Systemic Metabolomic Profiling of Acute Myeloid Leukemia Patients before and During Disease-Stabilizing Treatment Based on All-Trans Retinoic Acid, Valproic Acid, and Low-Dose Chemotherapy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. The Antileukemic Treatment Based on ATRA, Valproic Acid, and Low-Dose Cytotoxic Drugs
2.3. Preparation and Analysis of Serum Samples
2.4. Statistical Analyses
3. Results
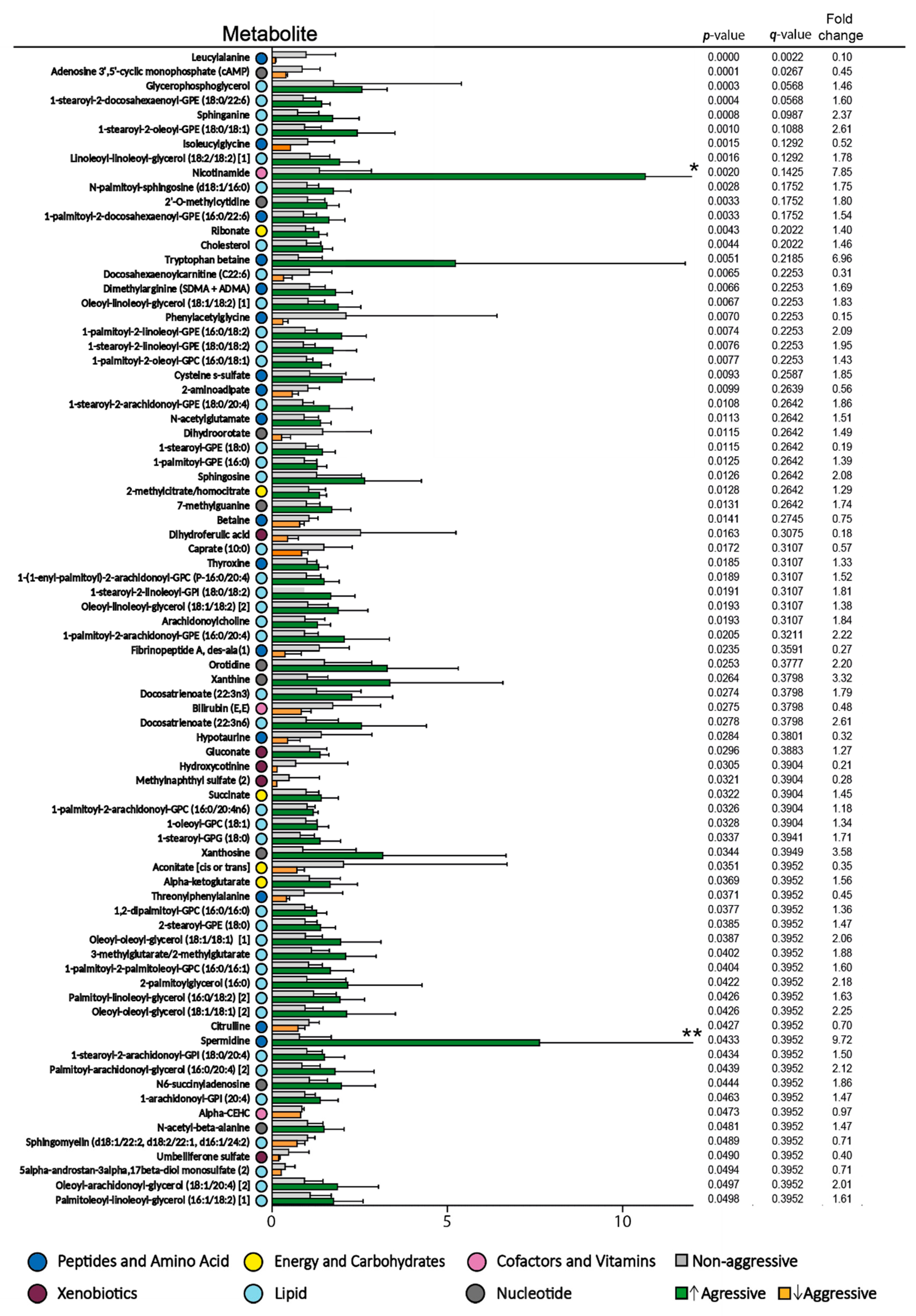
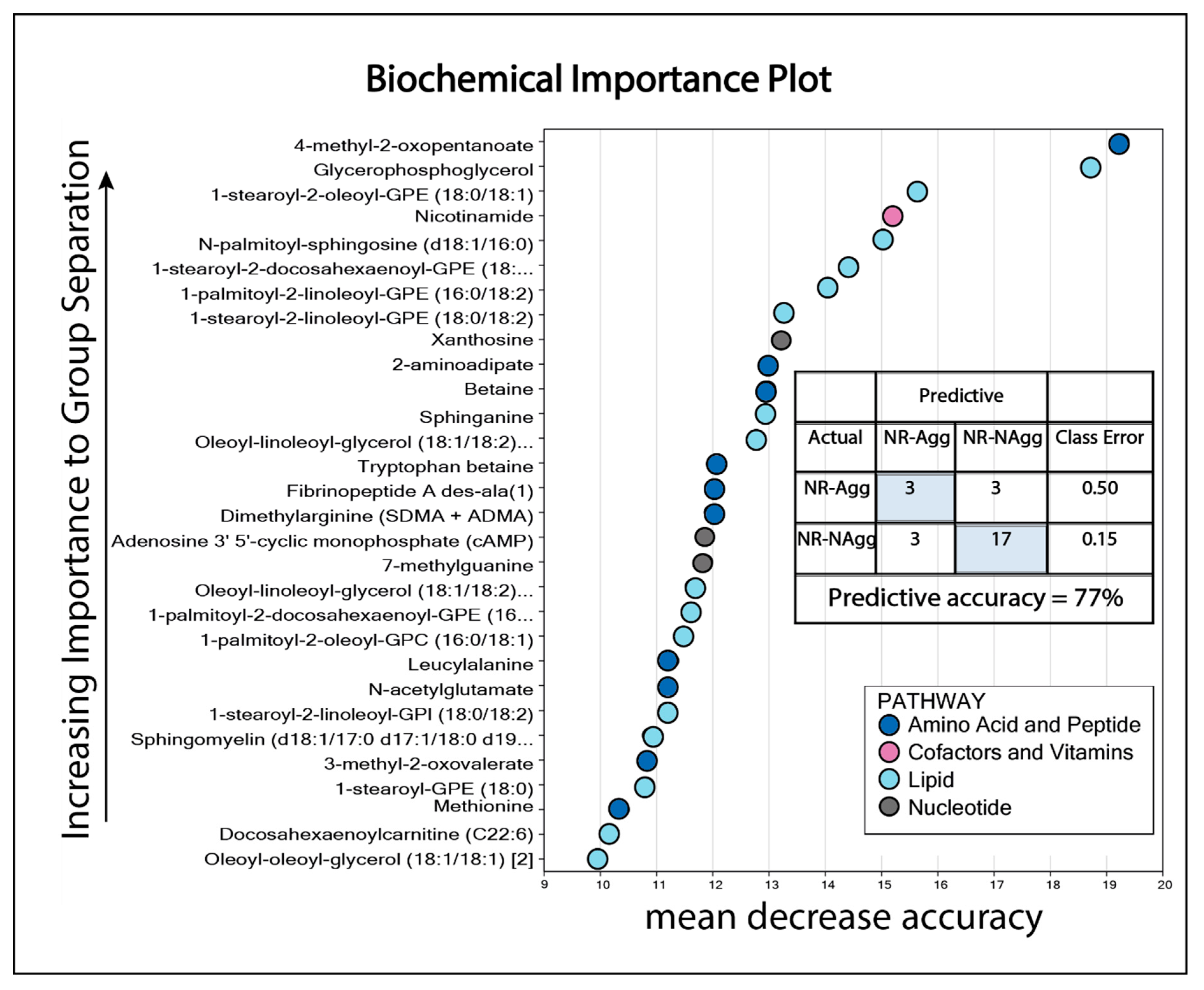
3.1. The Pretherapy Serum Metabolomic Profiles Could Not Predict Responsiveness to Antileukemic Treatment Based on ATRA, Valproic Acid, and Low-Dose Chemotherapy
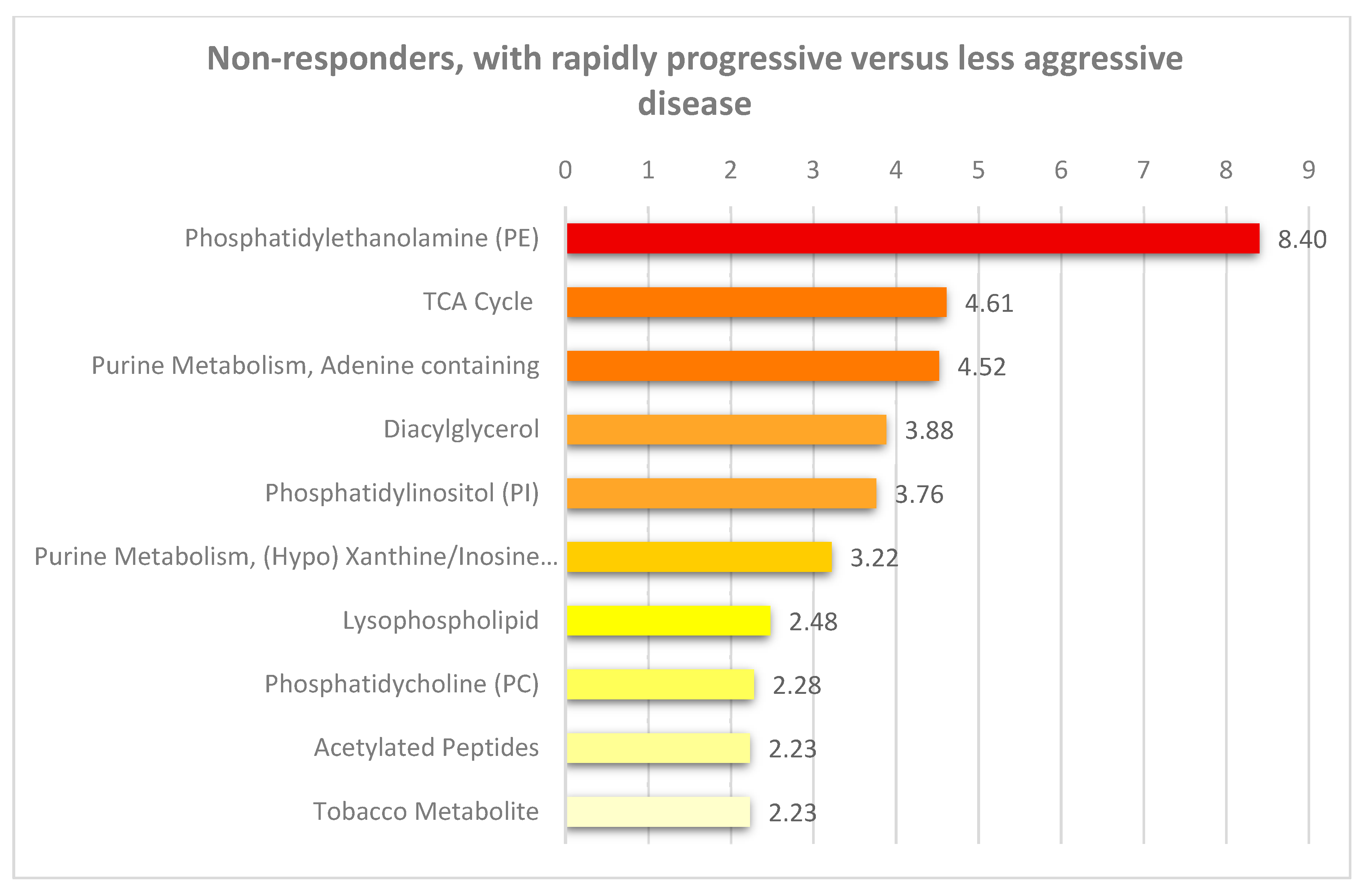
3.2. Pretherapy Differences in Fatty Acid Metabolism are Found between Non-Responders with Rapidly Progressive AML Compared with Other Non-Responders with Less Aggressive Disease
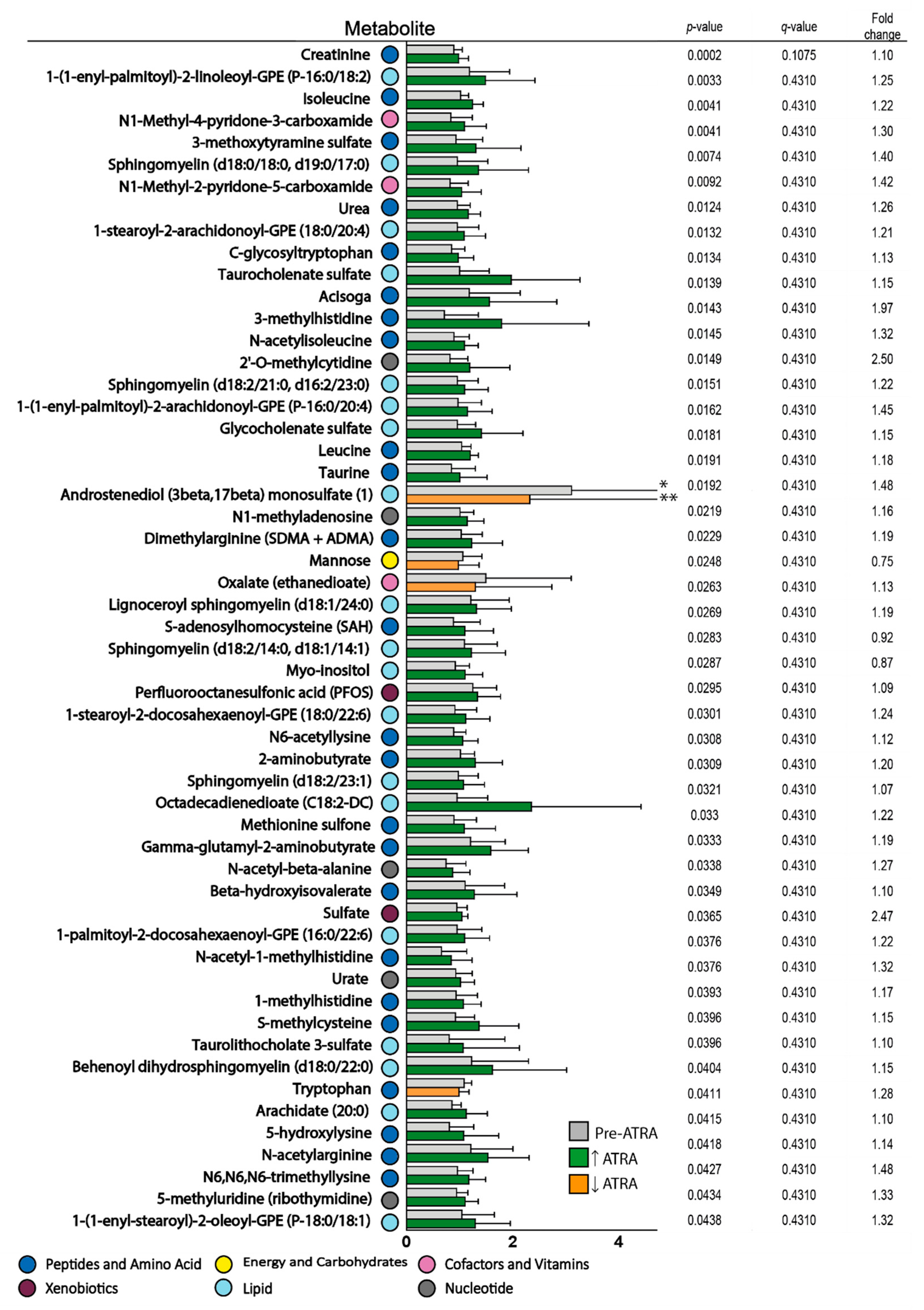
3.3. Effects of ATRA Monotherapy on Serum Metabolite Profiles
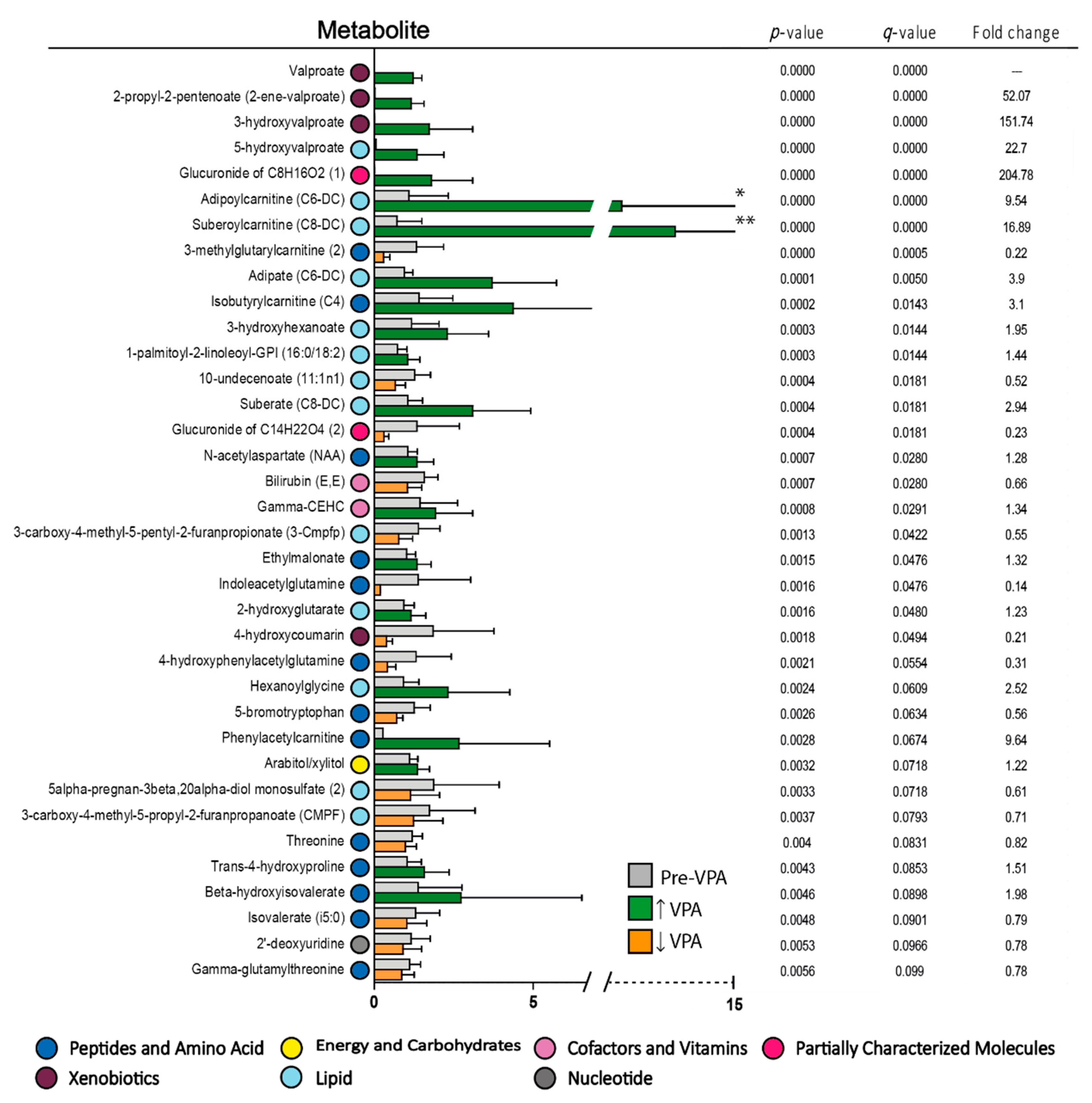
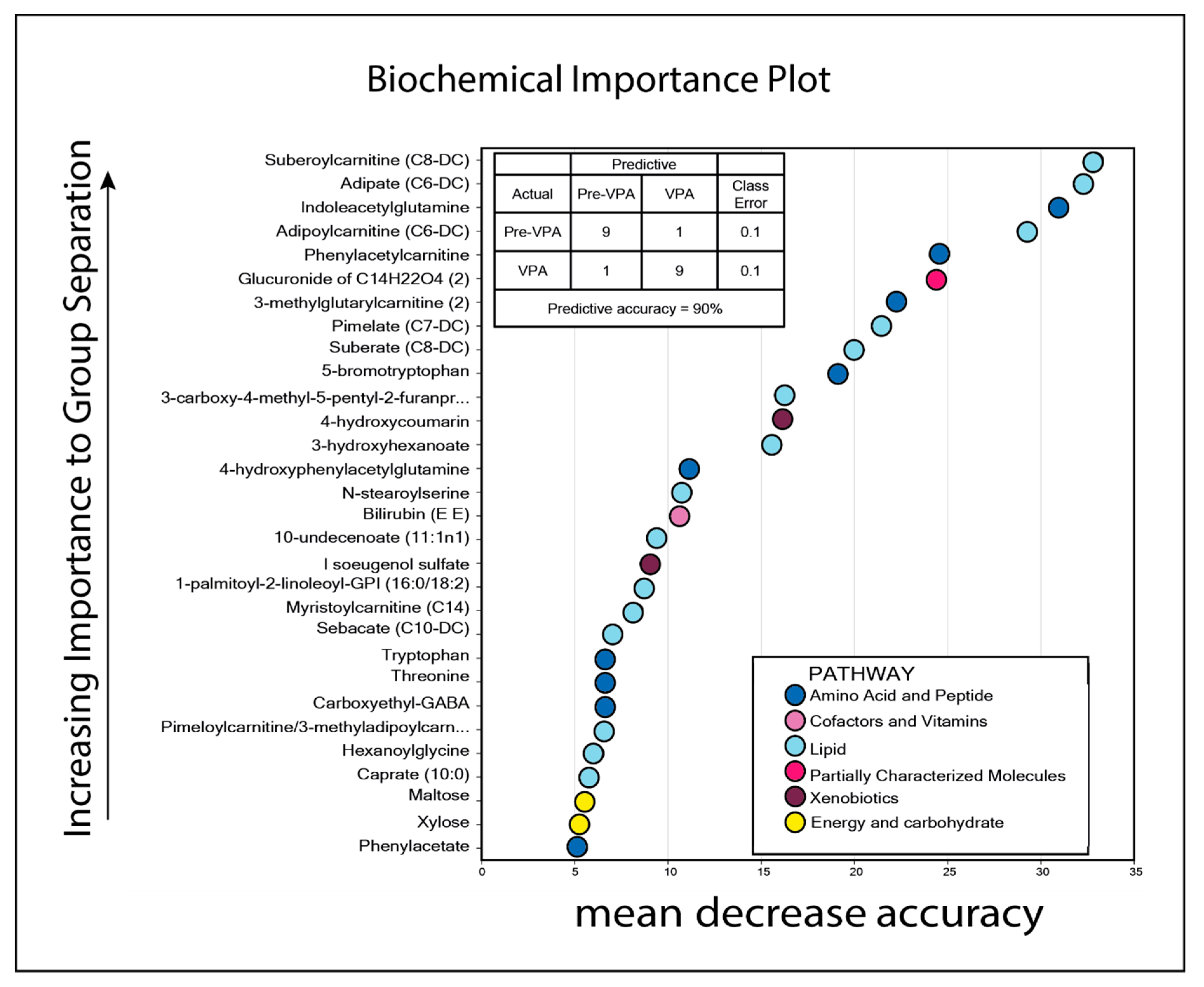
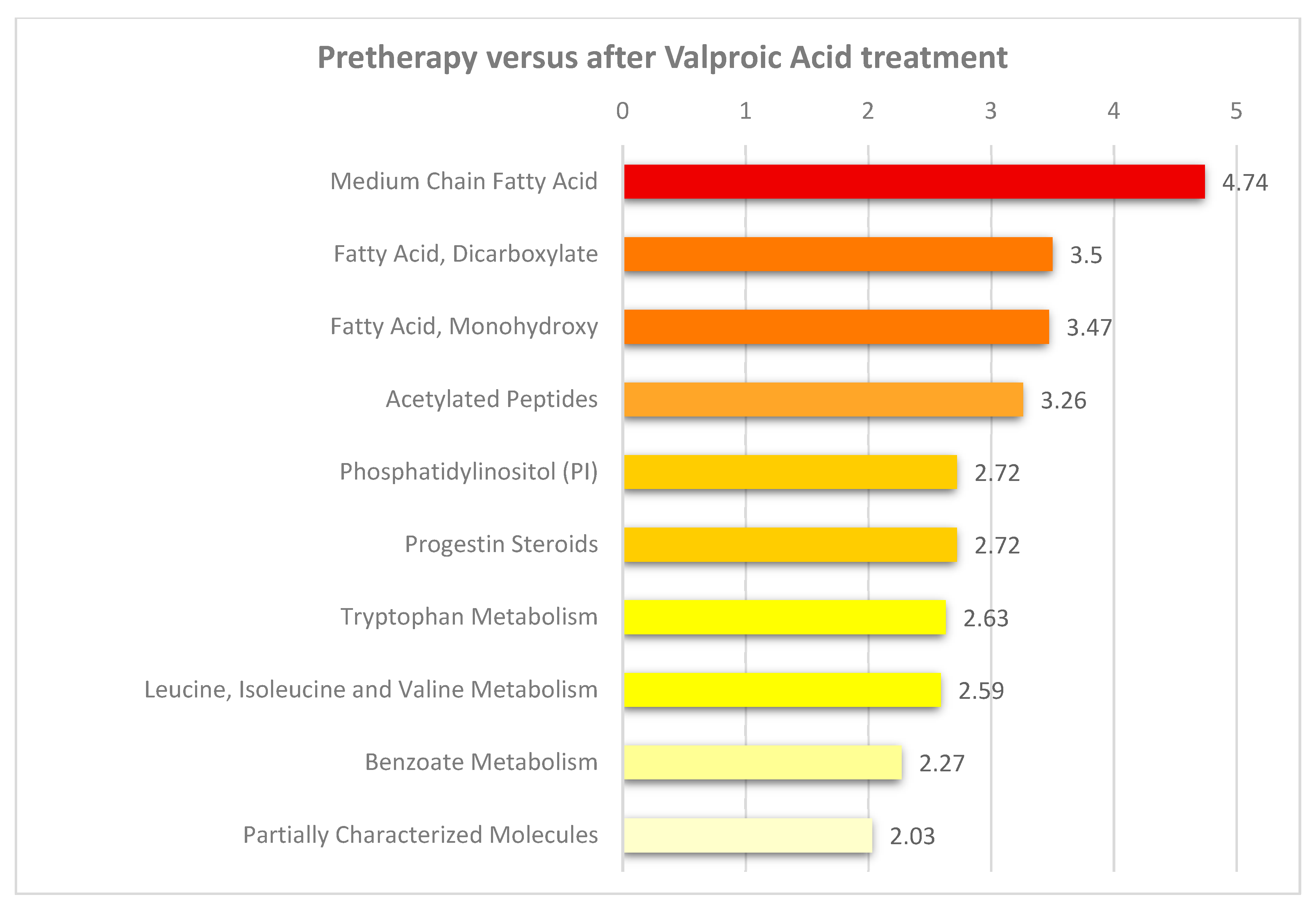
3.4. Valproic Acid Monotherapy Alters the Systemic Amino Acid and Lipid Metabolite Profiles in Both Responders and Non-Responders to the Antileukemic Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Steffen, B.; Muller-Tidow, C.; Schwable, J.; Berdel, W.E.; Serve, H. The molecular pathogenesis of acute myeloid leukemia. Crit. Rev. Oncol. Hemat. 2005, 56, 195–221. [Google Scholar] [CrossRef] [PubMed]
- Dohner, H.; Estey, E.H.; Amadori, S.; Appelbaum, F.R.; Buchner, T.; Burnett, A.K.; Dombret, H.; Fenaux, P.; Grimwade, D.; Larson, R.A.; et al. Diagnosis and management of acute myeloid leukemia in adults: Recommendations from an international expert panel, on behalf of the European LeukemiaNet. Blood 2010, 115, 453–474. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.R.A.; Reid, M.M.; Stark, A.N.; Bown, N.; Hamilton, P.J.; Proctor, S.J. De-novo acute myeloid-leukemia in patients over 55-years-old - a population-based study of incidence, treatment and outcome. Leukemia 1995, 9, 231–237. [Google Scholar] [PubMed]
- Dohner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute myeloid leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [PubMed]
- Kantarjian, H.; Ravandi, F.; O′Brien, S.; Cortes, J.; Faderl, S.; Garcia-Manero, G.; Jabbour, E.; Wierda, W.; Kadia, T.; Pierce, S.; et al. Intensive chemotherapy does not benefit most older patients (age 70 years or older) with acute myeloid leukemia. Blood 2010, 116, 4422–4429. [Google Scholar] [CrossRef] [PubMed]
- Giles, F.J.; Borthakur, G.; Ravandi, F.; Faderl, S.; Verstovsek, S.; Thomas, D.; Wierda, W.; Ferrajoli, A.; Kornblau, S.; Pierce, S.; et al. The haematopoietic cell transplantation comorbidity index score is predictive of early death and survival in patients over 60 years of age receiving induction therapy for acute myeloid leukaemia. Brit. J. Haematol. 2007, 136, 624–627. [Google Scholar] [CrossRef] [PubMed]
- Latagliata, R.; Bongarzoni, V.; Carmosino, I.; Mengarelli, A.; Breccia, M.; Borza, P.A.; D′Andrea, M.; D′Elia, G.M.; Mecarocci, S.; Morano, S.G.; et al. Acute myelogenous leukemia in elderly patients not eligible for intensive chemotherapy: The dark side of the moon. Ann. Oncol. 2006, 17, 281–285. [Google Scholar] [CrossRef]
- Thol, F.; Schlenk, R.F.; Heuser, M.; Ganser, A. How I treat refractory and early relapsed acute myeloid leukemia. Blood 2015, 126, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Delcuve, G.P.; Khan, D.H.; Davie, J.R. Roles of histone deacetylases in epigenetic regulation: Emerging paradigms from studies with inhibitors. Clin. Epigenetics 2012, 4. [Google Scholar] [CrossRef]
- Lee, J.; R, S.H. Cancer epigenetics: Mechanisms and crosstalk of a HDAC inhibitor, vorinostat. Chemotherapy 2013, 2, 14934. [Google Scholar]
- Quintas-Cardama, A.; Santos, F.P.S.; Garcia-Manero, G. Histone deacetylase inhibitors for the treatment of myelodysplastic syndrome and acute myeloid leukemia. Leukemia 2011, 25, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Tickenbrock, L.; Klein, H.U.; Trento, C.; Hascher, A.; Gollner, S.; Baumer, N.; Kuss, R.; Agrawal, S.; Bug, G.; Serve, H.; et al. Increased HDAC1 deposition at hematopoietic promoters in AML and its association with patient survival. Leuk. Res. 2011, 35, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Gottlicher, M.; Minucci, S.; Zhu, P.; Kramer, O.H.; Schimpf, A.; Giavara, S.; Sleeman, J.P.; Lo Coco, F.; Nervi, C.; Pelicci, P.G.; et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. Embo. J. 2001, 20, 6969–6978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimino, G.; Lo-Coco, F.; Fenu, S.; Travaglini, L.; Finolezzi, E.; Mancini, M.; Nanni, M.; Careddu, A.; Fazi, F.; Padula, F.; et al. Sequential valproic acid/all-trans retinoic acid treatment reprograms differentiation in refractory and high-risk acute myeloid leukemia. Cancer Res. 2006, 66, 8903–8911. [Google Scholar] [CrossRef] [PubMed]
- Leiva, M.; Moretti, S.; Soilihi, H.; Pallavicini, I.; Peres, L.; Mercurio, C.; Dal Zuffo, R.; Minucci, S.; de The, H. Valproic acid induces differentiation and transient tumor regression, but spares leukemia-initiating activity in mouse models of APL. Leukemia 2012, 26, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Klisovic, R.B.; Vukosavljevic, T.; Yu, J.H.; Paschka, P.; Huynh, L.; Pang, J.X.; Neviani, P.; Liu, Z.F.; Blum, W.; et al. Targeting AML1/ETO-histone deacetylase repressor complex: A novel mechanism for valproic acid-mediated gene expression and cellular differentiation in AML1/ETO-positive acute myeloid leukemia cells. J. Pharm. Exp. 2007, 321, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Stapnes, C.; Ryningen, A.; Hatfield, K.; Oyan, A.M.; Eide, G.E.; Corbascio, M.; Kalland, K.H.; Gjertsen, B.T.; Bruserud, O. Functional characteristics and gene expression profiles of primary acute myeloid leukaemia cells identify patient subgroups that differ in susceptibility to histone deacetylase inhibitors. Int. J. Oncol. 2007, 31, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Forthun, R.B.; Hellesøy, M.; Sulen, A.; Kopperud, R.K.; Sjøholt, G.; Bruserud, Ø.; McCormack, E.; Gjertsen, B.T. Modulation of phospho-proteins by interferon-alpha and valproic acid in acute myeloid leukemia. J. Cancer Res. Clin. Oncol. 2019, 145, 1729–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredly, H.; Ersvaer, E.; Kittang, A.O.; Tsykunova, G.; Gjertsen, B.T.; Bruserud, O. The combination of valproic acid, all-trans retinoic acid and low-dose cytarabine as disease-stabilizing treatment in acute myeloid leukemia. Clin. Epigenetics 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Fredly, H.; Gjertsen, B.T.; Bruserud, O. Histone deacetylase inhibition in the treatment of acute myeloid leukemia: The effects of valproic acid on leukemic cells, and the clinical and experimental evidence for combining valproic acid with other antileukemic agents. Clin. Epigenetics 2013, 5. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Chen, Z. Acute promyelocytic leukemia: From highly fatal to highly curable. Blood 2008, 111, 2505–2515. [Google Scholar] [CrossRef] [PubMed]
- Fredly, H.; Ersvaer, E.; Stapnes, C.; Gjertsen, B.T.; Bruserud, O. The combination of conventional chemotherapy with new targeted therapy in hematologic malignancies: The safety and efficiency of low- dose cytarabine supports its combination with new therapeutic agents in early clinical trials. Curr. Cancer Ther. Rev. 2009, 5, 243–255. [Google Scholar] [CrossRef]
- Dimberg, A.; Bahram, F.; Karlberg, I.; Larsson, L.G.; Nilsson, K.; Oberg, F. Retinoic acid-induced cell cycle arrest of human myeloid cell lines is associated with sequential down-regulation of c-Myc and cyclin E and posttranscriptional up-regulation of p27(Kip1). Blood 2002, 99, 2199–2206. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Redner, R.L. An ATRActive future for differentiation therapy in AML. Blood Rev. 2015, 29, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forthun, R.B.; SenGupta, T.; Skjeldam, H.K.; Lindvall, J.M.; McCormack, E.; Gjertsen, B.T.; Nilsen, H. Cross-species functional genomic analysis identifies resistance genes of the histone deacetylase inhibitor valproic acid. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Fredly, H.; Reikvam, H.; Gjertsen, B.T.; Bruserud, O. Disease-stabilizing treatment with all-trans retinoic acid and valproic acid in acute myeloid leukemia: Serum hsp70 and hsp90 levels and serum cytokine profiles are determined by the disease, patient age, and anti-leukemic treatment. Am. J. Hematol. 2012, 87, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryningen, A.; Stapnes, C.; Lassalle, P.; Corbascio, M.; Gjertsen, B.T.; Bruserud, O. A subset of patients with high-risk acute myelogenous leukemia shows improved peripheral blood cell counts when treated with the combination of valproic acid, theophylline and all-trans retinoic acid. Leuk Res. 2009, 33, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.; Wang, J.H.; Zhao, A.H.; Xu, X.; Wang, Y.H.; Chen, T.L.; Li, J.M.; Mi, J.Q.; Zhu, Y.M.; Liu, Y.F.; et al. A distinct glucose metabolism signature of acute myeloid leukemia with prognostic value. Blood 2014, 124, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Nepstad, I.; Reikvam, H.; Brenner, A.K.; Bruserud, O.; Hatfield, K.J. Resistance to the antiproliferative in vitro effect of PI3K-Akt-mTOR inhibition in primary human acute myeloid leukemia cells is associated with altered cell metabolism. Int. J. Mol. Sci. 2018, 19, 382. [Google Scholar] [CrossRef]
- Amengual, J.; Garcia-Carrizo, F.J.; Arreguin, A.; Musinovic, H.; Granados, N.; Palou, A.; Bonet, M.L.; Ribot, J. Retinoic acid increases fatty acid oxidation and irisin expression in skeletal muscle cells and impacts irisin in vivo. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 187–202. [Google Scholar] [CrossRef]
- Lheureux, P.E.; Hantson, P. Carnitine in the treatment of valproic acid-induced toxicity. Clin. Toxicol. (Phila. Pa.) 2009, 47, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Aires, C.C.; Soveral, G.; Luis, P.B.; ten Brink, H.J.; de Almeida, I.T.; Duran, M.; Wanders, R.J.; Silva, M.F. Pyruvate uptake is inhibited by valproic acid and metabolites in mitochondrial membranes. Febs. Lett. 2008, 582, 3359–3366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponchaut, S.; Van Hoof, F.; Veitch, K. In vitro effects of valproate and valproate metabolites on mitochondrial oxidations: Relevance of CoA sequestration to the observed inhibitions. Biochem. Pharm. 1992, 43, 2435–2442. [Google Scholar] [CrossRef]
- Silva, M.F.; Aires, C.C.; Luis, P.B.; Ruiter, J.P.; L, I.J.; Duran, M.; Wanders, R.J.; Tavares de Almeida, I. Valproic acid metabolism and its effects on mitochondrial fatty acid oxidation: A review. J. Inherit. Metab Dis. 2008, 31, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.F.B.; Ruiter, J.P.N.; Ijlst, L.; Jakobs, C.; Duran, M.; de Almeida, I.T.; Wanders, R.J.A. Differential effect of valproate and its Δ2- and Δ4-unsaturated metabolites, on the β-oxidation rate of long-chain and medium-chain fatty acids. Chem. -Biol. Interact. 2001, 137, 203–212. [Google Scholar] [CrossRef]
- Silva, M.F.; Selhorst, J.; Overmars, H.; van Gennip, A.H.; Maya, M.; Wanders, R.J.; de Almeida, I.T.; Duran, M. Characterization of plasma acylcarnitines in patients under valproate monotherapy using ESI-MS/MS. Clin. Biochem. 2001, 34, 635–638. [Google Scholar] [CrossRef]
- Ribot, J.; Arreguin, A.; Kuda, O.; Kopecky, J.; Palou, A.; Bonet, M.L. Novel markers of the metabolic impact of exogenous retinoic acid with a focus on acylcarnitines and amino acids. Int. J. Mol. Sci. 2019, 20, 3640. [Google Scholar] [CrossRef] [PubMed]
- Schlenk, R.F.; Lubbert, M.; Benner, A.; Lamparter, A.; Krauter, J.; Herr, W.; Martin, H.; Salih, H.R.; Kundgen, A.; Horst, H.A.; et al. All-trans retinoic acid as adjunct to intensive treatment in younger adult patients with acute myeloid leukemia: Results of the randomized AMLSG 07-04 study. Ann. Hematol. 2016, 95, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Lübbert, M.; Grishina, O.; Schmoor, C.; Schlenk, R.F.; Jost, E.; Krauter, J.; Heuser, M.; Thol, F.; Schittenhelm, M.M.; Salih, H.R.; et al. Results of the randomized phase II study decider (AMLSG 14-09) comparing decitabine (DAC) with or without valproic acid (VPA) and with or without all-trans retinoic acid (ATRA) add-on in newly diagnosed elderly non-fit AML patients. Blood 2016, 128, 589. [Google Scholar]
- Martelli, M.P.; Gionfriddo, I.; Mezzasoma, F.; Milano, F.; Pierangeli, S.; Mulas, F.; Pacini, R.; Tabarrini, A.; Pettirossi, V.; Rossi, R.; et al. Arsenic trioxide and all-trans retinoic acid target NPM1 mutant oncoprotein levels and induce apoptosis in NPM1-mutated AML cells. Blood 2015, 125, 3455–3465. [Google Scholar] [CrossRef]
- Ma, H.S.; Greenblatt, S.M.; Shirley, C.M.; Duffield, A.S.; Bruner, J.K.; Li, L.; Nguyen, B.; Jung, E.; Aplan, P.D.; Ghiaur, G.; et al. All-trans retinoic acid synergizes with FLT3 inhibition to eliminate FLT3/ITD+ leukemia stem cells in vitro and in vivo. Blood 2016, 127, 2867–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutzen, H.; Saland, E.; Larrue, C.; de Toni, F.; Gales, L.; Castelli, F.A.; Cathebas, M.; Zaghdoudi, S.; Stuani, L.; Kaoma, T.; et al. Isocitrate dehydrogenase 1 mutations prime the all-trans retinoic acid myeloid differentiation pathway in acute myeloid leukemia. J. Exp. Med. 2016, 213, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Tassara, M.; Dohner, K.; Brossart, P.; Held, G.; Gotze, K.; Horst, H.A.; Ringhoffer, M.; Kohne, C.H.; Kremers, S.; Raghavachar, A.; et al. Valproic acid in combination with all-trans retinoic acid and intensive therapy for acute myeloid leukemia in older patients. Blood 2014, 123, 4027–4036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davood, Z.A.; Shamsi, S.; Ghaedi, H.; Sahand, R.I.; Mojtaba, G.; Mahdi, T.; Reza, M.; Ebrahimi, M.J.; Miri-Moosavi, R.S.; Boosaliki, S.; et al. Valproic acid may exerts its cytotoxic effect through rassf1a expression induction in acute myeloid leukemia. Tumor Biol. 2016, 37, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Blagitko-Dorfs, N.; Schlosser, P.; Greve, G.; Pfeifer, D.; Meier, R.; Baude, A.; Brocks, D.; Plass, C.; Lubbert, M. Combination treatment of acute myeloid leukemia cells with DNMT and HDAC inhibitors: Predominant synergistic gene downregulation associated with gene body demethylation. Leukemia 2019, 33, 945–956. [Google Scholar] [CrossRef] [PubMed]
- van Gils, N.; Verhagen, H.; Smit, L. Reprogramming acute myeloid leukemia into sensitivity for retinoic-acid-driven differentiation. Exp. Hematol. 2017, 52, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Rucker, F.G.; Lang, K.M.; Futterer, M.; Komarica, V.; Schmid, M.; Dohner, H.; Schlenk, R.F.; Dohner, K.; Knudsen, S.; Bullinger, L. Molecular dissection of valproic acid effects in acute myeloid leukemia identifies predictive networks. Epigenetics 2016, 11, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Kreitz, J.; Schonfeld, C.; Seibert, M.; Stolp, V.; Alshamleh, I.; Oellerich, T.; Steffen, B.; Schwalbe, H.; Schnutgen, F.; Kurrle, N.; et al. Metabolic plasticity of acute myeloid leukemia. Cells 2019, 8, 805. [Google Scholar] [CrossRef]
- Castro, I.; Sampaio-Marques, B.; Ludovico, P. Targeting metabolic reprogramming in acute myeloid leukemia. Cells 2019, 8, 967. [Google Scholar] [CrossRef]
- Cheson, B.D.; Bennett, J.M.; Kantarjian, H.; Schiffer, C.A.; Nimer, S.D.; Löwenberg, B.; Stone, R.M.; Mittelman, M.; Sanz, G.F.; Wijermans, P.W.; et al. Myelodysplastic syndromes standardized response criteria: Further definition. Blood 2001, 98, 1985–1986. [Google Scholar] [CrossRef]
- Cheson, B.D.; Bennett, J.M.; Kopecky, K.J.; Buchner, T.; Willman, C.L.; Estey, E.H.; Schiffer, C.A.; Doehner, H.; Tallman, M.S.; Lister, T.A.; et al. Revised recommendations of the international working group for diagnosis, standardization of response criteria, treatment outcomes, and reporting standards for therapeutic trials in acute myeloid leukemia. J. Clin. Oncol. 2003, 21, 4642–4649. [Google Scholar] [CrossRef] [PubMed]
- Cheson, B.D.; Greenberg, P.L.; Bennett, J.M.; Lowenberg, B.; Wijermans, P.W.; Nimer, S.D.; Pinto, A.; Beran, M.; de Witte, T.M.; Stone, R.M.; et al. Clinical application and proposal for modification of the International Working Group (IWG) response criteria in myelodysplasia. Blood 2006, 108, 419–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, A.M.; Bridgewater, B.R.; Liu, Q.; Mitchell, M.W.; Robinson, R.J.; Dai, H.; Stewart, S.J.; DeHaven, C.D.; Miller, L.A.D. High resolution mass spectrometry improves data quantity and quality as compared to unit mass resolution mass spectrometry in high-throughput profiling metabolomics. Metabolomics 2014, 4, 132. [Google Scholar]
- Hernandez-Corbacho, M.J.; Salama, M.F.; Canals, D.; Senkal, C.E.; Obeid, L.M. Sphingolipids in mitochondria. Biochim. Et Biophys. Acta. Mol. Cell Biol. Lipids 2017, 1862, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef] [Green Version]
- Wanders, R.J.A.; Komen, J.; Kemp, S. Fatty acid omega-oxidation as a rescue pathway for fatty acid oxidation disorders in humans. Febs J. 2011, 278, 182–194. [Google Scholar] [CrossRef]
- Maher, M.; Diesch, J.; Casquero, R.; Buschbeck, M. Epigenetic-transcriptional regulation of fatty acid metabolism and its alterations in leukaemia. Front. Genet. 2018, 9, 405. [Google Scholar] [CrossRef]
- Yan, F.; Shen, N.; Pang, J.X.; Zhao, N.; Zhang, Y.W.; Bode, A.M.; Al-Kali, A.; Litzow, M.R.; Li, B.; Liu, S.J. A vicious loop of fatty acid-binding protein 4 and DNA methyltransferase 1 promotes acute myeloid leukemia and acts as a therapeutic target. Leukemia 2018, 32, 865–873. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [Green Version]
- Cheson, B.D.; Bennett, J.M.; Kantarjian, H.; Pinto, A.; Schiffer, C.A.; Nimer, S.D.; Lowenberg, B.; Beran, M.; de Witte, T.M.; Stone, R.M.; et al. Report of an international working group to standardize response criteria for myelodysplastic syndromes. Blood 2000, 96, 3671–3674. [Google Scholar] [PubMed]
- Deschler, B.; de Witte, T.; Mertelsmann, R.; Lübbert, M. Treatment decision-making for older patients with high-risk myelodysplastic syndrome or acute myeloid leukemia: Problems and approaches. 2006, 91, 1513–1522. Haematologica 2006, 91, 1513–1522. [Google Scholar] [PubMed]
- Menzin, J.; Lang, K.; Earle, C.C.; Kerney, D.; Mallick, R. The outcomes and costs of acute myeloid leukemia among the elderly. Arch. Intern. Med. 2002, 162, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Siminska, E.; Koba, M. Amino acid profiling as a method of discovering biomarkers for early diagnosis of cancer. Amino Acids 2016, 48, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.B.; Tang, S.Y. Folate status and colorectal cancer risk: A 2016 update. Mol. Asp. Med. 2017, 53, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Gao, F.; Ma, J.L.; Zhang, X.Z.; Li, Y.; Song, L.P.; Zhao, D.L. Analysis of the characteristics and prognosis of advanced non-small-cell lung cancer in older patients. Patient Prefer. Adher. 2015, 9, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Chen, T.X.; Fu, S.Z.; Sun, X.; Wang, L.Y.; Wang, J.; Lu, Y.F.; Ding, S.M.; Ruan, G.D.; Teng, L.S.; et al. Perioperative dynamics and significance of amino acid profiles in patients with cancer. J. Transl. Med. 2015, 13. [Google Scholar] [CrossRef]
- Argiles, J.M.; Busquets, S.; Stemmler, B.; Lopez-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef]
- Wu, Y.; Hurren, R.; MacLean, N.; Gronda, M.; Jitkova, Y.; Sukhai, M.A.; Minden, M.D.; Schimmer, A.D. Carnitine transporter CT2 (SLC22A16) is over-expressed in acute myeloid leukemia (AML) and target knockdown reduces growth and viability of AML cells. Apoptosis 2015, 20, 1099–1108. [Google Scholar] [CrossRef]
- Masoodi, M.; Lee, E.; Eiden, M.; Bahlo, A.; Shi, Y.; Ceddia, R.B.; Baccei, C.; Prasit, P.; Spaner, D.E. A role for oleoylethanolamide in chronic lymphocytic leukemia. Leukemia 2014, 28, 1381–1387. [Google Scholar] [CrossRef]
- Guzman, M.; Lo Verme, J.; Fu, J.; Oveisi, F.; Blazquez, C.; Piomelli, D. Oleoylethanolamide stimulates lipolysis by activating the nuclear receptor peroxisome proliferator-activated receptor alpha (PPAR-alpha). J. Biol. Chem. 2004, 279, 27849–27854. [Google Scholar] [CrossRef] [PubMed]
- Eder, K.; Schleser, S.; Becker, K.; Korting, R. Conjugated linoleic acids lower the release of eicosanoids and nitric oxide from human aortic endothelial cells. J. Nutr. 2003, 133, 4083–4089. [Google Scholar] [CrossRef] [PubMed]
- Ogretmen, B.; Hannun, Y.A. Biologically active sphingolipids in cancer pathogenesis and treatment. Nat. Rev. Cancer 2004, 4, 604. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Okimoto, R.A.; Purton, L.E.; Goodwin, M.; Haserlat, S.M.; Dayyani, F.; Sweetser, D.A.; McClatchey, A.I.; Bernard, O.A.; Look, A.T.; et al. Mutations in the neutral sphingomyelinase gene SMPD3 implicate the ceramide pathway in human leukemias. Blood 2008, 111, 4716–4722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagan, N.; Zoeller, R.A. Plasmalogens: Biosynthesis and functions. Prog. Lipid Res. 2001, 40, 199–229. [Google Scholar] [CrossRef]
- Morandat, S.; Bortolato, M.; Anker, G.; Doutheau, A.; Lagarde, M.; Chauvet, J.-P.; Roux, B. Plasmalogens protect unsaturated lipids against UV-induced oxidation in monolayer. Biochim. Et Biophys. Acta (Bba) - Biomembr. 2003, 1616, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Ozias, M.K.; Schalinske, K.L. All-trans-retinoic acid rapidly induces glycine N-methyltransferase in a dose-dependent manner and reduces circulating methionine and homocysteine levels in rats. J. Nutr. 2003, 133, 4090–4094. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Blok, B.A.; van Crevel, R.; Joosten, L.A.B.; Aaby, P.; Benn, C.S.; Netea, M.G. Vitamin A induces inhibitory histone methylation modifications and down-regulates trained immunity in human monocytes. J. Leukoc. Biol. 2015, 98, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Miftakhova, R.; Sandberg, T.; Hedblom, A.; Nevzorova, T.; Persson, J.L.; Bredberg, A. DNA methylation in ATRA-treated leukemia cell lines lacking a PML–RAR chromosome translocation. Anticancer Res. 2012, 32, 4715–4722. [Google Scholar]
- Yan, F.; Shen, N.; Pang, J.X.; Zhang, Y.W.; Rao, E.Y.; Bode, A.M.; Al-Kali, A.; Zhang, D.E.; Litzow, M.R.; Li, B.; et al. Fatty acid-binding protein FABP4 mechanistically links obesity with aggressive AML by enhancing aberrant DNA methylation in AML cells. Leukemia 2017, 31, 1434–1442. [Google Scholar] [CrossRef]
- Corbet, C.; Feron, O. Emerging roles of lipid metabolism in cancer progression. Curr. Opin. Clin. Nutr. 2017, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]
- German, N.J.; Yoon, H.; Yusuf, R.Z.; Murphy, J.P.; Finley, L.W.S.; Laurent, G.; Haas, W.; Satterstrom, F.K.; Guarnerio, J.; Zaganjor, E.; et al. PHD3 loss in cancer enables metabolic reliance on fatty acid oxidation via deactivation of ACC2. Mol. Cell. 2016, 63, 1006–1020. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Name | Classification | During VPA Therapy/Pretherapy | ||
|---|---|---|---|---|
| p-Value | q-Value | Fold Change | ||
| RESPONDERS | ||||
| *Valproate | Drug concentration | 0.0000 | 0.0000 | |
| *2-propyl-2-pentenoate (2-ene-valproate) | Valproic acid metabolite | 0.0000 | 0.0000 | 55.81 |
| *5-hydroxyvalproate | Valproic acid metabolite | 0.0000 | 0.0000 | 13.83 |
| *3-hydroxyvalproate | Valproic acid metabolite | 0.0000 | 0.0000 | 25.77 |
| *Glucuronide of C8H16O2 (1)* | Partially characterized | 0.0000 | 0.0000 | 28.34 |
| *Suberoylcarnitine (C8-DC) | Fatty acid metabolism, acyl carnitine | 0.0000 | 0.0000 | 30.82 |
| Phenylacetylcarnitine | Acetylated peptide | 0.0000 | 0.0000 | 17.30 |
| *Adipoylcarnitine (C6-DC) | Fatty acid metabolism | 0.0000 | 0.0000 | 17.06 |
| *3-methylglutarylcarnitine (2) | Leucine/isoleucin/valine metabolism | 0.0000 | 0.0000 | 0.18 |
| *Adipate (C6-DC) | Fatty acid, dicarboxylate | 0.0000 | 0.0027 | 3.41 |
| 10-undecenoate (11:1n1) | Medium chain fatty acid | 0.0001 | 0.0043 | 0.49 |
| *Isobutyrylca′rnitine (C4) | Leucine/isoleucin/valine metabolism | 0.0001 | 0.0093 | 3.06 |
| Glucuronide of C14H22O4 (2) | Partially characterized | 0.0003 | 0.0182 | 0.22 |
| 4-hydroxycinnamate sulfate | Tyrosine metabolism | 0.0004 | 0.0259 | 3.00 |
| Isoeugenol sulfate | Food component, plant | 0.0005 | 0.0264 | 0.04 |
| NON-RESPONDERS | ||||
| *Valproate | Drug concentration | 0.0000 | 0.0000 | |
| *2-propyl-2-pentenoate (2-ene-valproate) | Valproic acic metabolite | 0.0000 | 0.0000 | 48.32 |
| *5-hydroxyvalproate | Valproic acic metabolite | 0.0000 | 0.0000 | 31.58 |
| *3-hydroxyvalproate | Valproic acic metabolite | 0.0000 | 0.0000 | 93.38 |
| *Glucuronide of C8H16O2 (1) | Partially characterized | 0.0000 | 0.0000 | 16.16 |
| *Suberoylcarnitine (C8-DC) | Fatty acid, dicarboxylate | 0.0000 | 0.0000 | 12.96 |
| *Adipoylcarnitine (C6-DC) | Fatty acid metabolism | 0.0000 | 0.0000 | 7.34 |
| *Adipate (C6-DC) | Fatty acid, dicarboxylate | 0.0000 | 0.0001 | 4.33 |
| *3-methylglutarylcarnitine (2) | Leucine/isoleucin/valine metabolism | 0.0000 | 0.0001 | 0.25 |
| Suberate (C8-DC) | Fatty acid, dicarboxylate | 0.0000 | 0.0022 | 3.43 |
| 3-carboxy-4-methyl-5-pentyl-2-Furanpropionate (3-Cmpfp) | Fatty acid metabolism, dicarboxylate | 0.0001 | 0.0054 | 0.44 |
| Hexanoylglycine | Fatty acid metabolism, acyl glycine | 0.0001 | 0.0077 | 3.03 |
| 3-hydroxyhexanoate | Fatty acid metabolism, monohydroxy | 0.0001 | 0.0100 | 2.03 |
| Androstenediol (3beta,17beta) disulfate (1) | Androgenic steroid | 0.0003 | 0.0197 | 1.45 |
| *Isobutyrylcarnitine (C4) | Leucine/isoleucin/valine metabolism | 0.0004 | 0.0208 | 3.15 |
| Gamma-CEHC | Cofactors/vitamins | 0.0004 | 0.0234 | 1.56 |
| Isoursodeoxycholate | Secondary bile acid metabolism | 0.0005 | 0.0278 | 0.06 |
| Indoleacetylglutamine | Tryptophane metabolism | 0.0006 | 0.0298 | 0.09 |
| 4-hydroxyphenylacetylglutamine | Acetylated peptide | 0.0008 | 0.0352 | 0.28 |
| 1-palmitoyl-2-linoleoyl-GPI (16:0/18:2) | Phosphatidylinositole | 0.0008 | 0.0352 | 1.51 |
| 5-bromotryptophan | Tryptophane metabolism | 0.0010 | 0.0387 | 0.49 |
| 4-allylphenol sulfate | Food component, plant | 0.0010 | 0.0387 | 0.29 |
| N-acetyltyrosine | Tyrosine metabolism | 0.0011 | 0.0413 | 2.42 |
| Trans-4-hydroxyproline | Urea cycle. Proline metabolism | 0.0011 | 0.0411 | 1.73 |
| Galactonate | Carbohydrate metabolism | 0.0012 | 0.0430 | 0.38 |
| 3-aminoisobutyrate | Pyrimidine metabolism, thymine | 0.0013 | 0.0433 | 1.86 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grønningsæter, I.S.; Fredly, H.K.; Gjertsen, B.T.; Hatfield, K.J.; Bruserud, Ø. Systemic Metabolomic Profiling of Acute Myeloid Leukemia Patients before and During Disease-Stabilizing Treatment Based on All-Trans Retinoic Acid, Valproic Acid, and Low-Dose Chemotherapy. Cells 2019, 8, 1229. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8101229
Grønningsæter IS, Fredly HK, Gjertsen BT, Hatfield KJ, Bruserud Ø. Systemic Metabolomic Profiling of Acute Myeloid Leukemia Patients before and During Disease-Stabilizing Treatment Based on All-Trans Retinoic Acid, Valproic Acid, and Low-Dose Chemotherapy. Cells. 2019; 8(10):1229. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8101229
Chicago/Turabian StyleGrønningsæter, Ida Sofie, Hanne Kristin Fredly, Bjørn Tore Gjertsen, Kimberley Joanne Hatfield, and Øystein Bruserud. 2019. "Systemic Metabolomic Profiling of Acute Myeloid Leukemia Patients before and During Disease-Stabilizing Treatment Based on All-Trans Retinoic Acid, Valproic Acid, and Low-Dose Chemotherapy" Cells 8, no. 10: 1229. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8101229