Mitophagy in Alzheimer’s Disease and Other Age-Related Neurodegenerative Diseases


Department of Cell Biology and Neuroscience, Rutgers, The State University of New Jersey, Piscataway, NJ 08854, USA
*
Author to whom correspondence should be addressed.
Cells 2020, 9(1), 150; https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010150
Submission received: 19 December 2019
/
Revised: 3 January 2020
/
Accepted: 5 January 2020
/
Published: 8 January 2020
(This article belongs to the Section Mitochondria)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Mitochondrial dysfunction is a central aspect of aging and neurodegenerative diseases, including Alzheimer’s disease, Parkinson’s disease, amyotrophic lateral sclerosis, and Huntington’s disease. Mitochondria are the main cellular energy powerhouses, supplying most of ATP by oxidative phosphorylation, which is required to fuel essential neuronal functions. Efficient removal of aged and dysfunctional mitochondria through mitophagy, a cargo-selective autophagy, is crucial for mitochondrial maintenance and neuronal health. Mechanistic studies into mitophagy have highlighted an integrated and elaborate cellular network that can regulate mitochondrial turnover. In this review, we provide an updated overview of the recent discoveries and advancements on the mitophagy pathways and discuss the molecular mechanisms underlying mitophagy defects in Alzheimer’s disease and other age-related neurodegenerative diseases, as well as the therapeutic potential of mitophagy-enhancing strategies to combat these disorders.
1. Introduction
Mitochondria are termed the “powerhouses” of the cell, and generate the majority of the cell’s supply of adenosine triphosphate (ATP) through the oxidative phosphorylation system (OXPHOS) in which electrons produced by the citric acid cycle are transferred down the mitochondrial respiratory complexes. Neurons have particularly high and continuous energy demands so that mitochondrial function is essential for maintaining neuronal integrity and responsiveness [1,2,3,4,5,6,7]. Mitochondrial energy production fuels various critical neuronal functions, especially the ATP-dependent neurotransmission [1,3,8]. Along with regulating energy levels, mitochondria have a high capacity to sequester excessive Ca2+ and release Ca2+ so as to prolong residual levels at synaptic terminals [9,10]. Through this mechanism, mitochondria play essential roles in maintaining and regulating neurotransmission [11,12], as well as certain types of short-term synaptic plasticity [13,14]. In addition, mounting evidence has demonstrated the critical role of mitochondria in the maintenance of cellular homeostasis [15]. Glucose was shown to be an efficient energy source in neurons and glia that can consume energy produced in parallel by glycolysis and OXPHOS. However, upon neural network activation, the energy demand is robustly enhanced. Given ATP as the main energy source in neurons, mitochondrial energy metabolism thus may play a major role in supplying ATP to fuel these neuronal activities. Importantly, distinct mitochondrial energetic status might also have a significant impact on the cellular signaling pathways. Aged and dysfunctional mitochondria are defective in ATP production and Ca2+ buffering, leading to energy deficit and interruptions of neuronal function and health. Furthermore, damaged mitochondria trigger concomitant leakage of electrons and thus promote the production of harmful reactive oxygen species (ROS) that can damage nucleic acids, proteins, and membrane lipids [1,16,17,18]. Moreover, mitochondrial oxidative stress leads to the release of cytochrome c, a mitochondrial intermembrane space protein, into the cytosol, inducing DNA damage, caspase activation, and apoptosis [6].
A large body of work suggests that mitochondrial dysfunction underlies cognitive decline in neuronal aging and is one of the most notable hallmarks of age-associated neurodegenerative diseases. Mitochondrial damage causes energy deficit, oxidative stress, and impaired cellular signaling, which has been linked to the pathogenesis of neurodegeneration diseases [2,19,20,21]. Given that a mitochondrion’s half-life is estimated to be about 30 days [22,23], cells have developed the interconnected and elaborate pathways through the balance of mitochondrial biogenesis and efficient removal of damaged mitochondria to ensure the maintenance of mitochondrial integrity and bioenergetic functions. Mitophagy, a selective form of autophagy, constitutes a key pathway of mitochondrial quality control mechanisms involving sequestration of defective mitochondria into autophagosomes for subsequent lysosomal degradation [1,2,24,25]. Disruption of mitophagy has been indicated in aging and various diseases, including neurodegenerative disorders such as Alzheimer’s disease (AD), Parkinson’s disease (PD), amyotrophic lateral sclerosis (ALS), and Huntington’s disease (HD) [7,26]. This review aims to provide a thorough and timely overview of the mitophagy pathways, summarize the underlying mechanisms of mitophagy defects in AD and other age-related neurodegenerative diseases, and highlight the possible therapeutic strategies targeting mitophagy towards confronting mitochondrial dysfunction and neurodegeneration.
2. Overview of the Mitophagy Pathways
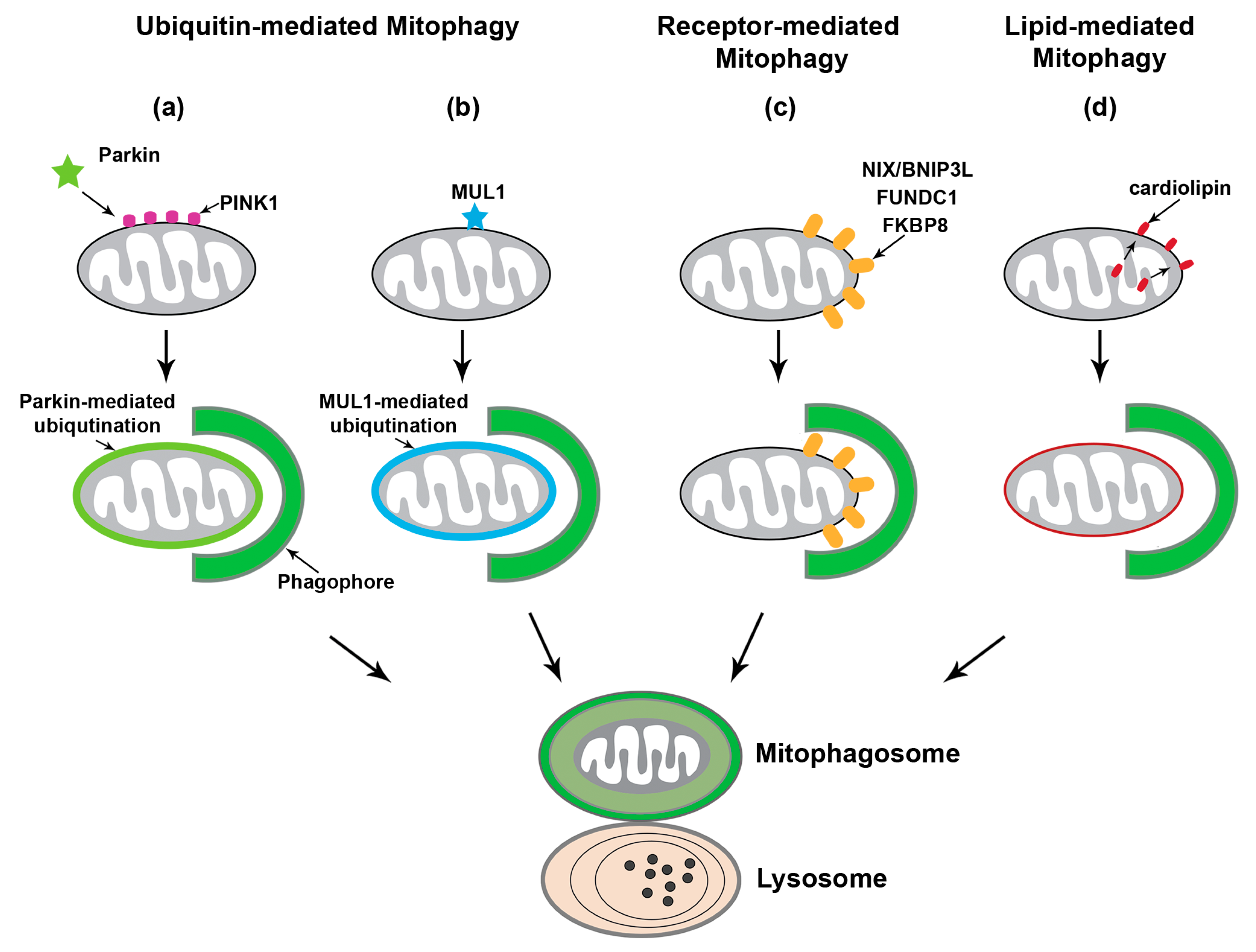
Mitophagy (mitochondrial autophagy) is the only known cellular pathway through which entire mitochondria are completely eliminated within lysosomes. Under physiological conditions, mitophagy plays an essential role in the basal mitochondrial turnover and maintenance. More importantly, mitophagy can also be robustly induced in response to a variety of pathological stimuli [7,25,26,27]. There are a number of mitophagy pathways that have been identified (Figure 1).
2.1. PINK1-Parkin-Mediated Mitophagy
PTEN-induced putative kinase protein 1 (PINK1)-Parkin-mediated mitophagy is the most heavily studied and the best-understood mitophagy pathway [28,29,30]. In brief, loss of mitochondrial membrane potential (Δψm) accumulates PINK1 on the outer membrane of mitochondria (OMM) to recruit and activate Parkin, an E3 ubiquitin ligase, through phosphorylation of ubiquitin [31,32,33,34,35,36,37,38]. Parkin then ubiquitinates a number of OMM proteins and subsequently activates the ubiquitin-proteasome system (UPS) to degrade these ubiquitinated OMM proteins [39,40,41,42,43]. This leads to recruitment of the autophagy machinery to promote the engulfment of damaged mitochondria by phagophore or isolation membranes and thus formation of mitophagosomes destined for removal via the lysosomal system. The roles of PINK1 and Parkin in mitochondrial quality control and mitophagy have been supported by multiple studies in Drosophila [28,44,45,46,47]. The PINK1-Parkin pathway was shown to facilitate mitophagy as well as selective mitochondrial respiratory chain turnover [28,44,45,46,47]. Furthermore, genetic and clinical data have provided clear evidence to support the notion that the PINK1-Parkin pathway is involved in the pathogenesis of PD [48,49]. However, recent in vivo studies indicate that PINK1 and Parkin are not critical for basal mitophagy in a range of tissues including the brain [50,51]. More recent studies have been focused on understanding the PINK1-Parkin-independent mitophagy pathways.
2.2. Ubiquitin-Mediated Mitophagy Independent of Parkin
Other E3 ubiquitin ligases that can also mediate removal dysfunctional mitochondria have been identified [52], which is with relation to PINK1-Parkin-independent mitophagy mechanisms. Mitochondrial ubiquitin ligase 1 (MUL1, also known as MAPL, GIDE, and MULAN) was reported to play a role in the regulation of mitophagy through multiple mechanisms. MUL1 interacts with mitochondrial fission GTPase protein dynamin-related protein 1 (Drp1) and mitochondrial fusion protein Mitofusin, both of which are the substrates of Parkin [53,54]. MUL1 has no effect on PINK1-Parkin-mediated mitophagy, but can suppress PINK1 or Parkin mutant phenotypes in both Drosophila and mouse neurons. This suppression is attributed to the ubiquitin-dependent degradation of Mitofusin. Interestingly, double mutants of MUL1 with either PINK1 or Parkin show much more severe phenotypes. Moreover, MUL1 contains an LC3-interacting region (LIR) motif in the RING domain through which MUL1 interacts with GABAA receptor-associated protein (GABARAP), a member of the Atg8 family that plays a key role in autophagy and mitophagy [55]. Thus, these observations collectively suggest that MUL1 functions in a pathway parallel to the PINK1-Parkin pathway. In addition to MUL1, a recent study reported PINK1-synphilin-1-SIAH-1 as another newly discovered Parkin-independent pathway that can promote PINK1-dependent mitophagy in the absence of Parkin [56].
2.3. Receptor-Mediated Mitophagy
The BCL-2 homology 3 (BH3)-containing protein NIP3-like X (NIX, also known as BNIP3L), an OMM protein, was reported to play an important role in mitochondrial turnover in erythrocytes [57]. NIX/BNIP3L contains an LIR motif at the amino-terminal that binds to LC3 on phagophore or isolation membranes, and is transcriptionally upregulated during erythrocyte differentiation [58]. Such a mechanism enables NIX/BNIP3L to serve as a selective mitophagy receptor and promote recruitment of the autophagy machinery to the surface of damaged mitochondria in erythroid cells. NIX/BNIP3L was also reported to be involved in hypoxia-induced mitophagy, during which forkhead box O3 (FOXO3) and hypoxia-inducible factor (HIF) transcriptionally regulate NIX/BNIP3L along with BNIP3 [59]. Noteworthy, overexpression of NIX/BNIP3L can restore mitophagy in skin fibroblasts from PD patients carrying mutations in PARK6 or PARK2 [60], suggesting an independent role of NIX/BNIP3L in PINK1-Parkin-mediated mitophagy. NIX/BNIP3L and BNIP3 were reported to be upregulated upon neuronal stress [61,62]. However, the extent to which NIX/BNIP3L and BNIP3 might participate in neuronal mitophagy remains unclear. FUN14 domain containing 1 (FUNDC1) also functions as a mitophagy receptor and regulates the autophagic clearance of mitochondria under hypoxic stress. Studies have demonstrated that the mitochondrial phosphatase phosphoglycerate mutase family member 5 (PGAM5) dephosphorylates FUNDC1 to activate mitophagy during hypoxia [63,64,65]. Additionally, FK506 Binding Protein 8 (FKBP8) was recently reported to have LIR domains and can mediate Parkin-independent mitophagy by recruiting LC3A [66]. Collectively, these observations suggest that specific mitophagy receptors on the OMM play an essential role in recruiting the autophagy machinery to damaged mitochondria for lysosomal clearance.
2.4. Lipid-Mediated Mitophagy
Recent studies have demonstrated that lipids can also act as an elimination signal to mediate recruitment of injured mitochondria to the autophagy pathway. Apart from ubiquitin- or receptor-mediated mitophagy, this pathway involves the direct interaction of LC3 with the phospholipid cardiolipin, and was originally observed in neuroblastoma cells and primary cortical neurons incubated with rotenone, staurosporine, or 6-hydroxydopamine [67]. Cardiolipin is primarily found in the inner membrane of mitochondria (IMM) and is externalized to the OMM upon mitochondrial damage. Three enzymatic translocations are needed for the externalization of cardiolipin, which are mediated by the phospholipid scramblase-3 of mitochondria and the inner and outer membrane spanning hexameric complex of mitochondrial nucleoside diphosphate kinase D (NDPK-D/NM23-H4) in SH-SY5Y cells or Tafazzin (TAZ) in mouse embryonic fibroblasts (MEFs), respectively [67,68,69]. Furthermore, cardiolipin interacts with LC3, and this interaction is facilitated by the negatively charged basic residues in LC3 and charged head group of cardiolipin. Thus, cardiolipin-mediated mitophagy is independent of PINK1 and Parkin. Importantly, cardiolipin downregulation or mutagenesis of LC3 at the sites predicted to interact with cardiolipin was shown to impair mitophagosome formation [67]. In addition, genome-wide screens indicate that F-box and WD40 domain protein 7 (FBXW7), sterol regulatory element binding transcription factor 1 (SREBF1), and other components of the lipogenesis pathway may play a role in the regulation of Parkin-mediated mitophagy [70]. Additionally, upon Drp1-mediated mitochondrial fission, ceramide was shown to promote autophagic recruitment of mitochondria through direct interaction of ceramide with LC3B-II [71].
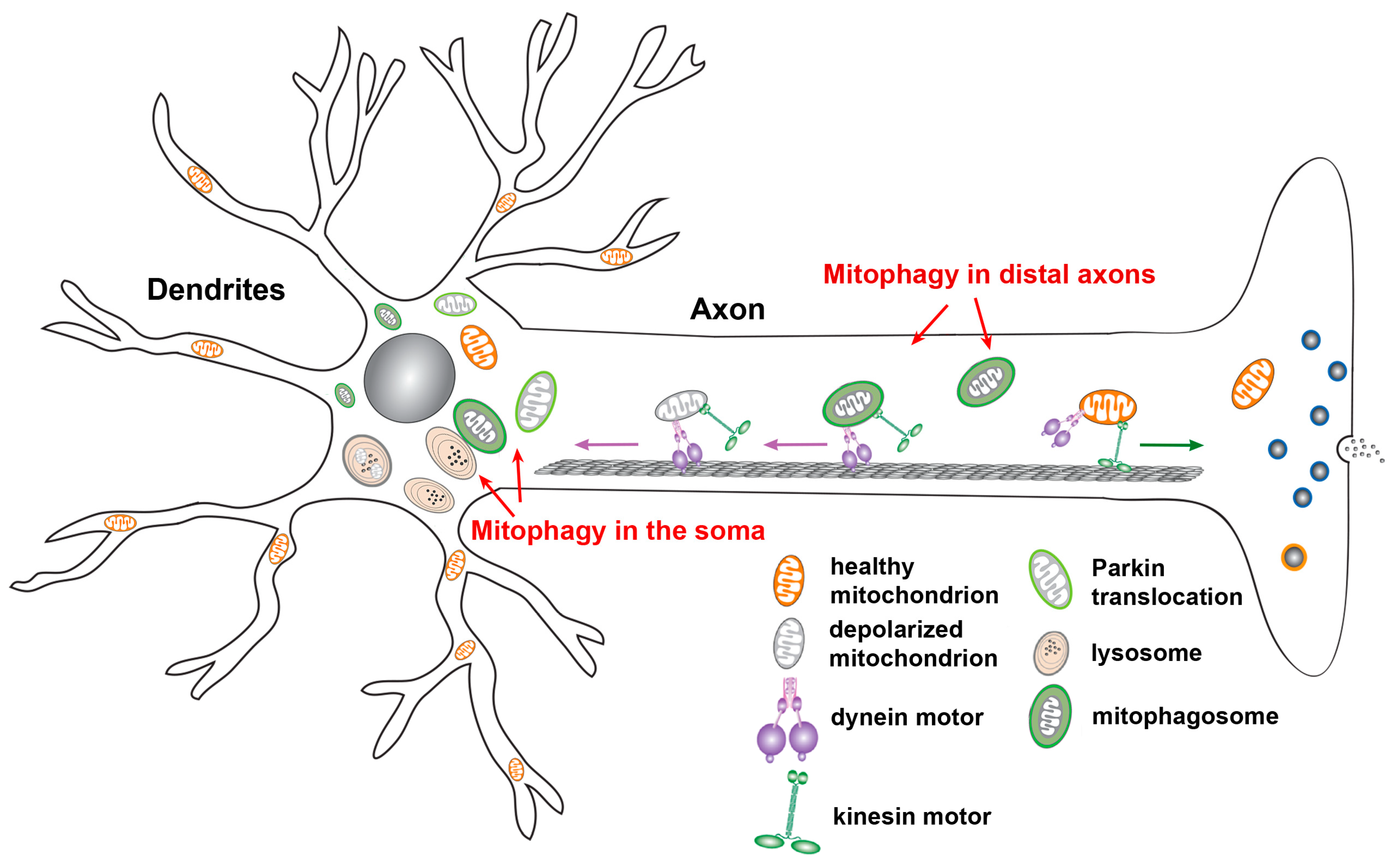
2.5. Neuronal Mitophagy
Neurons are highly polarized cells with unique properties in structure and function. Mitochondrial quality control mechanisms that efficiently sense and eliminate mitochondria damaged over usage, aging, or disease could be critical for neuronal health. Mitophagy is currently believed to constitute the major cellular pathway for mitochondrial quality control in neurons. While basal mitophagy is known to be required for the maintenance of neuronal homeostasis, mounting evidence has shown that mitophagy can be upregulated in response to various pathological stimuli (Figure 2). Cardiolipin-mediated mitophagy can be induced in primary cortical neurons treated with the mitochondrial complex I inhibitor rotenone [67]. As for Parkin-mediated mitophagy, Δψm dissipation triggers Parkin translocation onto depolarized mitochondria in neurons after treatment with CCCP, an Δψm uncoupler [72,73]. Interestingly, Parkin-targeted mitochondria primarily accumulate in the somatodendritic region of neurons where they undergo autophagic sequestration for lysosomal degradation. Moreover, mitophagy activation reduces anterograde transport, but increases retrograde transport of axonal mitochondria, suggesting that damaged mitochondria are trafficked back to the soma for mitophagic clearance. Parkin-dependent mitophagy was also discovered under AD-linked pathophysiological conditions in the absence of any Δψm dissipating reagent [74]. The spatial aspects of Parkin-dependent mitophagy were also observed in vivo. In particular, the PINK1 and Parkin mutant Drosophila exhibit abnormal tubular and reticular mitochondria restricted to the cell body, as well as normal morphology with reduced mitochondrial flux within axons [46,47]. In addition to Drosophila, the evidence from examination of Purkinje neurons in the mito-QC reporter mice suggests that the majority of mitochondrial turnover occurs in the Purkinje somata. This supports the view that damaged mitochondria or mitophagosomes are returned to the cell body for lysosomal clearance [75]. Collectively, these in vitro and in vivo observations consistently suggest that the soma is in the focus of neuronal mitophagy, a selective process with a function to restrict damaged mitochondria to the soma and thus limit the impact of impaired mitochondrial function on distal axons.
2.6. Mitophagy In Vivo
The mitophagy pathways have been extensively studied in vitro. To address the basal mitophagy in vivo, a number of transgenic mice expressing sensors to monitor the delivery of mitochondria to acidic organelles (lysosomes) have been developed [75,76]. These studies have demonstrated active mitochondrial delivery to acidic organelles in multiple tissues but with variable rates. A recent work further shows that the basal mitophagy is independent of the PINK1 pathway [51]. Consistently, studies from Drosophila expressing fluorescent mitophagy reporters, either mito-Keima or mito-QC, also reveal robust basal mitophagy in different tissues [50]. However, null mutations of either PINK1 or Parkin do not lead to altered rates of mitochondrial delivery into lysosomes, suggesting nonessential roles of PINK1 and Parkin in the basal mitophagy in vivo. These data are also consistent with the observations from mice with the deletion of PARK6 or PARK2. These mice lack strong phenotypes, such as dopaminergic neuron loss [77,78,79,80,81]. Importantly, the evidence of mitophagy activation is clear in the brain tissues of human patients with neurodegenerative diseases [74,82,83]. Given multiple distinct mechanisms that have been identified to target damaged mitochondria for autophagy, other PINK1-Parkin-independent pathways or other as yet undefined mechanisms likely play more important role in the basal neuronal mitophagy. Therefore, the involvement of these mitophagy pathways in the basal mitochondrial turnover and in response to specific disease-related stressors needs to be carefully determined in vivo.
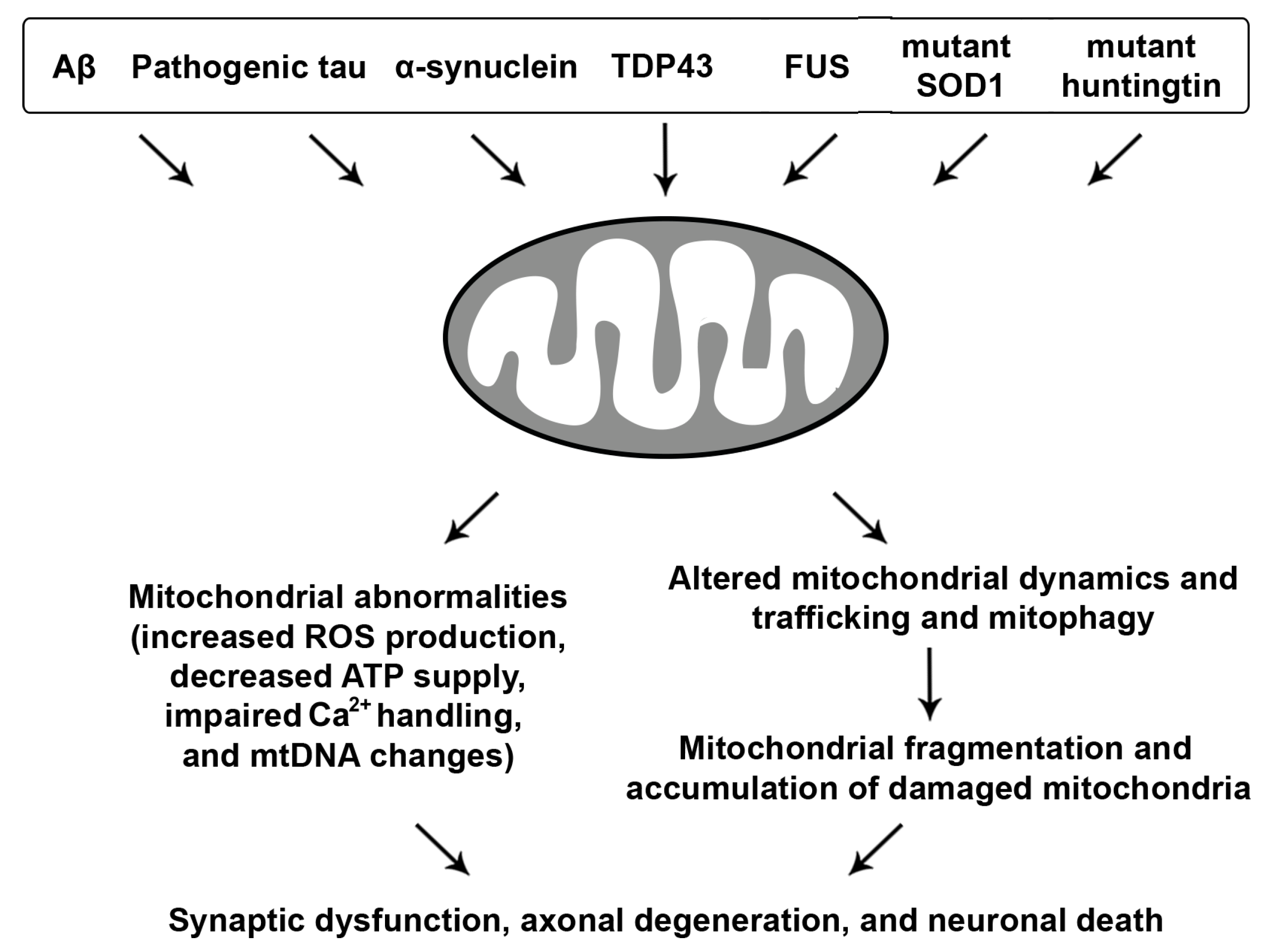
3. Mitochondrial Dysfunction in Neurodegenerative Diseases
Mitochondrial defects are a significant concern in the aging nervous system and have been consistently linked to age-related neurodegenerative diseases, suggesting that the underlying mechanisms might be somewhat shared (Figure 3).
3.1. Aβ and Tau-Linked Mitochondrial Abnormalities
AD is the most common form of neurodegenerative diseases in aging populations. Progression of the disease involves cognitive decline, memory loss, and neuronal death in the cerebral cortex and subcortical regions. AD patient brains are characterized by extracellular amyloid plaque deposits, composed of agglomerated amyloid β (Aβ) peptides, as well as intracellular accumulation of neurofibrillary tangles (NFTs), consisting of hyperphosphorylated tau (phospho-tau) protein. Mitochondrial disturbances have been suggested as a hallmark of AD as the patients exhibit early metabolic alterations prior to any histopathological or clinical manifestations [84]. Mitochondrial dysfunction, oxidative stress, and mitochondrial DNA (mtDNA) changes are prominent pathological features reported in AD postmortem brains [85,86,87,88,89,90,91,92,93,94,95]. Importantly, a growing body of evidence has indicated a major role of mitochondrial defects in the pathogenesis of AD [2,96,97,98].
The degree of cognitive dysfunction in AD was linked to the extent of Aβ accumulation within mitochondria and mitochondrial abnormalities [99]. Aβ has been proposed to be a key player in mediating mitochondrial damage. Aβ was found to impair multiple aspects of mitochondrial function [100,101,102], including function of the electron transport chain (ETC) [103], ROS production [104,105,106], mitochondrial dynamics [91,103,107,108], and mitochondrial transport [109,110,111]. The possible routes for Aβ to enter into mitochondria were thought to be through the translocase of the outer membrane (TOM) complex or mitochondrial-associated endoplasmic reticulum (ER) membrane (MAM) [112,113,114,115]. In addition to intracellular Aβ, mitochondria can also take up internalized extracellular Aβ [114,116]. Aβ1–42 treatment was shown to lead to the opening of mitochondrial permeability transition pore (mPTP) in cultured cortical neural progenitor cells. While transient mPTP opening decreases cell proliferation, prolonged mPTP opening irreversibly causes cell death [117]. Consistent with this observation, an interesting work in a live AD mouse model provided direct evidence that fragmented and defective mitochondria are limited to the vicinity of extracellular amyloid plaques that likely serve as a focal source to promote abnormal accumulation of Aβ within mitochondria and thus exacerbate Aβ-linked damage [118].
The mechanisms underlying Aβ-mediated mitochondrial toxicity have been carefully investigated by several studies. The interactions of Aβ with Aβ-binding alcohol dehydrogenase (ABAD), a mitochondrial matrix protein, and cyclophilin D (CypD), a component of the mitochondrial transition pore, were reported to mediate Aβ-induced cytotoxic effects [101,102]. In particular, ABAD was shown to be upregulated in AD neurons. Overexpression of ABAD can exacerbate Aβ-induced cellular oxidative stress and cell death. Aβ also forms a complex with CypD in the cortical regions of postmortem human AD patient brains and an AD mouse model [102]. Deletion of CypD in AD mice rescues the mitochondrial phenotypes including impaired Ca2+ uptake, mitochondrial swelling due to increased Ca2+, depolarized Δψm, elevated oxidative stress, decrease in ADP-induced respiration control rate, and reduced complex IV activity and ATP levels. Moreover, CypD deficiency can improve synaptic function as well as learning and memory in an AD mouse model [102]. These observations collectively suggest that Aβ-CypD interaction mediates AD-associated mitochondrial defects. Taken together, these pieces of evidence indicate that the aberrant accumulation of Aβ within mitochondria likely plays a causative role in impaired mitochondrial function in AD.
Pathogenic forms of tau can also induce mitochondrial damage. A number of studies have demonstrated that phospho-tau specifically impairs complex I of the mitochondrial respiratory chain, resulting in increased ROS production, loss of Δψm, lipid peroxidation, and reduced activities of detoxifying enzymes such as superoxide dismutase (SOD) [119,120]. Overexpression of the mutant human tau protein htauP301L was reported to reduce ATP levels and increase susceptibility to oxidative stress in cultured neuroblastoma cells [121]. Disrupted activity and altered composition of mitochondrial enzymes can also be detected in the P301S mouse model of tauopathy [122]. In the pR5 mice overexpressing the htauP301L, mitochondrial dysfunction was evidenced by impaired mitochondrial respiration and ATP synthesis, decreased complex I activity, and increased ROS levels [123,124]. Phospho-tau was also reported to directly interact with VDAC in AD brains. This interaction was proposed to impair mitochondrial function likely through blocking mitochondrial pores [125]. Furthermore, mitochondrial stress was shown, in turn, to enhance hyperphosphorylation of tau in a mouse model lacking SOD2 [126]. Inhibition of mitochondrial complex I activity reduces ATP levels, resulting in a redistribution of tau from the axon to the soma and subsequent cell death [127]. Thus, these observations suggest that the toxic effects of tau on mitochondria could be reciprocal and that mitochondrial deficiency might play a critical role in the development of tau pathology.
Both Aβ and pathogenic tau have deleterious effects on mitochondrial dynamics through which impact mitochondrial function. Studies on postmortem brain tissues from human patients with AD and mouse models have demonstrated increased levels of Drp1 and Fis1 and reduced levels of mitofusin 1 (Mfn1), Mfn2, and OPA1. Moreover, Aβ overproduction, phospho-tau accumulation, as well as abnormal interactions of Drp1 with Aβ or phospho-tau cause excessive mitochondrial fission and fragmentation, which tend to increase as AD progresses [91,97,108,125,128]. Cells overexpressing mutant tau associated with frontotemporal dementia (FTD) with Parkinsonism linked to chromosome 17 (FTDP-17) display decreased rates of mitochondrial fusion and fission and enhanced vulnerability to oxidative stress [121]. Strikingly, reduction of Drp1 expression can protect against mutated tau-induced mitochondrial dysfunction [129]. Collectively, these data suggest that the pathogenic forms of tau and Aβ could impair mitochondrial function either through direct interaction with VDAC, ABAD, or CypD, or indirectly through their toxic effects on mitochondrial dynamics.
3.2. Mitochondrial Defects with Synucleinopathies
PD is the second most common form of neurodegenerative disease, which is characterized by the aberrant accumulation of α-synuclein (α-syn) in the form of Lewy bodies, especially in the substantia nigra. α-syn is abundant throughout the central nervous system and Lewy bodies are a defining feature of many clinical phenotypes known as synucleinopathies [130,131]. Importantly, impaired mitochondrial function is also a pathological feature of both sporadic and familial PD [20,28,44,49,132,133,134]. The relationship between α-syn and mitochondria has been explored in many studies. Some evidence showed that mitochondria could be the main targets of α-syn. In particular, the oligomerization and aggregation of α-syn can cause deficits in the complex I activities, leading to reduced ATP levels, depolarized Δψm, and the release of cytochrome c into the cytosol to trigger apoptosis [135,136]. A number of studies have shown that α-syn is directly localized in mitochondria, and can be detected in isolated mitochondria from PD patient brains. Mitochondrial localization of α-syn has a negative impact on mitochondrial function, morphology, and dynamics [137,138,139,140,141,142,143]. α-syn has a cryptic mitochondrial targeting sequence located at its amino terminal region through which α-syn is constitutively imported into mitochondria and associates with the IMM. Such a mechanism leads to reduced complex I activities and elevated ROS levels in human dopaminergic neurons [140]. Moreover, oligomeric and dopamine-modified α-syn disrupts the association of the OMM translocase TOM20 and its coreceptor, TOM22, resulting in protein import impairment [144]. Thus, diminished import of mitochondrial proteins impairs mitochondrial function in nigrostriatal neurons, as reflected by deficient respiration, loss of Δψm, and enhanced production of ROS.
α-syn can also affect mitochondrial dynamics and mitophagy. Basically, α-syn is known to bind to the lipid membranes, especially the lipids of the ER membrane or the MAM through which ER interacts with mitochondria. Mutated α-syn decreases the ER-mitochondria contact or interaction, leading to MAM dysfunction and thus mitochondrial fragmentation [141]. Other studies into PD have demonstrated that α-syn causes mitochondrial fragmentation through either direct binding or as a result of increased Drp1 [142,145]. Cleavage of Opa1 was found in dopaminergic neurons with overexpression of α-syn, resulting in decreased mitochondrial fusion [145]. Consistently, suppression of Drp1-mediated mitochondrial fission was reported to protect cells from α-syn-induced cytotoxicity [146]. In addition, studies have demonstrated the direct binding of α-syn to cardiolipin [147,148]. Furthermore, PD-related SNCA-mutant neurons exhibit increased externalization of cardiolipin to the OMM. Externalized cardiolipin was shown to bind to and promote refolding α-syn fibrils. Importantly, the exposed cardiolipin initiates LC3 recruitment to mitochondria and thus enhances mitophagic turnover, leading to reduced mitochondrial volume and exacerbation of mutant α-syn-induced mitochondrial dysfunction [148]. On the other hand, mitophagy defects were also proposed to play a significant role in PD pathogenesis, especially augmenting α-syn accumulation and its mediated neurotoxicity [149,150,151].
3.3. ALS and FTD-Associated Mitochondrial Toxicity
ALS is a devastating disease characterized by motor neuron degeneration. A hallmark of ALS, as in the pathologies of other neurodegenerative diseases, is the abnormal accumulation of misfolded proteins and protein aggregates within the affected motor neurons. FTD affects the basal ganglia and cortical neurons, leading to cognitive deficits, language deficiency along with altered social behavior and conduct. Even though the affected neuron types are quite different, ALS and FTD show the similarities in genetic background and pathological processes and also share the common pathways of neurodegeneration [152,153]. Defects in oxidative phosphorylation, Ca2+ buffering, and increased ROS production have been linked to ALS pathogenesis [154]. Multiple studies in cell culture and in transgenic animal models of ALS reveal alterations in oxidative metabolism linked to changes in ETC activity and reduced ATP synthesis [155,156,157,158]. More importantly, mitochondria purified from ALS patients display impaired Ca2+ homeostasis and increased ROS levels. Such defects are coupled with oxidative damage including altered tyrosine nitration and protein carbonylation [159,160]. Indeed, glutamate-receptor mediated excitotoxicity was linked to overloaded mitochondrial Ca2+ and increased ROS levels in spinal motor neurons cultured from an ALS animal model [161].
Aggregation of the transactive response DNA-binding protein 43 kDa (TDP-43) and fused in sarcoma (FUS) is the pathological hallmarks of both ALS and FTD. Both TDP-43 and FUS are ribonuclear proteins and contain the glycine molecule-enriched prion-like domains that can increase the propensity of TDP-43 and FUS for aggregation as well as cell-to-cell transmission. Aged animals expressing mutant FUS exhibit abnormal accumulation of ubiquitin-positive aggregates, which correlates with neuron loss. These aggregates also stain positive for mitochondrial protein cytochrome c, suggesting that damaged mitochondria recruit the autophagy machinery for removal through mitophagy [162]. One study from a single postmortem analysis of an FUS mutation carrier uncovered similar defects. Additionally, C- and N-terminal fragments of TDP-43 were identified within mitochondria. Furthermore, animal models of TDP-43 pathology exhibit membranous organelle redistribution and clustering within cytoplasmic inclusions accompanied by morphological and ultrastructural alterations, as well as abnormal mitochondrial dynamics, trafficking, and quality control [163,164,165]. Thus, these data consistently indicate the phenotypes of mislocalized, fragmented, and defective mitochondria associated with ALS and FTD.
3.4. Mutant Htt-Induced Mitochondrial Damage
HD is a neurodegenerative genetic disease that affects muscle coordination and leads to cognitive decline and psychiatric symptoms [166]. This autosomal dominant inherited neurodegenerative disease is the most common genetic cause of abnormal involuntary movements called chorea, and is characterized by mutations in the huntingtin gene (HTT) that result in abnormal expansion of the cytosine–adenine–guanine (CAG) trinucleotide repeats in the HTT gene, encoding a polyglutamine (polyQ) tract at the N-terminal region of the huntingtin (Htt) protein. The N-terminus of Htt can be cleaved through protease activity, leading to formation of short and toxic polyQ peptides. The N-terminal fragments of Htt containing the polyQ tracts are more prone to aggregation and accumulate within the inclusions in the nucleus especially in the medium spiny neurons of the striatum [167,168,169,170,171]. Htt was reported to directly bind to Tim 23 on mitochondria, thus preventing the protein import into mitochondria. This defect could be reversed through overexpressing Tim 23 [172,173]. In addition, the aggregate accumulation can disrupt the ETC function [174]. Moreover, studies in HD patient brains found decreased activities of complex II, complex III, and complex IV. Reduced complex II activity was observed particularly in the striatum of HD patients. Such defects along with reduced ATP production were also demonstrated in other studies, which collectively point towards impaired OXPHOS and disrupted mitochondrial energy metabolism [175]. Importantly, overexpression of complex II reduces the mutant Htt-mediated toxic effects in striatal neurons. Moreover, alterations in mitochondrial dynamics were also reported in the striatum of HD patients, as well as in animal and cell models [176,177]. Such a defect is caused by abnormal interaction of mutant Htt with Drp1, leading to Drp1-enhanced mitochondrial fission and thus mitochondrial fragmentation as well as cellular dysregulation and death [178].
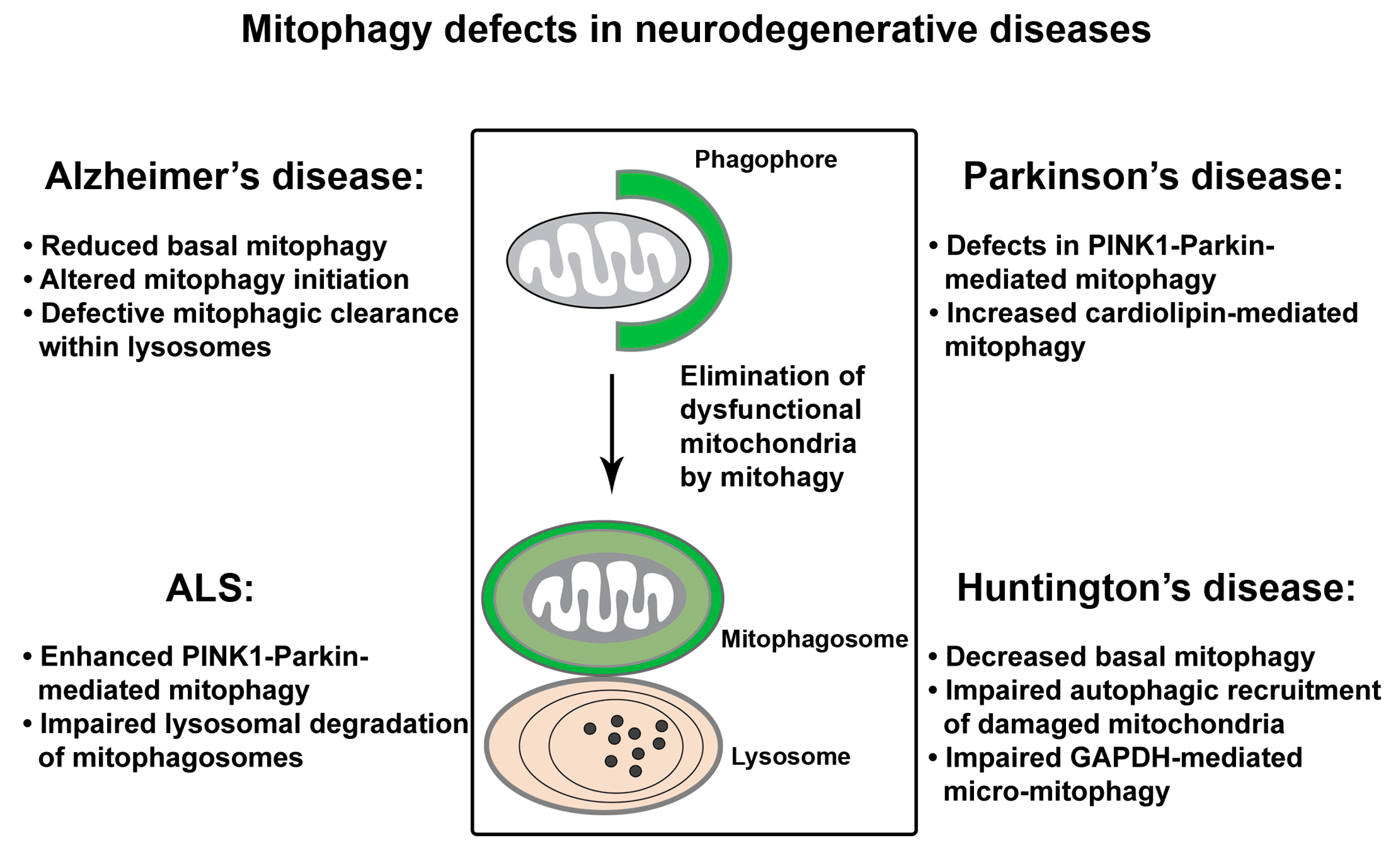
4. Mitophagy Defects in Neurodegenerative Diseases
Neurons have very high demand for ATP. Given that mitochondria are the major producer of ATP within cells, the nervous system is especially sensitive to mitochondrial damage. Inefficient elimination of injured mitochondria through mitophagy could be detrimental to neuronal health. Mitophagy deficit has been indicated in aging and the pathogenesis of age-associated neurodegenerative disorders. Studies into mitophagy suggest that defective mitophagy contributes to impaired mitochondrial function and neurodegeneration (Figure 4).
4.1. Mitophagy Defects in AD
Earlier studies revealed abnormal mitophagy in AD patient brains, as evidenced by autophagic accumulation of mitochondria in the soma of vulnerable AD neurons [87,179,180]. Among the multiple distinct mitophagy pathways, PINK1-Parkin-dependent mitophagy has been the focus of current studies in AD. We have shown that the Parkin pathway is robustly induced upon progressive Aβ accumulation and mitochondrial damage in human patient brains and animal models of AD [74]. Furthermore, cytosolic Parkin is depleted in AD brains over the disease’s progression, resulting in mitophagic pathology and augmented mitochondrial defects. Consistently, in the AD patient-derived skin fibroblasts and brain biopsies, another study reported diminished Parkin along with abnormal PINK1 accumulation [181]. Mitophagy can be restored in these cells by overexpression of Parkin, as reflected by decreased PINK1 and the recovery of Δψm coupled with reduced retention of defective mitochondria. Therefore, these findings indicate that impaired mitochondrial function and abnormal retention of dysfunctional mitochondria could be attributed to mitophagy defects in AD neurons. In addition, cardiolipin cluster-organized profile was shown to be lost in synaptic mitochondria purified from AD mouse models [182], occurring at the early disease stages and before nonsynaptic mitochondrial defects. This data suggests the possibility that cardiolipin-mediated mitophagy might be deficient in AD.
The degradation capacity of lysosomes is critical for mitophagic clearance, and defects in lysosomal proteolysis of autophagic cargoes can also impair the mitophagy function. Lysosomal deficit is a prominent feature in AD brains, linked to the pathogenesis of AD. Suppression of lysosomal proteolysis in wild-type (WT) mice was shown to mimic neuropathology of AD and exacerbate autophagic pathology and amyloidogenesis in AD mouse models [183,184]. Presenilin 1 mutations along with ApoE4, a key genetic risk factor of AD, are thought to disrupt lysosomal function [185]. Other factors, including Aβ peptides, phospho-tau, ROS, and oxidized lipids and lipoproteins, could also impair lysosomal proteolysis and result in a toxic accumulation, thus triggering apoptosis and neuronal death in AD. Our recent study proposes that AD-linked lysosomal deficit is also attributed to defects in protease targeting to lysosomes [186]. It is known that newly synthesized protease precursors need to be delivered from the trans-Golgi network (TGN) to the endo-lysosomal system for maturation, a process that relies on the presence of cation-independent mannose 6-phosphate receptor (CI-MPR) at the Golgi. The retromer complex mediates the retrieval of CI-MPR from late endosomes to the TGN and thus facilitates the trafficking of proteases to late endosomes and lysosomes [187]. Our study reveals that retromer dysfunction and defective CI-MPR recycling to the Golgi lead to defects in protease targeting to lysosomes. As a result, protease deficit within lysosomes impedes lysosomal proteolysis of defective mitochondria along with other autophagic cargoes in AD neurons [186]. Therefore, increased Parkin association with mitochondria, autophagic accumulation, as well as abnormal mitochondrial retention within lysosomes observed in AD neurons of patient brains and in cultured cells overexpressing mutant APP could also represent lysosomal deficiency [74,186]. Taken together, these observations indicate that defective mitophagy is likely involved in AD-linked neurodegeneration.
Pathogenic truncation of tau could impair mitophagy function. A recent work reported a stable association of an NH2-htau fragment with Parkin and Ubiquitin-C-terminal hydrolase L1 (UCHL-1) in cellular and animal AD models and human AD brains, leading to enhanced mitochondrial recruitment of Parkin and UCHL-1 and thus improper mitochondrial turnover [188]. Mitophagy suppression can restore synaptic mitochondrial density and partially, but significantly, protect against neuronal death induced by this NH2-htau. In contrast, another study proposed that human wild type tau (htau) is inserted into the mitochondrial membrane, thus inducing mitophagy impairment [189]. However, in a more recent study, both htau and htauP301L were shown to impair mitophagy in Caenorhabditis elegans (C. elegans) and neuroblastoma cells by reducing Parkin translocation onto mitochondria through a different mechanism. Instead of changes in the Δψm or the cytoskeleton, impaired Parkin recruitment to mitochondria is proposed to be caused by tau-mediated sequestration of Parkin in the cytosol [190]. Collectively, these data suggest that mitophagy is impaired under tauopathy conditions by distinct mechanisms.
In addition to defects in mitophagic clearance in response to Aβ and tau-induced mitochondrial damage, a recent study reveals a marked decrease in the basal level of mitophagy in postmortem hippocampal tissues from AD patients, cortical neurons derived from AD-induced pluripotent stem cell (iPSC), as well as AD mouse models [191]. This study further demonstrates defects in the activation of ULK1 and TBK1, the autophagy proteins that mediate autophagy/mitophagy initiation, thus leading to impaired mitophagy function. Furthermore, pharmacological reinstallation of mitophagy mitigates amyloid and tau pathologies, resulting in beneficial effects against memory loss in these AD mice. Therefore, these data support the view that defective mitophagy is likely an early event in AD brains and plays a causative role in the development of AD-linked neuropathology [191]. Further studies using neurons derived from iPSCs of sporadic AD or other similar models could be very critical to addressing whether mitophagy dysfunction serves as a key player in Aβ/tau proteinopathies.
4.2. Mitophagy Defects in PD
Dysfunctional mitophagy is closely linked to PD. Many PD-causing genes show the mitochondrial phenotypes [28,137,150,192,193,194,195,196,197]. In addition, PD patients have increased rates of mtDNA deletion in the substantia nigra, which further associates defective mitochondrial quality control with PD [198]. The important role for mitophagy in PD was first indicated from an ultrastructural study showing autophagic accumulation of mitochondria in neurons of the patients with PD and Lewy Body Dementia (LBD) [83]. Many other studies have demonstrated mitophagy abnormalities in a variety of experimental models representing genetic forms of toxic-environmental PD [67,199,200,201,202]. As previously discussed (see Section 2.1), cell-based and mechanistic studies directly link PINK1 and Parkin to mitophagy. However, while loss-of-function mutations in PARK6 (encoding PINK1) and PARK2 (encoding Parkin) genes are linked to familial PD [203], the role of the PINK1-Parkin-dependent pathway in vivo remains elusive. The PINK1 and Parkin pathway has been extensively studied in Drosophila. Mutant flies show dopaminergic degeneration, reduced lifespan, and locomotive defects [28,44,204,205,206]. Muscle cells of mutant flies exhibited swollen mitochondria with disrupted cristae, coupled with muscle degeneration [206,207,208,209]. PARK6 KO rats showed dopaminergic loss and motor defects [210]. Importantly, both Drosophila and rat model systems show mitochondrial dysfunction. However, mice with the deletion of PARK6 or PARK2 do not exhibit robust substantial PD-relevant phenotypes [77,79,80,81]. This might be due to compensations by other mechanisms that are sufficient enough to maintain neuronal homeostasis under physiological conditions. Strikingly, when crossing PARK2 KO mice with Mutator mice characterized by accelerated acquisition of mtDNA mutations, the resulting phenotypes have mitochondrial defects as well as dopaminergic neuronal loss [211]. Thus, this observation suggests that Parkin-mutant mice are susceptible to increased mtDNA damage. Given that both impaired mitochondrial function and mitophagy deficit are the upstream of neurodegeneration, the lack of robust phenotypes in mice suggest that the PINK1-Parkin pathway might be dispensable under physiological conditions, yet still necessary in response to stress/pathological stimuli for the functional maintenance and survival of PD-related dopaminergic neurons. Aside from PINK1-Parkin-dependent mitophagy, increased cardiolipin-mediated mitophagy was proposed to play a role in α-syn-induced mitochondrial dysfunction [148].
4.3. Mitophagy Defects in ALS
Impaired mitophagy was proposed to be involved in the denervation of neuromuscular junctions in an ALS mouse model [212]. Additionally, lysosomal dysfunction has been implicated in ALS. A recent work has provided a strong evidence showing that lysosomal deficits play a critical role in autophagy/mitophagy dysfunction and mitochondrial pathology in a mutant SOD1 transgenic mouse model of ALS [213]. Lysosomal deficits result in abnormal accumulation of autophagic vacuoles (AVs) engulfing damaged mitochondria within motor neuron axons of mutant SOD1 mice. More importantly, rescuing autophagy-lysosomal deficits was shown to enhance mitochondrial turnover, improve motor neuron survival, and ameliorate disease phenotype in mutant SOD1 mice. Given that autophagy/mitophagy is a lysosome-dependent pathway, defective mitophagy and mitochondrial pathology in ALS are attributed to defects in lysosomal proteolysis.
A more recent study uncovers that Parkin-dependent mitophagy is activated in the mutant SOD1 mouse model of ALS [214]. Mitophagy activation is known to induce Parkin-triggered and the UPS-mediated degradation of mitochondrial dynamics proteins Mfn2 and Mitochondrial Rho GTPase (Miro1) [42,215,216,217,218]. Consistently, increased mitophagy in the spinal cord of the mutant SOD1 mice is coupled with depletion of Parkin as well as mitochondrial dynamics proteins Mfn2 and Miro1 that are ubiquitinated by Parkin. Interestingly, genetic ablation of PARK2 protects against muscle denervation and motor neuron loss and attenuates the depletion of mitochondrial dynamics proteins, which delays disease progression and prolongs life span in mutant SOD1 mice. Thus, the results from this study suggest that Parkin could be a disease modifier of ALS, and chronic activation of Parkin-dependent mitophagy augments mitochondrial dysfunction by depleting mitochondrial dynamics proteins. Consistently, several other studies also reported a significant reduction of Miro1 in spinal cord tissue of ALS patients and animal models [219]. Moreover, it was shown that Miro1 reduction induced by ALS-linked mutant SOD is dependent on Parkin [220]. Miro1 is known as a component of the adaptor-motor complex essential for KIF5 motors to drive anterograde transport of mitochondria along axons [8]. Miro1-knockout mice exhibit upper motor neuron degeneration [221]. Thus, ALS-linked mitochondrial trafficking defect is likely caused by Miro1 deficiency as a result of Parkin-dependent enhancement of Miro1 turnover [220].
Compromised mitophagy may also induce ALS. Many of the genes linked to ALS encode proteins that play a critical role in autophagy/mitophagy, including OPTN and p62, as well as their kinase TBK1 [222,223,224]. However, it is poorly understood how the mutations in these genes are involved in the ALS pathology. Given the phosphorylation of OPTN and p62 by TBK1 to activate autophagy/mitophagy, aberrant accumulation of misfolded proteins and protein aggregates along with impaired mitochondrial turnover may both contribute to ALS-linked mitochondrial dysfunction and motor neuron death. Illuminating the role of these proteins in vivo will be critical in dissecting the molecular and cellular mechanisms leading to axonal degeneration and motor neuron loss.
4.4. Mitophagy Defects in HD
Mitochondrial dysfunction and autophagy failure have been linked to the pathogenesis of HD. Mutant Htt is known to be associated with mitochondria and to mediate mitochondrial damage. Defective mitophagy may also be involved in mitochondrial defects in HD. Decreased levels of the basal mitophagy were shown in the dentate gyrus of HD mice crossed with the mito-Keima mouse line [76]. In addition to its role in catalyzing the sixth step of glycolysis, a recent study proposed that GAPDH functions in micro-mitophagy—the direct engulfment of injured mitochondria by lysosomes [225]. In HD cell models, abnormal interaction of long polyQ tracts with mitochondrial GAPDH impairs GAPDH-mediated mitophagy, leading to mitochondrial dysfunction and increased cell death. Additionally, mutant Htt can interact with and affect the autophagy machinery [226]. A primary defect in the ability of autophagosomes to recognize and recruit cytosolic cargoes was reported in HD cells, leading to inefficient autophagic engulfment of substrates including mitochondria. Such a defect contributes to HD-associated accumulation of abnormal mitochondria [227]. Moreover, Htt was proposed to act as a scaffold protein for autophagy through the physical interaction of Htt with p62 and ULK1 proteins. This interaction allows Htt to facilitate p62-mediated cargo recognition efficiency, in particular, associating Lys-63-linked ubiquitin-modified substrates with LC3-II—the integral component of phagophore or isolation membranes. Thus, this study supports the possibility that polyQ expansion might compromise the role of Htt in autophagy [228]. Given the evidence for HD-linked autophagy impairment and mitophagic pathology, investigations into mitophagy status as well as detailed mechanisms are important for better understanding of HD pathogenesis.
5. Mitophagy-Targeted Therapeutic Interventions
From the above stated, it is clear that mitochondrial damage is a hallmark of major neurodegenerative diseases. Pharmacological agents that induce mitophagy with a goal of enhancing clearance of damaged mitochondria could be a promising strategy for achieving a significant therapeutic benefit [7,98,229,230]. Several mitophagy inducers, including NAD+ precursors, urolithin A (UA), the antibiotic actinonin (AC), and spermidine [231,232], have been examined and shown significant benefits in enhancing mitophagy, increasing mitochondrial resistance to oxidative stress, prolonging health span, and for neuronal protection in disease animal models and/or human cells. The levels of NAD+ are reduced in AD animal models, and elevation of cellular NAD+ levels through supplementation with NAD+ precursors such as nicotinamide, nicotinamide mononucleotide (NMN), and nicotinamide riboside (NR) is found to attenuate Aβ and tau pathologies and protect against cognitive dysfunction [233]. Such beneficial effects are attributed to the enhancement of the NAD+-dependent SIRT1 and SIRT3, expression of the transcription factor CREB, and enhanced activities of PI3K-Akt and MAPK/ERK1/2 [191,233,234,235,236]. Additionally, NAD+ replenishment was also shown to restore mitochondrial function and thus ameliorate dopaminergic neuron loss in iPSC and Drosophila models of PD [237]. These observations collectively indicate that the interventions to sustain NAD+ levels might be beneficial for AD and PD patients.
UA is an ellagitannins-derived metabolite, and can effectively induce neuronal mitophagy in both C. elegans and mouse brains [191]. Both UA and AC-mediated mitophagy activation is dependent on PINK1, Parkin, and NIX, and was shown to attenuate AD pathologies, inflammation, and learning and memory deficits [191]. Polyamines, including spermidine, can increase autophagy activity through affecting autophagy-related gene expression as well as enhance mitophagy through the mechanisms of mammalian target of rapamycin (mTOR) inhibition and 5′ AMP-activated protein kinase (AMPK) activation [232,238]. Moreover, spermidine was shown to activate the Ataxia-Telangiesctasia mutated (ATM)-dependent PINK1/Parkin signaling. Treatment with spermidine can lead to memory improvement and prolonged life span observed in C. elegans, Drosophila, and mice [231,239].
Other pharmacological strategies to enhance mitophagy through inducing mild bioenergetic stress or inhibiting mTOR activity have also been proven to be beneficial in either delaying or treating AD. Mitochondrial uncoupling agents such as 2,4-dinitrophenol (DNP) were reported to stimulate autophagy and preserve neuronal function in AD animal models [240]. Through inducing mild bioenergetic stress and stimulating ketogenesis, 2-deoxyglucose treatment was found to protect neurons against degeneration in a mitochondrial toxin-based PD model [241], as well as enhance mitochondrial function and stimulate autophagic clearance of Aβ [242]. The mTOR inhibitor rapamycin-mediated enhancement of autophagy/mitophagy and AMPK activation can induce mitochondrial clearance in a number of model organisms, including C. elegans, Drosophila, and mice [243,244]. Rapamycin was also shown to reduce Aβ pathology and ameliorate cognitive dysfunction in a mutant APP transgenic mouse model [245]. Similar to rapamycin, metformin can also stimulate mitophagy through inhibitions of mTOR and complex I activities and activations of AMPK, SIRT1, and Parkin-dependent mitophagy [246,247,248]. Therefore, these observations collectively provide a strong rationale for future research into the compounds that can enhance mitophagy in AD models, such as UA, AC, and spermidine [76,249,250].
In addition, mitophagy enhancement through activation of Parkin could be another promising strategy in some disease models. Nilotinib was originally identified as a tyrosine kinase inhibitor, and was recently reported to increase Parkin abundance and ubiquitination potentially through enhancing Parkin recycling via the proteasome system [251]. Nilotinib-mediated c-ABL inhibition can also prevent tyrosine phosphorylation of Parkin, leading to the release of Parkin auto-inhibition status. Such a mechanism was demonstrated to be protective in PD models [252]. Moreover, chronic treatment with nilotinib in APP transgenic mice can enhance Aβ clearance through increasing the interaction of Parkin with Beclin 1 [253]. As for ALS and FTD, nilotinib treatment was also reported to mitigate motor and cognitive deficits in TDP-43 transgenic mice [254].
6. Concluding Remarks
Mitochondrial health is vital for cellular and organismal homeostasis, and mitochondrial defects have long been linked to the pathogenesis of neurodegenerative diseases such as AD, PD, ALS, HD, and others. However, it is still unclear whether cellular mechanisms required for the maintenance of mitochondrial integrity and function are deficient in these diseases, thus exacerbating mitochondrial pathology. The quality control of mitochondria involves multiple levels of strategies to protect against mitochondrial damage and maintain a healthy mitochondrial population within cells. In neurons, mitophagy serves as a major pathway of the quality control mechanisms for the removal of aged and defective mitochondria through lysosomal proteolysis. The molecular and cellular mechanisms that govern mitophagy have been extensively studied in the past decade. However, mitophagy deficit has only been recognized recently as a key player involved in aging and neurodegeneration. Given the fact that mitochondrial deficit is clearly linked to neuronal dysfunction and the exacerbation of disease defects, protection of mitochondrial function could be a practical strategy to promote neuroprotection and modify disease pathology. Mitochondrially targeted antioxidants have been proposed. In particular, the antioxidant MitoQ, a redox active ubiquinone targeted to mitochondria, has been examined and demonstrated to have positive effects in multiple models of aging and neurodegenerative disorders [255,256,257,258]. Importantly, mitophagy could be another promising target for drug discovery strategy. Therefore, further detailed studies to elucidate mitophagy mechanisms not only advance our understanding of the mitochondrial phenotypes and disease pathogenesis, but also suggest potential therapeutic strategies to combat neurodegenerative diseases.
Author Contributions
Q.C. conceived ideas and wrote the manuscript. Y.Y.J. contributed to some sections related with AD. Q.C. is responsible for funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Institutes of Health (NIH) grants NS089737 and NS102780.
Acknowledgments
The authors thank L. Turkalj and J. Cheung for editing and other members of the Cai laboratory for their assistance and discussion. In addition to the cited references in the manuscript, we apologize for not being able to include and discuss other relevant studies due to the space limitation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sheng, Z.H.; Cai, Q. Mitochondrial transport in neurons: Impact on synaptic homeostasis and neurodegeneration. Nat. Rev. Neurosci. 2012, 13, 77–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Tammineni, P. Alterations in Mitochondrial Quality Control in Alzheimer’s Disease. Front. Cell Neurosci. 2016, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Tammineni, P. Mitochondrial Aspects of Synaptic Dysfunction in Alzheimer’s Disease. J. Alzheimers Dis. 2017, 57, 1087–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberlain, K.A.; Sheng, Z.H. Mechanisms for the maintenance and regulation of axonal energy supply. J. Neurosci. Res. 2019, 97, 897–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, D.G.; Budd, S.L. Mitochondria and neuronal survival. Physiol. Rev. 2000, 80, 315–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.; Chan, D.C. Mitochondrial dynamics and inheritance during cell division, development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 634–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, G.; Palikaras, K.; Lautrup, S.; Scheibye-Knudsen, M.; Tavernarakis, N.; Fang, E.F. Mitophagy and Neuroprotection. Trends Mol. Med. 2019. [Google Scholar] [CrossRef]
- Devine, M.J.; Kittler, J.T. Mitochondria at the neuronal presynapse in health and disease. Nat. Rev. Neurosci. 2018, 19, 63–80. [Google Scholar] [CrossRef]
- Werth, J.L.; Thayer, S.A. Mitochondria buffer physiological calcium loads in cultured rat dorsal root ganglion neurons. J. Neurosci. 1994, 14, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Zucker, R.S. Mitochondrial involvement in post-tetanic potentiation of synaptic transmission. Neuron 1997, 18, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Billups, B.; Forsythe, I.D. Presynaptic mitochondrial calcium sequestration influences transmission at mammalian central synapses. J. Neurosci. 2002, 22, 5840–5847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, G.; Barrett, E.F. Mitochondrial Ca2+ uptake prevents desynchronization of quantal release and minimizes depletion during repetitive stimulation of mouse motor nerve terminals. J. Physiol. 2003, 548, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Faas, G.C.; Saggau, P.; Craigen, W.J.; Sweatt, J.D. Mitochondrial regulation of synaptic plasticity in the hippocampus. J. Biol. Chem. 2003, 278, 17727–17734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.S.; Tian, J.H.; Pan, P.Y.; Zald, P.; Li, C.; Deng, C.; Sheng, Z.H. Docking of axonal mitochondria by syntaphilin controls their mobility and affects short-term facilitation. Cell 2008, 132, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.C. A mitochondrial bioenergetic etiology of disease. J. Clin. Investig. 2013, 123, 1405–1412. [Google Scholar] [CrossRef]
- Wallace, D.C. Mitochondrial diseases in man and mouse. Science 1999, 283, 1482–1488. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Court, F.A.; Coleman, M.P. Mitochondria as a central sensor for axonal degenerative stimuli. Trends Neurosci. 2012, 35, 364–372. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef]
- Bose, A.; Beal, M.F. Mitochondrial dysfunction in Parkinson’s disease. J. Neurochem. 2016, 139 (Suppl. 1), 216–231. [Google Scholar] [CrossRef]
- Fivenson, E.M.; Lautrup, S.; Sun, N.; Scheibye-Knudsen, M.; Stevnsner, T.; Nilsen, H.; Bohr, V.A.; Fang, E.F. Mitophagy in neurodegeneration and aging. Neurochem. Int. 2017, 109, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Gross, N.J.; Getz, G.S.; Rabinowitz, M. Apparent turnover of mitochondrial deoxyribonucleic acid and mitochondrial phospholipids in the tissues of the rat. J. Biol. Chem. 1969, 244, 1552–1562. [Google Scholar] [PubMed]
- Menzies, R.A.; Gold, P.H. The turnover of mitochondria in a variety of tissues of young adult and aged rats. J. Biol. Chem. 1971, 246, 2425–2429. [Google Scholar] [PubMed]
- Youle, R.J.; Narendra, D.P. Mechanisms of mitophagy. Nat. Rev. Mol. Cell Biol. 2011, 12, 9–14. [Google Scholar] [CrossRef]
- Pickles, S.; Vigie, P.; Youle, R.J. Mitophagy and Quality Control Mechanisms in Mitochondrial Maintenance. Curr. Biol. 2018, 28, R170–R185. [Google Scholar] [CrossRef]
- Chu, C.T. Mechanisms of selective autophagy and mitophagy: Implications for neurodegenerative diseases. Neurobiol. Dis. 2019, 122, 23–34. [Google Scholar] [CrossRef]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Mechanisms of mitophagy in cellular homeostasis, physiology and pathology. Nat. Cell Biol. 2018, 20, 1013–1022. [Google Scholar] [CrossRef]
- Clark, I.E.; Dodson, M.W.; Jiang, C.; Cao, J.H.; Huh, J.R.; Seol, J.H.; Yoo, S.J.; Hay, B.A.; Guo, M. Drosophila pink1 is required for mitochondrial function and interacts genetically with parkin. Nature 2006, 441, 1162–1166. [Google Scholar] [CrossRef]
- Gautier, C.A.; Kitada, T.; Shen, J. Loss of PINK1 causes mitochondrial functional defects and increased sensitivity to oxidative stress. Proc. Natl. Acad. Sci. USA 2008, 105, 11364–11369. [Google Scholar] [CrossRef] [Green Version]
- Narendra, D.; Tanaka, A.; Suen, D.F.; Youle, R.J. Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J. Cell. Biol. 2008, 183, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Kawajiri, S.; Saiki, S.; Sato, S.; Sato, F.; Hatano, T.; Eguchi, H.; Hattori, N. PINK1 is recruited to mitochondria with parkin and associates with LC3 in mitophagy. FEBS Lett. 2010, 584, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, N.; Sato, S.; Shiba, K.; Okatsu, K.; Saisho, K.; Gautier, C.A.; Sou, Y.S.; Saiki, S.; Kawajiri, S.; Sato, F.; et al. PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J. Cell Biol. 2010, 189, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Narendra, D.P.; Jin, S.M.; Tanaka, A.; Suen, D.F.; Gautier, C.A.; Shen, J.; Cookson, M.R.; Youle, R.J. PINK1 is selectively stabilized on impaired mitochondria to activate Parkin. PLoS Biol. 2010, 8, e1000298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba-Fukushima, K.; Imai, Y.; Yoshida, S.; Ishihama, Y.; Kanao, T.; Sato, S.; Hattori, N. PINK1-mediated phosphorylation of the Parkin ubiquitin-like domain primes mitochondrial translocation of Parkin and regulates mitophagy. Sci. Rep. 2012, 2, 1002. [Google Scholar] [CrossRef] [PubMed]
- Kane, L.A.; Lazarou, M.; Fogel, A.I.; Li, Y.; Yamano, K.; Sarraf, S.A.; Banerjee, S.; Youle, R.J. PINK1 phosphorylates ubiquitin to activate Parkin E3 ubiquitin ligase activity. J. Cell Biol. 2014, 205, 143–153. [Google Scholar] [CrossRef]
- Kazlauskaite, A.; Kondapalli, C.; Gourlay, R.; Campbell, D.G.; Ritorto, M.S.; Hofmann, K.; Alessi, D.R.; Knebel, A.; Trost, M.; Muqit, M.M. Parkin is activated by PINK1-dependent phosphorylation of ubiquitin at Ser65. Biochem. J. 2014, 460, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Kazlauskaite, A.; Kelly, V.; Johnson, C.; Baillie, C.; Hastie, C.J.; Peggie, M.; Macartney, T.; Woodroof, H.I.; Alessi, D.R.; Pedrioli, P.G.; et al. Phosphorylation of Parkin at Serine65 is essential for activation: Elaboration of a Miro1 substrate-based assay of Parkin E3 ligase activity. Open Biol. 2014, 4, 130213. [Google Scholar] [CrossRef] [Green Version]
- Koyano, F.; Okatsu, K.; Kosako, H.; Tamura, Y.; Go, E.; Kimura, M.; Kimura, Y.; Tsuchiya, H.; Yoshihara, H.; Hirokawa, T.; et al. Ubiquitin is phosphorylated by PINK1 to activate parkin. Nature 2014, 510, 162–166. [Google Scholar] [CrossRef]
- Geisler, S.; Holmstrom, K.M.; Skujat, D.; Fiesel, F.C.; Rothfuss, O.C.; Kahle, P.J.; Springer, W. PINK1/Parkin-mediated mitophagy is dependent on VDAC1 and p62/SQSTM1. Nat. Cell Biol. 2010, 12, 119–131. [Google Scholar] [CrossRef]
- Poole, A.C.; Thomas, R.E.; Yu, S.; Vincow, E.S.; Pallanck, L. The mitochondrial fusion-promoting factor mitofusin is a substrate of the PINK1/parkin pathway. PLoS ONE 2010, 5, e10054. [Google Scholar] [CrossRef] [Green Version]
- Ziviani, E.; Tao, R.N.; Whitworth, A.J. Drosophila parkin requires PINK1 for mitochondrial translocation and ubiquitinates mitofusin. Proc. Natl. Acad. Sci. USA 2010, 107, 5018–5023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, N.C.; Salazar, A.M.; Pham, A.H.; Sweredoski, M.J.; Kolawa, N.J.; Graham, R.L.; Hess, S.; Chan, D.C. Broad activation of the ubiquitin-proteasome system by Parkin is critical for mitophagy. Hum. Mol. Genet. 2011, 20, 1726–1737. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, S.R.; Kishi, C.; Ishihara, N.; Mizushima, N. Parkin mediates proteasome-dependent protein degradation and rupture of the outer mitochondrial membrane. J. Biol. Chem. 2011, 286, 19630–19640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lee, S.B.; Lee, S.; Kim, Y.; Song, S.; Kim, S.; Bae, E.; Kim, J.; Shong, M.; Kim, J.M.; et al. Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nature 2006, 441, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Vincow, E.S.; Merrihew, G.; Thomas, R.E.; Shulman, N.J.; Beyer, R.P.; MacCoss, M.J.; Pallanck, L.J. The PINK1-Parkin pathway promotes both mitophagy and selective respiratory chain turnover in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 6400–6405. [Google Scholar] [CrossRef] [Green Version]
- Devireddy, S.; Liu, A.; Lampe, T.; Hollenbeck, P.J. The Organization of Mitochondrial Quality Control and Life Cycle in the Nervous System In Vivo in the Absence of PINK1. J. Neurosci. 2015, 35, 9391–9401. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Tandarich, L.C.; Nguyen, K.; Hollenbeck, P.J. Compartmentalized Regulation of Parkin-Mediated Mitochondrial Quality Control in the Drosophila Nervous System In Vivo. J. Neurosci. 2016, 36, 7375–7391. [Google Scholar] [CrossRef] [Green Version]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Sanchez-Martinez, A.; Zarate, A.M.; Beninca, C.; Mayor, U.; Clague, M.J.; Whitworth, A.J. Basal mitophagy is widespread in Drosophila but minimally affected by loss of Pink1 or parkin. J. Cell Biol. 2018, 217, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- McWilliams, T.G.; Barini, E.; Pohjolan-Pirhonen, R.; Brooks, S.P.; Singh, F.; Burel, S.; Balk, K.; Kumar, A.; Montava-Garriga, L.; Prescott, A.R.; et al. Phosphorylation of Parkin at serine 65 is essential for its activation in vivo. Open Biol. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa, E.; Marchetti, S.; Ricci, J.E. No Parkin Zone: Mitophagy without Parkin. Trends Cell Biol. 2018, 28, 882–895. [Google Scholar] [CrossRef] [PubMed]
- Braschi, E.; Zunino, R.; McBride, H.M. MAPL is a new mitochondrial SUMO E3 ligase that regulates mitochondrial fission. EMBO Rep. 2009, 10, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Puri, R.; Yang, H.; Lizzio, M.A.; Wu, C.; Sheng, Z.H.; Guo, M. MUL1 acts in parallel to the PINK1/parkin pathway in regulating mitofusin and compensates for loss of PINK1/parkin. Elife 2014, 3, e01958. [Google Scholar] [CrossRef] [PubMed]
- Ambivero, C.T.; Cilenti, L.; Main, S.; Zervos, A.S. Mulan E3 ubiquitin ligase interacts with multiple E2 conjugating enzymes and participates in mitophagy by recruiting GABARAP. Cell Signal. 2014, 26, 2921–2929. [Google Scholar] [CrossRef] [PubMed]
- Szargel, R.; Shani, V.; Abd Elghani, F.; Mekies, L.N.; Liani, E.; Rott, R.; Engelender, S. The PINK1, synphilin-1 and SIAH-1 complex constitutes a novel mitophagy pathway. Hum. Mol. Genet. 2016, 25, 3476–3490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, H.; Thiagarajan, P.; Dasgupta, S.K.; Schumacher, A.; Prchal, J.T.; Chen, M.; Wang, J. Essential role for Nix in autophagic maturation of erythroid cells. Nature 2008, 454, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Novak, I.; Kirkin, V.; McEwan, D.G.; Zhang, J.; Wild, P.; Rozenknop, A.; Rogov, V.; Lohr, F.; Popovic, D.; Occhipinti, A.; et al. Nix is a selective autophagy receptor for mitochondrial clearance. EMBO Rep. 2010, 11, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Sakakibara, K.; Chen, Q.; Okamoto, K. Receptor-mediated mitophagy in yeast and mammalian systems. Cell Res. 2014, 24, 787–795. [Google Scholar] [CrossRef] [Green Version]
- Koentjoro, B.; Park, J.S.; Sue, C.M. Nix restores mitophagy and mitochondrial function to protect against PINK1/Parkin-related Parkinson’s disease. Sci. Rep. 2017, 7, 44373. [Google Scholar] [CrossRef]
- Schweers, R.L.; Zhang, J.; Randall, M.S.; Loyd, M.R.; Li, W.; Dorsey, F.C.; Kundu, M.; Opferman, J.T.; Cleveland, J.L.; Miller, J.L.; et al. NIX is required for programmed mitochondrial clearance during reticulocyte maturation. Proc. Natl. Acad. Sci. USA 2007, 104, 19500–19505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, S.H.; Ou, L.C.; Gean, P.W.; Hung, J.J.; Chang, W.C. Selective inhibition of early—But not late—Expressed HIF-1alpha is neuroprotective in rats after focal ischemic brain damage. Brain Pathol. 2011, 21, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Feng, D.; Chen, G.; Chen, M.; Zheng, Q.; Song, P.; Ma, Q.; Zhu, C.; Wang, R.; Qi, W.; et al. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 2012, 14, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Han, Z.; Feng, D.; Chen, Y.; Chen, L.; Wu, H.; Huang, L.; Zhou, C.; Cai, X.; Fu, C.; et al. A regulatory signaling loop comprising the PGAM5 phosphatase and CK2 controls receptor-mediated mitophagy. Mol. Cell 2014, 54, 362–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, M.; Wang, C.; Li, F.; Peng, J.; Wen, B.; Gong, Q.; Shi, Y.; Tang, Y. Structural insights into the recognition of phosphorylated FUNDC1 by LC3B in mitophagy. Protein Cell 2017, 8, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhujabal, Z.; Birgisdottir, A.B.; Sjottem, E.; Brenne, H.B.; Overvatn, A.; Habisov, S.; Kirkin, V.; Lamark, T.; Johansen, T. FKBP8 recruits LC3A to mediate Parkin-independent mitophagy. EMBO Rep. 2017, 18, 947–961. [Google Scholar] [CrossRef]
- Chu, C.T.; Ji, J.; Dagda, R.K.; Jiang, J.F.; Tyurina, Y.Y.; Kapralov, A.A.; Tyurin, V.A.; Yanamala, N.; Shrivastava, I.H.; Mohammadyani, D.; et al. Cardiolipin externalization to the outer mitochondrial membrane acts as an elimination signal for mitophagy in neuronal cells. Nat. Cell Biol. 2013, 15, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Kagan, V.E.; Jiang, J.; Huang, Z.; Tyurina, Y.Y.; Desbourdes, C.; Cottet-Rousselle, C.; Dar, H.H.; Verma, M.; Tyurin, V.A.; Kapralov, A.A.; et al. NDPK-D (NM23-H4)-mediated externalization of cardiolipin enables elimination of depolarized mitochondria by mitophagy. Cell Death Differ. 2016, 23, 1140–1151. [Google Scholar] [CrossRef] [Green Version]
- Schlattner, U.; Tokarska-Schlattner, M.; Ramirez, S.; Tyurina, Y.Y.; Amoscato, A.A.; Mohammadyani, D.; Huang, Z.; Jiang, J.; Yanamala, N.; Seffouh, A.; et al. Dual function of mitochondrial Nm23-H4 protein in phosphotransfer and intermembrane lipid transfer: A cardiolipin-dependent switch. J. Biol. Chem. 2013, 288, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Ivatt, R.M.; Sanchez-Martinez, A.; Godena, V.K.; Brown, S.; Ziviani, E.; Whitworth, A.J. Genome-wide RNAi screen identifies the Parkinson disease GWAS risk locus SREBF1 as a regulator of mitophagy. Proc. Natl. Acad. Sci. USA 2014, 111, 8494–8499. [Google Scholar] [CrossRef] [Green Version]
- Sentelle, R.D.; Senkal, C.E.; Jiang, W.; Ponnusamy, S.; Gencer, S.; Selvam, S.P.; Ramshesh, V.K.; Peterson, Y.K.; Lemasters, J.J.; Szulc, Z.M.; et al. Ceramide targets autophagosomes to mitochondria and induces lethal mitophagy. Nat. Chem. Biol. 2012, 8, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Zakaria, H.M.; Simone, A.; Sheng, Z.H. Spatial parkin translocation and degradation of damaged mitochondria via mitophagy in live cortical neurons. Curr. Biol. 2012, 22, 545–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Zakaria, H.M.; Sheng, Z.H. Long time-lapse imaging reveals unique features of PARK2/Parkin-mediated mitophagy in mature cortical neurons. Autophagy 2012, 8, 976–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Sun, X.; Starovoytov, V.; Cai, Q. Parkin-mediated mitophagy in mutant hAPP neurons and Alzheimer’s disease patient brains. Hum. Mol. Genet. 2015, 24, 2938–2951. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, T.G.; Prescott, A.R.; Allen, G.F.; Tamjar, J.; Munson, M.J.; Thomson, C.; Muqit, M.M.; Ganley, I.G. mito-QC illuminates mitophagy and mitochondrial architecture in vivo. J. Cell Biol. 2016, 214, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Yun, J.; Liu, J.; Malide, D.; Liu, C.; Rovira, I.I.; Holmstrom, K.M.; Fergusson, M.M.; Yoo, Y.H.; Combs, C.A.; et al. Measuring In Vivo Mitophagy. Mol. Cell 2015, 60, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, M.S.; Fleming, S.M.; Palacino, J.J.; Cepeda, C.; Lam, H.A.; Bhatnagar, A.; Meloni, E.G.; Wu, N.; Ackerson, L.C.; Klapstein, G.J.; et al. Parkin-deficient mice exhibit nigrostriatal deficits but not loss of dopaminergic neurons. J. Biol. Chem. 2003, 278, 43628–43635. [Google Scholar] [CrossRef] [Green Version]
- Itier, J.M.; Ibanez, P.; Mena, M.A.; Abbas, N.; Cohen-Salmon, C.; Bohme, G.A.; Laville, M.; Pratt, J.; Corti, O.; Pradier, L.; et al. Parkin gene inactivation alters behaviour and dopamine neurotransmission in the mouse. Hum. Mol. Genet. 2003, 12, 2277–2291. [Google Scholar] [CrossRef]
- Perez, F.A.; Palmiter, R.D. Parkin-deficient mice are not a robust model of parkinsonism. Proc. Natl. Acad. Sci. USA 2005, 102, 2174–2179. [Google Scholar] [CrossRef] [Green Version]
- Akundi, R.S.; Huang, Z.; Eason, J.; Pandya, J.D.; Zhi, L.; Cass, W.A.; Sullivan, P.G.; Bueler, H. Increased mitochondrial calcium sensitivity and abnormal expression of innate immunity genes precede dopaminergic defects in Pink1-deficient mice. PLoS ONE 2011, 6, e16038. [Google Scholar] [CrossRef] [Green Version]
- Kitada, T.; Pisani, A.; Porter, D.R.; Yamaguchi, H.; Tscherter, A.; Martella, G.; Bonsi, P.; Zhang, C.; Pothos, E.N.; Shen, J. Impaired dopamine release and synaptic plasticity in the striatum of PINK1-deficient mice. Proc. Natl. Acad. Sci. USA 2007, 104, 11441–11446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiesel, F.C.; Ando, M.; Hudec, R.; Hill, A.R.; Castanedes-Casey, M.; Caulfield, T.R.; Moussaud-Lamodiere, E.L.; Stankowski, J.N.; Bauer, P.O.; Lorenzo-Betancor, O.; et al. (Patho-)physiological relevance of PINK1-dependent ubiquitin phosphorylation. EMBO Rep. 2015, 16, 1114–1130. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.H.; Guo, F.; Shelburne, J.; Watkins, S.; Chu, C.T. Localization of phosphorylated ERK/MAP kinases to mitochondria and autophagosomes in Lewy body diseases. Brain Pathol. 2003, 13, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.E.; Shi, Q. A mitocentric view of Alzheimer’s disease suggests multi-faceted treatments. J. Alzheimers Dis. 2010, 20 (Suppl. 2), S591–S607. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.A.; Perry, G.; Richey, P.L.; Sayre, L.M.; Anderson, V.E.; Beal, M.F.; Kowall, N. Oxidative damage in Alzheimer’s. Nature 1996, 382, 120–121. [Google Scholar] [CrossRef]
- Maurer, I.; Zierz, S.; Moller, H.J. A selective defect of cytochrome c oxidase is present in brain of Alzheimer disease patients. Neurobiol. Aging 2000, 21, 455–462. [Google Scholar] [CrossRef]
- Hirai, K.; Aliev, G.; Nunomura, A.; Fujioka, H.; Russell, R.L.; Atwood, C.S.; Johnson, A.B.; Kress, Y.; Vinters, H.V.; Tabaton, M.; et al. Mitochondrial abnormalities in Alzheimer’s disease. J. Neurosci. 2001, 21, 3017–3023. [Google Scholar] [CrossRef] [Green Version]
- Devi, L.; Prabhu, B.M.; Galati, D.F.; Avadhani, N.G.; Anandatheerthavarada, H.K. Accumulation of amyloid precursor protein in the mitochondrial import channels of human Alzheimer’s disease brain is associated with mitochondrial dysfunction. J. Neurosci. 2006, 26, 9057–9068. [Google Scholar] [CrossRef] [Green Version]
- Caspersen, C.; Wang, N.; Yao, J.; Sosunov, A.; Chen, X.; Lustbader, J.W.; Xu, H.W.; Stern, D.; McKhann, G.; Yan, S.D. Mitochondrial Abeta: A potential focal point for neuronal metabolic dysfunction in Alzheimer’s disease. FASEB J. 2005, 19, 2040–2041. [Google Scholar] [CrossRef]
- Manczak, M.; Park, B.S.; Jung, Y.; Reddy, P.H. Differential expression of oxidative phosphorylation genes in patients with Alzheimer’s disease: Implications for early mitochondrial dysfunction and oxidative damage. Neuromolecular Med. 2004, 5, 147–162. [Google Scholar] [CrossRef]
- Manczak, M.; Calkins, M.J.; Reddy, P.H. Impaired mitochondrial dynamics and abnormal interaction of amyloid beta with mitochondrial protein Drp1 in neurons from patients with Alzheimer’s disease: Implications for neuronal damage. Hum. Mol. Genet. 2011, 20, 2495–2509. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.D., Jr.; Filley, C.M.; Parks, J.K. Cytochrome oxidase deficiency in Alzheimer’s disease. Neurology 1990, 40, 1302–1303. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Drake, J.; Pocernich, C.; Castegna, A. Evidence of oxidative damage in Alzheimer’s disease brain: Central role for amyloid beta-peptide. Trends Mol. Med. 2001, 7, 548–554. [Google Scholar] [CrossRef]
- Lin, M.T.; Simon, D.K.; Ahn, C.H.; Kim, L.M.; Beal, M.F. High aggregate burden of somatic mtDNA point mutations in aging and Alzheimer’s disease brain. Hum. Mol. Genet. 2002, 11, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Coskun, P.E.; Beal, M.F.; Wallace, D.C. Alzheimer’s brains harbor somatic mtDNA control-region mutations that suppress mitochondrial transcription and replication. Proc. Natl. Acad. Sci. USA 2004, 101, 10726–10731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.H.; Beal, M.F. Are mitochondria critical in the pathogenesis of Alzheimer’s disease? Brain Res. Brain Res. Rev. 2005, 49, 618–632. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Reddy, T.P.; Manczak, M.; Calkins, M.J.; Shirendeb, U.; Mao, P. Dynamin-related protein 1 and mitochondrial fragmentation in neurodegenerative diseases. Brain Res. Rev. 2011, 67, 103–118. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.S.; Adriaanse, B.A.; Greig, N.H.; Mattson, M.P.; Cader, M.Z.; Bohr, V.A.; Fang, E.F. Mitophagy and Alzheimer’s Disease: Cellular and Molecular Mechanisms. Trends Neurosci. 2017, 40, 151–166. [Google Scholar] [CrossRef] [Green Version]
- Dragicevic, N.; Mamcarz, M.; Zhu, Y.; Buzzeo, R.; Tan, J.; Arendash, G.W.; Bradshaw, P.C. Mitochondrial amyloid-beta levels are associated with the extent of mitochondrial dysfunction in different brain regions and the degree of cognitive impairment in Alzheimer’s transgenic mice. J. Alzheimers Dis. 2010, 20 (Suppl. 2), S535–S550. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Partin, J.; Begley, J.G. Amyloid beta-peptide induces apoptosis-related events in synapses and dendrites. Brain Res. 1998, 807, 167–176. [Google Scholar] [CrossRef]
- Lustbader, J.W.; Cirilli, M.; Lin, C.; Xu, H.W.; Takuma, K.; Wang, N.; Caspersen, C.; Chen, X.; Pollak, S.; Chaney, M.; et al. ABAD directly links Abeta to mitochondrial toxicity in Alzheimer’s disease. Science 2004, 304, 448–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Guo, L.; Fang, F.; Chen, D.; Sosunov, A.A.; McKhann, G.M.; Yan, Y.; Wang, C.; Zhang, H.; Molkentin, J.D.; et al. Cyclophilin D deficiency attenuates mitochondrial and neuronal perturbation and ameliorates learning and memory in Alzheimer’s disease. Nat. Med. 2008, 14, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Manczak, M.; Mao, P.; Calkins, M.J.; Cornea, A.; Reddy, A.P.; Murphy, M.P.; Szeto, H.H.; Park, B.; Reddy, P.H. Mitochondria-targeted antioxidants protect against amyloid-beta toxicity in Alzheimer’s disease neurons. J. Alzheimers Dis. 2010, 20 (Suppl. 2), S609–S631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, J.N.; Pang, Z.; Geddes, J.W.; Begley, J.G.; Germeyer, A.; Waeg, G.; Mattson, M.P. Impairment of glucose and glutamate transport and induction of mitochondrial oxidative stress and dysfunction in synaptosomes by amyloid beta-peptide: Role of the lipid peroxidation product 4-hydroxynonenal. J. Neurochem. 1997, 69, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Abramov, A.Y.; Canevari, L.; Duchen, M.R. Beta-amyloid peptides induce mitochondrial dysfunction and oxidative stress in astrocytes and death of neurons through activation of NADPH oxidase. J. Neurosci. 2004, 24, 565–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manczak, M.; Anekonda, T.S.; Henson, E.; Park, B.S.; Quinn, J.; Reddy, P.H. Mitochondria are a direct site of A beta accumulation in Alzheimer’s disease neurons: Implications for free radical generation and oxidative damage in disease progression. Hum. Mol. Genet. 2006, 15, 1437–1449. [Google Scholar] [CrossRef]
- Wang, X.; Su, B.; Siedlak, S.L.; Moreira, P.I.; Fujioka, H.; Wang, Y.; Casadesus, G.; Zhu, X. Amyloid-beta overproduction causes abnormal mitochondrial dynamics via differential modulation of mitochondrial fission/fusion proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 19318–19323. [Google Scholar] [CrossRef] [Green Version]
- Manczak, M.; Reddy, P.H. Abnormal interaction between the mitochondrial fission protein Drp1 and hyperphosphorylated tau in Alzheimer’s disease neurons: Implications for mitochondrial dysfunction and neuronal damage. Hum. Mol. Genet. 2012, 21, 2538–2547. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.; Tiwari, P.; Xie, Z.; Zheng, J.Q. Acute impairment of mitochondrial trafficking by beta-amyloid peptides in hippocampal neurons. J. Neurosci. 2006, 26, 10480–10487. [Google Scholar] [CrossRef]
- Du, H.; Guo, L.; Yan, S.; Sosunov, A.A.; McKhann, G.M.; Yan, S.S. Early deficits in synaptic mitochondria in an Alzheimer’s disease mouse model. Proc. Natl. Acad. Sci. USA 2010, 107, 18670–18675. [Google Scholar] [CrossRef] [Green Version]
- Calkins, M.J.; Manczak, M.; Mao, P.; Shirendeb, U.; Reddy, P.H. Impaired mitochondrial biogenesis, defective axonal transport of mitochondria, abnormal mitochondrial dynamics and synaptic degeneration in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2011, 20, 4515–4529. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, D.; Suski, J.M.; Oules, B.; Debayle, D.; Gay, A.S.; Lacas-Gervais, S.; Bussiere, R.; Bauer, C.; Pinton, P.; Paterlini-Brechot, P.; et al. Localization and Processing of the Amyloid-beta Protein Precursor in Mitochondria-Associated Membranes. J. Alzheimers Dis. 2017, 55, 1549–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiner, B.; Hedskog, L.; Wiehager, B.; Ankarcrona, M. Amyloid-beta peptides are generated in mitochondria-associated endoplasmic reticulum membranes. J. Alzheimers Dis. 2015, 43, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Hansson Petersen, C.A.; Alikhani, N.; Behbahani, H.; Wiehager, B.; Pavlov, P.F.; Alafuzoff, I.; Leinonen, V.; Ito, A.; Winblad, B.; Glaser, E.; et al. The amyloid beta-peptide is imported into mitochondria via the TOM import machinery and localized to mitochondrial cristae. Proc. Natl. Acad. Sci. USA 2008, 105, 13145–13150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinho, C.M.; Teixeira, P.F.; Glaser, E. Mitochondrial import and degradation of amyloid-beta peptide. Biochim. Biophys. Acta 2014, 1837, 1069–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedskog, L.; Pinho, C.M.; Filadi, R.; Ronnback, A.; Hertwig, L.; Wiehager, B.; Larssen, P.; Gellhaar, S.; Sandebring, A.; Westerlund, M.; et al. Modulation of the endoplasmic reticulum-mitochondria interface in Alzheimer’s disease and related models. Proc. Natl. Acad. Sci. USA 2013, 110, 7916–7921. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Ghosh, P.; Wan, R.; Ouyang, X.; Cheng, H.; Mattson, M.P.; Cheng, A. Permeability transition pore-mediated mitochondrial superoxide flashes mediate an early inhibitory effect of amyloid beta1-42 on neural progenitor cell proliferation. Neurobiol. Aging 2014, 35, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Guan, J.; Borrelli, L.A.; Xu, J.; Serrano-Pozo, A.; Bacskai, B.J. Mitochondrial alterations near amyloid plaques in an Alzheimer’s disease mouse model. J. Neurosci. 2013, 33, 17042–17051. [Google Scholar] [CrossRef] [Green Version]
- Eckert, A.; Nisbet, R.; Grimm, A.; Gotz, J. March separate, strike together—Role of phosphorylated TAU in mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1842, 1258–1266. [Google Scholar] [CrossRef] [Green Version]
- Spires-Jones, T.L.; Hyman, B.T. The intersection of amyloid beta and tau at synapses in Alzheimer’s disease. Neuron 2014, 82, 756–771. [Google Scholar] [CrossRef] [Green Version]
- Schulz, K.L.; Eckert, A.; Rhein, V.; Mai, S.; Haase, W.; Reichert, A.S.; Jendrach, M.; Muller, W.E.; Leuner, K. A new link to mitochondrial impairment in tauopathies. Mol. Neurobiol. 2012, 46, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Dumont, M.; Stack, C.; Elipenahli, C.; Jainuddin, S.; Gerges, M.; Starkova, N.N.; Yang, L.; Starkov, A.A.; Beal, F. Behavioral deficit, oxidative stress, and mitochondrial dysfunction precede tau pathology in P301S transgenic mice. FASEB J. 2011, 25, 4063–4072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotz, J.; Chen, F.; van Dorpe, J.; Nitsch, R.M. Formation of neurofibrillary tangles in P301l tau transgenic mice induced by Abeta 42 fibrils. Science 2001, 293, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- David, D.C.; Hauptmann, S.; Scherping, I.; Schuessel, K.; Keil, U.; Rizzu, P.; Ravid, R.; Drose, S.; Brandt, U.; Muller, W.E.; et al. Proteomic and functional analyses reveal a mitochondrial dysfunction in P301L tau transgenic mice. J. Biol. Chem. 2005, 280, 23802–23814. [Google Scholar] [CrossRef] [Green Version]
- Manczak, M.; Reddy, P.H. Abnormal interaction of VDAC1 with amyloid beta and phosphorylated tau causes mitochondrial dysfunction in Alzheimer’s disease. Hum. Mol. Genet. 2012, 21, 5131–5146. [Google Scholar] [CrossRef]
- Melov, S.; Adlard, P.A.; Morten, K.; Johnson, F.; Golden, T.R.; Hinerfeld, D.; Schilling, B.; Mavros, C.; Masters, C.L.; Volitakis, I.; et al. Mitochondrial oxidative stress causes hyperphosphorylation of tau. PLoS ONE 2007, 2, e536. [Google Scholar] [CrossRef]
- Escobar-Khondiker, M.; Hollerhage, M.; Muriel, M.P.; Champy, P.; Bach, A.; Depienne, C.; Respondek, G.; Yamada, E.S.; Lannuzel, A.; Yagi, T.; et al. Annonacin, a natural mitochondrial complex I inhibitor, causes tau pathology in cultured neurons. J. Neurosci. 2007, 27, 7827–7837. [Google Scholar] [CrossRef]
- Kandimalla, R.; Reddy, P.H. Multiple faces of dynamin-related protein 1 and its role in Alzheimer’s disease pathogenesis. Biochim. Biophys. Acta 2016, 1862, 814–828. [Google Scholar] [CrossRef]
- Kandimalla, R.; Manczak, M.; Fry, D.; Suneetha, Y.; Sesaki, H.; Reddy, P.H. Reduced dynamin-related protein 1 protects against phosphorylated Tau-induced mitochondrial dysfunction and synaptic damage in Alzheimer’s disease. Hum. Mol. Genet. 2016, 25, 4881–4897. [Google Scholar] [CrossRef] [Green Version]
- Massano, J.; Bhatia, K.P. Clinical approach to Parkinson’s disease: Features, diagnosis, and principles of management. Cold Spring Harb Perspect Med. 2012, 2, a008870. [Google Scholar] [CrossRef]
- Jellinger, K.A. Basic mechanisms of neurodegeneration: A critical update. J. Cell Mol. Med. 2010, 14, 457–487. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Hoffer, A.; Hoffer, B.; Qi, X. Mitochondria: A Therapeutic Target for Parkinson’s Disease? Int. J. Mol. Sci. 2015, 16, 20704–20730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osellame, L.D.; Rahim, A.A.; Hargreaves, I.P.; Gegg, M.E.; Richard-Londt, A.; Brandner, S.; Waddington, S.N.; Schapira, A.H.; Duchen, M.R. Mitochondria and quality control defects in a mouse model of Gaucher disease—Links to Parkinson’s disease. Cell Metab. 2013, 17, 941–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, A.K.; Ludtmann, M.H.; Angelova, P.R.; Simcox, E.M.; Horrocks, M.H.; Klenerman, D.; Gandhi, S.; Turnbull, D.M.; Abramov, A.Y. Aggregated alpha-synuclein and complex I deficiency: Exploration of their relationship in differentiated neurons. Cell Death Dis. 2015, 6, e1820. [Google Scholar] [CrossRef]
- Luth, E.S.; Stavrovskaya, I.G.; Bartels, T.; Kristal, B.S.; Selkoe, D.J. Soluble, prefibrillar alpha-synuclein oligomers promote complex I-dependent, Ca2+-induced mitochondrial dysfunction. J. Biol. Chem. 2014, 289, 21490–21507. [Google Scholar] [CrossRef] [Green Version]
- Chinta, S.J.; Mallajosyula, J.K.; Rane, A.; Andersen, J.K. Mitochondrial alpha-synuclein accumulation impairs complex I function in dopaminergic neurons and results in increased mitophagy in vivo. Neurosci. Lett. 2010, 486, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Hsu, L.J.; Sagara, Y.; Arroyo, A.; Rockenstein, E.; Sisk, A.; Mallory, M.; Wong, J.; Takenouchi, T.; Hashimoto, M.; Masliah, E. alpha-synuclein promotes mitochondrial deficit and oxidative stress. Am. J. Pathol. 2000, 157, 401–410. [Google Scholar] [CrossRef]
- Martin, L.J.; Pan, Y.; Price, A.C.; Sterling, W.; Copeland, N.G.; Jenkins, N.A.; Price, D.L.; Lee, M.K. Parkinson’s disease alpha-synuclein transgenic mice develop neuronal mitochondrial degeneration and cell death. J. Neurosci. 2006, 26, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Devi, L.; Raghavendran, V.; Prabhu, B.M.; Avadhani, N.G.; Anandatheerthavarada, H.K. Mitochondrial import and accumulation of alpha-synuclein impair complex I in human dopaminergic neuronal cultures and Parkinson disease brain. J. Biol. Chem. 2008, 283, 9089–9100. [Google Scholar] [CrossRef] [Green Version]
- Guardia-Laguarta, C.; Area-Gomez, E.; Rub, C.; Liu, Y.; Magrane, J.; Becker, D.; Voos, W.; Schon, E.A.; Przedborski, S. alpha-Synuclein is localized to mitochondria-associated ER membranes. J. Neurosci. 2014, 34, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Nemani, V.M.; Azarbal, F.; Skibinski, G.; Levy, J.M.; Egami, K.; Munishkina, L.; Zhang, J.; Gardner, B.; Wakabayashi, J.; et al. Direct membrane association drives mitochondrial fission by the Parkinson disease-associated protein alpha-synuclein. J. Biol. Chem. 2011, 286, 20710–20726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo Devoto, V.M.; Falzone, T.L. Mitochondrial dynamics in Parkinson’s disease: A role for alpha-synuclein? Dis. Model. Mech. 2017, 10, 1075–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Maio, R.; Barrett, P.J.; Hoffman, E.K.; Barrett, C.W.; Zharikov, A.; Borah, A.; Hu, X.; McCoy, J.; Chu, C.T.; Burton, E.A.; et al. alpha-Synuclein binds to TOM20 and inhibits mitochondrial protein import in Parkinson’s disease. Sci. Transl. Med. 2016, 8, 342ra378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.H.; Alaimo, A.; Gorojod, R.M.; Porte Alcon, S.; Fuentes, F.; Coluccio Leskow, F.; Kotler, M.L. Drp-1 dependent mitochondrial fragmentation and protective autophagy in dopaminergic SH-SY5Y cells overexpressing alpha-synuclein. Mol. Cell Neurosci. 2018, 88, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Bido, S.; Soria, F.N.; Fan, R.Z.; Bezard, E.; Tieu, K. Mitochondrial division inhibitor-1 is neuroprotective in the A53T-alpha-synuclein rat model of Parkinson’s disease. Sci. Rep. 2017, 7, 7495. [Google Scholar] [CrossRef] [Green Version]
- Zigoneanu, I.G.; Yang, Y.J.; Krois, A.S.; Haque, E.; Pielak, G.J. Interaction of alpha-synuclein with vesicles that mimic mitochondrial membranes. Biochim. Biophys. Acta 2012, 1818, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Ryan, T.; Bamm, V.V.; Stykel, M.G.; Coackley, C.L.; Humphries, K.M.; Jamieson-Williams, R.; Ambasudhan, R.; Mosser, D.D.; Lipton, S.A.; Harauz, G.; et al. Cardiolipin exposure on the outer mitochondrial membrane modulates alpha-synuclein. Nat. Commun. 2018, 9, 817. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Xie, Z.; Turkson, S.; Zhuang, X. A53T human alpha-synuclein overexpression in transgenic mice induces pervasive mitochondria macroautophagy defects preceding dopamine neuron degeneration. J. Neurosci. 2015, 35, 890–905. [Google Scholar] [CrossRef]
- Chung, S.Y.; Kishinevsky, S.; Mazzulli, J.R.; Graziotto, J.; Mrejeru, A.; Mosharov, E.V.; Puspita, L.; Valiulahi, P.; Sulzer, D.; Milner, T.A.; et al. Parkin and PINK1 Patient iPSC-Derived Midbrain Dopamine Neurons Exhibit Mitochondrial Dysfunction and alpha-Synuclein Accumulation. Stem Cell Rep. 2016, 7, 664–677. [Google Scholar] [CrossRef] [Green Version]
- Gispert, S.; Brehm, N.; Weil, J.; Seidel, K.; Rub, U.; Kern, B.; Walter, M.; Roeper, J.; Auburger, G. Potentiation of neurotoxicity in double-mutant mice with Pink1 ablation and A53T-SNCA overexpression. Hum. Mol. Genet. 2015, 24, 1061–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, R.; Kapogiannis, D.; Huey, E.D.; Momeni, P. FTD and ALS: A tale of two diseases. Curr. Alzheimer Res. 2011, 8, 273–294. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, J.H.; Hyman, T.; Dikic, I. Common Molecular Pathways in Amyotrophic Lateral Sclerosis and Frontotemporal Dementia. Trends Mol. Med. 2016, 22, 769–783. [Google Scholar] [CrossRef]
- Muyderman, H.; Chen, T. Mitochondrial dysfunction in amyotrophic lateral sclerosis—A valid pharmacological target? Br. J. Pharmacol. 2014, 171, 2191–2205. [Google Scholar] [CrossRef] [PubMed]
- Keep, M.; Elmer, E.; Fong, K.S.; Csiszar, K. Intrathecal cyclosporin prolongs survival of late-stage ALS mice. Brain Res. 2001, 894, 327–331. [Google Scholar] [CrossRef]
- Mattiazzi, M.; D’Aurelio, M.; Gajewski, C.D.; Martushova, K.; Kiaei, M.; Beal, M.F.; Manfredi, G. Mutated human SOD1 causes dysfunction of oxidative phosphorylation in mitochondria of transgenic mice. J. Biol. Chem. 2002, 277, 29626–29633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzies, F.M.; Grierson, A.J.; Cookson, M.R.; Heath, P.R.; Tomkins, J.; Figlewicz, D.A.; Ince, P.G.; Shaw, P.J. Selective loss of neurofilament expression in Cu/Zn superoxide dismutase (SOD1) linked amyotrophic lateral sclerosis. J. Neurochem. 2002, 82, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- Knott, A.B.; Perkins, G.; Schwarzenbacher, R.; Bossy-Wetzel, E. Mitochondrial fragmentation in neurodegeneration. Nat. Rev. Neurosci. 2008, 9, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Beal, M.F.; Ferrante, R.J.; Browne, S.E.; Matthews, R.T.; Kowall, N.W.; Brown, R.H., Jr. Increased 3-nitrotyrosine in both sporadic and familial amyotrophic lateral sclerosis. Ann. Neurol. 1997, 42, 644–654. [Google Scholar] [CrossRef]
- Beal, M.F. Coenzyme Q10 as a possible treatment for neurodegenerative diseases. Free Radic. Res. 2002, 36, 455–460. [Google Scholar] [CrossRef]
- Carriedo, S.G.; Sensi, S.L.; Yin, H.Z.; Weiss, J.H. AMPA exposures induce mitochondrial Ca(2+) overload and ROS generation in spinal motor neurons in vitro. J. Neurosci. 2000, 20, 240–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Zhou, H.; Tong, J.; Chen, H.; Liu, Y.J.; Wang, D.; Wei, X.; Xia, X.G. FUS transgenic rats develop the phenotypes of amyotrophic lateral sclerosis and frontotemporal lobar degeneration. PLoS Genet. 2011, 7, e1002011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, X.; Chiang, P.M.; Price, D.L.; Wong, P.C. Altered distributions of Gemini of coiled bodies and mitochondria in motor neurons of TDP-43 transgenic mice. Proc. Natl. Acad. Sci. USA 2010, 107, 16325–16330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.F.; Gendron, T.F.; Zhang, Y.J.; Lin, W.L.; D’Alton, S.; Sheng, H.; Casey, M.C.; Tong, J.; Knight, J.; Yu, X.; et al. Wild-type human TDP-43 expression causes TDP-43 phosphorylation, mitochondrial aggregation, motor deficits, and early mortality in transgenic mice. J. Neurosci. 2010, 30, 10851–10859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, S.A.; Itaman, S.; Khalid-Janney, C.M.; Sherard, J.A.; Dowell, J.A.; Cairns, N.J.; Gitcho, M.A. TDP-43 interacts with mitochondrial proteins critical for mitophagy and mitochondrial dynamics. Neurosci. Lett. 2018, 678, 8–15. [Google Scholar] [CrossRef] [PubMed]
- McColgan, P.; Tabrizi, S.J. Huntington’s disease: A clinical review. Eur. J. Neurol. 2018, 25, 24–34. [Google Scholar] [CrossRef]
- Feng, X.; Luo, S.; Lu, B. Conformation Polymorphism of Polyglutamine Proteins. Trends Biochem. Sci. 2018, 43, 424–435. [Google Scholar] [CrossRef]
- Lu, B.; Palacino, J. A novel human embryonic stem cell-derived Huntington’s disease neuronal model exhibits mutant huntingtin (mHTT) aggregates and soluble mHTT-dependent neurodegeneration. FASEB J. 2013, 27, 1820–1829. [Google Scholar] [CrossRef] [Green Version]
- Nagai, Y.; Inui, T.; Popiel, H.A.; Fujikake, N.; Hasegawa, K.; Urade, Y.; Goto, Y.; Naiki, H.; Toda, T. A toxic monomeric conformer of the polyglutamine protein. Nat. Struct. Mol. Biol. 2007, 14, 332–340. [Google Scholar] [CrossRef]
- Miller, J.; Arrasate, M.; Shaby, B.A.; Mitra, S.; Masliah, E.; Finkbeiner, S. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington’s disease molecular pathogenesis. J. Neurosci. 2010, 30, 10541–10550. [Google Scholar] [CrossRef] [Green Version]
- Arrasate, M.; Mitra, S.; Schweitzer, E.S.; Segal, M.R.; Finkbeiner, S. Inclusion body formation reduces levels of mutant huntingtin and the risk of neuronal death. Nature 2004, 431, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, H.; Baranov, S.V.; Baranova, O.V.; Kim, J.; Pan, Y.; Yablonska, S.; Carlisle, D.L.; Ferrante, R.J.; Kim, A.H.; Friedlander, R.M. Inhibition of mitochondrial protein import by mutant huntingtin. Nat. Neurosci. 2014, 17, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Orr, A.L.; Li, S.; Wang, C.E.; Li, H.; Wang, J.; Rong, J.; Xu, X.; Mastroberardino, P.G.; Greenamyre, J.T.; Li, X.J. N-terminal mutant huntingtin associates with mitochondria and impairs mitochondrial trafficking. J. Neurosci. 2008, 28, 2783–2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiano, M.; Diguet, E.; Malgorn, C.; D’Aurelio, M.; Galvan, L.; Petit, F.; Benhaim, L.; Guillermier, M.; Houitte, D.; Dufour, N.; et al. A role of mitochondrial complex II defects in genetic models of Huntington’s disease expressing N-terminal fragments of mutant huntingtin. Hum. Mol. Genet. 2013, 22, 3869–3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Iborra, S.; Vila, M.; Perier, C. Mitochondrial Quality Control in Neurodegenerative Diseases: Focus on Parkinson’s Disease and Huntington’s Disease. Front. Neurosci. 2018, 12, 342. [Google Scholar] [CrossRef] [Green Version]
- Shirendeb, U.; Reddy, A.P.; Manczak, M.; Calkins, M.J.; Mao, P.; Tagle, D.A.; Reddy, P.H. Abnormal mitochondrial dynamics, mitochondrial loss and mutant huntingtin oligomers in Huntington’s disease: Implications for selective neuronal damage. Hum. Mol. Genet. 2011, 20, 1438–1455. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Chen, J.; Petrilli, A.; Liot, G.; Klinglmayr, E.; Zhou, Y.; Poquiz, P.; Tjong, J.; Pouladi, M.A.; Hayden, M.R.; et al. Mutant huntingtin binds the mitochondrial fission GTPase dynamin-related protein-1 and increases its enzymatic activity. Nat. Med. 2011, 17, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Oliver, D.; Reddy, P.H. Dynamics of Dynamin-Related Protein 1 in Alzheimer’s Disease and Other Neurodegenerative Diseases. Cells 2019, 8, 961. [Google Scholar] [CrossRef] [Green Version]
- Moreira, P.I.; Siedlak, S.L.; Wang, X.; Santos, M.S.; Oliveira, C.R.; Tabaton, M.; Nunomura, A.; Szweda, L.I.; Aliev, G.; Smith, M.A.; et al. Increased autophagic degradation of mitochondria in Alzheimer disease. Autophagy 2007, 3, 614–615. [Google Scholar] [CrossRef] [Green Version]
- Moreira, P.I.; Siedlak, S.L.; Wang, X.; Santos, M.S.; Oliveira, C.R.; Tabaton, M.; Nunomura, A.; Szweda, L.I.; Aliev, G.; Smith, M.A.; et al. Autophagocytosis of mitochondria is prominent in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2007, 66, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Martin-Maestro, P.; Gargini, R.; Perry, G.; Avila, J.; Garcia-Escudero, V. PARK2 enhancement is able to compensate mitophagy alterations found in sporadic Alzheimer’s disease. Hum. Mol. 2016, 25, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro-Cardoso, V.F.; Oliveira, M.M.; Melo, T.; Domingues, M.R.; Moreira, P.I.; Ferreiro, E.; Peixoto, F.; Videira, R.A. Cardiolipin profile changes are associated to the early synaptic mitochondrial dysfunction in Alzheimer’s disease. J. Alzheimers Dis. 2015, 43, 1375–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Sato, Y.; Nixon, R.A. Lysosomal proteolysis inhibition selectively disrupts axonal transport of degradative organelles and causes an Alzheimer’s-like axonal dystrophy. J. Neurosci. 2011, 31, 7817–7830. [Google Scholar] [CrossRef] [PubMed]
- Nixon, R.A.; Yang, D.S. Autophagy failure in Alzheimer’s disease—Locating the primary defect. Neurobiol. Dis. 2011, 43, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nixon, R.A. The role of autophagy in neurodegenerative disease. Nat. Med. 2013, 19, 983–997. [Google Scholar] [CrossRef]
- Tammineni, P.; Jeong, Y.Y.; Feng, T.; Aikal, D.; Cai, Q. Impaired axonal retrograde trafficking of the retromer complex augments lysosomal deficits in Alzheimer’s disease neurons. Hum. Mol. Genet. 2017, 26, 4352–4366. [Google Scholar] [CrossRef]
- Small, S.A.; Petsko, G.A. Retromer in Alzheimer disease, Parkinson disease and other neurological disorders. Nat. Rev. Neurosci. 2015, 16, 126–132. [Google Scholar] [CrossRef]
- Corsetti, V.; Florenzano, F.; Atlante, A.; Bobba, A.; Ciotti, M.T.; Natale, F.; Della Valle, F.; Borreca, A.; Manca, A.; Meli, G.; et al. NH2-truncated human tau induces deregulated mitophagy in neurons by aberrant recruitment of Parkin and UCHL-1: Implications in Alzheimer’s disease. Hum. Mol. Genet. 2015, 24, 3058–3081. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Li, X.C.; Wang, Z.H.; Luo, Y.; Zhang, X.; Liu, X.P.; Feng, Q.; Wang, Q.; Yue, Z.; Chen, Z.; et al. Tau accumulation impairs mitophagy via increasing mitochondrial membrane potential and reducing mitochondrial Parkin. Oncotarget 2016, 7, 17356–17368. [Google Scholar] [CrossRef] [Green Version]
- Cummins, N.; Tweedie, A.; Zuryn, S.; Bertran-Gonzalez, J.; Gotz, J. Disease-associated tau impairs mitophagy by inhibiting Parkin translocation to mitochondria. EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Fang, E.F.; Hou, Y.; Palikaras, K.; Adriaanse, B.A.; Kerr, J.S.; Yang, B.; Lautrup, S.; Hasan-Olive, M.M.; Caponio, D.; Dan, X.; et al. Mitophagy inhibits amyloid-beta and tau pathology and reverses cognitive deficits in models of Alzheimer’s disease. Nat. Neurosci. 2019, 22, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, V.; Gustafsson, G.; Sanders, L.H.; Howlett, E.H.; Sigvardson, J.; Kasrayan, A.; Ingelsson, M.; Bergstrom, J.; Erlandsson, A. Extensive uptake of alpha-synuclein oligomers in astrocytes results in sustained intracellular deposits and mitochondrial damage. Mol. Cell Neurosci. 2017, 82, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Pukass, K.; Goldbaum, O.; Richter-Landsberg, C. Mitochondrial impairment and oxidative stress compromise autophagosomal degradation of alpha-synuclein in oligodendroglial cells. J. Neurochem. 2015, 135, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherer, T.B.; Betarbet, R.; Stout, A.K.; Lund, S.; Baptista, M.; Panov, A.V.; Cookson, M.R.; Greenamyre, J.T. An in vitro model of Parkinson’s disease: Linking mitochondrial impairment to altered alpha-synuclein metabolism and oxidative damage. J. Neurosci. 2002, 22, 7006–7015. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.H.; Shaltouki, A.; Gonzalez, A.E.; Bettencourt da Cruz, A.; Burbulla, L.F.; St Lawrence, E.; Schule, B.; Krainc, D.; Palmer, T.D.; Wang, X. Functional Impairment in Miro Degradation and Mitophagy Is a Shared Feature in Familial and Sporadic Parkinson’s Disease. Cell Stem Cell 2016, 19, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Burchell, V.S.; Nelson, D.E.; Sanchez-Martinez, A.; Delgado-Camprubi, M.; Ivatt, R.M.; Pogson, J.H.; Randle, S.J.; Wray, S.; Lewis, P.A.; Houlden, H.; et al. The Parkinson’s disease-linked proteins Fbxo7 and Parkin interact to mediate mitophagy. Nat. Neurosci. 2013, 16, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.D.; Xie, S.P.; Sathiyamoorthy, S.; Saw, W.T.; Sing, T.Y.; Ng, S.H.; Chua, H.P.; Tang, A.M.; Shaffra, F.; Li, Z.; et al. F-box protein 7 mutations promote protein aggregation in mitochondria and inhibit mitophagy. Hum. Mol. Genet. 2015, 24, 6314–6330. [Google Scholar] [CrossRef] [Green Version]
- Bender, A.; Krishnan, K.J.; Morris, C.M.; Taylor, G.A.; Reeve, A.K.; Perry, R.H.; Jaros, E.; Hersheson, J.S.; Betts, J.; Klopstock, T.; et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat. Genet. 2006, 38, 515–517. [Google Scholar] [CrossRef]
- Dagda, R.K.; Cherra, S.J., 3rd; Kulich, S.M.; Tandon, A.; Park, D.; Chu, C.T. Loss of PINK1 function promotes mitophagy through effects on oxidative stress and mitochondrial fission. J. Biol. Chem. 2009, 284, 13843–13855. [Google Scholar] [CrossRef] [Green Version]
- Dagda, R.K.; Zhu, J.; Kulich, S.M.; Chu, C.T. Mitochondrially localized ERK2 regulates mitophagy and autophagic cell stress: Implications for Parkinson’s disease. Autophagy 2008, 4, 770–782. [Google Scholar] [CrossRef] [Green Version]
- Osellame, L.D.; Duchen, M.R. Defective quality control mechanisms and accumulation of damaged mitochondria link Gaucher and Parkinson diseases. Autophagy 2013, 9, 1633–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.H.; Horbinski, C.; Guo, F.; Watkins, S.; Uchiyama, Y.; Chu, C.T. Regulation of autophagy by extracellular signal-regulated protein kinases during 1-methyl-4-phenylpyridinium-induced cell death. Am. J. Pathol. 2007, 170, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.T. Multiple pathways for mitophagy: A neurodegenerative conundrum for Parkinson’s disease. Neurosci. Lett. 2019, 697, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, A.J.; Theodore, D.A.; Greene, J.C.; Benes, H.; Wes, P.D.; Pallanck, L.J. Increased glutathione S-transferase activity rescues dopaminergic neuron loss in a Drosophila model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 8024–8029. [Google Scholar] [CrossRef] [Green Version]
- Cha, G.H.; Kim, S.; Park, J.; Lee, E.; Kim, M.; Lee, S.B.; Kim, J.M.; Chung, J.; Cho, K.S. Parkin negatively regulates JNK pathway in the dopaminergic neurons of Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 10345–10350. [Google Scholar] [CrossRef] [Green Version]
- Greene, J.C.; Whitworth, A.J.; Kuo, I.; Andrews, L.A.; Feany, M.B.; Pallanck, L.J. Mitochondrial pathology and apoptotic muscle degeneration in Drosophila parkin mutants. Proc. Natl. Acad. Sci. USA 2003, 100, 4078–4083. [Google Scholar] [CrossRef] [Green Version]
- Pesah, Y.; Pham, T.; Burgess, H.; Middlebrooks, B.; Verstreken, P.; Zhou, Y.; Harding, M.; Bellen, H.; Mardon, G. Drosophila parkin mutants have decreased mass and cell size and increased sensitivity to oxygen radical stress. Development 2004, 131, 2183–2194. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Gehrke, S.; Imai, Y.; Huang, Z.; Ouyang, Y.; Wang, J.W.; Yang, L.; Beal, M.F.; Vogel, H.; Lu, B. Mitochondrial pathology and muscle and dopaminergic neuron degeneration caused by inactivation of Drosophila Pink1 is rescued by Parkin. Proc. Natl. Acad. Sci. USA 2006, 103, 10793–10798. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, T.; Vilain, S.; Vints, K.; Gounko, N.; Verstreken, P.; Vandenberghe, W. Deficiency of parkin and PINK1 impairs age-dependent mitophagy in Drosophila. Elife 2018, 7. [Google Scholar] [CrossRef]
- Dave, K.D.; De Silva, S.; Sheth, N.P.; Ramboz, S.; Beck, M.J.; Quang, C.; Switzer, R.C., 3rd; Ahmad, S.O.; Sunkin, S.M.; Walker, D.; et al. Phenotypic characterization of recessive gene knockout rat models of Parkinson’s disease. Neurobiol. Dis. 2014, 70, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Pickrell, A.M.; Huang, C.H.; Kennedy, S.R.; Ordureau, A.; Sideris, D.P.; Hoekstra, J.G.; Harper, J.W.; Youle, R.J. Endogenous Parkin Preserves Dopaminergic Substantia Nigral Neurons following Mitochondrial DNA Mutagenic Stress. Neuron 2015, 87, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Rogers, R.S.; Tungtur, S.; Tanaka, T.; Nadeau, L.L.; Badawi, Y.; Wang, H.; Ni, H.M.; Ding, W.X.; Nishimune, H. Impaired Mitophagy Plays a Role in Denervation of Neuromuscular Junctions in ALS Mice. Front. Neurosci. 2017, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhou, B.; Lin, M.Y.; Wang, S.; Foust, K.D.; Sheng, Z.H. Endolysosomal Deficits Augment Mitochondria Pathology in Spinal Motor Neurons of Asymptomatic fALS Mice. Neuron 2015, 87, 355–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomo, G.M.; Granatiero, V.; Kawamata, H.; Konrad, C.; Kim, M.; Arreguin, A.J.; Zhao, D.; Milner, T.A.; Manfredi, G. Parkin is a disease modifier in the mutant SOD1 mouse model of ALS. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Birsa, N.; Norkett, R.; Wauer, T.; Mevissen, T.E.; Wu, H.C.; Foltynie, T.; Bhatia, K.; Hirst, W.D.; Komander, D.; Plun-Favreau, H.; et al. Lysine 27 ubiquitination of the mitochondrial transport protein Miro is dependent on serine 65 of the Parkin ubiquitin ligase. J. Biol. Chem. 2014, 289, 14569–14582. [Google Scholar] [CrossRef] [Green Version]
- Bingol, B.; Tea, J.S.; Phu, L.; Reichelt, M.; Bakalarski, C.E.; Song, Q.H.; Foreman, O.; Kirkpatrick, D.S.; Sheng, M.G. The mitochondrial deubiquitinase USP30 opposes parkin-mediated mitophagy. Nature 2014, 510, 370. [Google Scholar] [CrossRef]
- Liu, S.; Sawada, T.; Lee, S.; Yu, W.; Silverio, G.; Alapatt, P.; Millan, I.; Shen, A.; Saxton, W.; Kanao, T.; et al. Parkinson’s disease-associated kinase PINK1 regulates Miro protein level and axonal transport of mitochondria. PLoS Genet. 2012, 8, e1002537. [Google Scholar] [CrossRef]
- Wang, X.; Winter, D.; Ashrafi, G.; Schlehe, J.; Wong, Y.L.; Selkoe, D.; Rice, S.; Steen, J.; LaVoie, M.J.; Schwarz, T.L. PINK1 and Parkin target Miro for phosphorylation and degradation to arrest mitochondrial motility. Cell 2011, 147, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wang, W.; Siedlak, S.L.; Liu, Y.; Liu, J.; Jiang, K.; Perry, G.; Zhu, X.; Wang, X. Miro1 deficiency in amyotrophic lateral sclerosis. Front. Aging Neurosci. 2015, 7, 100. [Google Scholar] [CrossRef] [Green Version]
- Moller, A.; Bauer, C.S.; Cohen, R.N.; Webster, C.P.; De Vos, K.J. Amyotrophic lateral sclerosis-associated mutant SOD1 inhibits anterograde axonal transport of mitochondria by reducing Miro1 levels. Hum. Mol. Genet. 2017, 26, 4668–4679. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Oh, S.S.; Weaver, D.; Lewandowska, A.; Maxfield, D.; Schuler, M.H.; Smith, N.K.; Macfarlane, J.; Saunders, G.; Palmer, C.A.; et al. Loss of Miro1-directed mitochondrial movement results in a novel murine model for neuron disease. Proc. Natl. Acad. Sci. USA 2014, 111, E3631–E3640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, H.; Morino, H.; Ito, H.; Izumi, Y.; Kato, H.; Watanabe, Y.; Kinoshita, Y.; Kamada, M.; Nodera, H.; Suzuki, H.; et al. Mutations of optineurin in amyotrophic lateral sclerosis. Nature 2010, 465, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Freischmidt, A.; Wieland, T.; Richter, B.; Ruf, W.; Schaeffer, V.; Muller, K.; Marroquin, N.; Nordin, F.; Hubers, A.; Weydt, P.; et al. Haploinsufficiency of TBK1 causes familial ALS and fronto-temporal dementia. Nat. Neurosci. 2015, 18, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Fecto, F.; Yan, J.; Vemula, S.P.; Liu, E.; Yang, Y.; Chen, W.; Zheng, J.G.; Shi, Y.; Siddique, N.; Arrat, H.; et al. SQSTM1 mutations in familial and sporadic amyotrophic lateral sclerosis. Arch. Neurol. 2011, 68, 1440–1446. [Google Scholar] [CrossRef]
- Hwang, S.; Disatnik, M.H.; Mochly-Rosen, D. Impaired GAPDH-induced mitophagy contributes to the pathology of Huntington’s disease. EMBO Mol. Med. 2015, 7, 1307–1326. [Google Scholar] [CrossRef]
- Ross, C.A.; Tabrizi, S.J. Huntington’s disease: From molecular pathogenesis to clinical treatment. Lancet Neurol. 2011, 10, 83–98. [Google Scholar] [CrossRef]
- Martinez-Vicente, M.; Talloczy, Z.; Wong, E.; Tang, G.; Koga, H.; Kaushik, S.; de Vries, R.; Arias, E.; Harris, S.; Sulzer, D.; et al. Cargo recognition failure is responsible for inefficient autophagy in Huntington’s disease. Nat. Neurosci. 2010, 13, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.N.; Xu, Z.; Patel, B.; Chen, Z.; Chen, D.; Tito, A.; David, G.; Sun, Y.; Stimming, E.F.; Bellen, H.J.; et al. Huntingtin functions as a scaffold for selective macroautophagy. Nat. Cell Biol. 2015, 17, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Gleichmann, M.; Cheng, A. Mitochondria in neuroplasticity and neurological disorders. Neuron 2008, 60, 748–766. [Google Scholar] [CrossRef] [Green Version]
- Fang, E.F.; Lautrup, S.; Hou, Y.; Demarest, T.G.; Croteau, D.L.; Mattson, M.P.; Bohr, V.A. NAD(+) in Aging: Molecular Mechanisms and Translational Implications. Trends Mol. Med. 2017, 23, 899–916. [Google Scholar] [CrossRef]
- Gupta, V.K.; Scheunemann, L.; Eisenberg, T.; Mertel, S.; Bhukel, A.; Koemans, T.S.; Kramer, J.M.; Liu, K.S.; Schroeder, S.; Stunnenberg, H.G.; et al. Restoring polyamines protects from age-induced memory impairment in an autophagy-dependent manner. Nat. Neurosci. 2013, 16, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in health and disease. Science 2018, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, B.; Pan, Y.; Vempati, P.; Zhao, W.; Knable, L.; Ho, L.; Wang, J.; Sastre, M.; Ono, K.; Sauve, A.A.; et al. Nicotinamide riboside restores cognition through an upregulation of proliferator-activated receptor-gamma coactivator 1alpha regulated beta-secretase 1 degradation and mitochondrial gene expression in Alzheimer’s mouse models. Neurobiol. Aging 2013, 34, 1581–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Lautrup, S.; Cordonnier, S.; Wang, Y.; Croteau, D.L.; Zavala, E.; Zhang, Y.; Moritoh, K.; O’Connell, J.F.; Baptiste, B.A.; et al. NAD(+) supplementation normalizes key Alzheimer’s features and DNA damage responses in a new AD mouse model with introduced DNA repair deficiency. Proc. Natl. Acad. Sci. USA 2018, 115, E1876–E1885. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Pitta, M.; Jiang, H.; Lee, J.H.; Zhang, G.; Chen, X.; Kawamoto, E.M.; Mattson, M.P. Nicotinamide forestalls pathology and cognitive decline in Alzheimer mice: Evidence for improved neuronal bioenergetics and autophagy procession. Neurobiol. Aging 2013, 34, 1564–1580. [Google Scholar] [CrossRef] [Green Version]
- Turunc Bayrakdar, E.; Uyanikgil, Y.; Kanit, L.; Koylu, E.; Yalcin, A. Nicotinamide treatment reduces the levels of oxidative stress, apoptosis, and PARP-1 activity in Abeta(1-42)-induced rat model of Alzheimer’s disease. Free Radic. Res. 2014, 48, 146–158. [Google Scholar] [CrossRef]
- Schondorf, D.C.; Ivanyuk, D.; Baden, P.; Sanchez-Martinez, A.; De Cicco, S.; Yu, C.; Giunta, I.; Schwarz, L.K.; Di Napoli, G.; Panagiotakopoulou, V.; et al. The NAD+ Precursor Nicotinamide Riboside Rescues Mitochondrial Defects and Neuronal Loss in iPSC and Fly Models of Parkinson’s Disease. Cell Rep. 2018, 23, 2976–2988. [Google Scholar] [CrossRef]
- Fan, J.; Yang, X.; Li, J.; Shu, Z.; Dai, J.; Liu, X.; Li, B.; Jia, S.; Kou, X.; Yang, Y.; et al. Spermidine coupled with exercise rescues skeletal muscle atrophy from D-gal-induced aging rats through enhanced autophagy and reduced apoptosis via AMPK-FOXO3a signal pathway. Oncotarget 2017, 8, 17475–17490. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Buttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Geisler, J.G.; Marosi, K.; Halpern, J.; Mattson, M.P. DNP, mitochondrial uncoupling, and neuroprotection: A little dab’ll do ya. Alzheimers Dement. 2017, 13, 582–591. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Mattson, M.P. Dietary restriction and 2-deoxyglucose administration improve behavioral outcome and reduce degeneration of dopaminergic neurons in models of Parkinson’s disease. J. Neurosci. Res. 1999, 57, 195–206. [Google Scholar] [CrossRef]
- Yao, J.; Chen, S.; Mao, Z.; Cadenas, E.; Brinton, R.D. 2-Deoxy-D-glucose treatment induces ketogenesis, sustains mitochondrial function, and reduces pathology in female mouse model of Alzheimer’s disease. PLoS ONE 2011, 6, e21788. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.S.; Ingram, D.K. Manipulation of health span and function by dietary caloric restriction mimetics. Ann. N. Y. Acad. Sci. 2016, 1363, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, B.; Vacher, C.; Berger, Z.; Davies, J.E.; Luo, S.; Oroz, L.G.; Scaravilli, F.; Easton, D.F.; Duden, R.; O’Kane, C.J.; et al. Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat. Genet. 2004, 36, 585–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spilman, P.; Podlutskaya, N.; Hart, M.J.; Debnath, J.; Gorostiza, O.; Bredesen, D.; Richardson, A.; Strong, R.; Galvan, V. Inhibition of mTOR by rapamycin abolishes cognitive deficits and reduces amyloid-beta levels in a mouse model of Alzheimer’s disease. PLoS ONE 2010, 5, e9979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, J.J.; Hellberg, K.; Turner, M.; Talbott, G.; Kolar, M.J.; Ross, D.S.; Hoxhaj, G.; Saghatelian, A.; Shaw, R.J.; Manning, B.D. Metformin Inhibits Hepatic mTORC1 Signaling via Dose-Dependent Mechanisms Involving AMPK and the TSC Complex. Cell Metab. 2017, 25, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Chen, H.; Li, J.; Li, T.; Zheng, B.; Zheng, Y.; Jin, H.; He, Y.; Gu, Q.; Xu, X. Sirtuin 1-mediated cellular metabolic memory of high glucose via the LKB1/AMPK/ROS pathway and therapeutic effects of metformin. Diabetes 2012, 61, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.M.; Lee, W.K.; Lee, Y.H.; Kang, E.S.; Cha, B.S.; Lee, B.W. Metformin Restores Parkin-Mediated Mitophagy, Suppressed by Cytosolic p53. Int. J. Mol. Sci. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Ryu, D.; Mouchiroud, L.; Andreux, P.A.; Katsyuba, E.; Moullan, N.; Nicolet-Dit-Felix, A.A.; Williams, E.G.; Jha, P.; Lo Sasso, G.; Huzard, D.; et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 2016, 22, 879–888. [Google Scholar] [CrossRef]
- Morselli, E.; Marino, G.; Bennetzen, M.V.; Eisenberg, T.; Megalou, E.; Schroeder, S.; Cabrera, S.; Benit, P.; Rustin, P.; Criollo, A.; et al. Spermidine and resveratrol induce autophagy by distinct pathways converging on the acetylproteome. J. Cell Biol. 2011, 192, 615–629. [Google Scholar] [CrossRef] [Green Version]
- Lonskaya, I.; Hebron, M.L.; Desforges, N.M.; Schachter, J.B.; Moussa, C.E. Nilotinib-induced autophagic changes increase endogenous parkin level and ubiquitination, leading to amyloid clearance. J. Mol. Med. (Berl.) 2014, 92, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.S.; Lee, Y.; Shin, J.H.; Karuppagounder, S.S.; Gadad, B.S.; Koleske, A.J.; Pletnikova, O.; Troncoso, J.C.; Dawson, V.L.; Dawson, T.M. Phosphorylation by the c-Abl protein tyrosine kinase inhibits parkin’s ubiquitination and protective function. Proc. Natl. Acad. Sci. USA 2010, 107, 16691–16696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonskaya, I.; Hebron, M.L.; Desforges, N.M.; Franjie, A.; Moussa, C.E. Tyrosine kinase inhibition increases functional parkin-Beclin-1 interaction and enhances amyloid clearance and cognitive performance. EMBO Mol. Med. 2013, 5, 1247–1262. [Google Scholar] [CrossRef] [PubMed]
- Wenqiang, C.; Lonskaya, I.; Hebron, M.L.; Ibrahim, Z.; Olszewski, R.T.; Neale, J.H.; Moussa, C.E. Parkin-mediated reduction of nuclear and soluble TDP-43 reverses behavioral decline in symptomatic mice. Hum. Mol. Genet. 2014, 23, 4960–4969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelso, G.F.; Porteous, C.M.; Coulter, C.V.; Hughes, G.; Porteous, W.K.; Ledgerwood, E.C.; Smith, R.A.; Murphy, M.P. Selective targeting of a redox-active ubiquinone to mitochondria within cells: Antioxidant and antiapoptotic properties. J. Biol. Chem. 2001, 276, 4588–4596. [Google Scholar] [CrossRef] [Green Version]
- Miquel, E.; Cassina, A.; Martinez-Palma, L.; Souza, J.M.; Bolatto, C.; Rodriguez-Bottero, S.; Logan, A.; Smith, R.A.; Murphy, M.P.; Barbeito, L.; et al. Neuroprotective effects of the mitochondria-targeted antioxidant MitoQ in a model of inherited amyotrophic lateral sclerosis. Free Radic. Biol. Med. 2014, 70, 204–213. [Google Scholar] [CrossRef]
- McManus, M.J.; Murphy, M.P.; Franklin, J.L. The mitochondria-targeted antioxidant MitoQ prevents loss of spatial memory retention and early neuropathology in a transgenic mouse model of Alzheimer’s disease. J. Neurosci. 2011, 31, 15703–15715. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.F.; Gruber, J.; Cheah, I.K.; Goo, C.K.; Cheong, W.F.; Shui, G.; Sit, K.P.; Wenk, M.R.; Halliwell, B. The mitochondria-targeted antioxidant MitoQ extends lifespan and improves healthspan of a transgenic Caenorhabditis elegans model of Alzheimer disease. Free Radic. Biol. Med. 2014, 71, 390–401. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The mitophagy pathways. Upon mitochondrial damage, mitophagy can be induced through three major mechanisms: ubiquitin-mediated mitophagy including PTEN-induced putative kinase protein 1 (PINK1)-Parkin-dependent mitophagy, outer mitochondrial membrane (OMM) receptor-mediated mitophagy, and lipid-mediated mitophagy. (a) PINK1-Parkin-mediated mitophagy initiates with PINK1 stabilization on the OMM of damaged mitochondria to recruit Parkin, an E3 ubiquitin ligase. Phospho-ubiquitination of substrates on the OMM by PINK1/Parkin recruits the autophagy machinery and thus promotes the engulfment of damaged mitochondria by growing phagophore or isolation membranes. (b) Other E3 ubiquitin ligases have been reported to regulate mitophagy independent of PINK1 and Parkin. MUL1 has similar substrates to Parkin and can directly bind to GABAA receptor-associated protein (GABARAP), suggesting that MUL1 can function independently to facilitate autophagic engulfment. (c) Several autophagy receptors are anchored within the OMM, including NIP3-like protein X (NIX; also known as BNIP3L), FUN14 domain containing 1 (FUNDC1), and FK506 Binding Protein 8 (FKBP8). Binding of NIX/BNIP3L or FUNDC1 to LC3 or GABARAP on the phagophore mediates targeting dysfunctional mitochondria for autophagy. (d) Externalization of cardiolipin, normally found in the inner mitochondrial membrane (IMM) phospholipid, to the OMM is a unique mechanism for lipid-mediated mitophagy. Cardiolipin initiates mitophagy through its direct interaction with LC3.
Figure 1.
The mitophagy pathways. Upon mitochondrial damage, mitophagy can be induced through three major mechanisms: ubiquitin-mediated mitophagy including PTEN-induced putative kinase protein 1 (PINK1)-Parkin-dependent mitophagy, outer mitochondrial membrane (OMM) receptor-mediated mitophagy, and lipid-mediated mitophagy. (a) PINK1-Parkin-mediated mitophagy initiates with PINK1 stabilization on the OMM of damaged mitochondria to recruit Parkin, an E3 ubiquitin ligase. Phospho-ubiquitination of substrates on the OMM by PINK1/Parkin recruits the autophagy machinery and thus promotes the engulfment of damaged mitochondria by growing phagophore or isolation membranes. (b) Other E3 ubiquitin ligases have been reported to regulate mitophagy independent of PINK1 and Parkin. MUL1 has similar substrates to Parkin and can directly bind to GABAA receptor-associated protein (GABARAP), suggesting that MUL1 can function independently to facilitate autophagic engulfment. (c) Several autophagy receptors are anchored within the OMM, including NIP3-like protein X (NIX; also known as BNIP3L), FUN14 domain containing 1 (FUNDC1), and FK506 Binding Protein 8 (FKBP8). Binding of NIX/BNIP3L or FUNDC1 to LC3 or GABARAP on the phagophore mediates targeting dysfunctional mitochondria for autophagy. (d) Externalization of cardiolipin, normally found in the inner mitochondrial membrane (IMM) phospholipid, to the OMM is a unique mechanism for lipid-mediated mitophagy. Cardiolipin initiates mitophagy through its direct interaction with LC3.

Figure 2.
Neuronal mitophagy. Upon mitochondrial membrane potential (Δψm) dissipation, Parkin is recruited to depolarized mitochondria, triggering mitochondrial engulfment by autophagosomes. Parkin-targeted mitochondria accumulate in the somatodendritic regions of neurons. Such compartmental restriction is attributed to altered mitochondrial motility along axons, as evidenced by decreased anterograde transport and relatively increased retrograde transport of mitochondria. This spatial process allows neurons to efficiently remove damaged mitochondria from axonal terminals and facilitate mitophagic clearance in the soma, where mature lysosomes are mainly located. Studies have also shown that autophagosomes containing engulfed mitochondria move in an exclusively retrograde direction from distal axons toward the soma for maturation and for more efficient cargo degradation within acidic lysosomes in the soma. Figure is modified from [2].
Figure 2.
Neuronal mitophagy. Upon mitochondrial membrane potential (Δψm) dissipation, Parkin is recruited to depolarized mitochondria, triggering mitochondrial engulfment by autophagosomes. Parkin-targeted mitochondria accumulate in the somatodendritic regions of neurons. Such compartmental restriction is attributed to altered mitochondrial motility along axons, as evidenced by decreased anterograde transport and relatively increased retrograde transport of mitochondria. This spatial process allows neurons to efficiently remove damaged mitochondria from axonal terminals and facilitate mitophagic clearance in the soma, where mature lysosomes are mainly located. Studies have also shown that autophagosomes containing engulfed mitochondria move in an exclusively retrograde direction from distal axons toward the soma for maturation and for more efficient cargo degradation within acidic lysosomes in the soma. Figure is modified from [2].

Figure 3.
Neurodegenerative disease-associated mitochondrial toxicity. Misfolded proteins, oligomers, protein aggregates, or fibrils linked to major neurodegenerative diseases induce mitochondrial abnormalities, leading to increased reactive oxygen species (ROS) levels, loss of mitochondrial membrane potential (Δψm), decreased oxidative phosphorylation (OXPHOS) and ATP production, impaired Ca2+ buffering, and enhanced mitochondrial DNA (mtDNA) changes. Moreover, impairments in mitochondrial dynamics and trafficking as well as mitophagy result in excessive mitochondrial fission and fragmentation and aberrant accumulation of dysfunctional mitochondria, all of which collectively contribute to synaptic dysfunction, axonal degeneration, and neuronal death.
Figure 3.
Neurodegenerative disease-associated mitochondrial toxicity. Misfolded proteins, oligomers, protein aggregates, or fibrils linked to major neurodegenerative diseases induce mitochondrial abnormalities, leading to increased reactive oxygen species (ROS) levels, loss of mitochondrial membrane potential (Δψm), decreased oxidative phosphorylation (OXPHOS) and ATP production, impaired Ca2+ buffering, and enhanced mitochondrial DNA (mtDNA) changes. Moreover, impairments in mitochondrial dynamics and trafficking as well as mitophagy result in excessive mitochondrial fission and fragmentation and aberrant accumulation of dysfunctional mitochondria, all of which collectively contribute to synaptic dysfunction, axonal degeneration, and neuronal death.

Figure 4.
Mitophagy defects in neurodegenerative diseases. A growing body of evidence indicate that mitophagy function is impaired in major neurodegenerative diseases. In Alzheimer’s disease (AD), robust activation of Parkin-mediated mitophagy was observed in patient brains of familiar and sporadic AD and the cell and animal models. However, under tauopathy conditions, excessive or defective Parkin-dependent mitochondrial turnover was reported in different studies, respectively. In addition, impaired lysosomal proteolysis and reduced levels of the basal mitophagy collectively contribute to mitophagy dysfunction in AD. In Parkinson’s disease (PD), PINK1-Parkin-dependent mitophagy is necessary for the function and survival of PD-related dopaminergic neurons in response to stress/pathological stimuli. The role for increased cardiolipin-mediated mitophagy in mutant α-syn-induced mitochondrial dysfunction has also been proposed. As for amyotrophic lateral sclerosis (ALS), enhanced Parkin-mediated mitophagy was demonstrated in an ALS mouse model. Mitophagy defects and mitochondrial pathology are also attributed to lysosomal deficit in ALS affected motor neurons. In Huntington’s disease (HD), decreased basal levels of mitophagy, defects in autophagic recognition and recruitment of damaged mitochondria, and impaired GAPDH-mediated micro-mitophagy lead to pathological mitophagy and mitochondrial dysfunction.
Figure 4.
Mitophagy defects in neurodegenerative diseases. A growing body of evidence indicate that mitophagy function is impaired in major neurodegenerative diseases. In Alzheimer’s disease (AD), robust activation of Parkin-mediated mitophagy was observed in patient brains of familiar and sporadic AD and the cell and animal models. However, under tauopathy conditions, excessive or defective Parkin-dependent mitochondrial turnover was reported in different studies, respectively. In addition, impaired lysosomal proteolysis and reduced levels of the basal mitophagy collectively contribute to mitophagy dysfunction in AD. In Parkinson’s disease (PD), PINK1-Parkin-dependent mitophagy is necessary for the function and survival of PD-related dopaminergic neurons in response to stress/pathological stimuli. The role for increased cardiolipin-mediated mitophagy in mutant α-syn-induced mitochondrial dysfunction has also been proposed. As for amyotrophic lateral sclerosis (ALS), enhanced Parkin-mediated mitophagy was demonstrated in an ALS mouse model. Mitophagy defects and mitochondrial pathology are also attributed to lysosomal deficit in ALS affected motor neurons. In Huntington’s disease (HD), decreased basal levels of mitophagy, defects in autophagic recognition and recruitment of damaged mitochondria, and impaired GAPDH-mediated micro-mitophagy lead to pathological mitophagy and mitochondrial dysfunction.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cai, Q.; Jeong, Y.Y. Mitophagy in Alzheimer’s Disease and Other Age-Related Neurodegenerative Diseases. Cells 2020, 9, 150. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010150
AMA Style
Cai Q, Jeong YY. Mitophagy in Alzheimer’s Disease and Other Age-Related Neurodegenerative Diseases. Cells. 2020; 9(1):150. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010150
Chicago/Turabian StyleCai, Qian, and Yu Young Jeong. 2020. "Mitophagy in Alzheimer’s Disease and Other Age-Related Neurodegenerative Diseases" Cells 9, no. 1: 150. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9010150
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.