Involvement of Enteric Glia in Small Intestine Neuromuscular Dysfunction of Toll-Like Receptor 4-Deficient Mice

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Fluoroacetate Treatment
2.3. Neuromuscular Function Studies
2.4. Immunohistochemistry on Ileal whole Mount Preparations
2.5. Confocal Image Acquisition and Analysis
2.6. RNA Isolation and Quantitative RT-PCR
2.7. Chemicals
2.8. Statistical Analysis
3. Results
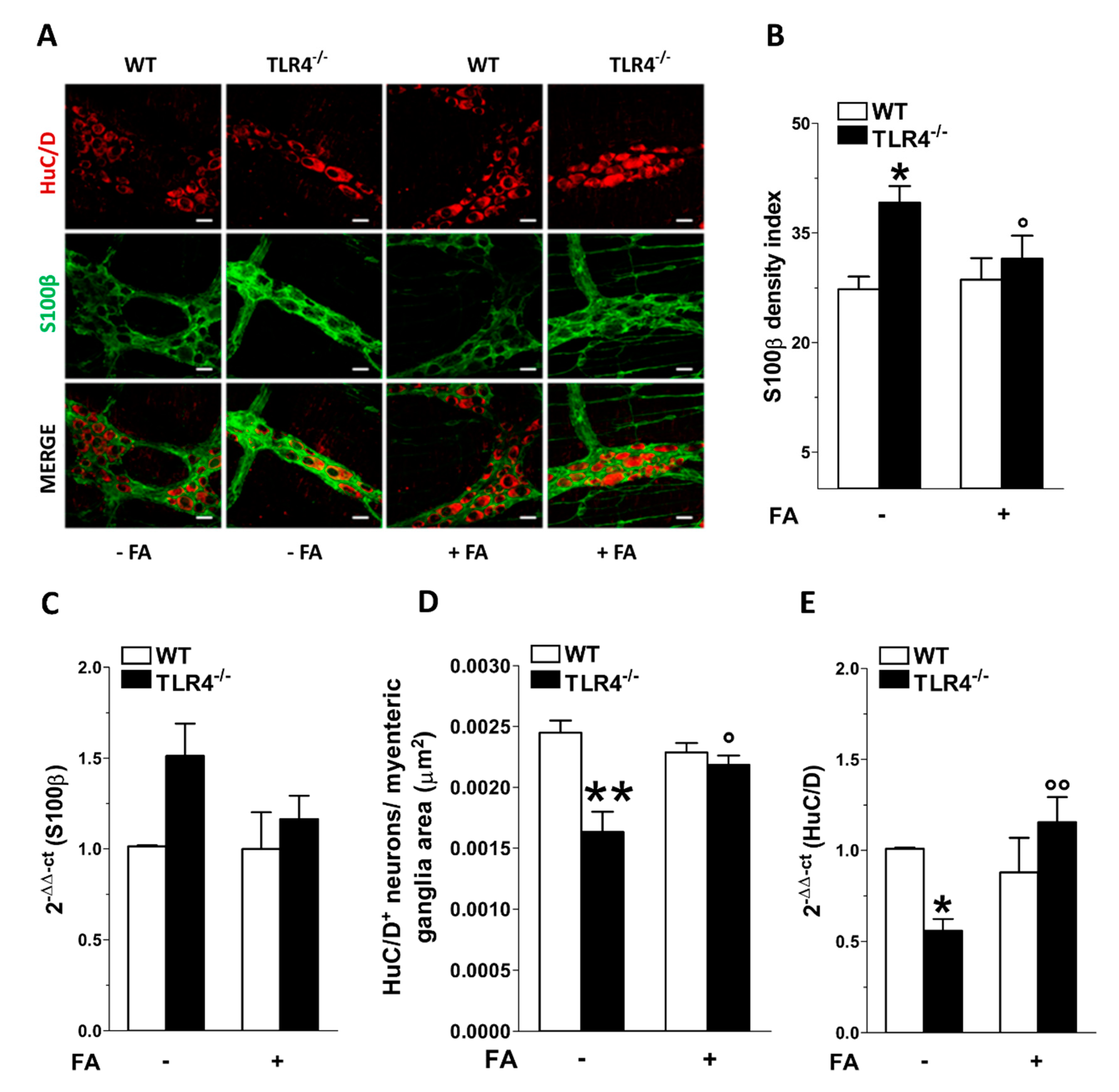
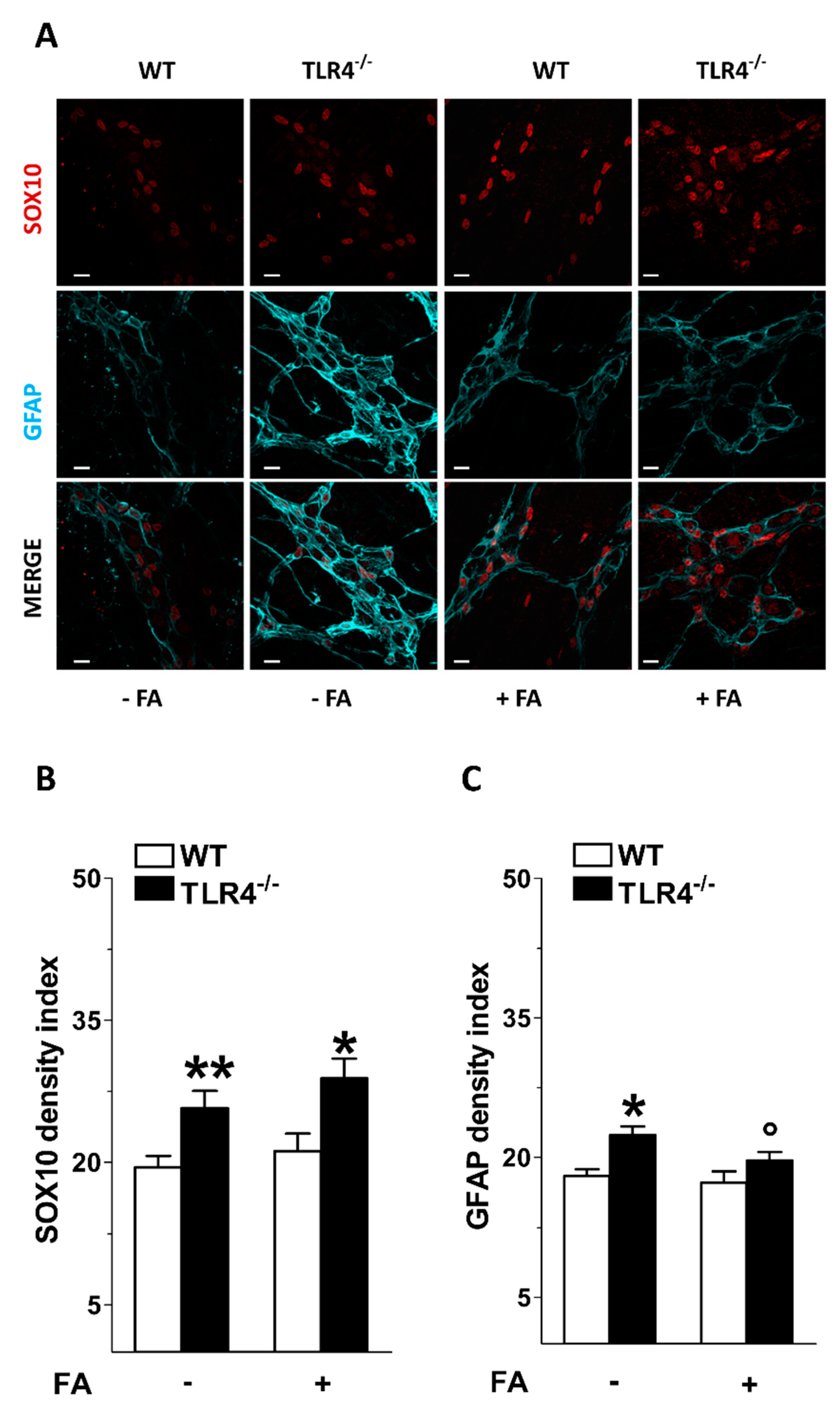
3.1. In Vitro Fluoroacetate Treatment Reduces Enteric Reactive Gliosis in TLR4−/− Mice
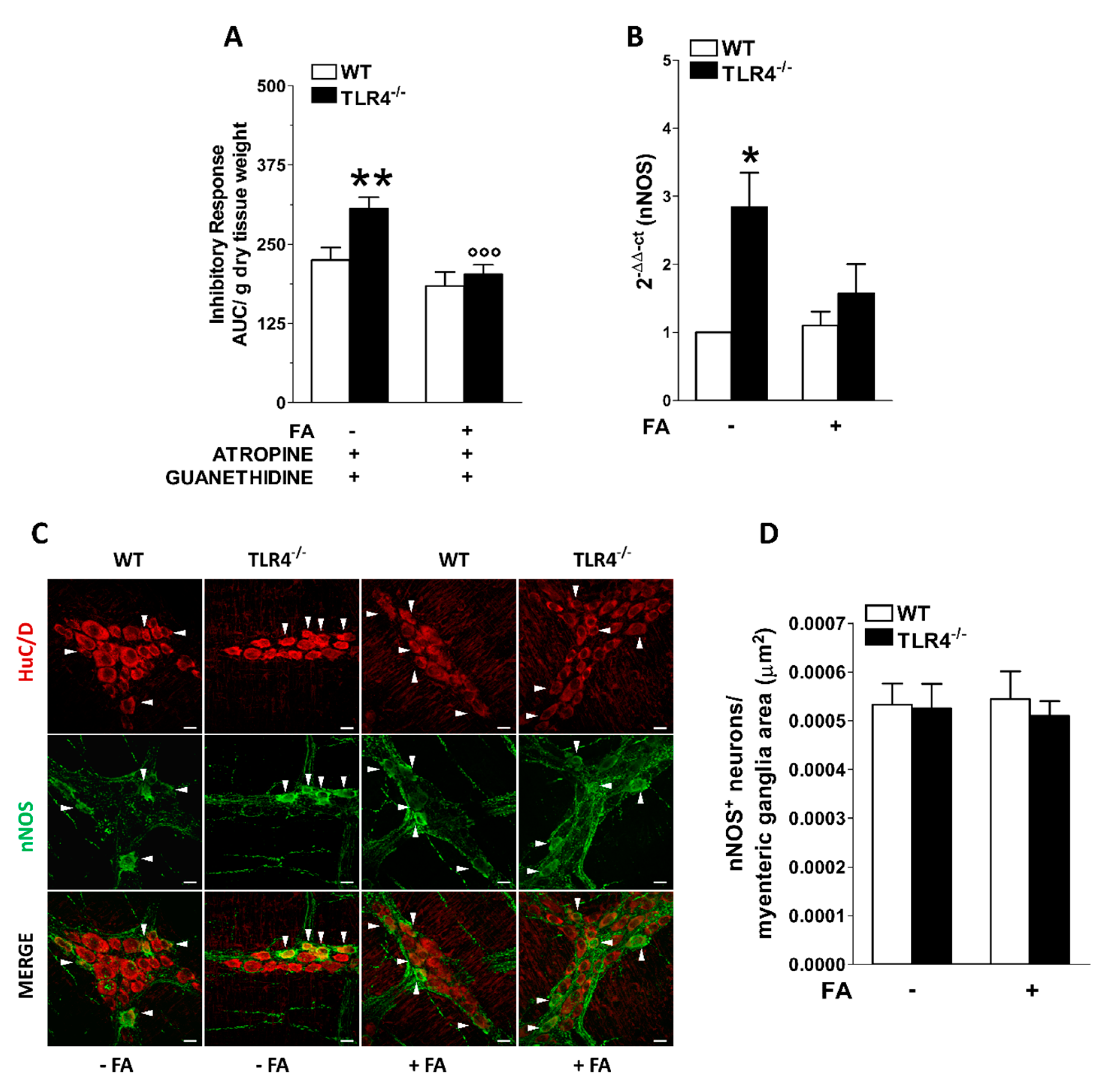
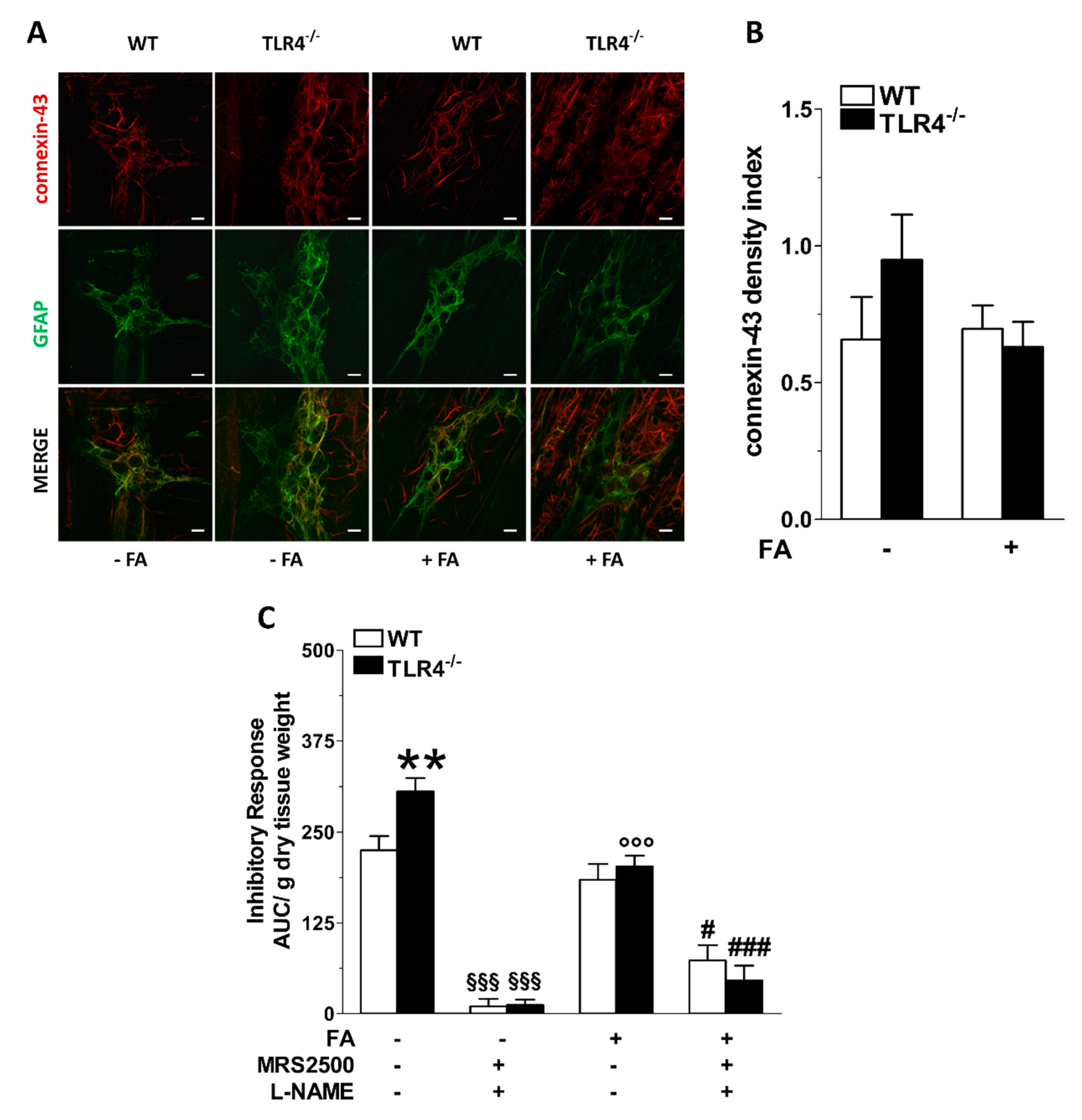
3.2. Influence of Enteric Glial Cells (EGCs) on Nitrergic Neurotransmission in Small Intestine of TLR4−/− Mice
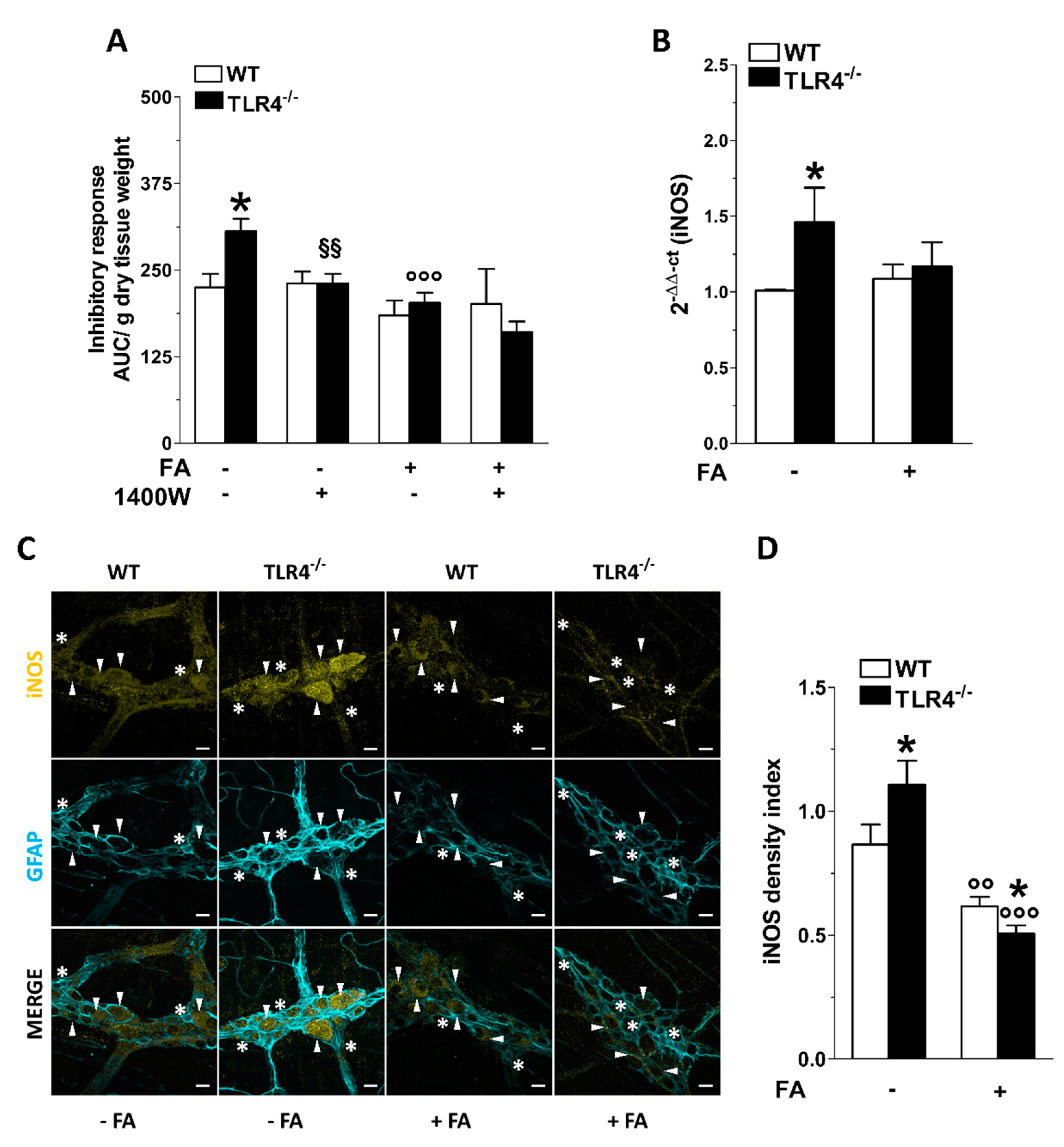
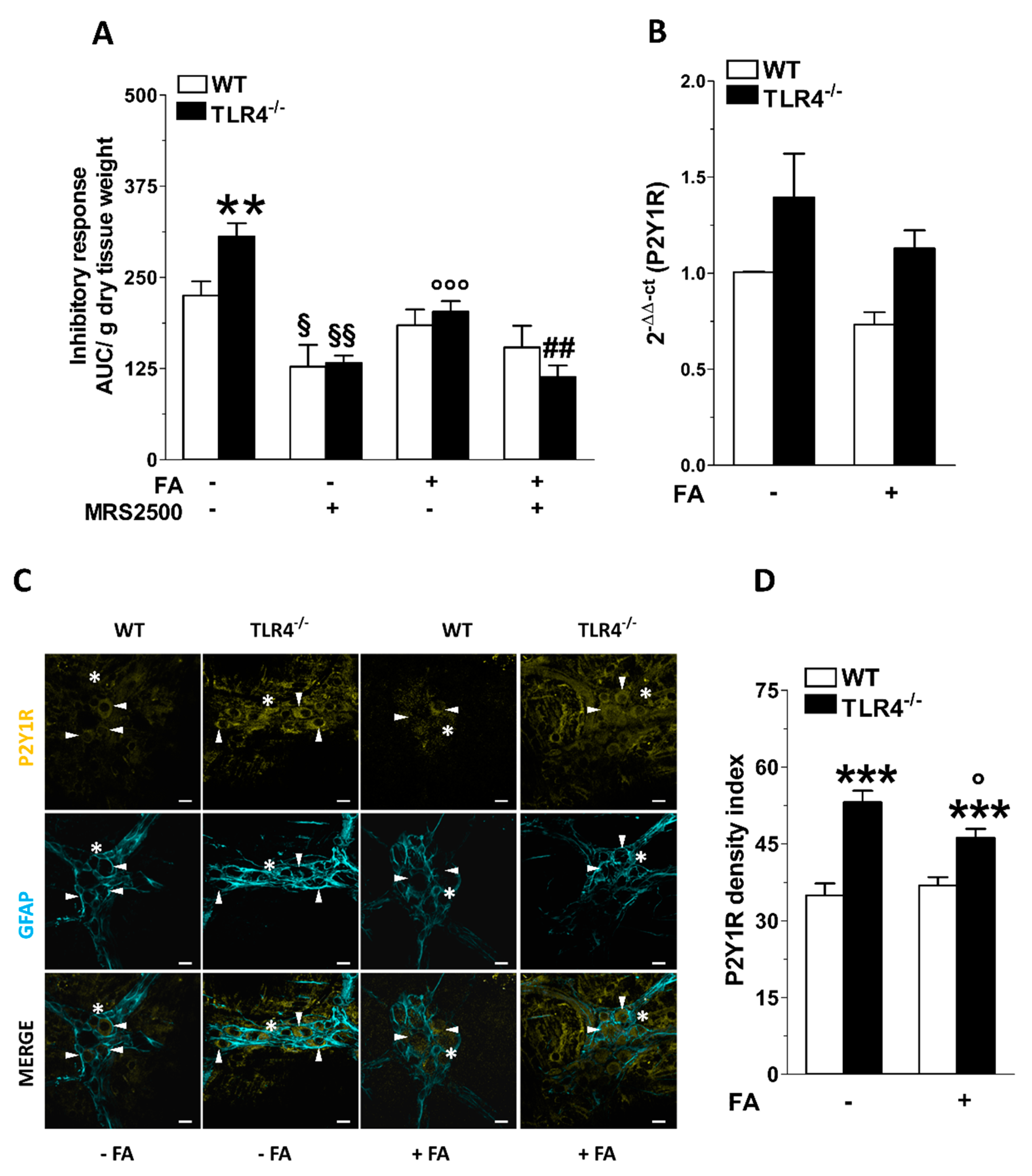
3.3. Inhibition of Enteric Glial Cells (EGC) Metabolism Affects Purinergic-Mediated Inhibitory Response of TLR4−/− Ileal Segments
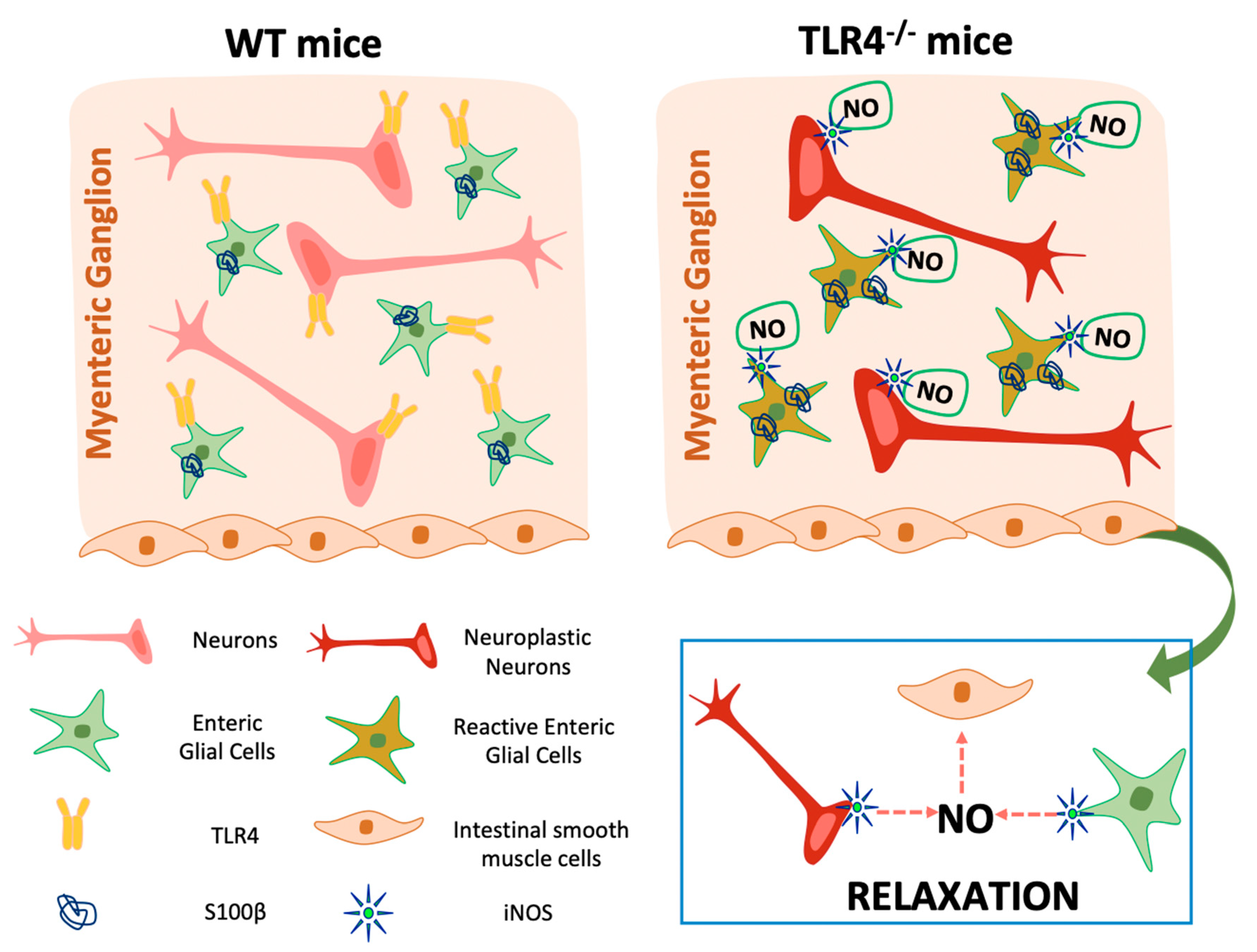
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neunlist, M.; Rolli-Derkinderen, M.; Latorre, R.; Van Landeghem, L.; Coron, E.; Derkinderen, P.; De Giorgio, R. Enteric glial cells: Recent developments and future directions. Gastroenterology 2014, 147, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Capoccia, E.; Cirillo, C.; Gigli, S.; Pesce, M.; D’Alessandro, A.; Cuomo, R.; Sarnelli, G.; Steardo, L.; Esposito, G. Enteric glia: A new player in inflammatory bowel diseases. Int. J. Immunopathol. Pharmacol. 2015, 28, 443–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulbransen, B.D.; Sharkey, K.A. Novel functional roles for enteric glia in the gastrointestinal tract. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.B.; Li, Y.Q. Enteric glial cells and their role in the intestinal epithelial barrier. World J. Gastroenterol. 2014, 20, 11273–11280. [Google Scholar] [CrossRef]
- Ochoa-Cortes, F.; Turco, F.; Linan-Rico, A.; Soghomonyan, S.; Whitaker, E.; Wehner, S.; Cuomo, R.; Christofi, F.L. Enteric Glial Cells: A New Frontier in Neurogastroenterology and Clinical Target for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2016, 22, 433–449. [Google Scholar] [CrossRef]
- Cornet, A.; Savidge, T.C.; Cabarrocas, J.; Deng, W.L.; Colombel, J.F.; Lassmann, H.; Desreumaux, P.; Liblau, R.S. Enterocolitis induced by autoimmune targeting of enteric glial cells: A possible mechanism in Crohn’s disease? Proc. Natl. Acad. Sci. USA 2001, 98, 13306–13311. [Google Scholar] [CrossRef] [Green Version]
- von Boyen, G.B.; Schulte, N.; Pflüger, C.; Spaniol, U.; Hartmann, C.; Steinkamp, M. Distribution of enteric glia and GDNF during gut inflammation. BMC Gastroenterol. 2011, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Wedel, T.; Büsing, V.; Heinrichs, G.; Nohroudi, K.; Bruch, H.P.; Roblick, U.J.; Böttner, M. Diverticular disease is associated with an enteric neuropathy as revealed by morphometric analysis. Neurogastroenterol. Motil. 2010, 22, 407–414. [Google Scholar] [CrossRef]
- Wedel, T.; Krammer, H.J.; Kühnel, W.; Sigge, W. Alterations of the enteric nervous system in neonatal necrotizing enterocolitis revealed by whole-mount immunohistochemistry. Pediatr. Pathol. Lab. Med. 1998, 18, 57–70. [Google Scholar] [CrossRef]
- da Silveira, A.B.; Freitas, M.A.; de Oliveira, E.C.; Neto, S.G.; Luquetti, A.O.; Furness, J.B.; Correa-Oliveira, R.; Reis, D.D. Glial fibrillary acidic protein and S-100 colocalization in the enteroglial cells in dilated and nondilated portions of colon from chagasic patients. Hum. Pathol. 2009, 40, 244–251. [Google Scholar] [CrossRef]
- Stenkamp-Strahm, C.M.; Kappmeyer, A.J.; Schmalz, J.T.; Gericke, M.; Balemba, O. High-fat diet ingestion correlates with neuropathy in the duodenum myenteric plexus of obese mice with symptoms of type 2 diabetes. Cell Tissue Res. 2013, 354, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Devos, D.; Lebouvier, T.; Lardeux, B.; Biraud, M.; Rouaud, T.; Pouclet, H.; Coron, E.; Bruley des Varannes, S.; Naveilhan, P.; Nguyen, J.M.; et al. Colonic inflammation in Parkinson’s disease. Neurobiol. Dis. 2013, 50, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Ferri, G.L.; Probert, L.; Cocchia, D.; Michetti, F.; Marangos, P.J.; Polak, J.M. Evidence for the presence of S-100 protein in the glial component of the human enteric nervous system. Nature 1982, 297, 409–410. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R. Glial cells in the enteric nervous system contain glial fibrillary acidic protein. Nature 1980, 286, 736–737. [Google Scholar] [CrossRef]
- Laranjeira, C.; Pachnis, V. Enteric nervous system development: Recent progress and future challenges. Auton. Neurosci. 2009, 151, 61–69. [Google Scholar] [CrossRef]
- Boesmans, W.; Lasrado, R.; Vanden Berghe, P.; Pachnis, V. Heterogeneity and phenotypic plasticity of glial cells in the mammalian enteric nervous system. Glia 2015, 63, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Shaffer, S.M.; Dunagin, M.C.; Torborg, S.R.; Torre, E.A.; Emert, B.; Krepler, C.; Beqiri, M.; Sproesser, K.; Brafford, P.A.; Xiao, M.; et al. Rare cell variability and drug-induced reprogramming as a mode of cancer drug resistance. Nature 2017, 546, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Burda, J.E.; Radulovic, M.; Yoon, H.; Scarisbrick, I.A. Critical role for PAR1 in kallikrein 6-mediated oligodendrogliopathy. Glia 2013, 61, 1456–1470. [Google Scholar] [CrossRef]
- Brown, I.A.; McClain, J.L.; Watson, R.E.; Patel, B.A.; Gulbransen, B.D. Enteric glia mediate neuron death in colitis through purinergic pathways that require connexin-43 and nitric oxide. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Delvalle, N.M.; Fried, D.E.; Rivera-Lopez, G.; Gaudette, L.; Gulbransen, B.D. Cholinergic activation of enteric glia is a physiological mechanism that contributes to the regulation of gastrointestinal motility. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G473–G483. [Google Scholar] [CrossRef] [PubMed]
- Fonnum, F.; Johnsen, A.; Hassel, B. Use of fluorocitrate and fluoroacetate in the study of brain metabolism. Glia 1997, 21, 106–113. [Google Scholar] [CrossRef]
- Nasser, Y.; Fernandez, E.; Keenan, C.M.; Ho, W.; Oland, L.D.; Tibbles, L.A.; Schemann, M.; MacNaughton, W.K.; Rühl, A.; Sharkey, K.A. Role of enteric glia in intestinal physiology: Effects of the gliotoxin fluorocitrate on motor and secretory function. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G912–G927. [Google Scholar] [CrossRef] [PubMed]
- MacEachern, S.J.; Patel, B.A.; Keenan, C.M.; Dicay, M.; Chapman, K.; McCafferty, D.M.; Savidge, T.C.; Beck, P.L.; MacNaughton, W.K.; Sharkey, K.A. Inhibiting inducible nitric oxide synthase in enteric glia restores electrogenic ion transport in mice with colitis. Gastroenterology 2015, 149, 445–455.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubišić, V.; Gulbransen, B.D. Enteric glia: The most alimentary of all glia. J. Physiol. 2017, 595, 557–570. [Google Scholar]
- McClain, J.L.; Gulbransen, B.D. The acute inhibition of enteric glial metabolism with fluoroacetate alters calcium signaling, hemichannel function, and the expression of key proteins. J. Neurophysiol. 2017, 117, 365–375. [Google Scholar] [CrossRef]
- Anitha, M.; Vijay-Kumar, M.; Sitaraman, S.V.; Gewirtz, A.T.; Srinivasan, S. Gut microbial products regulate murine gastrointestinal motility via Toll-like receptor 4 signaling. Gastroenterology 2012, 143, 1006.e6–1016.e4. [Google Scholar] [CrossRef] [Green Version]
- Turco, F.; Sarnelli, G.; Cirillo, C.; Palumbo, I.; De Giorgi, F.; D’Alessandro, A.; Cammarota, M.; Giuliano, M.; Cuomo, R. Enteroglial-derived S100B protein integrates bacteria-induced Toll-like receptor signalling in human enteric glial cells. Gut 2014, 63, 105–115. [Google Scholar] [CrossRef]
- Caputi, V.; Marsilio, I.; Cerantola, S.; Roozfarakh, M.; Lante, I.; Galuppini, F.; Rugge, M.; Napoli, E.; Giulivi, C.; Orso, G.; et al. Toll-Like Receptor 4 Modulates Small Intestine Neuromuscular Function through Nitrergic and Purinergic Pathways. Front. Pharmacol. 2017, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- McGrath, J.C.; Lilley, E. Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [Green Version]
- Curtis, M.J.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Giembycz, M.A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. Experimental design and analysis and their reporting II: Updated and simplified guidance for authors and peer reviewers. Br. J. Pharmacol. 2018, 175, 987–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoppellaro, C.; Bin, A.; Brun, P.; Banzato, S.; Macchi, V.; Castagliuolo, I.; Giron, M.C. Adenosine-mediated enteric neuromuscular function is affected during herpes simplex virus type 1 infection of rat enteric nervous system. PLoS ONE 2013, 8, e72648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputi, V.; Marsilio, I.; Filpa, V.; Cerantola, S.; Orso, G.; Bistoletti, M.; Paccagnella, N.; De Martin, S.; Montopoli, M.; Dall’Acqua, S.; et al. Antibiotic-induced dysbiosis of the microbiota impairs gut neuromuscular function in juvenile mice. Br. J. Pharmacol. 2017, 174, 3623–3639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, P.; Giron, M.C.; Qesari, M.; Porzionato, A.; Caputi, V.; Zoppellaro, C.; Banzato, S.; Grillo, A.R.; Spagnol, L.; De Caro, R.; et al. Toll-like receptor 2 regulates intestinal inflammation by controlling integrity of the enteric nervous system. Gastroenterology 2013, 145, 1323–1333. [Google Scholar] [CrossRef] [Green Version]
- Filpa, V.; Bistoletti, M.; Caon, I.; Moro, E.; Grimaldi, A.; Moretto, P.; Baj, A.; Giron, M.C.; Karousou, E.; Viola, M.; et al. Changes in hyaluronan deposition in the rat myenteric plexus after experimentally-induced colitis. Sci. Rep. 2017, 7, 17644. [Google Scholar] [CrossRef]
- Bistoletti, M.; Caputi, V.; Baranzini, N.; Marchesi, N.; Filpa, V.; Marsilio, I.; Cerantola, S.; Terova, G.; Baj, A.; Grimaldi, A.; et al. Antibiotic treatment-induced dysbiosis differently affects BDNF and TrkB expression in the brain and in the gut of juvenile mice. PLoS ONE 2019, 14, e0212856. [Google Scholar] [CrossRef] [Green Version]
- Zulato, E.; Ciccarese, F.; Agnusdei, V.; Pinazza, M.; Nardo, G.; Iorio, E.; Curtarello, M.; Silic-Benussi, M.; Rossi, E.; Venturoli, C.; et al. LKB1 loss is associated with glutathione deficiency under oxidative stress and sensitivity of cancer cells to cytotoxic drugs and γ-irradiation. Biochem. Pharmacol. 2018, 156, 479–490. [Google Scholar] [CrossRef]
- Paquola, A.; Mañé, N.; Giron, M.C.; Jimenez, M. Diadenosine tetraphosphate activates P2Y1 receptors that cause smooth muscle relaxation in the mouse colon. Eur. J. Pharmacol. 2019, 855, 160–166. [Google Scholar] [CrossRef]
- Scemes, E.; Duval, N.; Meda, P. Reduced expression of P2Y1 receptors in connexin43-null mice alters calcium signaling and migration of neural progenitor cells. J. Neurosci. 2003, 23, 11444–11452. [Google Scholar] [CrossRef] [Green Version]
- Laranjeira, C.; Sandgren, K.; Kessaris, N.; Richardson, W.; Potocnik, A.; Vanden Berghe, P.; Pachnis, V. Glial cells in the mouse enteric nervous system can undergo neurogenesis in response to injury. J. Clin. Invest. 2011, 121, 3412–3424. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.; Ganz, J.; Bayrer, J.; Becker, L.; Bogunovic, M.; Rao, M. Advances in Enteric Neurobiology: The “Brain” in the Gut in Health and Disease. J. Neurosci. 2018, 38, 9346–9354. [Google Scholar] [CrossRef] [Green Version]
- Bódi, N.; Szalai, Z.; Bagyánszki, M. Nitrergic Enteric Neurons in Health and Disease-Focus on Animal Models. Int. J. Mol. Sci. 2019, 20, 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Zhu, B.; Xue, X.D. Role of neuronal nitric oxide synthase and inducible nitric oxide synthase in intestinal injury in neonatal rats. World J. Gastroenterol. 2006, 12, 4364–4368. [Google Scholar] [CrossRef] [PubMed]
- Eskandari, M.K.; Kalff, J.C.; Billiar, T.R.; Lee, K.K.; Bauer, A.J. LPS-induced muscularis macrophage nitric oxide suppresses rat jejunal circular muscle activity. Am. J. Physiol. 1999, 277, G478–G486. [Google Scholar] [CrossRef]
- Seago, N.D.; Thompson, J.H.; Zhang, X.J.; Eloby-Childress, S.; Sadowska-Krowicka, H.; Rossi, J.L.; Currie, M.G.; Manning, P.T.; Clark, D.A.; Miller, M.J. Inducible nitric oxide synthase and guinea-pig ileitis induced by adjuvant. Mediators Inflamm. 1995, 4, 19–24. [Google Scholar] [CrossRef]
- Green, C.L.; Ho, W.; Sharkey, K.A.; McKay, D.M. Dextran sodium sulfate-induced colitis reveals nicotinic modulation of ion transport via iNOS-derived NO. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G706–G714. [Google Scholar] [CrossRef] [Green Version]
- Aubé, A.C.; Cabarrocas, J.; Bauer, J.; Philippe, D.; Aubert, P.; Doulay, F.; Liblau, R.; Galmiche, J.P.; Neunlist, M. Changes in enteric neurone phenotype and intestinal functions in a transgenic mouse model of enteric glia disruption. Gut 2006, 55, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Mañé, N.; Gil, V.; Martínez-Cutillas, M.; Clavé, P.; Gallego, D.; Jiménez, M. Differential functional role of purinergic and nitrergic inhibitory cotransmitters in human colonic relaxation. Acta Physiol. (Oxf). 2014, 212, 293–305. [Google Scholar] [CrossRef]
- Giaroni, C. Purinergic signalling and development of the autonomic nervous system. Auton. Neurosci. 2015, 191, 67–77. [Google Scholar] [CrossRef]
- Franke, H.; Krügel, U.; Schmidt, R.; Grosche, J.; Reichenbach, A.; Illes, P. P2 receptor-types involved in astrogliosis in vivo. Br. J. Pharmacol. 2001, 134, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Grasa, L.; Gil, V.; Gallego, D.; Martín, M.T.; Jiménez, M. P2Y(1) receptors mediate inhibitory neuromuscular transmission in the rat colon. Br. J. Pharmacol. 2009, 158, 1641–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClain, J.; Grubišić, V.; Fried, D.; Gomez-Suarez, R.A.; Leinninger, G.M.; Sévigny, J.; Parpura, V.; Gulbransen, B.D. Ca2+ responses in enteric glia are mediated by connexin-43 hemichannels and modulate colonic transit in mice. Gastroenterology 2014, 146, 497.e1–507.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zizzo, M.G.; Mastropaolo, M.; Grählert, J.; Mulè, F.; Serio, R. Pharmacological characterization of uracil nucleotide-preferring P2Y receptors modulating intestinal motility: A study on mouse ileum. Purinergic Signal 2012, 8, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Incà, R.; Paccagnella, M.; Cardin, R.; Pathak, S.; Baldo, V.; Giron, M.C.; Sturniolo, G.C. 5-ASA colonic mucosal concentrations resulting from different pharmaceutical formulations in ulcerative colitis. World J. Gastroenterol. 2013, 19, 5665–5670. [Google Scholar] [CrossRef] [PubMed]
- Gulbransen, B.D.; Sharkey, K.A. Purinergic neuron-to-glia signaling in the enteric nervous system. Gastroenterology 2009, 136, 1349–1358. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host Species | Dilution | Catalog Number | Source |
|---|---|---|---|---|
| Primary Antisera (Clone) | ||||
| HuC/D (16A11) | Mouse biotin-conjugated | 1:100 | A-21272 | Thermo Fisher Scientific (Monza, Italy) |
| nNOS (polyclonal) | Rabbit | 1:100 | 61-700 | Thermo Fisher Scientific |
| GFAP (polyclonal) | Chicken | 1:100 | ab4674 | Abcam (Cambridge, UK) |
| S100β (EP1576Y) | Rabbit | 1:50 | ab52642 | Abcam |
| iNOS (polyclonal) | Rabbit | 1:100 | sc-8310 | Santa Cruz Biotechnology (Heidelberg, Germany) |
| P2Y1 (polyclonal) | Rabbit | 1:50 | APR-009 | Alomone Labs (Jerusalem, Israel) |
| connexin-43 (polyclonal) | Rabbit | 1:50 | ACC-201 | Alomone Labs |
| SOX10 (EPR4007) | Rabbit | 1:50 | ab155279 | Abcam |
| Secondary Antisera | ||||
| Goat anti-rabbit IgG Alexa 488-conjugated | - | 1:1000 | A-11008 | Thermo Fisher Scientific |
| Goat anti-chicken IgY Alexa 555-conjugated | - | 1:1000 | A-11039 | Thermo Fisher Scientific |
| Streptavidin Alexa 555-conjugated | - | 1:1000 | S21381 | Thermo Fisher Scientific |
| Gene | Sequence 5′–3′ |
|---|---|
| HuC/D | F-AAGAGTCCCCTGTCGCTCA R-TACACGAAGATGCACCAGCC |
| S100β | F-GACTCCAGCAGCAAAGGTGA R-ATCTTCGTCCAGCGTCTCCA |
| iNOS | F-CAGCTGGGCTGTACAAACCTT R-CATTGGAAGTGAAGCGTTTCG |
| nNOS | F-GTGGCCATCGTGTCCTACCATAC R-GTTTCGAGGCAGGTGGAAGCTA |
| P2Y1R | F-AGTGTGTGCCACCTGAGTGA R-ACCCTTGAGCTTGGAATGGAAT |
| β-actin | F-TGACAGGATGCAGAAGGAGA R-TAGAGCCACCAATCCACACA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerantola, S.; Caputi, V.; Marsilio, I.; Ridolfi, M.; Faggin, S.; Bistoletti, M.; Giaroni, C.; Giron, M.C. Involvement of Enteric Glia in Small Intestine Neuromuscular Dysfunction of Toll-Like Receptor 4-Deficient Mice. Cells 2020, 9, 838. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9040838
Cerantola S, Caputi V, Marsilio I, Ridolfi M, Faggin S, Bistoletti M, Giaroni C, Giron MC. Involvement of Enteric Glia in Small Intestine Neuromuscular Dysfunction of Toll-Like Receptor 4-Deficient Mice. Cells. 2020; 9(4):838. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9040838
Chicago/Turabian StyleCerantola, Silvia, Valentina Caputi, Ilaria Marsilio, Manuela Ridolfi, Sofia Faggin, Michela Bistoletti, Cristina Giaroni, and Maria Cecilia Giron. 2020. "Involvement of Enteric Glia in Small Intestine Neuromuscular Dysfunction of Toll-Like Receptor 4-Deficient Mice" Cells 9, no. 4: 838. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9040838