DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling


Laboratory of Aging Physiology and Molecular Evolution, Department of Biology, Ghent University, 9000 Ghent, Belgium
*
Author to whom correspondence should be addressed.
Cells 2020, 9(1), 109; https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010109
Submission received: 28 November 2019
/
Revised: 30 December 2019
/
Accepted: 31 December 2019
/
Published: 2 January 2020
(This article belongs to the Special Issue The FoxO Transcription Factors and Metabolic Regulation)
{kind=link}
Abstract
:DAF-16, the only forkhead box transcription factors class O (FoxO) homolog in Caenorhabditis elegans, integrates signals from upstream pathways to elicit transcriptional changes in many genes involved in aging, development, stress, metabolism, and immunity. The major regulator of DAF-16 activity is the insulin/insulin-like growth factor 1 (IGF-1) signaling (IIS) pathway, reduction of which leads to lifespan extension in worms, flies, mice, and humans. In C. elegans daf-2 mutants, reduced IIS leads to a heterochronic activation of a dauer survival program during adulthood. This program includes elevated antioxidant defense and a metabolic shift toward accumulation of carbohydrates (i.e., trehalose and glycogen) and triglycerides, and activation of the glyoxylate shunt, which could allow fat-to-carbohydrate conversion. The longevity of daf-2 mutants seems to be partially supported by endogenous trehalose, a nonreducing disaccharide that mammals cannot synthesize, which points toward considerable differences in downstream mechanisms by which IIS regulates aging in distinct groups.
1. Introduction
Over the past decades, the free-living nematode Caenorhabditis elegans has been developed into a prominent model organism in aging research. This is primarily due to its short lifespan, the ease of culturing and experimentation [1], the fully sequenced and well-annotated genome [2,3], and ample availability of forward and reverse genetic screens [4]. Identification of DAF-16, the C. elegans homolog of the forkhead box transcription factors class O (FoxO), as one of the key regulators of the aging process [5,6,7] prompted massive attention and a wealth of studies aimed at identifying its upstream regulators, downstream targets and the mechanisms by which it controls aging.
Originally, daf-16 was identified in studies on the genetic basis of dauer formation [8,9]. The dauer larva is the most specialized diapause stage of C. elegans and it is crucial for survival in the spatially and temporally restricted microbe-rich patches of rotting plant material, which this worm naturally inhabits [8,9,10,11]. Under adverse conditions, L1/early L2 larvae develop into a dauer, an alternative L3 stage. Dauer entry is triggered by a continuously secreted dauer (“crowding”) pheromone and this process is further modulated by temperature and food availability [12,13]. Dauer larvae are long-lived and considered non-aging (as the post-dauer lifespan is not affected by the time spent in dauer stage) [14] and highly resistant to various types of stress [15]. When the environment becomes hospitable again, dauers molt into L4 larvae and development is continued as usual.
2. Identification of DAF-16: The Genetics of Dauer Arrest
Early studies to identify genes regulating dauer arrest characterized two classes of daf (abnormal dauer formation) mutants: dauer constitutive (daf-c) and dauer defective (daf-d) [8,9]. The daf-16 mutations were classified as daf-d because these mutants failed to form dauers under dauer-inducing conditions [8]. daf-2 mutants, on the other hand, were daf-c since they formed dauers even under favorable conditions [8]. Furthermore, dauer entry in daf-2 mutants was partially or entirely suppressed by daf-16 mutations [16]. In the same decade, the first single-gene mutation causing lifespan extension in C. elegans was identified: age-1 (allelic with daf-23) [17,18,19,20]. In the early 1990s, mutation in daf-2 was found to double C. elegans lifespan and this effect required functional daf-16, thus linking the genetics of dauer formation and longevity [5]. In the years to come, molecular cloning and characterization showed that daf-2 encodes a receptor tyrosine kinase homolog [21], age-1 a phosphatidyl inositol 3-kinase (PI3K) homolog [20], and daf-16 a FoxO transcription factor [6,7]; all acting together in an insulin/insulin-like growth factor 1 (IGF-1) signaling (IIS) pathway.
3. DAF-16/FoxO Structure and Isoforms
The forkhead box (Fox) gene family encodes transcription factors characterized by a winged-helix DNA-binding domain (‘forkhead box’) and is evolutionarily conserved in species ranging from yeast to humans [22,23]. These transcription factors have regulatory roles in a plethora of cellular processes, such as differentiation, apoptosis, DNA repair, cell cycle arrest, metabolism, stress and immune response, and longevity [24,25,26]. In mammals, there are 19 subclasses of Fox transcription factors (A to S) based on sequence homology [23] and the FoxO subclass contains four members encoded by distinctive genes: FoxO1 (FKHR), FoxO3 (FKHRL1), FoxO4 (AFX), and FoxO6 [23,27]. In invertebrates, only one member of the FoxO subfamily has been characterized: DAF-16 in C. elegans [6,7] and dFOXO in Drosophila melanogaster [28].
In C. elegans, five different DAF-16 isoforms are transcribed from three distinctive promoters of a single genetic locus: daf-16a1/a2, daf-16b, and daf-16d/f [29,30,31,32]. These isoforms differ in their tissue distribution and function as indicated by multiple studies using transcriptional and translational reporters, promoter swapping, mutants, and RNA interference (RNAi) [29,30,31,33,34]. While daf-16a1/a2 and daf-16d/f are expressed in almost all tissues [29,30,31], daf-16b expression is primarily enriched in the pharynx, somatic gonad, and neurons [29,31,33], and this distribution is driven by divergence at the isoforms’ N-termini [31,34]. Furthermore, daf-16a1/a2 and daf-16d/f have dominant roles in controlling dauer arrest and lifespan [29,31,34], while daf-16b is required for pharynx remodeling during dauer formation [29,31] and neurite outgrowth during development [33].
To regulate the expression of its targets, DAF-16 binds to a consensus DNA motif TTGTTTAC, known as the DAF-16 binding element (DBE) [35]. Analysis of the 1 kb region upstream of the putative DAF-16 targets revealed that the DBE is overrepresented in promoter regions of both Class I (upregulated in daf-2 mutants) and Class II (downregulated in daf-2 mutants) genes, suggesting that DAF-16 also functions as a transcriptional repressor [36]. However, by using DamID, Schuster et al. [37] demonstrated that DAF-16 acts solely as a transcriptional activator of Class I genes through binding to DBE. A microarray study by Murphy et al. [36] identified a second overrepresented motif: the GATA-like sequence CTTATCA, later named DAF-16 associated element (DAE) [38]. Rather than DAF-16, DAE is bound by PQM-1, a transcriptional factor with a key role in controlling the expression of Class II genes and their downregulation in daf-2 mutants, thus accompanying DAF-16’s role in IIS-mediated longevity [39].
4. The IIS Pathway: A Central Regulator of DAF-16 Activity
The insulin/IGF-1 signaling pathway is an evolutionarily conserved phosphorylation cascade that controls growth, metabolism, and reproduction in response to nutrient availability in all eukaryotes. Moreover, this pathway has a well-established role in regulating aging as reduced IIS leads to lifespan extension in C. elegans [5,18], D. melanogaster [40,41], and Mus musculus [42,43].
The IIS cascade is activated by binding of insulin-like peptides (ILPs) to the membrane receptor tyrosine kinase. The C. elegans genome encodes 40 putative ILPs that are largely expressed in neurons and interneurons [44,45,46]. However, the differential roles of these peptides are not yet fully explored. These ILPs are not generally redundant, rather, they have distinct functions and regulate each other transcriptionally within a complex signaling network that affects C. elegans development, stress resistance, and longevity [47]. A recent study functionally characterized all 40 ILPs in vivo, by creating transgenic lines overexpressing individual peptides pan-neuronally and scoring multiple phenotypes [48]. The study unraveled functions of 35 ILPs and classified them as agonists, antagonists, or pleiotropic to IIS. Moreover, eight ILPs were shown to have specific functions (e.g., INS-15, INS-21, and INS-22 are IIS antagonists with roles in dauer formation, fat accumulation, and lifespan of L1 arrested larvae, respectively) [48]. D. melanogaster has eight ILPs (DILPs) encoded by distinct genes [49,50], while in mammals the ILP superfamily contains 10 members of which only insulin, IGF-1, and IGF-2 are related to C. elegans ILPs and bind to tyrosine kinase receptors [51,52,53,54,55]. In striking contrast to the number of ILPs, there is only one insulin receptor (IR) in C. elegans—DAF-2. In humans, on the other hand, there are several tyrosine kinase receptors (IR, IGF-IR, IGF-IIR, and a hybrid insulin receptor-related receptor—IRR) that bind insulin, IGF-1, and IGF-2 with different affinities to elicit diverse and complex physiological effects [56].
Upon insulin-like ligand binding, the C. elegans DAF-2 receptor auto-phosphorylates, which results in the recruitment and activation of the p110 catalytic subunit of PI3K/AGE-1 [20]. AGE-1 catalyzes conversion of phosphatidylinositol 4,5-bisphosphate (PIP2) into phosphatidylinositol 3,4,5-trisphosphate (PIP3) [57]. In humans, the p110 catalytic subunit is recruited to the membrane by PI3K adaptor subunits that bind to the phosphotyrosines of the insulin receptor substrate (IRS) scaffold proteins [58]. The C. elegans genome encodes the IRS and PI3K adapter unit homologs, ist-1 and aap-1, respectively [59]. However, aap-1 and ist-1 are deemed dispensable for AGE-1/p110 catalytic subunit activation by DAF-2, at least in a wild-type background [59]. Elevated levels of PIP3 result in activation of the 3-phosphoinositide-dependent kinase-1 homolog PDK-1 [60], which, in turn, phosphorylates and activates the Akt/PKB homologs AKT-1 and AKT-2 [61] and the serum- and glucocorticoid-inducible kinase SGK-1 [62]. In mammals, both AKT and SGK can inhibit FoxO3A by phosphorylation but differ in preference for regulatory sites [63,64]. In C. elegans, the regulatory role of SGK-1 is less clear. While SGK-1 phosphorylates DAF-16 in vitro [62], it does not influence DAF-16 subcellular localization in vivo, but probably exerts its regulatory function by affecting other proteins that interact with DAF-16 [65]. Phosphorylation of DAF-16 by AKT-1 and AKT-2 promotes its sequestration in cytoplasm through the association with 14-3-3 scaffold proteins PAR-5 and FTT-2 [66,67]. Under the conditions of reduced IIS due to stress or mutations in daf-2 or any of the pathway kinases, DAF-16 translocates into the nucleus where it modulates transcription of its targets [36,38,68,69].
5. IIS-Independent Regulation of DAF-16 Activity
To fulfill its diverse roles, DAF-16 integrates signals from multiple upstream pathways that act in parallel to IIS. c-Jun N-terminal kinase JNK-1, a member of the mitogen-activated protein kinase (MAPK) family, and CST-1, a Ste20-like kinase 1 (MST1) homolog, directly phosphorylate DAF-16, leading to its nuclear translocation and activation in response to heat and oxidative stress [70,71]. Furthermore, overexpression of both jnk-1 and cst-1 results in a daf-16-dependent lifespan extension [70,71]. Another kinase that activates DAF-16 via direct phosphorylation is the AMP-activated protein kinase (AMPK) [72]. AMPK was shown to phosphorylate DAF-16 in vitro at least at six different residues, but it does not increase its nuclear translocation [72,73]. In turn, DAF-16 is a direct transcriptional activator of aakg-4, which encodes the regulatory γ subunit of AMPK, thus suggesting the existence of a positive feedback loop that further activates DAF-16 and the expression of its target genes [73]. This notion is additionally supported by the upregulation of aakg-4 in daf-2 mutants and lifespan shortening of daf-2 due to aakg-4 RNAi or mutation [73]. The nutrient-sensing target of rapamycin (TOR) signaling pathway also converges on DAF-16. More specifically, genetic inhibition of TORC1 leads to increased transcription of daf-16 and the nuclear translocation of a single DAF-16 isoform, DAF-16d/f, resulting in lifespan extension [74]. In addition to DAF-16, this longevity phenotype also requires activation of SKN-1 [74]. Under the conditions of intermittent fasting, however, TORC1 promotes DAF-16 nuclear translocation and activation of its target genes in a LET-363/TOR- and RHEB-1-dependent manner [75]. Finally, removal of germline cells results in nuclear translocation and activation of DAF-16 in the intestine and this is facilitated by kri-1, which codes for an intestinal ankyrin-repeat protein [76], and the microRNA mir-71 [77].
6. Identification of DAF-16 Targets
In order to uncover underlying molecular mechanisms of C. elegans IIS mutant longevity, several approaches have been used with the aim to first identify DAF-16 transcriptional targets (reviewed in [78]). Bioinformatic approach by Lee et al. [79] identified 17 orthologs in C. elegans and D. melanogaster that contained the consensus DBE within 1 kb of their promoters. Subsequent functional analysis of these candidates in C. elegans by RNAi revealed that they regulate longevity, dauer formation, and fat storage [79]. Likewise, initial transcriptomic studies based on expression microarrays and serial analysis of gene expression (SAGE) identified DAF-16-dependent upregulation of genes involved in metabolism, cellular stress, and antimicrobial response [36,38,80,81]. A subset of these genes was functionally assayed for lifespan phenotypes by RNAi, and despite the fact that the vast majority had a significant effect, not a single treatment completely abolished the longevity of daf-2 mutants like daf-16 RNAi, pointing toward a complex network of multiple effector genes to regulate aging [36]. In subsequent studies, chromatin immunoprecipitation (chIP) [82,83,84] and DamID [37] were performed to distinguish between direct and indirect targets of DAF-16. Oh et al. identified 88 putative targets that had at least one DAF-16 binding site in their promoter region using chIP on non-synchronized cultures of daf-2 worms [82]. A DamID approach detected 65 ‘high-confidence’ targets, i.e., genes that are targets of DAF-16 and are regulated by IIS, which were also enriched for Class I genes identified in the previous microarray study [37]. Many of the differentially expressed DAF-16 targets identified in daf-2 mutants by genome-wide studies are shared with the transcriptome of dauer larvae [38,68,85]. These observations are supported by multiple proteomic [86,87,88,89,90] and metabolomic [91,92] studies, which also highlighted the extensive remodeling of metabolism in daf-2 mutants, reminiscent of hypometabolic dauers. This is not surprising given that dauers are also long-lived and supports the idea that daf-2 mutants could potentially rely on the heterochronically activated dauer survival program for their longevity assurance. In line with this is the discovery that the TGF-β signaling pathway regulates both longevity and dauer formation and this occurs partially through interactions with IIS and changes in DAF-16 cellular localization [93]. Further evidence that supports the notion that daf-2 are “adult dauers” is that, at 20 °C, adults of the Class 2 daf-2(e1370) mutant show mild dauer-like characteristics, such as body darkening, gonad shrinkage, and early-life decline in spontaneous movement [94,95,96]. On the contrary, Ewald et al. have demonstrated that the lifespan extension due to reduced IIS can be independent of the dauer program [95]. This is the case when daf-2(e1370) are grown at 15 °C or the daf-2 RNAi is performed at 15, 20, or 25 °C, all of which are the conditions where dauer-like traits are absent and require SKN-1 for lifespan extension [95]. Thus, the requirement of the heterochronically activated dauer program for daf-2 longevity appears to be context-dependent. In the following sections, we will discuss aspects of dauer physiology manifested in daf-2 mutants and their importance for daf-2 longevity.
7. DAF-16-Mediated Enhanced Stress Resistance in Long-Lived daf-2 Mutants
In C. elegans, reduced IIS confers extension of lifespan as well as elevated resistance to various types of stress, including heat [97], oxidative [98,99], UV [100], osmotic [101], hypoxic [102,103], and heavy metal [104] stress. Likewise, long-lived IIS mutants also show increased resistance to bacterial pathogens [105], reduced bacterial colonization, and enhanced clearance of pathogenic bacteria [106]. This wide range of stress responses is mediated by joint activity of DAF-16 with transcription factors such as HSF-1 [107,108], SKN-1 [109,110,111], SMK-1 [112] and, with regards to innate immunity, the p38 MAPK pathway [113]. Could increase in stress resistance be the key mechanism of IIS mutant longevity given the correlation between these two phenotypes? Concordant with this idea, numerous studies have demonstrated upregulation of superoxide dismutases (SODs), catalases, and glutathione S-transferases in long-lived IIS mutants [36,80,98,114,115]. However, both double deletion of sod-2 and sod-3 (mitochondrial) [116] and sod-1 and sod-5 (cytosolic) [117] render IIS mutants sensitive to oxidative stress but do not abolish their longevity. Deletion of sod-4 (extracellular) even further extends daf-2 lifespan [117]. Moreover, a quintuple mutant with a complete loss of SOD activity has a normal lifespan despite being increasingly sensitive not only to oxidative but also osmotic, heat, and cold stress [118]. Finally, a recent study has shown a similar effect in daf-2 worms: complete lack of SOD activity fully suppresses resistance to oxidative and heat stress but has a negligible effect on longevity [119]. Altogether these results imply that stress resistance and longevity can be experimentally uncoupled and that elevated activity of antioxidant enzymes is dispensable for daf-2 longevity. If not supporting longevity, what could possibly be the role of increased antioxidant defense? Taking into consideration C. elegans ecology, high antioxidant activity in dauers is crucial for the survival of anhydrobiosis and rehydration, which are accompanied by high levels of reactive oxygen species [120]. In daf-2 mutants that do not naturally undergo these cycles of anhydrobiotic survival, elevated antioxidant levels could potentially be only an integral part of the heterochronically activated dauer program that is unrelated to lifespan extension.
8. The Metabolism in daf-2 Mutants is Extensively Remodeled
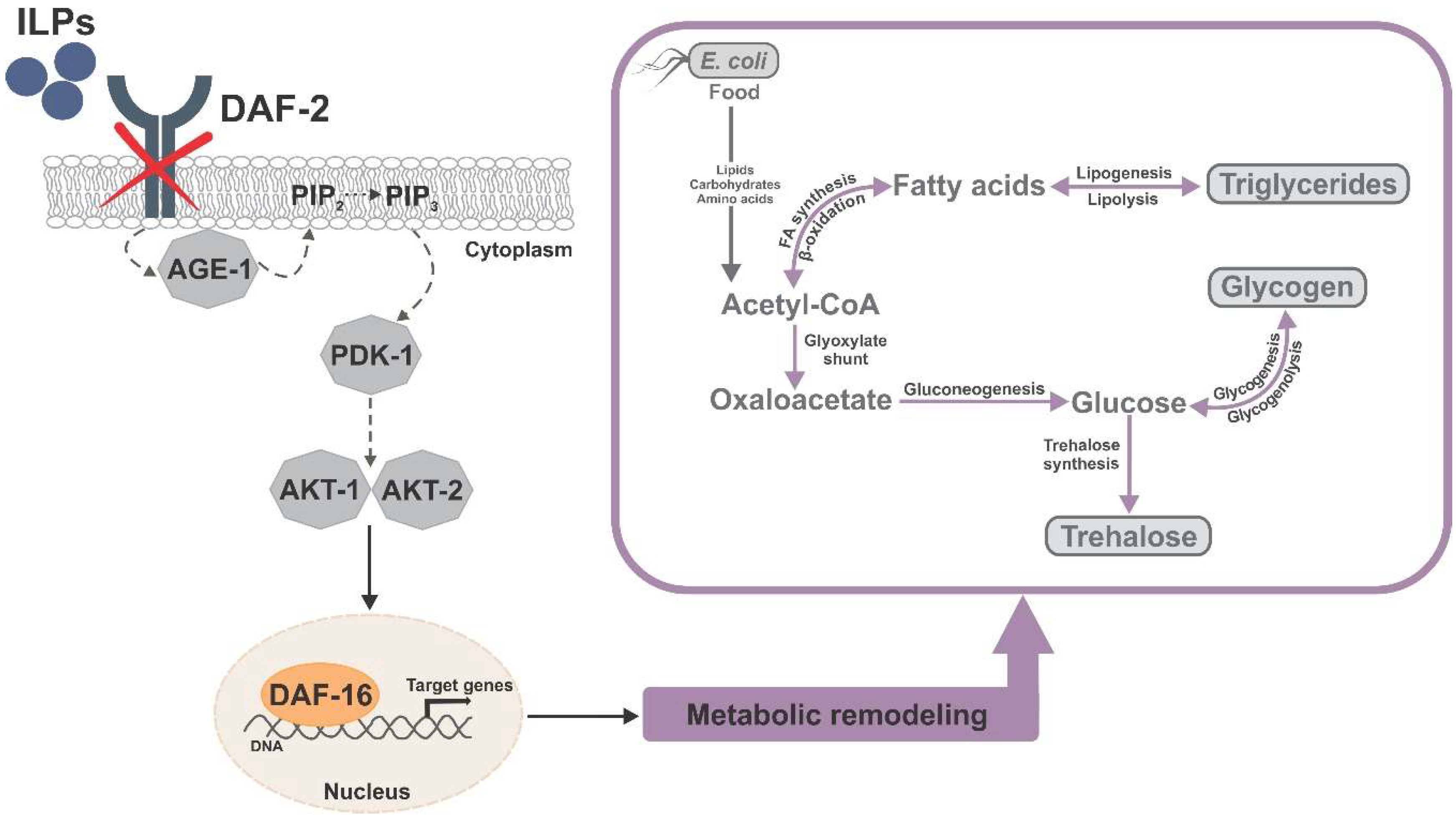
In addition to elevated stress resistance, another aspect of dauer physiology is reiterated in daf-2 mutants: a massive restructuring of intermediary metabolism [86,89,91] (Figure 1). In dauers, nonessential and energy-costly metabolic functions, such as feeding, growth, and reproduction, are suppressed [121], and all the internal energy stores are used for long-term structural maintenance and stress resistance [122]. Prior to dauer formation, young larvae accumulate triglycerides as intestinal and hypodermal lipid droplets [123], an energy reserve that can be consumed in a slow and strictly controlled manner to ensure survival of extended periods without food [123,124]. Additionally, dauers accumulate glycogen, which could serve as a readily available short-term energy source for locomotion and nictation, especially in the first weeks after dauer formation when they are highly motile [122].
Similarly, daf-2 mutants have large lipid [7,21,89,125] and glycogen stores [89,126] in their intestine and hypodermis, as shown by different histological, biochemical, and ultrastructural studies. Transmission electron microscopy (TEM) images also indicate higher amounts of glycogen in daf-2 body-wall muscles [89]. Furthermore, daf-2 mutants show increase in de novo fatty acid synthesis, as demonstrated by a 13C isotope-labeling approach, and this effect is entirely dependent on DAF-16 activity [127]. This finding appears to be in contrast with upregulation of many genes involved in β-oxidation in daf-2 worms at protein level [89]. Namely, fatty acid β-oxidation is inhibited by high concentrations of malonyl-CoA coming from increased fatty acid synthesis. These discrepancies could be explained by the different stages of worms used in the isotope-labeling and proteomic studies, i.e., late L4 larval stage and day 2 adults, respectively [89,127]. Indeed, age-dependent changes in lipid content in daf-2 mutants point toward a metabolic switch from lipogenesis during development and early adulthood (up to day 2) to tightly controlled lipolysis throughout the remaining life [89], reminiscent of non-feeding dauer larvae [123,124]. Concordant with this notion is also the reduced level of the fatty acid synthase homolog FASN-1 in adult daf-2 worms [89]. However, it is crucial to highlight that the described increase in lipid synthesis and storage refers to mutants in a canonical allele daf-2(e1370) [7,89,125,127] and that certain alternative lifespan-extending alleles such as e1368 and m577 show no such phenotype [127], suggesting that the bulk increase in lipid content per se is not directly correlated with longevity in IIS daf-2 mutants. On the other hand, altering lipid composition toward increase in unsaturated fatty acids could potentially play a role. Consistent with this, IIS longevity mutants show DAF-16-dependent upregulation of fatty acid desaturase FAT-7 [36] and elevated levels of monounsaturated fatty acids (MUFAs) [92,128,129] as well as MUFA-rich triglycerides [129].
Glycogen is essential for survival of acute hyperosmotic stress in C. elegans as a readily available reservoir for the production of glycerol in a pathway mediated by AMPK [130]. Its protective role was also demonstrated in daf-2 mutants in conditions of osmotic stress and anoxia [126,131]. Lifespan effects of glycogen in worms fed on standard and glucose-rich diet were evaluated by knocking down genes involved in glycogen synthesis and breakdown, i.e., glycogen synthase (gsy-1) and glycogen phosphorylase (pyg-1) [132]. gsy-1 knockdown depletes glycogen reserves, however, it causes only a negligible effect on daf-2 longevity on both diets [132]. pyg-1 RNAi results in a further increase in glycogen content in daf-2 worms (relative to controls without a knockdown), however, this treatment drastically shortens lifespan only on a high glucose diet, suggesting that accumulating excess glucose in form of glycogen in these conditions accelerates aging [132]. These observations were recently confirmed in a study that has also demonstrated that reduction of gsy-1 leads to a metabolic shift toward an increase in stored trehalose, which is crucial for alleviating detrimental effects of high dietary glucose in both wild type and daf-2 worms [133]. Moreover, this beneficent effect of trehalose requires the functional DAF-16d/f isoform [133]. The role of glycogen in daf-2 physiology is not entirely clear. It appears that these worms live even slightly longer when glycogen is depleted by gsy-1 knockdown [133] (in contrast with [132]), yet, GSY-1 is one of the most upregulated proteins in daf-2 mutants, in agreement with the large glycogen accumulations observed in these worms [89]. It is plausible that accumulation of this sugar is a result of the activated dauer program with the sole purpose of protection against stress.
Another sugar that could possibly be the main player in daf-2 longevity is the nonreducing disaccharide trehalose. It is a major circulating sugar in insects, where it is used as an energy source for fueling glycolysis [134] and its protective role against desiccation [135,136], heat [137], and osmotic [101,138] stress is shown in diverse organisms. In C. elegans and the anhydrobiotic nematode Aphelenchus avenae, there are two trehalose phosphate synthase genes, tps-1 and tps-2, responsible for synthesis of trehalose [139,140]. These genes are the downstream targets of daf-2/daf-16 signaling and are highly upregulated in dauers and daf-2 mutants [36,80,89], in accordance with increased trehalose levels in these worms [91,101,141]. In dauer larvae, trehalose is essential for survival of extreme desiccation [135]. Furthermore, this effect requires a fully active glyoxylate cycle that allows utilization of fat as a major carbon source for trehalose synthesis [142]. C. elegans has a single protein with two enzymatic functions (isocitrate lyase and malate synthase) that is responsible for the glyoxylate shunt reactions [143] and exists in two isoforms: ICL-1 (GEI-7) and the poorly characterized C08F11.14 [126,142]. ICL-1 is expressed in mitochondria and it catalyzes conversion of isocitrate and acetyl-CoA into glyoxylate and succinate and subsequent conversion of glyoxylate into malate. While succinate can further fuel the tricarboxylic acid (TCA) cycle, malate can be converted into oxaloacetate and used for the gluconeogenesis to generate trehalose [142,144]. Certainly, impairment of ICL-1 function results in a severe impediment of dauer desiccation tolerance due to a reduced amount of trehalose in these worms [142]. Likewise, RNAi knockdown of icl-1 shortens lifespan in the daf-2 mutants mu150 and e1370, by up to ~23% and ~27%, respectively, probably by a similar mechanism [145]. These observations are supported by heavy upregulation of icl-1 in dauers [68,142,146] and daf-2 mutants [36,89,145] and a recent metabolic model that predicts greater production of trehalose from fatty acids by activated glyoxylate cycle in microaerobic conditions [147]. Finally, simultaneous RNAi knockdown of tps-1 and tps-2 shortens lifespan in daf-2 mutants, also indicating that the endogenous trehalose is at least partially responsible for daf-2 longevity [141].
The protective role of trehalose is ascribed to the preservation of membrane organization through maintaining native packaging of lipids, prevention of protein denaturation, and aggregation and clearance of formed aggregates through induction of autophagy [89,120,133,135,148,149]. Moreover, trehalose was shown to jointly act with a group of intrinsically disordered proteins (late embryogenesis abundant, LEA) to exert its protective function both in vitro and in vivo [148]. There is only one LEA homolog in the C. elegans genome, encoded by lea-1, and it is essential for dauer desiccation tolerance [120]. Not surprisingly, lea-1 is shown to be upregulated in daf-2 worms, yet again pointing toward the heterochronic activation of the dauer survival program in these mutants [86,89]. Considering the importance of trehalose for longevity and its role in alleviating detrimental effects of glucose-rich diet in C. elegans [133,141,145], it would be interesting to explore the tissue dependency of these effects and the mechanisms of trehalose transport between different tissues.
The described restructuring of intermediary metabolism in long-lived daf-2 mutants is an evolutionarily conserved strategy of small organisms with restricted mobility, such as fungi [150], some plants [151], and invertebrates [135,152], to survive desiccation. This tolerance relies heavily on mechanisms that are absent in vertebrates (e.g., trehalose synthesis, glyoxylate shunt activation, LEA proteins) and that, at least partially, contribute to daf-2 longevity. Even though the FoxO transcription factors have been linked to human longevity, the underlying mechanisms by which this is achieved could differ substantially between worms and humans due to differences in the details of their physiology, organismal complexity, and environmental challenges these organisms face in nature [153,154].
9. Conclusions
The C. elegans FoxO homolog DAF-16 is the main output of IIS with a central role in regulating aging, which is an evolutionarily conserved function. However, DAF-16-mediated changes in daf-2 mutants are reminiscent of the hypometabolic dauer larvae and the longevity of these worms seems to rely, at least in part, on mechanisms that are not present in humans, such as synthesis of trehalose and activation of the glyoxylate cycle. Furthermore, it appears that certain aspects of daf-2 physiology are a consequence of a heterochronically activated dauer program with a role in stress resistance, but not longevity. On the other hand, the discovery of DAF-16 in C. elegans and its conserved role in aging across metazoans stemmed from studies on the genetics of dauer arrest. In addition to dauer formation, IIS also regulates many other aspects of C. elegans physiology (e.g., metabolism, autophagy, proteostasis), which have been linked to aging and diseases. Moreover, IIS-mediated longevity in daf-2 mutants can also be achieved independently of a genetic dauer program, in a SKN-1-dependent manner [95]. Finally, certain single nucleotide polymorphisms in the DAF-16 ortholog, FoxO3A, have been associated with human longevity [155]. Taken together, these findings suggest that uncovering more details of IIS-mediated longevity in C. elegans could provide potentially valuable targets for treatment of aging and age-related diseases in humans.
Author Contributions
Writing—original draft preparation, A.Z.; writing—review and editing, B.P.B. All authors have read and agreed to the published version of the manuscript.
Funding
A.Z. is funded by the Special Research Fund (BOF) of Ghent University, project BOF2-4J.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stiernagle, T. Maintenance of C. elegans. Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 31 December 2019).
- C. elegans Sequencing Consortium. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar] [CrossRef]
- Kamath, R.S.; Fraser, A.G.; Dong, Y.; Poulin, G.; Durbin, R.; Gotta, M.; Kanapin, A.; Le Bot, N.; Moreno, S.; Sohrmann, M.; et al. Systematic functional analysis of the Caenorhabditis elegans genome using rnai. Nature 2003, 421, 231–237. [Google Scholar] [CrossRef]
- Kutscher, L.M.; Shaham, S. Forward and Reverse Mutagenesis in C. elegans. Available online: http://www.wormbook.org/chapters/www_frmutagenesis/frmutagenesis.html (accessed on 31 December 2019).
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Dorman, J.B.; Rodan, A.; Kenyon, C. Daf-16: An hnf-3/forkhead family member that can function to double the life-span of Caenorhabditis elegans. Science 1997, 278, 1319–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogg, S.; Paradis, S.; Gottlieb, S.; Patterson, G.I.; Lee, L.; Tissenbaum, H.A.; Ruvkun, G. The fork head transcription factor daf-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 1997, 389, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.S.; Brown, S.J.; Riddle, D.L. Sensory control of dauer larva formation in Caenorhabditis elegans. J. Comp. Neurol. 1981, 198, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Riddle, D.L.; Swanson, M.M.; Albert, P.S. Interacting genes in nematode dauer larva formation. Nature 1981, 290, 668–671. [Google Scholar] [CrossRef]
- Braendle, C.; Milloz, J.; Félix, M.A. Mechanisms and evolution of environmental responses in Caenorhabditis elegans. In Current Topics in Developmental Biology; Academic Press: Cambridge, MA, USA, 2007; Volume 80, pp. 171–207. [Google Scholar]
- Félix, M.-A.; Braendle, C. The natural history of Caenorhabditis elegans. Curr. Biol. 2010, 20, R965–R969. [Google Scholar] [CrossRef]
- Golden, J.W.; Riddle, D.L. The Caenorhabditis elegans dauer larva: Developmental effects of pheromone, food, and temperature. Dev. Biol. 1984, 102, 368–378. [Google Scholar] [CrossRef]
- Hu, P.J. Dauer. Available online: http://www.wormbook.org/chapters/www_dauer/dauer.html (accessed on 31 December 2019).
- Klass, M.; Hirsh, D. Non-ageing developmental variant of Caenorhabditis elegans. Nature 1976, 260, 523–525. [Google Scholar] [CrossRef]
- Anderson, G.L. Responses of dauerlarvae of Caenorhabditis elegans (nematoda: Rhabditidae) to thermal stress and oxygen deprivation. Can. J. Zool. 1978, 56, 1786–1791. [Google Scholar] [CrossRef]
- Vowels, J.J.; Thomas, J.H. Genetic analysis of chemosensory control of dauer formation in Caenorhabditis elegans. Genetics 1992, 130, 105–123. [Google Scholar] [PubMed]
- Klass, M.R. A method for the isolation of longevity mutants in the nematode Caenorhabditis elegans and initial results. Mech. Ageing Dev. 1983, 22, 279–286. [Google Scholar] [CrossRef]
- Friedman, D.B.; Johnson, T.E. A mutation in the age-1 gene in Caenorhabditis elegans lengthens life and reduces hermaphrodite fertility. Genetics 1988, 118, 75–86. [Google Scholar] [PubMed]
- Malone, E.A.; Inoue, T.; Thomas, J.H. Genetic analysis of the roles of daf-28 and age-1 in regulating Caenorhabditis elegans dauer formation. Genetics 1996, 143, 1193–1205. [Google Scholar] [PubMed]
- Morris, J.Z.; Tissenbaum, H.A.; Ruvkun, G. A phosphatidylinositol-3-oh kinase family member regulating longevity and diapause in Caenorhabditis elegans. Nature 1996, 382, 536–539. [Google Scholar] [CrossRef]
- Kimura, K.D.; Tissenbaum, H.A.; Liu, Y.; Ruvkun, G. Daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 1997, 277, 942–946. [Google Scholar] [CrossRef]
- Weigel, D.; Jürgens, G.; Küttner, F.; Seifert, E.; Jäckle, H. The homeotic gene fork head encodes a nuclear protein and is expressed in the terminal regions of the Drosophila embryo. Cell 1989, 57, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Hannenhalli, S.; Kaestner, K.H. The evolution of fox genes and their role in development and disease. Nat. Rev. Genet. 2009, 10, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Accili, D.; Arden, K.C. Foxos at the crossroads of cellular metabolism, differentiation, and transformation. Cell 2004, 117, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Greer, E.L.; Brunet, A. Foxo transcription factors at the interface between longevity and tumor suppression. Oncogene 2005, 24, 7410–7425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Tindall, D.J. Dynamic foxo transcription factors. J. Cell Sci. 2007, 120, 2479–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaestner, K.H.; Knochel, W.; Martinez, D.E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 2000, 14, 142–146. [Google Scholar]
- Kramer, J.M.; Davidge, J.T.; Lockyer, J.M.; Staveley, B.E. Expression of Drosophila foxo regulates growth and can phenocopy starvation. BMC Dev. Biol. 2003, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.Y.N.; Hench, J.; Ruvkun, G. Regulation of C. elegans daf-16 and its human ortholog fkhrl1 by the daf-2 insulin-like signaling pathway. Curr. Biol. 2001, 11, 1950–1957. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.; Hsin, H.; Libina, N.; Kenyon, C. Regulation of the Caenorhabditis elegans longevity protein daf-16 by insulin/igf-1 and germline signaling. Nat. Genet. 2001, 28, 139–145. [Google Scholar] [CrossRef]
- Kwon, E.-S.; Narasimhan, S.D.; Yen, K.; Tissenbaum, H.A. A new daf-16 isoform regulates longevity. Nature 2010, 466, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.; Kwon, E.-S.; Conte, D.; Liu, H.; Gilchrist, M.J.; MacNeil, L.T.; Tissenbaum, H.A. Transcriptional regulation of Caenorhabditis elegans foxo/daf-16 modulates lifespan. Longev. Healthspan 2014, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Christensen, R.; De La Torre-Ubieta, L.; Bonni, A.; Colón-Ramos, D.A. A conserved pten/foxo pathway regulates neuronal morphology during C. elegans development. Development 2011, 138, 5257–5367. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.T.-Y.; Guo, C.; Itani, O.A.; Budaitis, B.G.; Williams, T.W.; Hopkins, C.E.; McEachin, R.C.; Pande, M.; Grant, A.R.; Yoshina, S.; et al. Longevity genes revealed by integrative analysis of isoform-specific daf-16/foxo mutants of Caenorhabditis elegans. Genetics 2015, 201, 613–629. [Google Scholar] [CrossRef] [Green Version]
- Furuyama, T.; Nakazawa, T.; Nakano, I.; Mori, N. Identification of the differential distribution patterns of mrnas and consensus binding sequences for mouse daf-16 homologues. Biochem. J. 2000, 349, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of daf-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Schuster, E.; McElwee, J.J.; Tullet, J.M.A.; Doonan, R.; Matthijssens, F.; Reece-Hoyes, J.S.; Hope, I.A.; Vanfleteren, J.R.; Thornton, J.M.; Gems, D. Damid in C. elegans reveals longevity-associated targets of daf-16/foxo. Mol. Syst. Biol. 2010, 6, 399. [Google Scholar] [CrossRef]
- McElwee, J.J.; Schuster, E.; Blanc, E.; Thomas, J.H.; Gems, D. Shared transcriptional signature in Caenorhabditis elegans dauer larvae and long-lived daf-2 mutants implicates detoxification system in longevity assurance. J. Biol. Chem. 2004, 279, 44533–44543. [Google Scholar] [CrossRef] [Green Version]
- Tepper, R.G.; Ashraf, J.; Kaletsky, R.; Kleemann, G.; Murphy, C.T.; Bussemaker, H.J. Pqm-1 complements daf-16 as a key transcriptional regulator of daf-2-mediated development and longevity. Cell 2013, 154, 676–690. [Google Scholar] [CrossRef] [Green Version]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of life-span by loss of chico, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef] [Green Version]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Holzenberger, M.; Dupont, J.; Ducos, B.; Leneuve, P.; Géloën, A.; Even, P.C.; Cervera, P.; Le Bouc, Y. Igf-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature 2003, 421, 182–187. [Google Scholar] [CrossRef]
- Blüher, M.; Kahn, B.B.; Kahn, C.R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef] [Green Version]
- Pierce, S.B.; Costa, M.; Wisotzkey, R.; Devadhar, S.; Homburger, S.A.; Buchman, A.R.; Ferguson, K.C.; Heller, J.; Platt, D.M.; Pasquinelli, A.A.; et al. Regulation of daf-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes Dev. 2001, 15, 672–686. [Google Scholar] [CrossRef] [Green Version]
- Cornils, A.; Gloeck, M.; Chen, Z.; Zhang, Y.; Alcedo, J. Specific insulin-like peptides encode sensory information to regulate distinct developmental processes. Development 2011, 138, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Kodama, E.; Kuhara, A.; Mohri-Shiomi, A.; Kimura, K.D.; Okumura, M.; Tomioka, M.; Iino, Y.; Mori, I. Insulin-like signaling and the neural circuit for integrative behavior in C. elegans. Genes Dev. 2006, 20, 2955–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes De Abreu, D.A.; Caballero, A.; Fardel, P.; Stroustrup, N.; Chen, Z.; Lee, K.; Keyes, W.D.; Nash, Z.M.; López-Moyado, I.F.; Vaggi, F.; et al. An insulin-to-insulin regulatory network orchestrates phenotypic specificity in development and physiology. PLoS Genet. 2014, 10, e1004225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Chiu, H.; Boudreau, J.; Papanicolaou, T.; Bendena, W.; Chin-Sang, I. A functional study of all 40 Caenorhabditis elegans insulin-like peptides. J. Biol. Chem. 2018, 293, 16912–16922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogiolo, W.; Stocker, H.; Ikeya, T.; Rintelen, F.; Fernandez, R.; Hafen, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol. 2001, 11, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Garelli, A.; Gontijo, A.M.; Miguela, V.; Caparros, E.; Dominguez, M. Imaginal discs secrete insulin-like peptide 8 to mediate plasticity of growth and maturation. Science 2012, 336, 579–582. [Google Scholar] [CrossRef]
- Bedarkar, S.; Turnell, W.G.; Blundell, T.L.; Schwabe, C. Relaxin has conformational homology with insulin. Nature 1977, 270, 449–451. [Google Scholar] [CrossRef]
- Rinderknecht, E.; Humbel, R.E. The amino acid sequence of human insulin-like growth factor i and its structural homology with proinsulin. J. Biol. Chem. 1978, 253, 2769–2776. [Google Scholar]
- Bell, G.I.; Pictet, R.L.; Rutter, W.J.; Cordell, B.; Tischer, E.; Goodman, H.M. Sequence of the human insulin gene. Nature 1980, 284, 26–32. [Google Scholar] [CrossRef]
- Büllesbach, E.E.; Schwabe, C. A novel leydig cell cdna-derived protein is a relaxin-like factor. J. Biol. Chem. 1995, 270, 16011–16015. [Google Scholar] [CrossRef] [Green Version]
- Chassin, D.; Laurent, A.; Janneau, J.-L.; Berger, R.; Bellet, D. Cloning of a new member of the insulin gene superfamily (insl4) expressed in human placenta. Genomics 1995, 29, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Clayton, P.E.; Banerjee, I.; Murray, P.G.; Renehan, A.G. Growth hormone, the insulin-like growth factor axis, insulin and cancer risk. Nat. Rev. Endocrinol. 2011, 7, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Weinkove, D.; Halstead, J.R.; Gems, D.; Divecha, N. Long-term starvation and ageing induce age-1/pi 3-kinase-dependent translocation of daf-16/foxo to the cytoplasm. BMC Biol. 2006, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical nodes in signalling pathways: Insights into insulin action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Wolkow, C.A.; Muñoz, M.J.; Riddle, D.L.; Ruvkun, G. Insulin receptor substrate and p55 orthologous adaptor proteins function in the Caenorhabditis elegans daf-2/insulin-like signaling pathway. J. Biol. Chem. 2002, 277, 49591–49597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, S.; Ailion, M.; Toker, A.; Thomas, J.H.; Ruvkun, G. A pdk1 homolog is necessary and sufficient to transduce age-1 pi3 kinase signals that regulate diapause in Caenorhabditis elegans. Genes Dev. 1999, 13, 1438–1452. [Google Scholar] [CrossRef]
- Paradis, S.; Ruvkun, G. Caenorhabditis elegans akt/pkb transduces insulin receptor-like signals from age-1 pi3 kinase to the daf-16 transcription factor. Genes Dev. 1998, 12, 2488–2498. [Google Scholar] [CrossRef] [Green Version]
- Hertweck, M.; Göbel, C.; Baumeister, R.C. Elegans sgk-1 is the critical component in the akt/pkb kinase complex to control stress response and life span. Dev. Cell 2004, 6, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Brunet, A.; Park, J.; Tran, H.; Hu, L.S.; Hemmings, B.A.; Greenberg, M.E. Protein kinase sgk mediates survival signals by phosphorylating the forkhead transcription factor fkhrl1 (foxo3a). Mol. Cell Biol. 2001, 21, 952–965. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.T.-Y.; Guo, C.; Dumas, K.J.; Ashrafi, K.; Hu, P.J. Effects of Caenorhabditis elegans sgk-1 mutations on lifespan, stress resistance, and daf-16/foxo regulation. Aging Cell 2013, 12, 932–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdichevsky, A.; Viswanathan, M.; Horvitz, H.R.; Guarente, L.C. Elegans sir-2.1 interacts with 14-3-3 proteins to activate daf-16 and extend life span. Cell 2006, 125, 1165–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Tewari, M.; Vidal, M.; Sylvia Lee, S. The 14-3-3 protein ftt-2 regulates daf-16 in Caenorhabditis elegans. Dev. Biol. 2007, 301, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kim, S.K. Global analysis of dauer gene expression in Caenorhabditis elegans. Development 2003, 130, 1621–1634. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.A.; Kawli, T.; Tan, M.-W. Pseudomonas aeruginosa suppresses host immunity by activating the daf-2 insulin-like signaling pathway in Caenorhabditis elegans. PLoS Pathog. 2008, 4, e1000175. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. Jnk regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/daf-16. Proc. Natl. Acad. Sci. USA 2005, 102, 4494–4499. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, M.K.; Yuan, Z.; Boag, P.R.; Yang, Y.; Villén, J.; Becker, E.B.E.; DiBacco, S.; De La Iglesia, N.; Gygi, S.; Blackwell, T.K.; et al. A conserved mst-foxo signaling pathway mediates oxidative-stress responses and extends life span. Cell 2006, 125, 987–1001. [Google Scholar] [CrossRef] [Green Version]
- Greer, E.L.; Dowlatshahi, D.; Banko, M.R.; Villen, J.; Hoang, K.; Blanchard, D.; Gygi, S.P.; Brunet, A. An ampk-foxo pathway mediates longevity induced by a novel method of dietary restriction in C. elegans. Curr. Biol. 2007, 17, 1646–1656. [Google Scholar] [CrossRef] [Green Version]
- Tullet, J.M.A.; Araiz, C.; Sanders, M.J.; Au, C.; Benedetto, A.; Papatheodorou, I.; Clark, E.; Schmeisser, K.; Jones, D.; Schuster, E.F.; et al. Daf-16/foxo directly regulates an atypical amp-activated protein kinase gamma isoform to mediate the effects of insulin/igf-1 signaling on aging in Caenorhabditis elegans. PLoS Genet. 2014, 10, e1004109. [Google Scholar] [CrossRef]
- Robida-Stubbs, S.; Glover-Cutter, K.; Lamming, D.W.; Mizunuma, M.; Narasimhan, S.D.; Neumann-Haefelin, E.; Sabatini, D.M.; Blackwell, T.K. Tor signaling and rapamycin influence longevity by regulating skn-1/nrf and daf-16/foxo. Cell Metab. 2012, 15, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Honjoh, S.; Yamamoto, T.; Uno, M.; Nishida, E. Signalling through rheb-1 mediates intermittent fasting-induced longevity in C. elegans. Nature 2009, 457, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.R.; Kenyon, C. Germ-cell loss extends C. elegans life span through regulation of daf-16 by kri-1 and lipophilic-hormone signaling. Cell 2006, 124, 1055–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulias, K.; Horvitz, H.R. The C. elegans microrna mir-71 acts in neurons to promote germline-mediated longevity through regulation of daf-16/foxo. Cell Metab. 2012, 15, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, C.T. The search for daf-16/foxo transcriptional targets: Approaches and discoveries. Exp. Gerontol. 2006, 41, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Kennedy, S.; Tolonen, A.C.; Ruvkun, G. Daf-16 target genes that control C. elegans life-span and metabolism. Science 2003, 300, 644–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McElwee, J.; Bubb, K.; Thomas, J.H. Transcriptional outputs of the Caenorhabditis elegans forkhead protein daf-16. Aging Cell 2003, 2, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Halaschek-Wiener, J.; Khattra, J.S.; McKay, S.; Pouzyrev, A.; Stott, J.M.; Yang, G.S.; Holt, R.A.; Jones, S.J.M.; Marra, M.A.; Brooks-Wilson, A.R.; et al. Analysis of long-lived C. elegans daf-2 mutants using serial analysis of gene expression. Genome Res. 2005, 15, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.W.; Mukhopadhyay, A.; Dixit, B.L.; Raha, T.; Green, M.R.; Tissenbaum, H.A. Identification of direct daf-16 targets controlling longevity, metabolism and diapause by chromatin immunoprecipitation. Nat. Genet. 2006, 38, 251–257. [Google Scholar]
- Kumar, N.; Jain, V.; Singh, A.; Jagtap, U.; Verma, S.; Mukhopadhyay, A. Genome-wide endogenous daf-16/foxo recruitment dynamics during lowered insulin signalling in C. elegans. Oncotarget 2015, 6, 41418–41433. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.E.; Kundaje, A.; Brunet, A. Characterization of the direct targets of foxo transcription factors throughout evolution. Aging Cell 2016, 15, 673–685. [Google Scholar] [CrossRef]
- McElwee, J.J.; Schuster, E.; Blanc, E.; Thornton, J.; Gems, D. Diapause-associated metabolic traits reiterated in long-lived daf-2 mutants in the nematode Caenorhabditis elegans. Mech. Ageing Dev. 2006, 127, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.-Q.; Venable, J.D.; Au, N.; Xu, T.; Park, S.K.; Cociorva, D.; Johnson, J.R.; Dillin, A.; Yates, J.R. Quantitative mass spectrometry identifies insulin signaling targets in C. elegans. Science 2007, 317, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.M.; Staffa, K.; Perally, S.; LaCourse, E.J.; Brophy, P.M.; Hamilton, J.V. Proteomic analyses of Caenorhabditis elegans dauer larvae and long-lived daf-2 mutants implicates a shared detoxification system in longevity assurance. J. Proteome Res. 2010, 9, 2871–2881. [Google Scholar] [CrossRef] [PubMed]
- Stout, G.J.; Stigter, E.C.A.; Essers, P.B.; Mulder, K.W.; Kolkman, A.; Snijders, D.S.; Van Den Broek, N.J.F.; Betist, M.C.; Korswagen, H.C.; MacInnes, A.W.; et al. Insulin/igf-1-mediated longevity is marked by reduced protein metabolism. Mol. Syst. Biol. 2013, 9, 679. [Google Scholar] [CrossRef]
- Depuydt, G.; Xie, F.; Petyuk, V.A.; Smolders, A.; Brewer, H.M.; Camp, D.G., 2nd; Smith, R.D.; Braeckman, B.P. Lc-ms proteomics analysis of the insulin/igf-1-deficient Caenorhabditis elegans daf-2(e1370) mutant reveals extensive restructuring of intermediary metabolism. J. Proteome Res. 2014, 13, 1938–1956. [Google Scholar] [CrossRef]
- Walther, D.M.; Kasturi, P.; Zheng, M.; Pinkert, S.; Vecchi, G.; Ciryam, P.; Morimoto, R.I.; Dobson, C.M.; Vendruscolo, M.; Mann, M.; et al. Widespread proteome remodeling and aggregation in aging C. elegans. Cell 2015, 161, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, S.; Bundy, J.G.; Davies, S.K.; Viney, J.M.; Swire, J.S.; Leroi, A.M. A metabolic signature of long life in Caenorhabditis elegans. BMC Biol. 2010, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.W.; Smith, R.L.; Van Weeghel, M.; Kamble, R.; Janssens, G.E.; Houtkooper, R.H. Identification of key pathways and metabolic fingerprints of longevity in C. elegans. Exp. Gerontol. 2018, 113, 128–140. [Google Scholar] [CrossRef]
- Shaw, W.M.; Luo, S.; Landis, J.; Ashraf, J.; Murphy, C.T. The C. elegans tgf-beta dauer pathway regulates longevity via insulin signaling. Curr. Biol. 2007, 17, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Gems, D.; Sutton, A.J.; Sundermeyer, M.L.; Albert, P.S.; King, K.V.; Edgley, M.L.; Larsen, P.L.; Riddle, D.L. Two pleiotropic classes of daf-2 mutation affect larval arrest, adult behavior, reproduction and longevity in Caenorhabditis elegans. Genetics 1998, 150, 129–155. [Google Scholar]
- Ewald, C.Y.; Landis, J.N.; Porter Abate, J.; Murphy, C.T.; Blackwell, T.K. Dauer-independent insulin/igf-1-signalling implicates collagen remodelling in longevity. Nature 2015, 519, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewald, C.Y.; Castillo-Quan, J.I.; Blackwell, T.K. Untangling longevity, dauer, and healthspan in Caenorhabditis elegans insulin/igf-1-signalling. Gerontology 2018, 64, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Lithgow, G.J.; White, T.M.; Melov, S.; Johnson, T.E. Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proc. Natl. Acad. Sci. USA 1995, 92, 7540–7544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanfleteren, J.R. Oxidative stress and ageing in Caenorhabditis elegans. Biochem. J. 1993, 292, 605–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, Y.; Honda, S. The daf-2 gene network for longevity regulates oxidative stress resistance and mn-superoxide dismutase gene expression in Caenorhabditis elegans. FASEB J. 1999, 13, 1385–1393. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Johnson, T.E. A genetic pathway conferring life extension and resistance to uv stress in Caenorhabditis elegans. Genetics 1996, 143, 1207–1218. [Google Scholar]
- Lamitina, S.T.; Strange, K. Transcriptional targets of daf-16 insulin signaling pathway protect C. elegans from extreme hypertonic stress. Am. J. Physiol. Cell Physiol. 2005, 288, C467–C474. [Google Scholar] [CrossRef]
- Scott, B.A.; Avidan, M.S.; Crowder, C.M. Regulation of hypoxic death in C. elegans by the insulin/igf receptor homolog daf-2. Science 2002, 296, 2388–2391. [Google Scholar] [CrossRef]
- Mabon, M.E.; Scott, B.A.; Crowder, C.M. Divergent mechanisms controlling hypoxic sensitivity and lifespan by the daf-2/insulin/igf-receptor pathway. PLoS ONE 2009, 4, e7937. [Google Scholar] [CrossRef]
- Barsyte, D.; Lovejoy, D.A.; Lithgow, G.J. Longevity and heavy metal resistance in daf-2 and age-1 long-lived mutants of Caenorhabditis elegans. FASEB J. 2001, 15, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Garsin, D.A.; Villanueva, J.M.; Begun, J.; Kim, D.H.; Sifri, C.D.; Calderwood, S.B.; Ruvkun, G.; Ausubel, F.M. Long-lived C. elegans daf-2 mutants are resistant to bacterial pathogens. Science 2003, 300, 1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, E.A.; Chen, W.C.; Tan, M.-W. The daf-2 insulin-like signaling pathway independently regulates aging and immunity in C. elegans. Aging Cell 2008, 7, 879–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, W.-C.; Ching, T.-T.; Lee, H.C.; Mousigian, C.; Hsu, A.-L. Hsf-1 regulators ddl-1/2 link insulin-like signaling to heat-shock responses and modulation of longevity. Cell 2012, 148, 322–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.; Aballay, A. Heat-shock transcription factor (hsf)-1 pathway required for Caenorhabditis elegans immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 13092–13097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.H.; Blackwell, T.K. Skn-1 links C. elegans mesendodermal specification to a conserved oxidative stress response. Genes Dev. 2003, 17, 1882–1893. [Google Scholar] [CrossRef] [Green Version]
- Tullet, J.M.A.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor skn-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Papp, D.; Csermely, P.; Sőti, C. A role for skn-1/nrf in pathogen resistance and immunosenescence in Caenorhabditis elegans. PLoS Pathog. 2012, 8, e1002673. [Google Scholar] [CrossRef] [Green Version]
- Wolff, S.; Ma, H.; Burch, D.; Maciel, G.A.; Hunter, T.; Dillin, A. Smk-1, an essential regulator of daf-16-mediated longevity. Cell 2006, 124, 1039–1053. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.-W.; et al. A conserved p38 map kinase pathway in Caenorhabditis elegans innate immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef] [Green Version]
- Larsen, P.L. Aging and resistance to oxidative damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1993, 90, 8905–8909. [Google Scholar] [CrossRef] [Green Version]
- Houthoofd, K.; Fidalgo, M.A.; Hoogewijs, D.; Braeckman, B.P.; Lenaerts, I.; Brys, K.; Matthijssens, F.; De Vreese, A.; Van Eygen, S.; Muñoz, M.J.; et al. Metabolism, physiology and stress defense in three aging ins/igf-1 mutants of the nematode Caenorhabditis elegans. Aging Cell 2005, 4, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, Y.; Tanaka, M.; Honda, S. Modulation of longevity and diapause by redox regulation mechanisms under the insulin-like signaling control in Caenorhabditis elegans. Exp. Gerontol. 2008, 43, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Doonan, R.; McElwee, J.J.; Matthijssens, F.; Walker, G.A.; Houthoofd, K.; Back, P.; Matscheski, A.; Vanfleteren, J.R.; Gems, D. Against the oxidative damage theory of aging: Superoxide dismutases protect against oxidative stress but have little or no effect on life span in Caenorhabditis elegans. Genes Dev. 2008, 22, 3236–3241. [Google Scholar] [CrossRef] [Green Version]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef] [Green Version]
- Dues, D.J.; Andrews, E.K.; Senchuk, M.M.; Van Raamsdonk, J.M. Resistance to stress can be experimentally dissociated from longevity. J. Gerontol. A Biol. 2018, 74, 1206–1214. [Google Scholar] [CrossRef] [Green Version]
- Erkut, C.; Vasilj, A.; Boland, S.; Habermann, B.; Shevchenko, A.; Kurzchalia, T.V. Molecular strategies of the Caenorhabditis elegans dauer larva to survive extreme desiccation. PLoS ONE 2013, 8, e82473. [Google Scholar] [CrossRef] [Green Version]
- Lant, B.; Storey, K.B. An overview of stress response and hypometabolic strategies in Caenorhabditis elegans: Conserved and contrasting signals with the mammalian system. Int. J. Biol. Sci. 2010, 6, 9–50. [Google Scholar] [CrossRef] [Green Version]
- Burnell, A.M.; Houthoofd, K.; O’Hanlon, K.; Vanfleteren, J.R. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Exp. Gerontol. 2005, 40, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Roy, R. Amp-activated kinase regulates lipid droplet localization and stability of adipose triglyceride lipase in C. elegans dauer larvae. PLoS ONE 2015, 10, e0130480. [Google Scholar] [CrossRef] [Green Version]
- Narbonne, P.; Roy, R. Caenorhabditis elegans dauers need lkb1/ampk to ration lipid reserves and ensure long-term survival. Nature 2009, 457, 210–214. [Google Scholar] [CrossRef]
- Yen, K.; Le, T.T.; Bansal, A.; Narasimhan, S.D.; Cheng, J.-X.; Tissenbaum, H.A. A comparative study of fat storage quantitation in nematode Caenorhabditis elegans using label and label-free methods. PLoS ONE 2010, 5, e12810. [Google Scholar] [CrossRef] [PubMed]
- Frazier, H.N., III; Roth, M.B. Adaptive sugar provisioning controls survival of C. elegans embryos in adverse environments. Curr. Biol. 2009, 19, 859–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, C.L.; Van Gilst, M.R. A 13C isotope labeling strategy reveals the influence of insulin signaling on lipogenesis in C. elegans. Cell Metab. 2008, 8, 266–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shmookler Reis, R.J.; Xu, L.; Lee, H.; Chae, M.; Thaden, J.J.; Bharill, P.; Tazearslan, C.; Siegel, E.; Alla, R.; Zimniak, P.; et al. Modulation of lipid biosynthesis contributes to stress resistance and longevity of C. elegans mutants. Aging 2011, 3, 125–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, C.; Krumsiek, J.; Lehrbach, N.J.; Murfitt, S.A.; Miska, E.A.; Griffin, J.L. A study of Caenorhabditis elegans daf-2 mutants by metabolomics and differential correlation networks. Mol. Biosyst. 2013, 9, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Possik, E.; Ajisebutu, A.; Manteghi, S.; Gingras, M.-C.; Vijayaraghavan, T.; Flamand, M.; Coull, B.; Schmeisser, K.; Duchaine, T.; Van Steensel, M.; et al. Flcn and ampk confer resistance to hyperosmotic stress via remodeling of glycogen stores. PLoS Genet. 2015, 11, e1005520. [Google Scholar] [CrossRef] [Green Version]
- LaMacchia, J.C.; Frazier, H.N., 3rd; Roth, M.B. Glycogen fuels survival during hyposmotic-anoxic stress in Caenorhabditis elegans. Genetics 2015, 201, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Gusarov, I.; Pani, B.; Gautier, L.; Smolentseva, O.; Eremina, S.; Shamovsky, I.; Katkova-Zhukotskaya, O.; Mironov, A.; Nudler, E. Glycogen controls Caenorhabditis elegans lifespan and resistance to oxidative stress. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Seo, Y.; Kingsley, S.; Walker, G.; Mondoux, M.A.; Tissenbaum, H.A. Metabolic shift from glycogen to trehalose promotes lifespan and healthspan in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2018, 115, E2791. [Google Scholar] [CrossRef] [Green Version]
- Clegg, J.S.; Evans, D.R. The physiology of blood trehalose and its function during flight in the blowfly. J. Exp. Biol. 1961, 38, 771–792. [Google Scholar]
- Erkut, C.; Penkov, S.; Khesbak, H.; Vorkel, D.; Verbavatz, J.-M.; Fahmy, K.; Kurzchalia, T.V. Trehalose renders the dauer larva of Caenorhabditis elegans resistant to extreme desiccation. Curr. Biol. 2011, 21, 1331–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia, H.; Young, L.; Fox, D.; Bertozzi, C.R.; Koshland, D. Increasing intracellular trehalose is sufficient to confer desiccation tolerance to Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2015, 112, 6122–6127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, B.; Haltiwanger, R.S.; Futcher, B. Synergy between trehalose and hsp104 for thermotolerance in Saccharomyces cerevisiae. Genetics 1996, 144, 923–933. [Google Scholar] [PubMed]
- Page-Sharp, M.; Behm, C.A.; Smith, G.D. Involvement of the compatible solutes trehalose and sucrose in the response to salt stress of a cyanobacterial scytonema species isolated from desert soils. Biochim. Biophys. Acta 1999, 1472, 519–528. [Google Scholar] [CrossRef]
- Pellerone, F.I.; Archer, S.K.; Behm, C.A.; Grant, W.N.; Lacey, M.J.; Somerville, A.C. Trehalose metabolism genes in Caenorhabditis elegans and filarial nematodes. Int. J. Parasitol. 2003, 33, 1195–1206. [Google Scholar] [CrossRef]
- Goyal, K.; Browne, J.A.; Burnell, A.M.; Tunnacliffe, A. Dehydration-induced tps gene transcripts from an anhydrobiotic nematode contain novel spliced leaders and encode atypical gt-20 family proteins. Biochimie 2005, 87, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Honda, Y.; Tanaka, M.; Honda, S. Trehalose extends longevity in the nematode Caenorhabditis elegans. Aging Cell 2010, 9, 558–569. [Google Scholar] [CrossRef]
- Erkut, C.; Gade, V.R.; Laxman, S.; Kurzchalia, T.V. The glyoxylate shunt is essential for desiccation tolerance in C. elegans and budding yeast. eLife 2016, 5, e13614. [Google Scholar] [CrossRef]
- Liu, F.; Thatcher, J.D.; Barral, J.M.; Epstein, H.F. Bifunctional glyoxylate cycle protein of Caenorhabditis elegans: A developmentally regulated protein of intestine and muscle. Dev. Biol. 1995, 169, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Braeckman, B.P.; Houthoofd, K.; Vanfleteren, J.R. Intermediary Metabolism. Available online: http://www.wormbook.org/chapters/www_intermetabolism/intermetabolism.html (accessed on 31 December 2019).
- Shen, E.-Z.; Song, C.-Q.; Lin, Y.; Zhang, W.-H.; Su, P.-F.; Liu, W.-Y.; Zhang, P.; Xu, J.; Lin, N.; Zhan, C.; et al. Mitoflash frequency in early adulthood predicts lifespan in Caenorhabditis elegans. Nature 2014, 508, 128–132. [Google Scholar] [CrossRef]
- Holt, S.J.; Riddle, D.L. Sage surveys C. elegans carbohydrate metabolism: Evidence for an anaerobic shift in the long-lived dauer larva. Mech. Ageing Dev. 2003, 124, 779–800. [Google Scholar] [CrossRef]
- Yilmaz, L.S.; Walhout, A.J. A Caenorhabditis elegans genome-scale metabolic network model. Cell Syst. 2016, 2, 297–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabortee, S.; Boschetti, C.; Walton, L.J.; Sarkar, S.; Rubinsztein, D.C.; Tunnacliffe, A. Hydrophilic protein associated with desiccation tolerance exhibits broad protein stabilization function. Proc. Natl. Acad. Sci. USA 2007, 104, 18073–18078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.; Davies, J.E.; Huang, Z.; Tunnacliffe, A.; Rubinsztein, D.C. Trehalose, a novel mtor-independent autophagy enhancer, accelerates the clearance of mutant huntingtin and α-synuclein. J. Biol. Chem. 2007, 282, 5641–5652. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.M.; Campbell, L.T.; Lodge, J.K. Cryptococcus neoformans, a fungus under stress. Curr. Opin. Microbiol. 2007, 10, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Challabathula, D.; Bartels, D. Desiccation tolerance in resurrection plants: New insights from transcriptome, proteome and metabolome analysis. Front. Plant. Sci. 2013, 4, 482. [Google Scholar]
- Alpert, P. Constraints of tolerance: Why are desiccation-tolerant organisms so small or rare? J. Exp. Biol. 2006, 209, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.; Lithgow, G.J.; Link, W. Long live foxo: Unraveling the role of foxo proteins in aging and longevity. Aging Cell 2016, 15, 196–207. [Google Scholar] [CrossRef]
- Stefanetti, R.J.; Voisin, S.; Russell, A.; Lamon, S. Recent advances in understanding the role of foxo3. F1000Res 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Morris, B.J.; Willcox, D.C.; Donlon, T.A.; Willcox, B.J. Foxo3: A major gene for human longevity—A mini-review. Gerontology 2015, 61, 515–525. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Simplified graphic summary of the DAF-16-dependent metabolic remodeling in the Caenorhabditis elegans daf-2 mutant, with emphasis on the major carbon stores and their interconversion.
Figure 1.
Simplified graphic summary of the DAF-16-dependent metabolic remodeling in the Caenorhabditis elegans daf-2 mutant, with emphasis on the major carbon stores and their interconversion.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zečić, A.; Braeckman, B.P. DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling. Cells 2020, 9, 109. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010109
AMA Style
Zečić A, Braeckman BP. DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling. Cells. 2020; 9(1):109. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010109
Chicago/Turabian StyleZečić, Aleksandra, and Bart P. Braeckman. 2020. "DAF-16/FoxO in Caenorhabditis elegans and Its Role in Metabolic Remodeling" Cells 9, no. 1: 109. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010109
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.