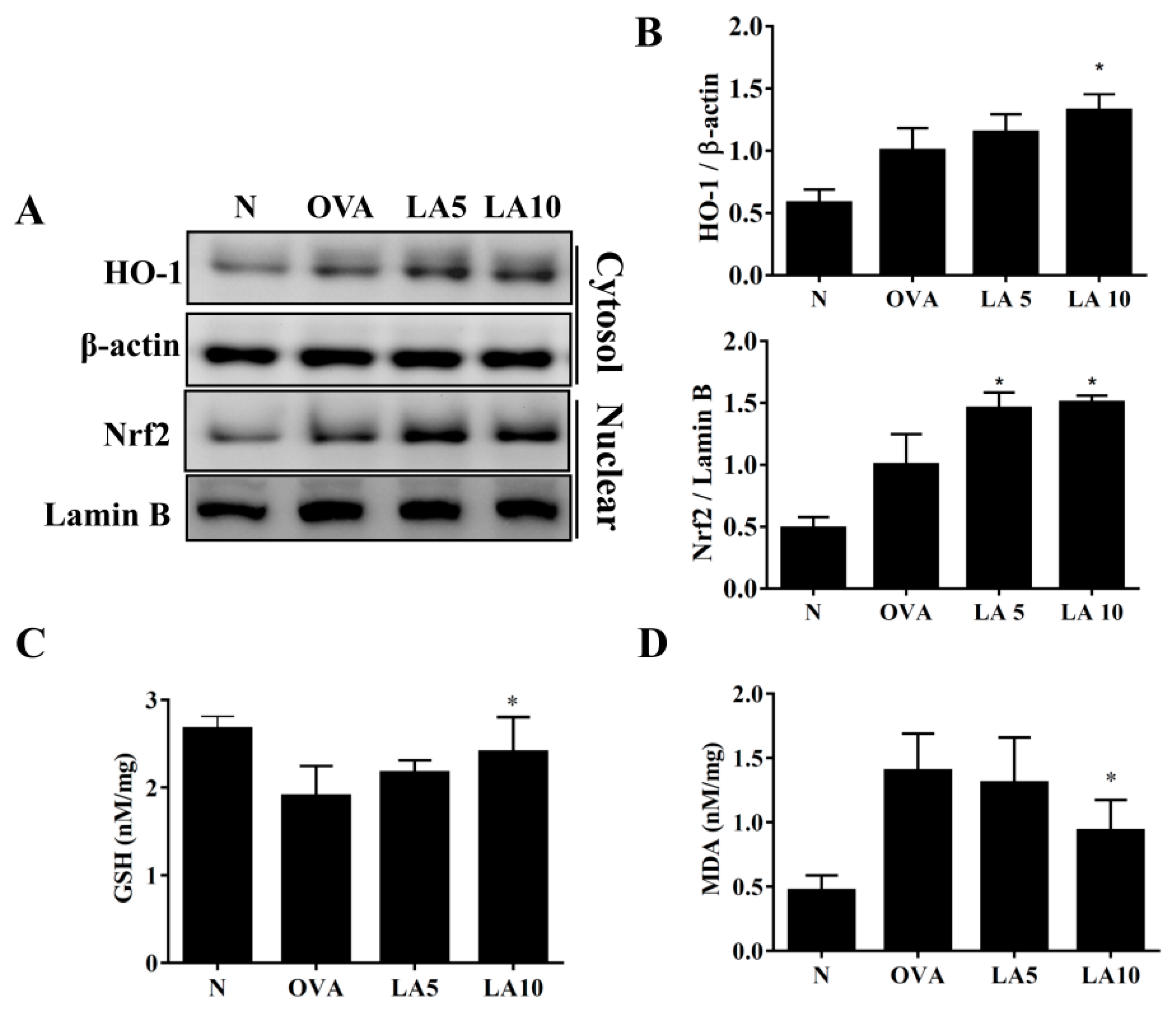
3.6. Licochalcone A Modulated GSH and MDA Activity in the Lungs
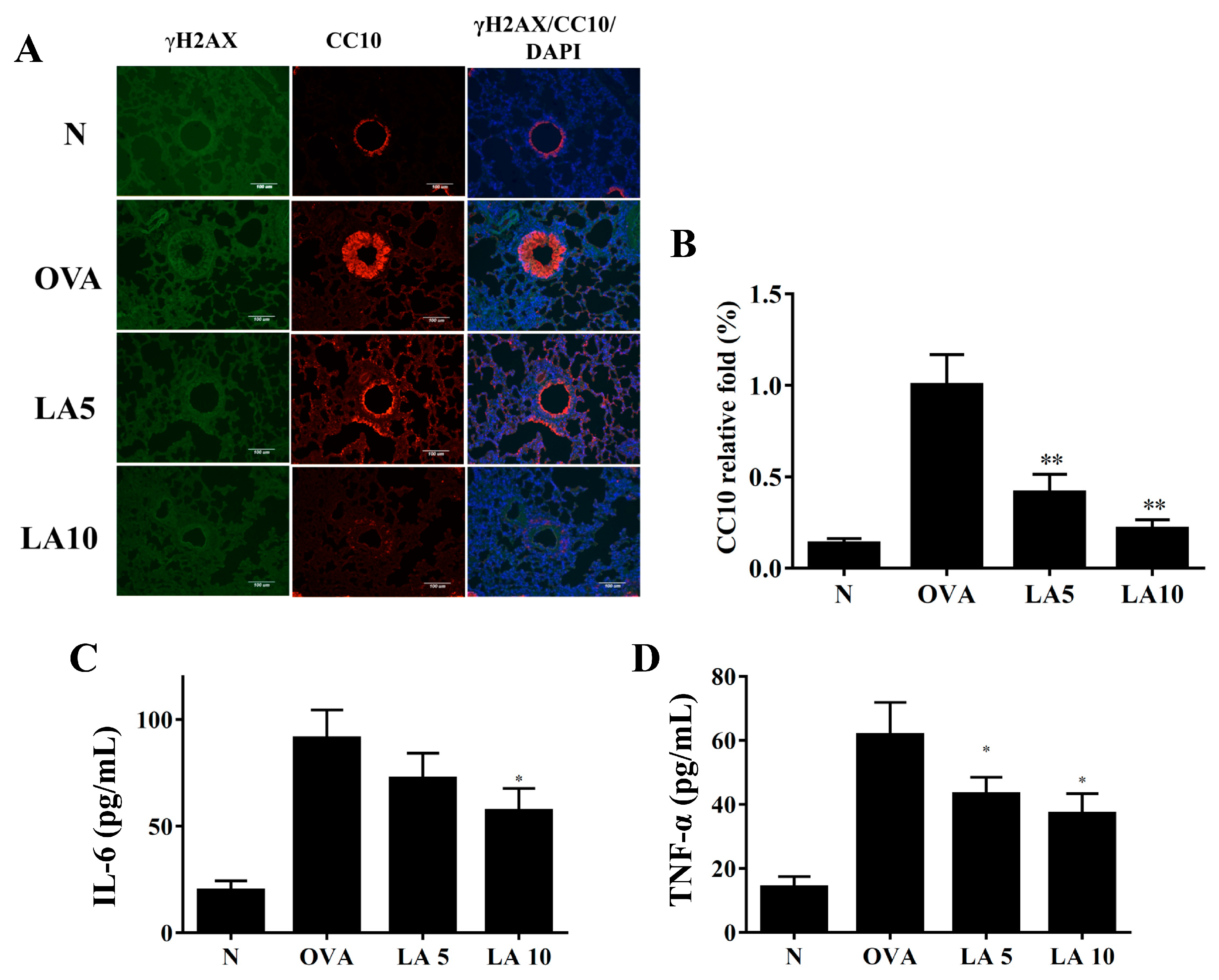
3.7. Licochalcone A Did Not Improve DNA Damage in Ovalbumin (OVA)-Sensitized Asthmatic Mice
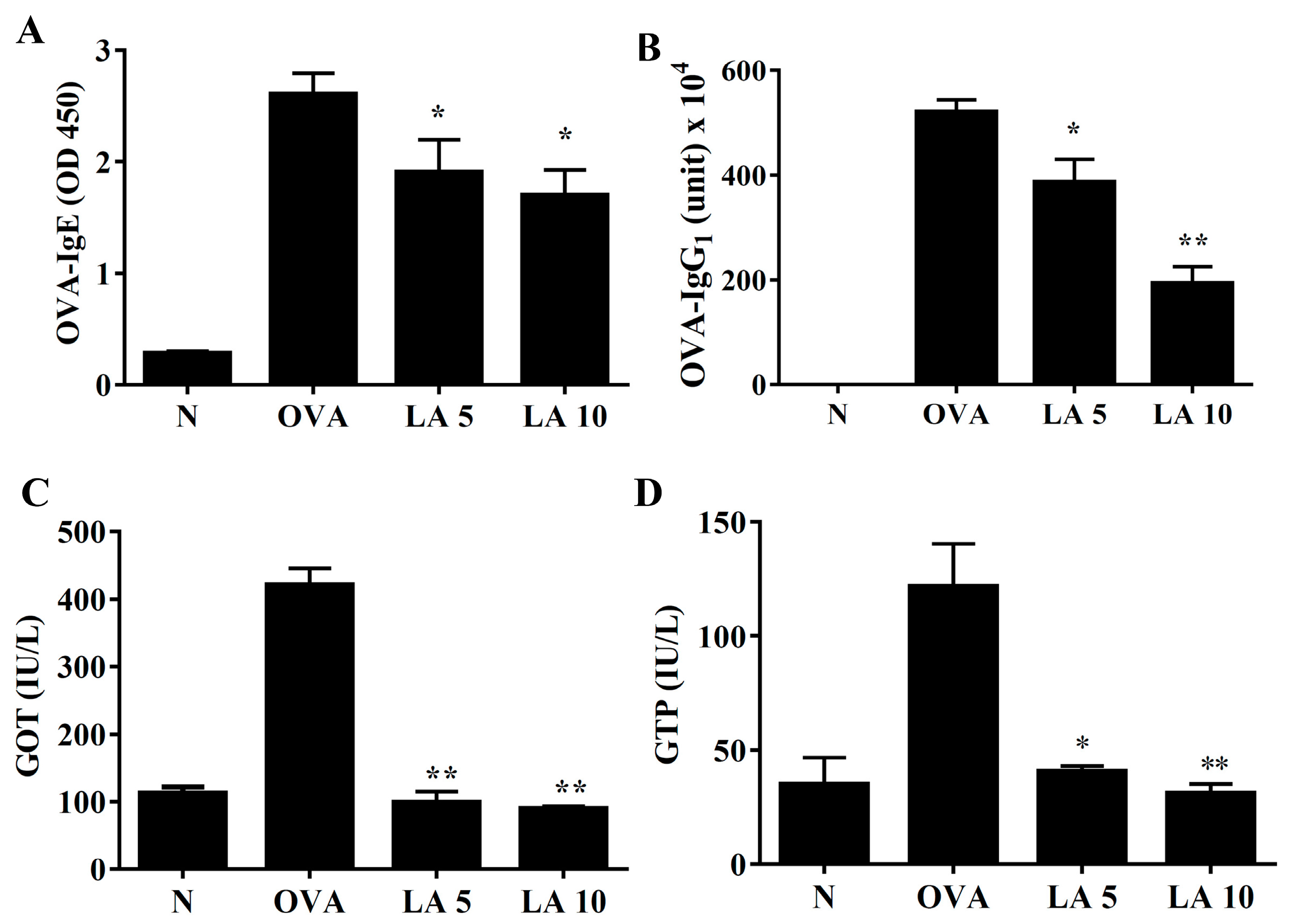
3.8. Licochalcone A Modulated Serum OVA-Specific Antibody
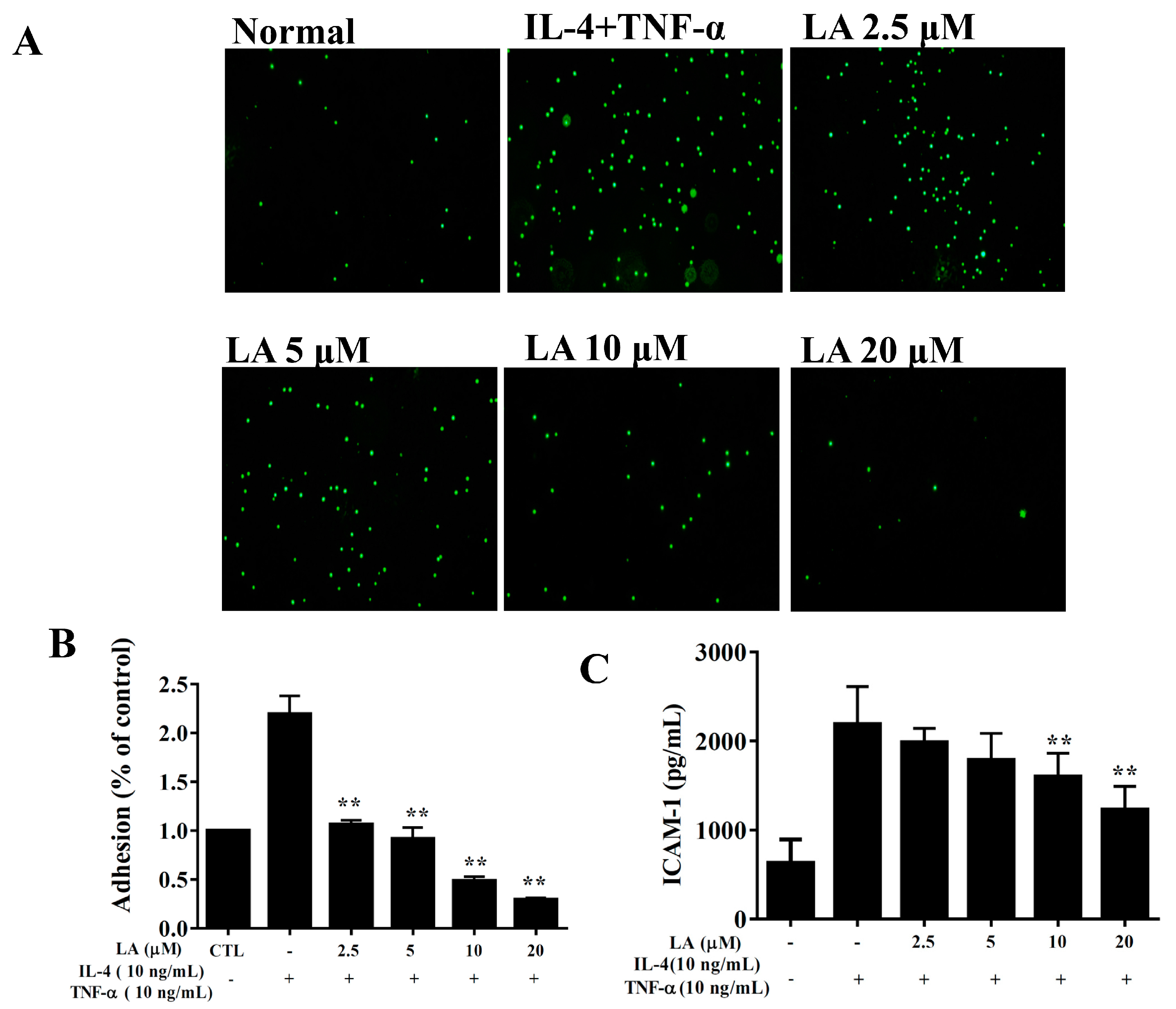
3.9. Licochalcone A Suppressed Inflammatory Mediators and Cell Adhesion in BEAS-2B Cells
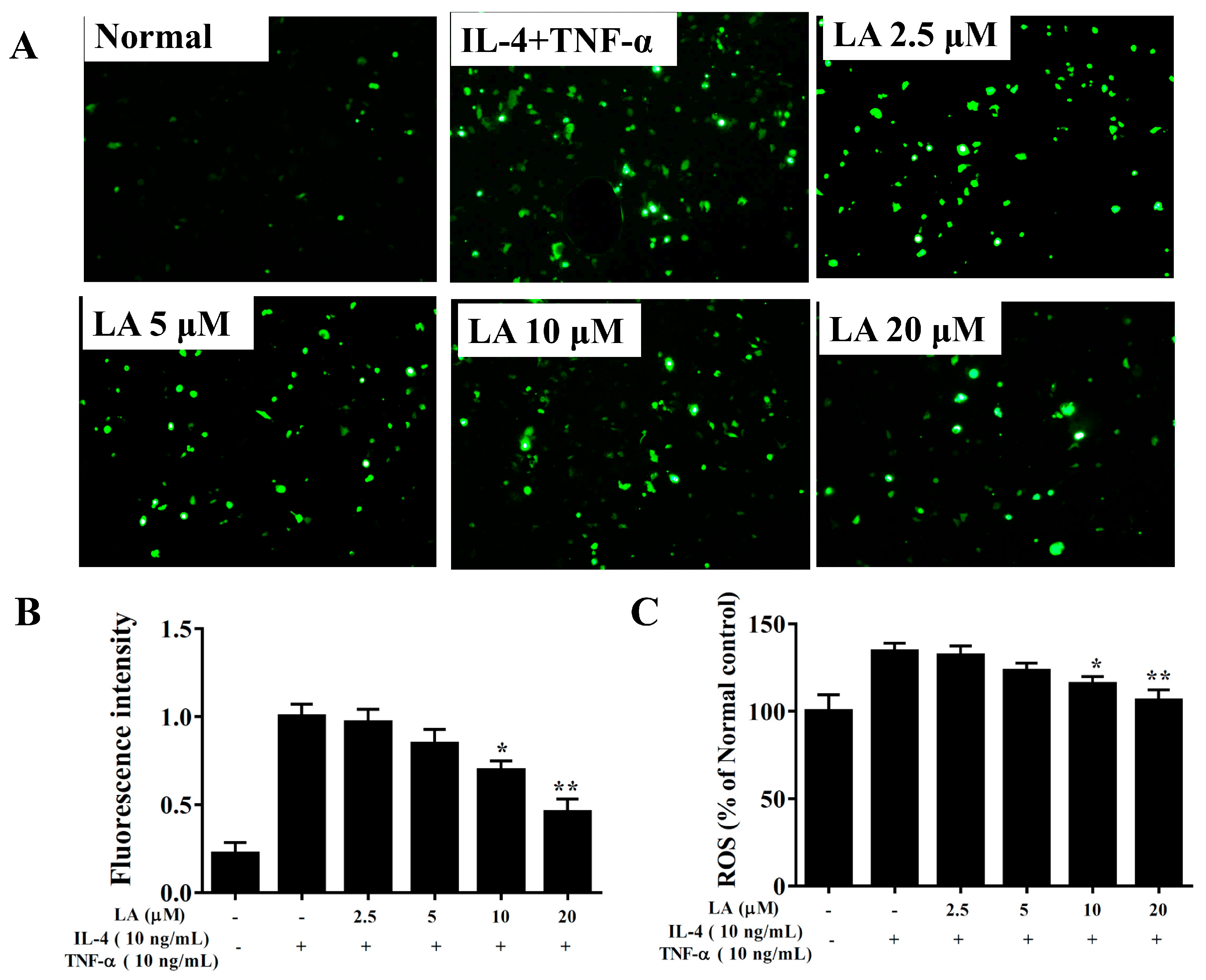
3.10. Effect of Licochalcone A on ROS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aghasafari, P.; George, U.; Pidaparti, R. A review of inflammatory mechanism in airway diseases. Inflamm. Res. 2019, 68, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.; Baptist, A.P.; Divekar, R.; Parikh, N.; Seggev, J.S.; Yusin, J.S.; Nyenhuis, S.M. Asthma in the older adult. J. Asthma. 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Ray, A.; Raundhal, M.; Oriss, T.B.; Ray, P.; Wenzel, S.E. Current concepts of severe asthma. J. Clin. Invest. 2016, 126, 2394–2403. [Google Scholar] [CrossRef] [PubMed]
- KleinJan, A. Airway inflammation in asthma: Key players beyond the Th2 pathway. Curr. Opin. Pulm. Med. 2016, 22, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Parulekar, A.D.; Diamant, Z.; Hanania, N.A. Role of biologics targeting type 2 airway inflammation in asthma: What have we learned so far? Curr. Opin. Pulm. Med. 2017, 23, 3–11. [Google Scholar] [CrossRef]
- Parulekar, A.D.; Kao, C.C.; Diamant, Z.; Hanania, N.A. Targeting the interleukin-4 and interleukin-13 pathways in severe asthma: Current knowledge and future needs. Curr. Opin. Pulm. Med. 2018, 24, 50–55. [Google Scholar] [CrossRef] [PubMed]
- de Groot, L.E.S.; van der Veen, T.A.; Martinez, F.O.; Hamann, J.; Lutter, R.; Melgert, B.N. Oxidative stress and macrophages: Driving forces behind exacerbations of asthma and chronic obstructive pulmonary disease? Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L369–L384. [Google Scholar] [CrossRef] [PubMed]
- Nakagome, K.; Nagata, M. Involvement and possible role of eosinophils in asthma exacerbation. Front. Immunol. 2018, 9, 2220. [Google Scholar] [CrossRef] [PubMed]
- Persson, C. Airways exudation of plasma macromolecules: Innate defense, epithelial regeneration, and asthma. J. Allergy Clin. Immunol. 2019, 143, 1271–1286. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, C.M.; Saglani, S. Epithelial cytokines and pulmonary allergic inflammation. Curr. Opin. Immunol. 2015, 34, 52–58. [Google Scholar] [CrossRef]
- Suzuki, M.; Bandoski, C.; Bartlett, J.D. Fluoride induces oxidative damage and SIRT1/autophagy through ROS-mediated JNK signaling. Free Radic. Biol. Med. 2015, 89, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatta, D.M.P.; Franceschelli, S.; Felaco, M.; Speranza, L. Biological effects of licochalcones. Mini Rev. Med. Chem. 2019, 19, 647–656. [Google Scholar]
- Furusawa, J.; Funakoshi-Tago, M.; Mashino, T.; Tago, K.; Inoue, H.; Sonoda, Y.; Kasahara, T. Glycyrrhiza inflata-derived chalcones, Licochalcone A, Licochalcone B and Licochalcone D, inhibit phosphorylation of NF-kappaB p65 in LPS signaling pathway. Int. Immunopharmacol. 2009, 9, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Kuribayashi, K.; Iida, S.; Nakajima, Y.; Funaguchi, N.; Tabata, C.; Fukuoka, K.; Fujimori, Y.; Ihaku, D.; Nakano, T. Suppression of heme oxygenase-1 activity reduces airway hyperresponsiveness and inflammation in a mouse model of asthma. J. Asthma. 2015, 52, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Su, H.H.; Fang, L.W.; Wu, S.J.; Liou, C.J. Licochalcone A inhibits cellular motility by suppressing E-cadherin and MAPK signaling in breast cancer. Cells 2019, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ma, Y.; Wei, X.; Fan, T. Neuroprotective effect of licochalcone A against oxygen-glucose deprivation/reperfusion in rat primary cortical neurons by attenuating oxidative stress injury and inflammatory response via the SIRT1/Nrf2 pathway. J. Cell Biochem. 2018, 119, 3210–3219. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Jiang, L.; Wei, M.; Yang, X.; Guan, M.; Xie, X.; Wei, J.; Liu, D.; Wang, D. Attenuation of allergic airway inflammation in a murine model of asthma by Licochalcone A. Immunopharmacol. Immunotoxicol. 2013, 35, 653–661. [Google Scholar] [CrossRef]
- Liou, C.J.; Huang, W.C. Dehydroepiandrosterone suppresses eosinophil infiltration and airway hyperresponsiveness via modulation of chemokines and Th2 cytokines in ovalbumin-sensitized mice. J. Clin. Immunol. 2011, 31, 656–665. [Google Scholar] [CrossRef]
- Liou, C.J.; Cheng, C.Y.; Yeh, K.W.; Wu, Y.H.; Huang, W.C. Protective effects of casticin from Vitex trifolia alleviate eosinophilic airway inflammation and oxidative stress in a murine asthma model. Front. Pharmacol. 2018, 9, 635. [Google Scholar] [CrossRef]
- Huang, W.C.; Fang, L.W.; Liou, C.J. Phloretin attenuates allergic airway inflammation and oxidative stress in asthmatic mice. Front. Immunol. 2017, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Lai, C.L.; Liang, Y.T.; Hung, H.C.; Liu, H.C.; Liou, C.J. Phloretin attenuates LPS-induced acute lung injury in mice via modulation of the NF-kappaB and MAPK pathways. Int. Immunopharmacol. 2016, 40, 98–105. [Google Scholar] [CrossRef]
- Huang, W.C.; Chan, C.C.; Wu, S.J.; Chen, L.C.; Shen, J.J.; Kuo, M.L.; Chen, M.C.; Liou, C.J. Matrine attenuates allergic airway inflammation and eosinophil infiltration by suppressing eotaxin and Th2 cytokine production in asthmatic mice. J. Ethnopharmacol. 2014, 151, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Myou, S.; Leff, A.R.; Myo, S.; Boetticher, E.; Tong, J.; Meliton, A.Y.; Liu, J.; Munoz, N.M.; Zhu, X. Blockade of inflammation and airway hyperresponsiveness in immune-sensitized mice by dominant-negative phosphoinositide 3-kinase-TAT. J. Exp. Med. 2003, 198, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.Y.; Huang, W.C.; Liou, C.J.; Chen, L.C.; Shen, J.J.; Kuo, M.L. Tomatidine attenuates airway hyperresponsiveness and inflammation by suppressing Th2 cytokines in a mouse model of asthma. Mediat. Inflamm. 2017, 2017, 5261803. [Google Scholar] [CrossRef] [PubMed]
- Liou, C.J.; Huan, Y.L.; Huang, W.C.; Yeh, K.W.; Huang, T.Y.; Lin, C.F. Water extract of Helminthostachys zeylanica attenuates LPS-induced acute lung injury in mice by modulating NF-κB and MAPK pathways. J. Ethnopharmacol. 2017, 199, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.L.; Huang, W.C.; Cheng, S.C.; Liou, C.J. Fisetin inhibits the generation of inflammatory mediators in interleukin-1beta-induced human lung epithelial cells by suppressing the NF-kappaB and ERK1/2 pathways. Int. Immunopharmacol. 2018, 60, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Wu, S.J.; Tu, R.S.; Lai, Y.R.; Liou, C.J. Phloretin inhibits interleukin-1beta-induced COX-2 and ICAM-1 expression through inhibition of MAPK, Akt, and NF-kappaB signaling in human lung epithelial cells. Food Funct. 2015, 6, 1960–1967. [Google Scholar] [CrossRef]
- Liou, C.J.; Lee, Y.K.; Ting, N.C.; Chen, Y.L.; Shen, S.C.; Wu, S.J.; Huang, W.C. Protective effects of Licochalcone A ameliorates obesity and non-alcoholic fatty liver disease via promotion of the Sirt-1/AMPK pathway in mice fed a high-fat diet. Cells 2019, 8, 447. [Google Scholar] [CrossRef]
- Huang, W.C.; Dai, Y.W.; Peng, H.L.; Kang, C.W.; Kuo, C.Y.; Liou, C.J. Phloretin ameliorates chemokines and ICAM-1 expression via blocking of the NF-kappaB pathway in the TNF-alpha-induced HaCaT human keratinocytes. Int. Immunopharmacol. 2015, 27, 32–37. [Google Scholar] [CrossRef]
- Saglani, S.; Lloyd, C.M. Eosinophils in the pathogenesis of paediatric severe asthma. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Antus, B. Oxidative Stress Markers in Sputum. Oxid. Med. Cell Longev. 2016, 2016, 2930434. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Wu, J.; Di, C.; Xia, Z. Heme oxygenase-1 exerts a protective role in ovalbumin-induced neutrophilic airway inflammation by inhibiting Th17 cell-mediated immune response. J. Biol. Chem. 2013, 288, 34612–34626. [Google Scholar] [CrossRef] [PubMed]
- Zahiruddin, A.S.; Grant, J.A.; Sur, S. Role of epigenetics and DNA-damage in asthma. Curr. Opin. Allergy Clin. Immunol. 2018, 18, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, J.; Shu, J.; Li, H.; Ren, Z. Oxidative damage and DNA damage in lungs of an ovalbumin-induced asthmatic murine model. J. Thorac. Dis. 2018, 10, 4819–4830. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.K.; Loh, X.Y.; Peh, H.Y.; Tan, W.N.; Tan, W.S.; Li, N.; Tay, I.J.; Wong, W.S.; Engelward, B.P. House dust mite-induced asthma causes oxidative damage and DNA double-strand breaks in the lungs. J. Allergy Clin. Immunol. 2016, 138, 84–96. [Google Scholar] [CrossRef]
- Bortolotto, L.F.; Barbosa, F.R.; Silva, G.; Bitencourt, T.A.; Beleboni, R.O.; Baek, S.J.; Marins, M.; Fachin, A.L. Cytotoxicity of trans-chalcone and licochalcone A against breast cancer cells is due to apoptosis induction and cell cycle arrest. Biomed. Pharmacother. 2017, 85, 425–433. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Z.; Meng, R.; Shi, C.; Guo, N. Antioxidative and anticancer properties of Licochalcone A from licorice. J. Ethnopharmacol. 2017, 198, 331–337. [Google Scholar] [CrossRef]
- Chu, X.; Ci, X.; Wei, M.; Yang, X.; Cao, Q.; Guan, M.; Li, H.; Deng, Y.; Feng, H.; Deng, X. Licochalcone a inhibits lipopolysaccharide-induced inflammatory response in vitro and in vivo. J. Agric. Food Chem. 2012, 60, 3947–3954. [Google Scholar] [CrossRef]
- Dandekar, A.; Mendez, R.; Zhang, K. Cross talk between ER stress, oxidative stress, and inflammation in health and disease. Methods Mol. Biol. 2015, 1292, 205–214. [Google Scholar]
- Hiemstra, P.S.; McCray, P.B., Jr.; Bals, R. The innate immune function of airway epithelial cells in inflammatory lung disease. Eur. Respir. J. 2015, 45, 1150–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laforgia, N.; Di Mauro, A.; Favia Guarnieri, G.; Varvara, D.; De Cosmo, L.; Panza, R.; Capozza, M.; Baldassarre, M.E.; Resta, N. The role of oxidative stress in the pathomechanism of congenital malformations. Oxid. Med. Cell Longev. 2018, 2018, 7404082. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhu, L.; Zhang, D.; Li, N.; Li, Q.; Dai, P.; Mao, Y.; Li, X.; Ma, J.; Huang, S. Oxidative stress-related biomarkers in Postmenopausal osteoporosis: A systematic review and meta-analyses. Dis. Markers 2016, 2016, 7067984. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Xiao, Q.; Zhou, J.; Feng, H.; Liu, G.; Ci, X. Licochalcone A upregulates Nrf2 antioxidant pathway and thereby alleviates acetaminophen-induced hepatotoxicity. Front. Pharmacol. 2018, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.J.; Lee, C.M.; Lin, T.H.; Lin, H.Y.; Lee, S.Y.; Mesri, M.; Chang, K.H.; Lin, J.Y.; Lee-Chen, G.J.; Chen, C.M. Chinese herbal medicine Glycyrrhiza inflata reduces abeta aggregation and exerts neuroprotection through anti-oxidation and anti-inflammation. Am. J. Chin. Med. 2018, 4, 1–25. [Google Scholar]
- Hou, X.; Yang, S.; Zheng, Y. Licochalcone A attenuates abdominal aortic aneurysm induced by angiotensin II via regulating the miR-181b/SIRT1/HO-1 signaling. J. Cell Physiol. 2019, 234, 7560–7568. [Google Scholar] [CrossRef]
- Sonar, S.S.; Ehmke, M.; Marsh, L.M.; Dietze, J.; Dudda, J.C.; Conrad, M.L.; Renz, H.; Nockher, W.A. Clara cells drive eosinophil accumulation in allergic asthma. Eur. Respir. J. 2012, 39, 429–438. [Google Scholar] [CrossRef]
- Charron, C.B.; Pakhale, S. The role of airway hyperresponsiveness measured by methacholine challenge test in defining asthma severity in asthma-obesity syndrome. Curr. Opin. Allergy Clin. Immunol. 2016, 16, 218–223. [Google Scholar] [CrossRef]
- Belanger, K.K.; Ameredes, B.T.; Boldogh, I.; Aguilera-Aguirre, L. The potential role of 8-oxoguanine DNA glycosylase-driven DNA base excision repair in exercise-induced asthma. Mediat. Inflamm. 2016, 2016, 3762561. [Google Scholar] [CrossRef]
- Fatani, S.H. Biomarkers of oxidative stress in acute and chronic bronchial asthma. J. Asthma. 2014, 51, 578–584. [Google Scholar] [CrossRef]
- Yancey, S.W.; Keene, O.N.; Albers, F.C.; Ortega, H.; Bates, S.; Bleecker, E.R.; Pavord, I. Biomarkers for severe eosinophilic asthma. J. Allergy Clin. Immunol. 2017, 140, 1509–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royce, S.G.; Cheng, V.; Samuel, C.S.; Tang, M.L. The regulation of fibrosis in airway remodeling in asthma. Mol. Cell Endocrinol. 2012, 351, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.G.; Thomas, B.; Koff, J. TGF-beta: Master regulator of inflammation and fibrosis. Respirology 2018, 23, 1096–1097. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Gene Number |
|---|---|---|---|
| COX-2 | ACCAGCAGTTCCAGTATCAGA | CAGGAGGATGGAGTTGTTGTAG | NM_011198 |
| MUC5AC | AATGCTGGTGCCTGTGTCT | CCTCCTATGCCATCTGTTGTG | NM_017511 |
| IL-6 | AGGACCAAGACCATCCAATTCA | GCTTAGGCATAACGCACTAGG | NM_031168 |
| IFN-γ | CAGCAACAACATAAGCGTCATT | ACCTCAAACTTGGCAATACTCA | NM_000619.2 |
| IL-13 | GCTCCAGCATTGAAGCAGTG | CGTGGCAGACAGGAGTGTT | NM_008355.3 |
| IL-5 | ATCCTCCTGCCTCCTCTTCC | GGTTCCATCTCCAGCACTTCA | NM_000879 |
| IL-4 | TCCGTGCTTGAAGAAGAACTC | GTGATGTGGACTTGGACTCATT | NM_021283.2 |
| CCL24 | AGGCAGTGAGAACCAAGT | GCGTCAATACCTATGTCCAA | NM_019577.4 |
| CCL11 | GGCTTCATGTAGTTCCAGAT | CCATTGTGTTCCTCAATAATCC | NM_011330.3 |
| β-actin | AAGACCTCTATGCCAACACAGT | AGCCAGAGCAGTAATCTCCTTC | NM_007393.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.-C.; Liu, C.-Y.; Shen, S.-C.; Chen, L.-C.; Yeh, K.-W.; Liu, S.-H.; Liou, C.-J. Protective Effects of Licochalcone A Improve Airway Hyper-Responsiveness and Oxidative Stress in a Mouse Model of Asthma. Cells 2019, 8, 617. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8060617
Huang W-C, Liu C-Y, Shen S-C, Chen L-C, Yeh K-W, Liu S-H, Liou C-J. Protective Effects of Licochalcone A Improve Airway Hyper-Responsiveness and Oxidative Stress in a Mouse Model of Asthma. Cells. 2019; 8(6):617. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8060617
Chicago/Turabian StyleHuang, Wen-Chung, Chien-Yu Liu, Szu-Chuan Shen, Li-Chen Chen, Kuo-Wei Yeh, Shih-Hai Liu, and Chian-Jiun Liou. 2019. "Protective Effects of Licochalcone A Improve Airway Hyper-Responsiveness and Oxidative Stress in a Mouse Model of Asthma" Cells 8, no. 6: 617. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8060617