Mapping Research in the Obesity, Adipose Tissue, and MicroRNA Field: A Bibliometric Analysis

 , ,
, ,  and
and
Abstract
:1. Introduction
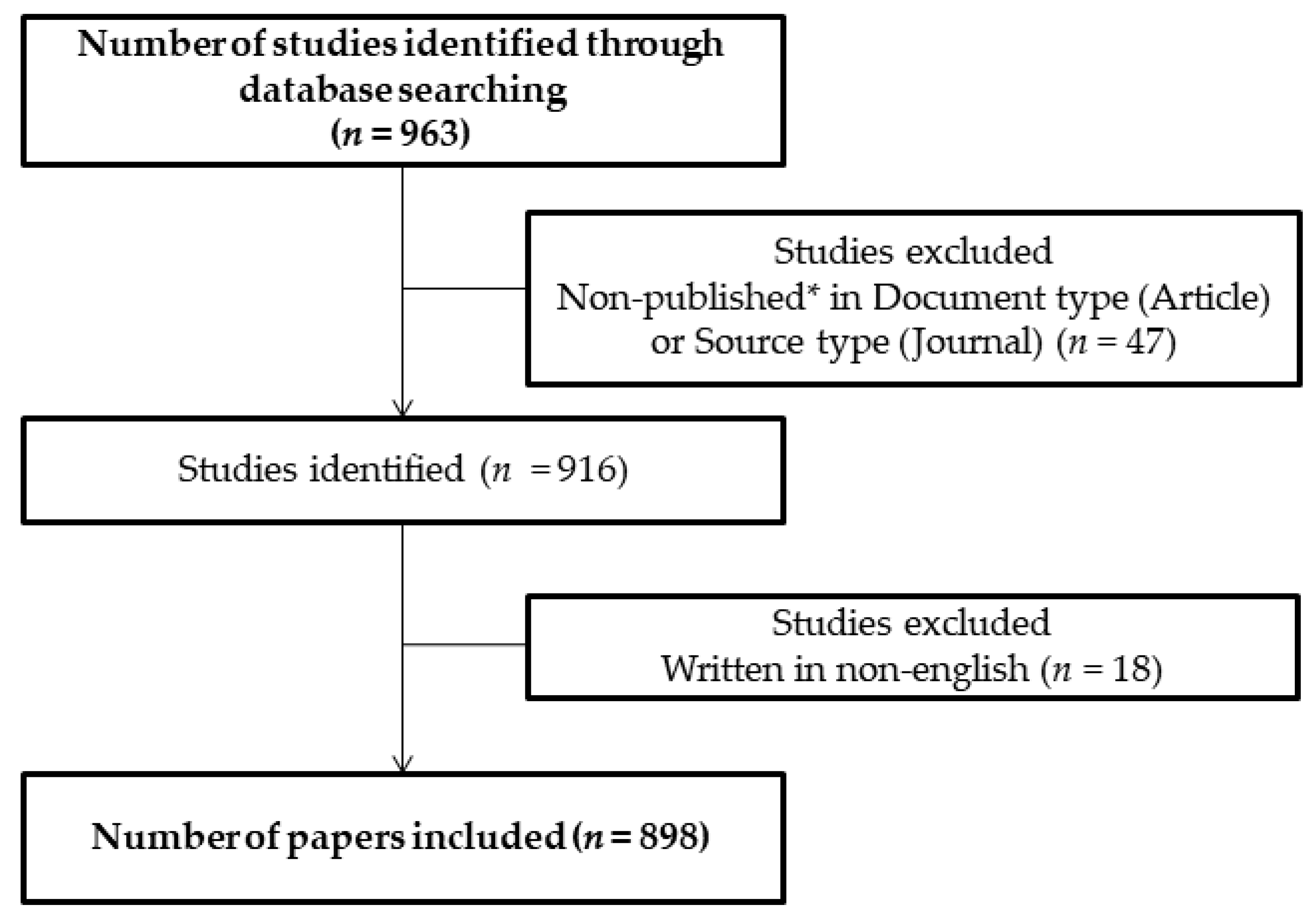
2. Materials and Methods
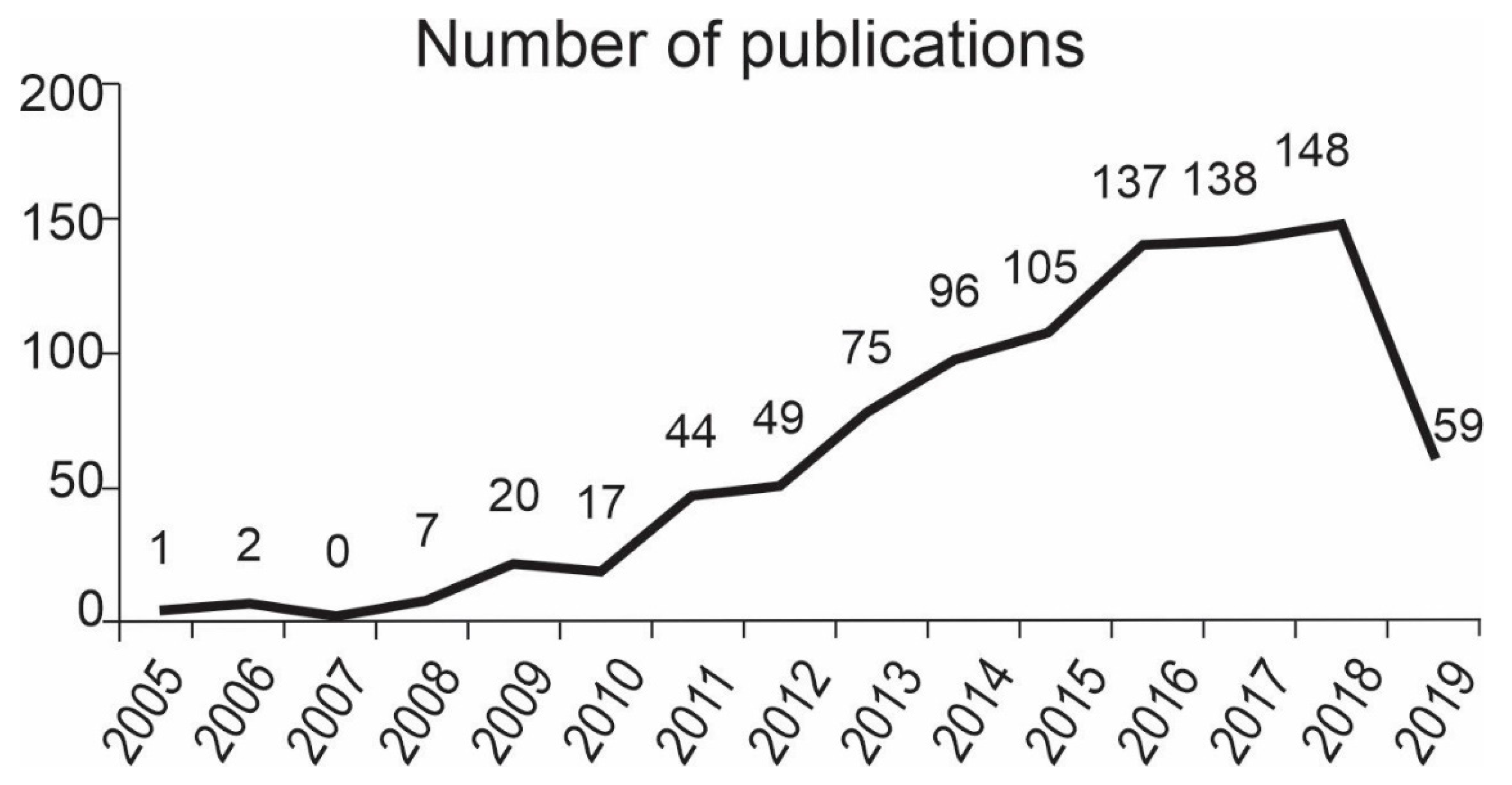
3. General Information
4. Bibliometric Variables
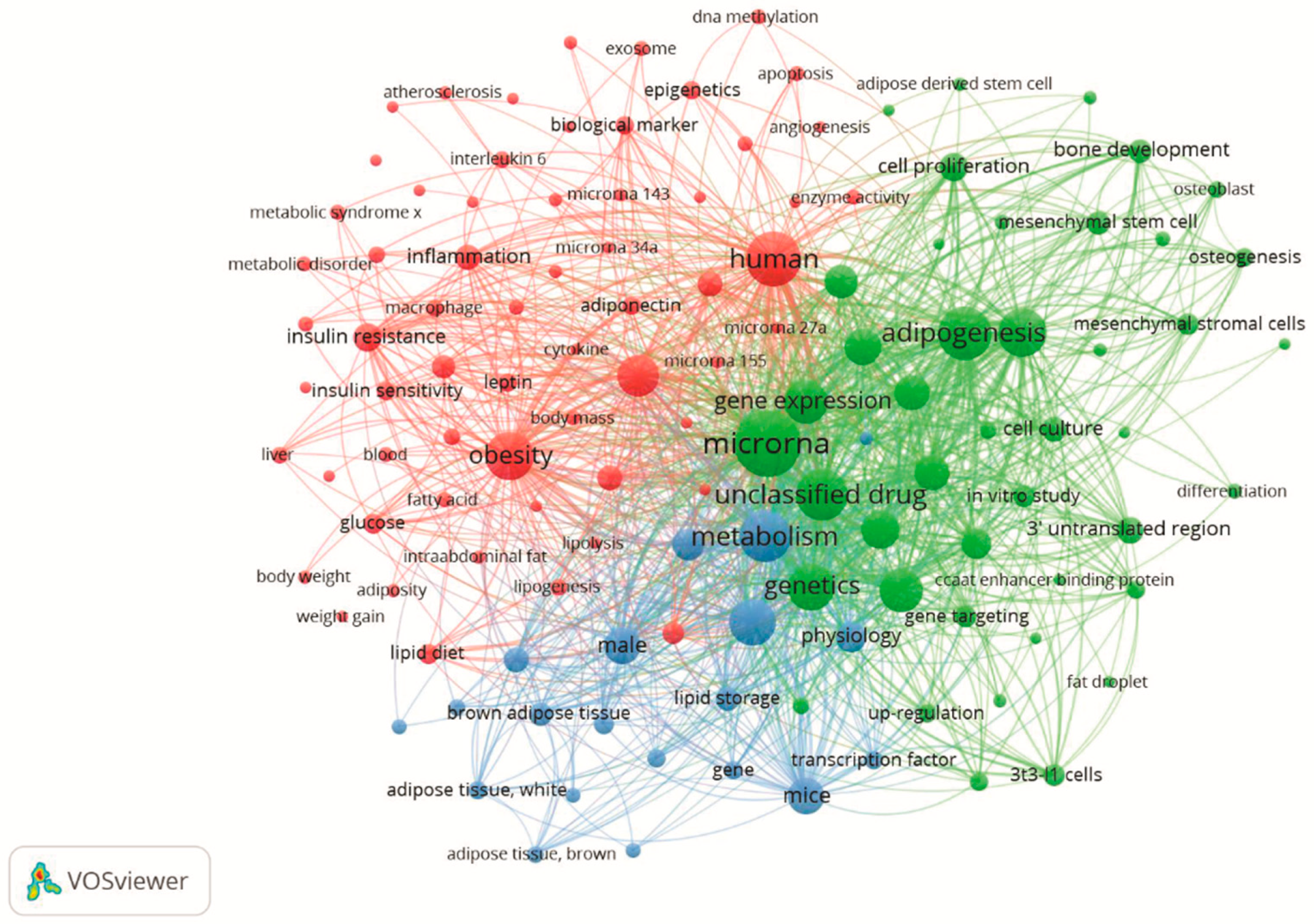
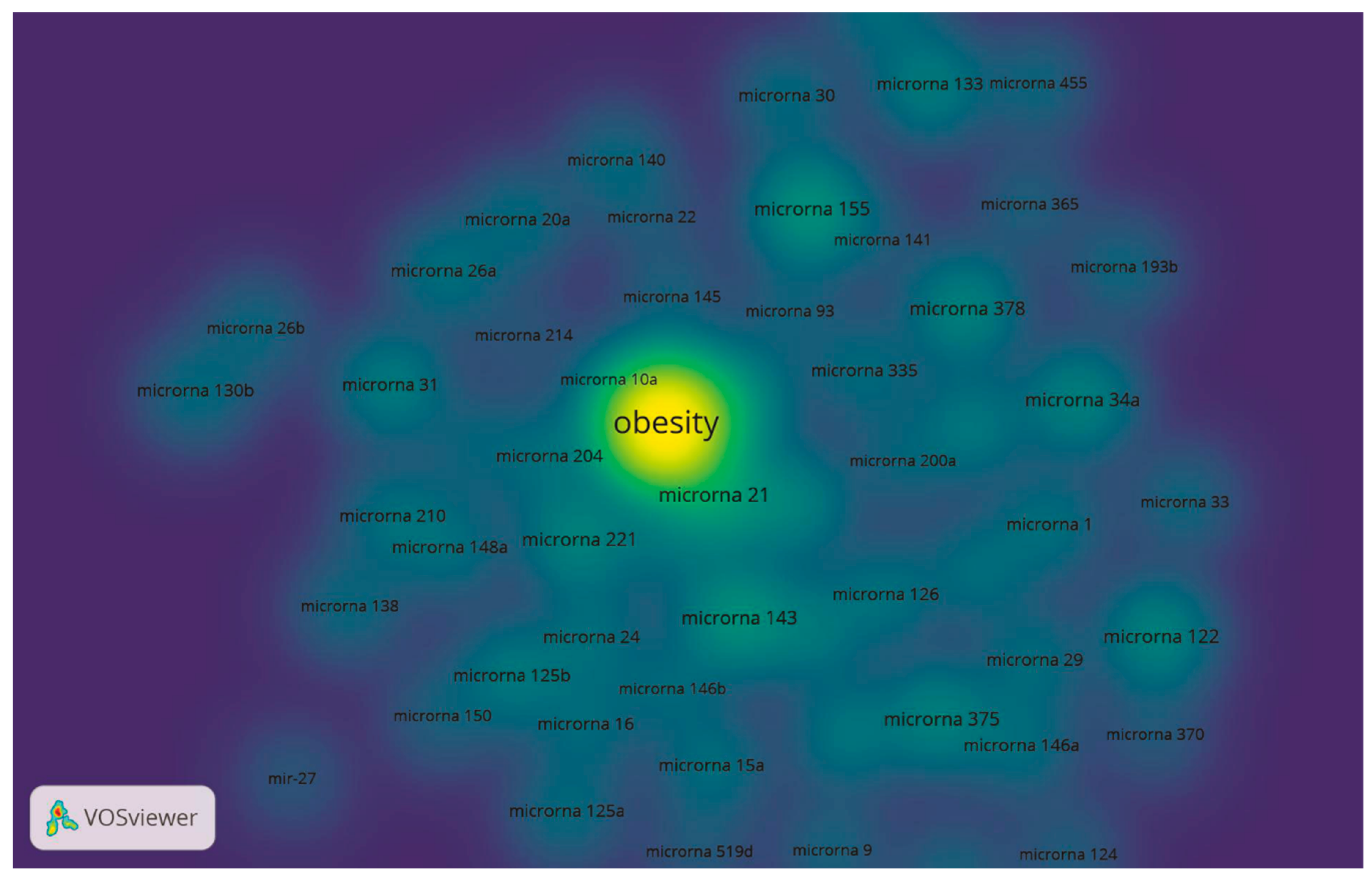
5. Hotspots
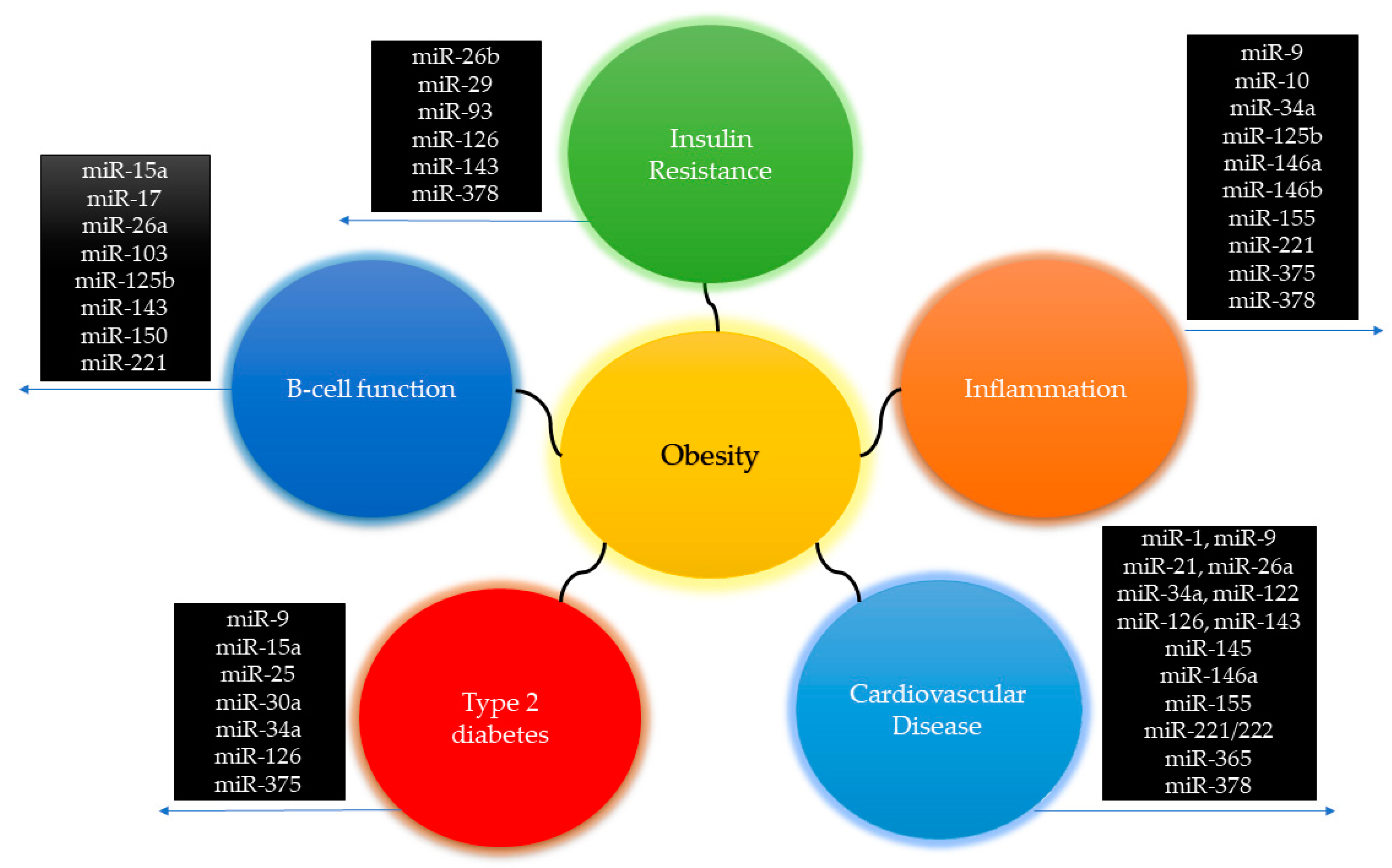
5.1. Red Cluster: Obesity, Inflammation, Type 2 Diabetes and Cardiovascular Disease: A Perspective through miRNAs
5.2. Green Cluster: Adipogenic Process and miRNAs
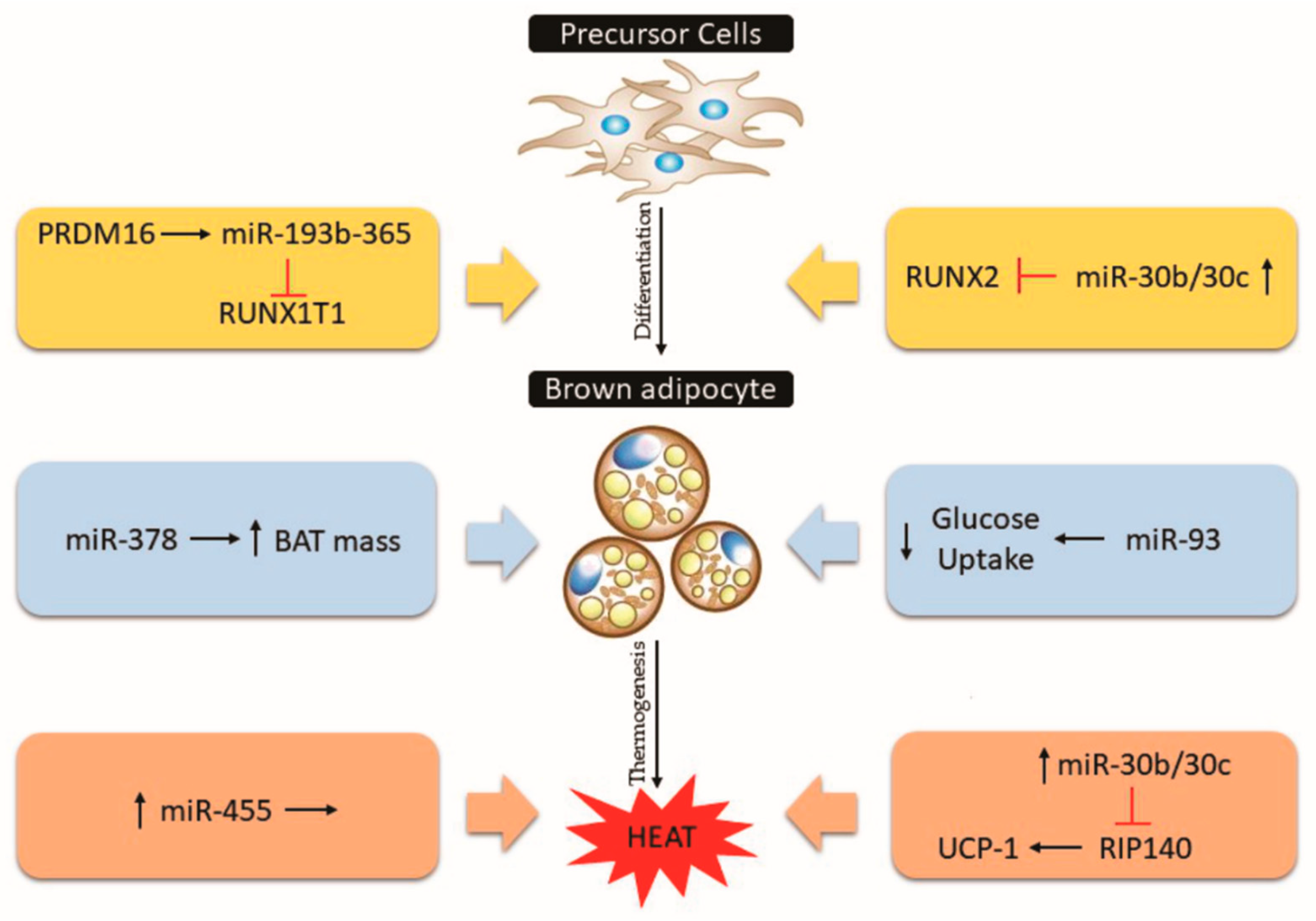
5.3. Blue Cluster: MiRNAs in the Control of Brown Adipose Tissue and Thermogenesis
5.3.1. MiR-193b/miR-365, miR-30, miR-455, miR-378
5.3.2. MiR-93, miR-155, miR-34a
6. Strengths and Limitations
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; Naghavi, M.; et al. GBD 2015 Obesity Collaborators: Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, Regional, and National Prevalence of Overweight and Obesity in Children and Adults during 1980–2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2014, 6736, 766–781. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Obesity and Overweight: Fact Sheet 2016. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 21 November 2019).
- WOD. World Obesity Day 2017. Available online: https:// www.obesityday.worldobesity.org/world-obesity-day-2017 (accessed on 21 November 2019).
- Schoettl, T.; Fischer, I.P.; Ussar, S. Heterogeneity of Adipose Tissue in Development and Metabolic Function. J. Exp. Biol. 2018, 221, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gesta, S.; Kahn, C.R. White Adipose Tissue. In Adipose Tissue Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 149–199. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. What We Talk about When We Talk about Fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, C.A.; Da Silva, K.A.; Rocha, M.C.; Sene-Fiorese, M.; Nonaka, K.O.; Malavazi, I.; De Freitas Anibal, F.; De Oliveira Duarte, A.C.G. Exercise and Omentin: Their Role in the Crosstalk between Muscle and Adipose Tissues in Type 2 Diabetes Mellitusrat Models. Front. Physiol. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-Derived Circulating MiRNAs Regulate Gene Expression in Other Tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef]
- Ahima, R.S.; Lazar, M.A. Adipokines and the Peripheral and Neural Control of Energy Balance. Mol. Endocrinol. 2008, 22, 1023–1031. [Google Scholar] [CrossRef]
- Wensveen, F.M.; Jelenčić, V.; Valentić, S.; Šestan, M.; Wensveen, T.T.; Theurich, S.; Glasner, A.; Mendrila, D.; Štimac, D.; Wunderlich, F.T.; et al. NK Cells Link Obesity-Induced Adipose Stress to Inflammation and Insulin Resistance. Nat. Immunol. 2015, 16, 376–385. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Carobbio, S.; Vidal-Puig, A. Adipose Tissue Plasticity: How Fat Depots Respond Differently to Pathophysiological Cues. Diabetologia 2016, 59, 1075–1088. [Google Scholar] [CrossRef] [Green Version]
- Ellies, L.G.; Johnson, A.; Olefsky, J.M. Obesity, Inflammation, and Insulin Resistance. Obes. Inflamm. Cancer 2013, 1, 1–23. [Google Scholar] [CrossRef]
- Landsberg, L.; Aronne, L.J.; Beilin, L.J.; Burke, V.; Igel, L.I.; Lloyd-Jones, D.; Sowers, J. Obesity-Related Hypertension: Pathogenesis, Cardiovascular Risk, and Treatment-a Position Paper of the the Obesity Society and the American Society of Hypertension. Obesity 2013, 15, 14–33. [Google Scholar] [CrossRef] [PubMed]
- De Pergola, G.; Silvestris, F. Obesity as a Major Risk Factor for Cancer. J. Obes. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Kahn, C.R. Brown Fat as a Therapy for Obesity and Diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 143–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown Adipose Tissue Activity Controls Triglyceride Clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Sidossis, L.; Kajimura, S. Brown and Beige Fat in Humans: Thermogenic Adipocytes That Control Energy and Glucose Homeostasis. J. Clin. Investig. 2015, 125, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Heeren, J. Adipose Tissue Browning and Metabolic Health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Hobert, O. Gene Regulation by Transcription Factors and MicroRNAs. Science 2008, 319, 1785–1786. [Google Scholar] [CrossRef]
- Pasquinelli, A.E. MicroRNAs and Their Targets: Recognition, Regulation and an Emerging Reciprocal Relationship. Nat. Rev. Genet. 2012, 13, 271–282. [Google Scholar] [CrossRef]
- Arner, P.; Kulyté, A. MicroRNA Regulatory Networks in Human Adipose Tissue and Obesity. Nat. Rev. Endocrinol. 2015, 11, 276–288. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological Functions of MicroRNAs: A Review. J. Physiol. Biochem. 2011, 67, 129–139. [Google Scholar] [CrossRef] [PubMed]
- del Carmen Martínez-Jiménez, V.; Méndez-Mancilla, A.; Patricia Portales-Pérez, D. MiRNAs in Nutrition, Obesity, and Cancer: The Biology of MiRNAs in Metabolic Disorders and Its Relationship with Cancer Development. Mol. Nutr. Food Res. 2018, 62, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Lim, B.; Lodish, H.F. MicroRNAs Induced during Adipogenesis That Accelerate Fat Cell Development Are Downregulated in Obesity. Diabetes 2009, 58, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in Metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef]
- Landrier, J.F.; Derghal, A.; Karkeni, E.; Mounien, L. MicroRNAs in Endocrine Disorders. Rsc. Drug Discov. Ser. 2019, 254–274. [Google Scholar] [CrossRef]
- Iacomino, G.; Siani, A. Role of MicroRNAs in Obesity and Obesity-Related Diseases. Genes Nutr. 2017, 12, 1–16. [Google Scholar] [CrossRef]
- Rottiers, V.; Näär, A.M. MicroRNAs in Metabolism and Metabolic Disorders. Nat. Rev. Mol. Cell Biol. 2012, 13, 239–251. [Google Scholar] [CrossRef]
- Estep, M.; Armistead, D.; Hossain, N.; Elarainy, H.; Goodman, Z.; Baranova, A.; Chandhoke, V.; Younossi, Z.M. Differential Expression of MiRNAs in the Visceral Adipose Tissue of Patients with Non-Alcoholic Fatty Liver Disease. Aliment. Pharm. 2010, 32, 487–497. [Google Scholar] [CrossRef]
- Dumortier, O.; Hinault, C.; Van Obberghen, E. MicroRNAs and Metabolism Crosstalk in Energy Homeostasis. Cell Metab. 2013, 18, 312–324. [Google Scholar] [CrossRef] [Green Version]
- Marques-Rocha, J.L.; Samblas, M.; Milagro, F.I.; Bressan, J.; Martínez, J.A.; Marti, A. Noncoding RNAs, Cytokines, and Inflammation-Related Diseases. Faseb J. 2015, 29, 3595–3611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.L.; Shi, C.M.; Xu, G.F.; Chen, L.; Zhu, L.L.; Zhu, L.; Guo, X.R.; Xu, M.Y.; Ji, C.B. TNF-α, IL-6, and Leptin Increase the Expression of MiR-378, an Adipogenesis-Related MicroRNA in Human Adipocytes. Cell Biochem. Biophys. 2014, 70, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Ma, M.; Liang, T.; Guo, L. Role of MicroRNAs in Obesity-Induced Metabolic Disorder and Immune Response. J. Immunol. Res. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heneghan, H.M.; Miller, N.; Kerin, M.J. Role of MicroRNAs in Obesity and the Metabolic Syndrome. Obes. Rev. 2010, 11, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, J.; Cui, J.; Hou, J.; Zheng, X.; Li, C.; Liu, L. Micrornas Regulate Adipocyte Differentiation. Cell Biol. Int. 2013, 37, 533–546. [Google Scholar] [CrossRef]
- Kilic, I.D.; Dodurga, Y.; Uludag, B.; Alihanoglu, Y.I.; Yildiz, B.S.; Enli, Y.; Secme, M.; Bostanci, H.E. MicroRNA -143 and -223 in Obesity. Gene 2015, 560, 140–142. [Google Scholar] [CrossRef]
- Esau, C.; Kang, X.; Peralta, E.; Hanson, E.; Marcusson, E.G.; Ravichandran, L.V.; Sun, Y.; Koo, S.; Perera, R.J.; Jain, R.; et al. MicroRNA-143 Regulates Adipocyte Differentiation. J. Biol. Chem. 2004, 279, 52361–52365. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.-Y.; Wen, G.-B.; Feng, S.-D.; Tuo, Q.-H.; Ou, H.-S.; Yao, C.H.; Zhu, B.-Y.; Gao, Z.-P.; Zhang, L.; Liao, D.-F. MicroRNA-375 Promotes 3T3-L1 Adipocyte Differentiation through Modulation of Extracellular Signal-Regulated Kinase Signalling. Clin. Exp. Pharm. Physiol. 2011, 38, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Koh, E.H.; Chernis, N.; Saha, P.K.; Xiao, L.; Bader, D.A.; Zhu, B.; Rajapakshe, K.; Hamilton, M.P.; Liu, X.; Perera, D.; et al. MiR-30a Remodels Subcutaneous Adipose Tissue Inflammation to Improve Insulin Sensitivity in Obesity. Diabetes 2018, 67, 2541–2553. [Google Scholar] [CrossRef] [Green Version]
- Klöting, N.; Berthold, S.; Kovacs, P.; Schön, M.R.; Fasshauer, M.; Ruschke, K.; Stumvoll, M.; Blüher, M. MicroRNA Expression in Human Omental and Subcutaneous Adipose Tissue. PLoS ONE 2009, 4, e4699. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Choudhury, N.; Uddin, S.; Hossain, L.; Baur, L.A. Longitudinal Trends in Global Obesity Research and Collaboration: A Review Using Bibliometric Metadata. Obes. Rev. 2016, 17, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Teles, R.H.G.; Moralles, H.F.; Cominetti, M.R. Global Trends in Nanomedicine Research on Triple Negative Breast Cancer: A Bibliometric Analysis. Int. J. Nanomed. 2018, 13, 2321–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackl, H.; Burkard, T.R.; Sturn, A.; Rubio, R.; Schleiffer, A.; Tian, S.; Quackenbush, J.; Eisenhaber, F.; Trajanoski, Z. Molecular Processes during Fat Cell Development Revealed by Gene Expression Profiling and Functional Annotation. Genome Biol. 2005, 6, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Zhang, M.; Tong, M.; Yang, L.; Pang, L.; Chen, L.; Xu, G.; Chi, X.; Hong, Q.; Ni, Y.; et al. MiR-148a Is Associated with Obesity and Modulates Adipocyte Differentiation of Mesenchymal Stem Cells through Wnt Signaling. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, W.; Horn, P.; Castoldi, M.; Diehlmann, A.; Bork, S.; Saffrich, R.; Benes, V.; Blake, J.; Pfister, S.; Eckstein, V.; et al. Replicative Senescence of Mesenchymal Stem Cells: A Continuous and Organized Process. PLoS ONE 2008, 3, e2213. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.; Pulgarín, A.; Escalona, M.I. Viewing Information Literacy Concepts: A Comparison of Two Branches of Knowledge. Scientometrics 2014, 98, 2311–2329. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Kajimura, S. Metabolic Adaptation and Maladaptation in Adipose Tissue. Nat. Metab. 2019, 1, 189–200. [Google Scholar] [CrossRef]
- Fonseca, V.; Desouza, C.; Asnani, S.; Jialal, I. Nontraditional Risk Factors for Cardiovascular Disease in Diabetes. Endocr. Rev. 2004, 25, 153–175. [Google Scholar] [CrossRef] [Green Version]
- Hulsmans, M.; De Keyzer, D.; Holvoet, P. MicroRNAs Regulating Oxidative Stress and Inflammation in Relation to Obesity and Atherosclerosis. Faseb J. 2011, 25, 2515–2527. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Pivarcsi, A. MicroRNAs in Inflammation. Int. Rev. Immunol. 2009, 1, 535–561. [Google Scholar] [CrossRef]
- Javidan, A.; Jiang, W.; Okuyama, M.; Thiagarajan, D.; Yang, L.; Moorleghen, J.J.; Muniappan, L.; Subramanian, V. MiR-146a Deficiency Accelerates Hepatic Inflammation Without Influencing Diet-Induced Obesity in Mice. Sci. Rep. 2019, 9, 12626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Cárdenas, M.; Uresti-Rivera, E.E.; Cortés-García, J.D.; Briones-Espinoza, M.; Ruíz-Rodríguez, V.M.; Reynaga-Hernández, E.; Mendez-Mancilla, A.; Portales-Pérez, D.P. Increased Levels of Adipose Tissue-Resident Th17 Cells in Obesity Associated with MiR-326. Immunol. Lett. 2019, 211, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Karkeni, E.; Astier, J.; Tourniaire, F.; El Abed, M.; Romier, B.; Gouranton, E.; Wan, L.; Borel, P.; Salles, J.; Walrand, S.; et al. Obesity-Associated Inflammation Induces MicroRNA-155 Expression in Adipocytes and Adipose Tissue: Outcome on Adipocyte Function. J. Clin. Endocrinol. Metab. 2016, 101, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
- Ceppi, M.; Pereira, A.M.; Dunand-Sauthier, I.; Barras, E.; Reith, W.; Santos, M.A.; Pierre, P. MicroRNA-155 Modulates the Interleukin-1 Signaling Pathway in Activated Human Monocyte-Derived Dendritic Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2735–2740. [Google Scholar] [CrossRef] [Green Version]
- López, P.; Castro, A.; Flórez, M.; Miranda, K.; Aranda, P.; Sánchez-González, C.; Llopis, J.; Arredondo, M. MiR-155 and MiR-122 Expression of Spermatozoa in Obese Subjects. Front. Genet. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Zhang, J.; Chen, X.; Zhang, J.; Ding, X.; You, X.; Fan, L.; Chen, C.; Zhou, Y. MicroRNA-155 Mediates Obesity-Induced Renal Inflammation and Dysfunction. Inflammation 2019, 42, 994–1003. [Google Scholar] [CrossRef]
- Pan, Y.; Hui, X.; Chong Hoo, R.L.; Ye, D.; Cheung Chan, C.Y.; Feng, T.; Wang, Y.; Ling Lam, K.S.; Xu, A. Adipocyte-Secreted Exosomal MicroRNA-34a Inhibits M2 Macrophage Polarization to Promote Obesity-Induced Adipose Inflammation. J. Clin. Invest. 2019, 129, 834–849. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Feng, T.; Chan, Y.-C.; Xu, A.; Lam, K. OR01-3 MicroRNA-34a-Mediated FGF21 Resistance in the Adipose Tissue Contributes to Insulin Resistance and Hypoadiponectinemia in Diet-Induced Obesity. J. Endocr. Soc. 2019, 3 (Suppl. 1). [Google Scholar] [CrossRef]
- Xiao, H.; Li, H.; Yu, G.; Xiao, W.; Hu, J.; Tang, K.; Zeng, J.; He, W.; Zeng, G.; Ye, Z.; et al. MicroRNA-10b Promotes Migration and Invasion through KLF4 and HOXD10 in Human Bladder Cancer. Oncol. Rep. 2014, 31, 1832–1838. [Google Scholar] [CrossRef] [Green Version]
- Boldin, M.P.; Baltimore, D. MicroRNAs, New Effectors and Regulators of NF-ΚB. Immunol. Rev. 2012, 246, 205–220. [Google Scholar] [CrossRef]
- Estep, J.M.; Goodman, Z.; Sharma, H.; Younossi, E.; Elarainy, H.; Baranova, A.; Younossi, Z. Adipocytokine Expression Associated with MiRNA Regulation and Diagnosis of NASH in Obese Patients with NAFLD. Liver Int. 2015, 35, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B. Mitochondrial Dysfunction and Type 2 Diabetes. Science 2005, 307, 384–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-W.; You, Y.-H.; Jung, S.; Suh-Kim, H.; Lee, I.-K.; Cho, J.-H.; Yoon, K.-H. MiRNA-30a-5p-Mediated Silencing of Beta2/NeuroD Expression Is an Important Initial Event of Glucotoxicity-Induced Beta Cell Dysfunction in Rodent Models. Diabetologia 2013, 56, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karolina, D.S.; Sepramaniam, S.; Tan, H.Z.; Armugam, A.; Jeyaseelan, K. MiR-25 and MiR-92a Regulate Insulin i Biosynthesis in Rats. RNA Biol. 2013, 10, 1365–1378. [Google Scholar] [CrossRef] [Green Version]
- LaPierre, M.P.; Stoffel, M. MicroRNAs as Stress Regulators in Pancreatic Beta Cells and Diabetes. Mol. Metab. 2017, 6, 1010–1023. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Katsiki, N.; Behnam, B.; Iranpanah, H.; Sahebkar, A. MicroRNAs and Type 2 Diabetes Mellitus: Molecular Mechanisms and the Effect of Antidiabetic Drug Treatment. Metabolism 2018, 1, 48–55. [Google Scholar] [CrossRef]
- Landrier, J.-F.; Derghal, A.; Mounien, L. MicroRNAs in Obesity and Related Metabolic Disorders. Cells 2019, 8, 859. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Lucena, R.; Camargo, A.; Alcalá-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; van Ommen, B.; Delgado-Lista, J.; Ordovás, J.M.; Pérez-Martínez, P.; Rangel-Zúñiga, O.A.; et al. A Plasma Circulating MiRNAs Profile Predicts Type 2 Diabetes Mellitus and Prediabetes: From the CORDIOPREV Study. Exp. Mol. Med. 2018, 50, 168. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Fan, J.; Chen, N. A Novel Regulator of Type II Diabetes: MicroRNA-143. Trends Endocrinol. Metab. 2018, 29, 380–388. [Google Scholar] [CrossRef]
- Ortega, F.J.; Moreno-Navarrete, J.M.; Pardo, G.; Sabater, M.; Hummel, M.; Ferrer, A.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. MiRNA Expression Profile of Human Subcutaneous Adipose and during Adipocyte Differentiation. PLoS ONE 2010, 5, e9022. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Zhou, Y.; Deng, Z.; Zhang, H.; Wu, Y.; Song, T.; Yang, Y.; Wei, H.; Peng, J. MiR-221 Negatively Regulates Inflammation and Insulin Sensitivity in White Adipose Tissue by Repression of Sirtuin-1 (SIRT1). J. Cell. Biochem. 2018, 119, 6418–6428. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, D.; Roy, U.; Garg, S.; Ghosh, S.; Pathak, S.; Kolthur-Seetharam, U. Sirt1 and Mir-9 Expression Is Regulated during Glucose-Stimulated Insulin Secretion in Pancreatic β-Islets. Febs J. 2011, 278, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Gok, O.; Karaali, Z.; Ergen, A.; Ekmekci, S.; Abaci, N. Serum Sirtuin 1 Protein as a Potential Biomarker for Type 2 Diabetes: Increased Expression of Sirtuin 1 and the Correlation with MicroRNAs. J. Res. Med. Sci. 2019, 25, 24–56. [Google Scholar] [CrossRef]
- Raman, S.; FitzGerald, U.; Murphy, J.M. Interplay of Inflammatory Mediators with Epigenetics and Cartilage Modifications in Osteoarthritis. Front. Bioeng. Biotechnol. 2018, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Koya, D. SIRT1 in Type 2 Diabetes: Mechanisms and Therapeutic Potential. Diabetes Metab. J. 2013, 37, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and Cardiovascular Disease: Pathophysiology, Evaluation, and Effect of Weight Loss: An Update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef] [Green Version]
- Quiat, D.; Olson, E.N. Review Series MicroRNAs in Cardiovascular Disease: From Pathogenesis to Prevention and Treatment. J. Clin. Invest. 2013, 123, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Hulsmans, M.; Holvoet, P. MicroRNAs as Early Biomarkers in Obesity and Related Metabolic and Cardiovascular Diseases. Curr. Pharm. Des. 2013, 19, 5704–5717. [Google Scholar] [CrossRef]
- Martinelli, R.; Nardelli, C.; Pilone, V.; Buonomo, T.; Liguori, R.; Castanò, I.; Buono, P.; Masone, S.; Persico, G.; Forestieri, P.; et al. MiR-519d Overexpression Is Associated with Human Obesity. Obesity 2010, 18, 2170–2176. [Google Scholar] [CrossRef]
- Kushner, R.F. Weight Loss Strategies for Treatment of Obesity. Prog. Cardiovasc. Dis. 2014, 56, 465–472. [Google Scholar] [CrossRef]
- Nardelli, C.; Iaffaldano, L.; Pilone, V.; Labruna, G.; Ferrigno, M.; Carlomagno, N.; Dodaro, C.A.; Forestieri, P.; Buono, P.; Salvatore, F.; et al. Changes in the MicroRNA Profile Observed in the Subcutaneous Adipose Tissue of Obese Patients after Laparoscopic Adjustable Gastric Banding. J. Obes. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.J.; Mercader, J.M.; Catalán, V.; Moreno-Navarrete, J.M.; Pueyo, N.; Sabater, M.; Gómez-Ambrosi, J.; Anglada, R.; Fernández-Formoso, J.A.; Ricart, W.; et al. Targeting the Circulating MicroRNA Signature of Obesity. Clin. Chem. 2013, 59, 781–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willeit, P.; Skroblin, P.; Moschen, A.R.; Yin, X.; Kaudewitz, D.; Zampetaki, A.; Barwari, T.; Whitehead, M.; Ramírez, C.M.; Goedeke, L.; et al. Circulating MicroRNA-122 Is Associated with the Risk of New-Onset Metabolic Syndrome and Type 2 Diabetes. Diabetes 2017, 66, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. MiR-122 Regulation of Lipid Metabolism Revealed by in Vivo Antisense Targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.D.; Cismowski, M.J.; Serpico, M.; Pusateri, A.; Brigstock, D.R. Elevation of Circulating MicroRNA Levels in Obese Children Compared to Healthy Controls. Clin. Obes. 2017, 7, 216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lv, C.; Li, L.; Chen, S.; Liu, S.; Wang, C.; Su, B. Plasma MiR-126 Is a Potential Biomarker for Early Prediction of Type 2 Diabetes Mellitus in Susceptible Individuals. Biomed. Res. Int. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma MicroRNA Profiling Reveals Loss of Endothelial MiR-126 and Other MicroRNAs in Type 2 Diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef]
- Fichtlscherer, S.; Zeiher, A.M.; Dimmeler, S. Circulating MicroRNAs: Biomarkers or Mediators of Cardiovascular Diseases? Arter. Thromb. Vasc. Biol. 2011, 31, 2383–2390. [Google Scholar] [CrossRef] [Green Version]
- Gomes, J.L.P.; Fernandes, T.; Soci, U.P.R.; Silveira, A.C.; Barretti, D.L.M.; Negrão, C.E.; Oliveira, E.M. Obesity Downregulates MicroRNA-126 Inducing Capillary Rarefaction in Skeletal Muscle: Effects of Aerobic Exercise Training. Oxid. Med. Cell. Longev. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of Tissue-Specific MicroRNAs from Mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef] [Green Version]
- Shan, Z.X.; Lin, Q.X.; Deng, C.Y.; Zhu, J.N.; Mai, L.P.; Liu, J.L.; Fu, Y.H.; Liu, X.Y.; Li, Y.X.; Zhang, Y.Y.; et al. MiR-1/MiR-206 Regulate Hsp60 Expression Contributing to Glucose-Mediated Apoptosis in Cardiomyocytes. Febs Lett. 2010, 584, 3592–3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From Stem Cell to Adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trauner, M.; Arrese, M.; Wagner, M. Fatty Liver and Lipotoxicity. Biochim. Et Biophys. Acta - Mol. Cell Biol. Lipids 2010, 1801, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and Metabolic Health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Huang, N.; Wang, J.; Xie, W.; Lyu, Q.; Wu, J.; He, J.; Qiu, W.; Xu, N.; Zhang, Y. MiR-378a-3p Enhances Adipogenesis by Targeting Mitogen-Activated Protein Kinase 1. Biochem. Biophys. Res. Commun. 2015, 457, 37–42. [Google Scholar] [CrossRef]
- Brandão, B.B.; Guerra, B.A.; Mori, M.A. Shortcuts to a Functional Adipose Tissue: The Role of Small Non-Coding RNAs. Redox Biol. 2017, 12, 82–102. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, Y.C.; Wang, J.; Kong, J.; Qi, Y.; Quigg, R.J.; Li, X. MiR-17-92 Cluster Accelerates Adipocyte Differentiation by Negatively Regulating Tumor-Suppressor Rb2/P130. Proc. Natl. Acad. Sci. 2008, 105, 2889–2894. [Google Scholar] [CrossRef] [Green Version]
- Jeong Kim, Y.; Jin Hwang, S.; Chan Bae, Y.; Sup Jung, J. MiR-21 Regulates Adipogenic Differentiation through the Modulation of TGF-β Signaling in Mesenchymal Stem Cells Derived from Human Adipose Tissue. Stem Cells 2009, 27, 3093–3102. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Li, H.; Song, Y.; Zhao, Y.; Zhai, L.; Wang, H.; Zhong, R.; Tang, H.; Zhu, D. MiR-378 Activates the Pyruvate-PEP Futile Cycle and Enhances Lipolysis to Ameliorate Obesity in Mice. EBioMedicine 2016, 5, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Ishida, M.; Shimabukuro, M.; Yagi, S.; Nishimoto, S.; Kozuka, C.; Fukuda, D.; Soeki, T.; Masuzaki, H.; Tsutsui, M.; Sata, M. MicroRNA-378 Regulates Adiponectin Expression in Adipose Tissue: A New Plausible Mechanism. PLoS ONE 2014, 9, e111537. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Spiegelman, B.M. Fat and Beyond: The Diverse Biology of PPARγ. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef] [PubMed]
- Kota, B.P.; Huang, T.H.W.; Roufogalis, B.D. An Overview on Biological Mechanisms of PPARs. Pharm. Res. 2005, 51, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Farmer, S.R. Transcriptional Control of Adipocyte Formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. Pparγ Signaling and Metabolism: The Good, the Bad and the Future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Jennewein, C.; Von Knethen, A.; Schmid, T.; Brüne, B. MicroRNA-27b Contributes to Lipopolysaccharide-Mediated Peroxisome Proliferator-Activated Receptor γ (PPARγ) MRNA Destabilization. J. Biol. Chem. 2010, 285, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Karbiener, M.; Fischer, C.; Nowitsch, S.; Opriessnig, P.; Papak, C.; Ailhaud, G.; Dani, C.; Amri, E.-Z.; Scheideler, M. MicroRNA MiR-27b Impairs Human Adipocyte Differentiation and Targets PPARγ. Biochem. Biophys. Res. Commun. 2009, 390, 247–251. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, A.Y.; Lee, H.W.; Son, Y.H.; Lee, G.Y.; Lee, J.W.; Lee, Y.S.; Kim, J.B. MiR-27a Is a Negative Regulator of Adipocyte Differentiation via Suppressing PPARγ Expression. Biochem. Biophys. Res. Commun. 2010, 392, 323–328. [Google Scholar] [CrossRef]
- Lin, Q.; Gao, Z.; Alarcon, R.M.; Ye, J.; Yun, Z. A Role of MiR-27 in the Regulation of Adipogenesis. Febs J. 2009, 276, 2348–2358. [Google Scholar] [CrossRef]
- Lee, E.K.; Lee, M.J.; Abdelmohsen, K.; Kim, W.; Kim, M.M.; Srikantan, S.; Martindale, J.L.; Hutchison, E.R.; Kim, H.H.; Marasa, B.S.; et al. MiR-130 Suppresses Adipogenesis by Inhibiting Peroxisome Proliferator-Activated Receptor Expression. Mol. Cell. Biol. 2011, 31, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.A.; Thomou, T.; Boucher, J.; Lee, K.Y.; Lallukka, S.; Kim, J.K.; Torriani, M.; Yki-Järvinen, H.; Grinspoon, S.K.; Cypess, A.M.; et al. Altered MiRNA Processing Disrupts Brown/White Adipocyte Determination and Associates with Lipodystrophy. J. Clin. Invest. 2014, 124, 3339–3351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, M.A.; Ludwig, R.G.; Garcia-Martin, R.; Brandão, B.B.; Kahn, C.R. Extracellular MiRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Raghavan, P.; Thomou, T.; Boucher, J.; Robida-Stubbs, S.; MacOtela, Y.; Russell, S.J.; Kirkland, J.L.; Blackwell, T.K.; Kahn, C.R. Role of MicroRNA Processing in Adipose Tissue in Stress Defense and Longevity. Cell Metab. 2012, 16, 336–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Xie, H.; Mori, M.A.; Alexander, R.; Yuan, B.; Hattangadi, S.M.; Liu, Q.; Kahn, C.R.; Lodish, H.F. Mir193b-365 Is Essential for Brown Fat Differentiation. Nat. Cell Biol 2011, 13, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, P.; Kajimura, S.; Yang, W.; Chin, S.; Rohas, L.M.; Uldry, M.; Tavernier, G.; Langin, D.; Spiegelman, B.M. Transcriptional Control of Brown Fat Determination by PRDM16. Cell Metab. 2007, 6, 38–54. [Google Scholar] [CrossRef] [Green Version]
- Trajkovski, M.; Ahmed, K.; Esau, C.C.; Stoffel, M. MyomiR-133 Regulates Brown Fat Differentiation through Prdm16. Nat. Cell Biol. 2012, 14, 1330–1335. [Google Scholar] [CrossRef]
- Feuermann, Y.; Kang, K.; Gavrilova, O.; Haetscher, N.; Jang, S.J.; Yoo, K.H.; Jiang, C.; Gonzalez, F.J.; Robinson, G.W.; Hennighausen, L. MiR-193b and MiR-365-1 Are Not Required for the Development and Function of Brown Fat in the Mouse. Rna Biol. 2013, 10, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Zaragosi, L.E.; Wdziekonski, B.; Brigand, K.L.; Villageois, P.; Mari, B.; Waldmann, R.; Dani, C.; Barbry, P. Small RNA Sequencing Reveals MiR-642a-3p as a Novel Adipocyte-Specific MicroRNA and MiR-30 as a Key Regulator of Human Adipogenesis. Genome Biol. 2011, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Wang, M.; Xiao, T.; Yin, B.; He, L.; Meng, W.; Dong, M.; Liu, F. MiR-30 Promotes Thermogenesis and the Development of Beige Fat by Targeting RIP140. Diabetes 2015, 64, 2056–2068. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhao, L.; Xing, L.; Chen, D. MicroRNA-204 Regulates Runx2 Protein Expression and Mesenchymal Progenitor Cell Differentiation. Stem Cells 2010, 28, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Kiskinis, E.; Hallberg, M.; Christian, M.; Olofsson, M.; Dilworth, S.M.; White, R.; Parker, M.G. RIP140 Directs Histone and DNA Methylation to Silence Ucp1 Expression in White Adipocytes. Embo J. 2007, 26, 4831–4840. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guan, M.; Townsend, K.L.; Huang, T.L.; An, D.; Yan, X.; Xue, R.; Schulz, T.J.; Winnay, J.; Mori, M.; et al. MicroRNA-455 Regulates Brown Adipogenesis via a Novel HIF1 an- AMPK - PGC 1α Signaling Network. Embo Rep. 2015, 16, 1378–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, Y.H.; Kokkotou, E.; Schulz, T.J.; Huang, T.L.; Winnay, J.N.; Taniguchi, C.M.; Tran, T.T.; Suzuki, R.; Espinoza, D.O.; Yamamoto, Y.; et al. New Role of Bone Morphogenetic Protein 7 in Brown Adipogenesis and Energy Expenditure. Nature 2008, 454, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Schulz, T.J.; Huang, P.; Huang, T.L.; Xue, R.; McDougall, L.E.; Townsend, K.L.; Cypess, A.M.; Mishina, Y.; Gussoni, E.; Tseng, Y.H. Brown-Fat Paucity Due to Impaired BMP Signalling Induces Compensatory Browning of White Fat. Nature 2013, 495, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Mao, C.; Quattrochi, B.; Friedline, R.H.; Zhu, L.J.; Jung, D.Y.; Kim, J.K.; Lewis, B.; Wang, Y.X. MicroRNA-378 Controls Classical Brown Fat Expansion to Counteract Obesity. Nat. Commun. 2014, 5, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Francis, S.H.; Blount, M.A.; Corbin, J.D. Mammalian Cyclic Nucleotide Phosphodiesterases: Molecular Mechanisms and Physiological Functions. Physiol. Rev. 2011, 91, 651–690. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Heneidi, S.; Lee, J.M.; Layman, L.C.; Stepp, D.W.; Gamboa, G.M.; Chen, B.S.; Chazenbalk, G.; Azziz, R. Mirna-93 Inhibits Glut4 and Is Overexpressed in Adipose Tissue of Polycystic Ovary Syndrome Patients and Women with Insulin Resistance. Diabetes 2013, 62, 2278–2286. [Google Scholar] [CrossRef] [Green Version]
- Foshay, K.M.; Gallicano, G.I. MiR-17 Family MiRNAs Are Expressed during Early Mammalian Development and Regulate Stem Cell Differentiation. Dev. Biol. 2009, 326, 431–443. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zuo, J.; Zhang, Y.; Xie, Y.; Hu, F.; Chen, L.; Liu, B.; Liu, F. Identification of MiR-106b-93 as a Negative Regulator of Brown Adipocyte Differentiation. Biochem. Biophys. Res. Commun. 2013, 438, 575–580. [Google Scholar] [CrossRef]
- He, M.; Xu, Z.; Ding, T.; Kuang, D.M.; Zheng, L. MicroRNA-155 Regulates Inflammatory Cytokine Production in Tumor-Associated Macrophages via Targeting C/EBPβ. Cell. Mol. Immunol. 2009, 6, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Worm, J.; Stenvang, J.; Petri, A.; Frederiksen, K.S.; Obad, S.; Elmén, J.; Hedtjärn, M.; Straarup, E.M.; Hansen, J.B.; Kauppinen, S. Silencing of MicroRNA-155 in Mice during Acute Inflammatory Response Leads to Derepression of c/Ebp Beta and down-Regulation of G-CSF. Nucleic Acids Res. 2009, 37, 5784–5792. [Google Scholar] [CrossRef]
- Skårn, M.; Namløs, H.M.; Noordhuis, P.; Wang, M.Y.; Meza-Zepeda, L.A.; Myklebost, O. Adipocyte Differentiation of Human Bone Marrow-Derived Stromal Cells Is Modulated by MicroRNA-155, MicroRNA-221, and MicroRNA-222. Stem Cells Dev. 2012, 21, 873–883. [Google Scholar] [CrossRef]
- Chen, Y.; Siegel, F.; Kipschull, S.; Haas, B.; Fröhlich, H.; Meister, G.; Pfeifer, A. MiR-155 Regulates Differentiation of Brown and Beige Adipocytes via a Bistable Circuit. Nat. Commun. 2013, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, T.; Seok, S.; Choi, S.; Huang, Z.; Suino-Powell, K.; Eric Xu, H.; Kemper, B.; Kemper, J.K. MicroRNA 34a Inhibits Beige and Brown Fat Formation in Obesity in Part by Suppressing Adipocyte Fibroblast Growth Factor 21 Signaling and SIRT1 Function. Mol. Cell. Biol. 2014, 34, 4130–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavery, C.A.; Kurowska-Stolarska, M.; Holmes, W.M.; Donnelly, I.; Caslake, M.; Collier, A.; Baker, A.H.; Miller, A.M. MiR-34a −/− Mice Are Susceptible to Diet-Induced Obesity. Obesity 2016, 24, 1741–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Articles | 898 |
|---|---|
| Citations | 25,556 |
| Average citations per articles | 28.45 |
| Authors | 4106 |
| Articles per author | 4.57 |
| Coauthor per article | 6.98 |
| Collaboration Index | 4.66 |
| Sources (journals) | 358 |
| Keywords | 7261 |
| Collaboration Index | 4.66 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manoel Alves, J.; Handerson Gomes Teles, R.; do Valle Gomes Gatto, C.; Muñoz, V.R.; Regina Cominetti, M.; Garcia de Oliveira Duarte, A.C. Mapping Research in the Obesity, Adipose Tissue, and MicroRNA Field: A Bibliometric Analysis. Cells 2019, 8, 1581. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121581
Manoel Alves J, Handerson Gomes Teles R, do Valle Gomes Gatto C, Muñoz VR, Regina Cominetti M, Garcia de Oliveira Duarte AC. Mapping Research in the Obesity, Adipose Tissue, and MicroRNA Field: A Bibliometric Analysis. Cells. 2019; 8(12):1581. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121581
Chicago/Turabian StyleManoel Alves, João, Ramon Handerson Gomes Teles, Camila do Valle Gomes Gatto, Vitor Rosetto Muñoz, Márcia Regina Cominetti, and Ana Cláudia Garcia de Oliveira Duarte. 2019. "Mapping Research in the Obesity, Adipose Tissue, and MicroRNA Field: A Bibliometric Analysis" Cells 8, no. 12: 1581. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121581