Y-Box Binding Protein 1 Expression in Trophoblast Cells Promotes Fetal and Placental Development

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Experimental Design and Sample Collection
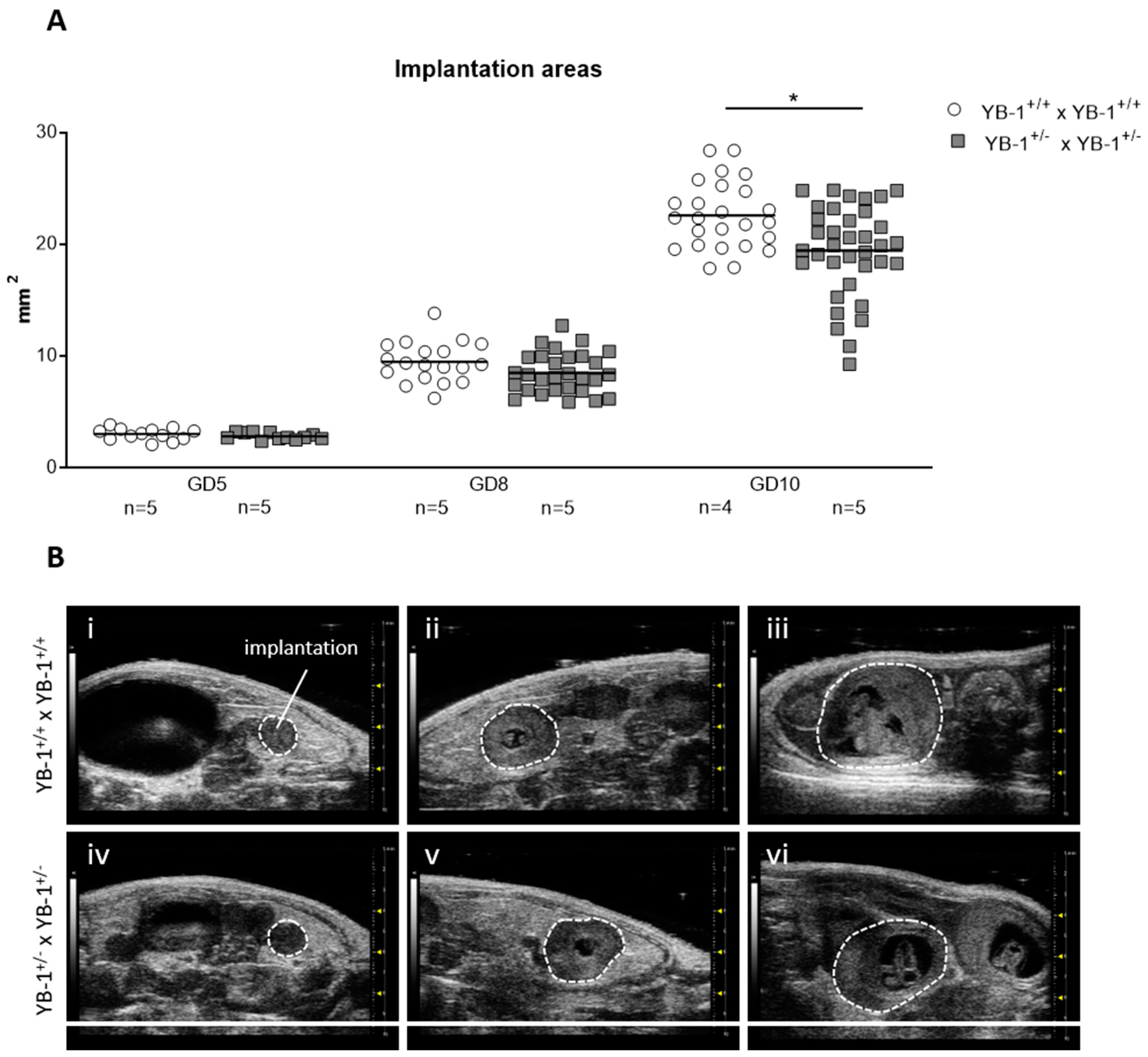
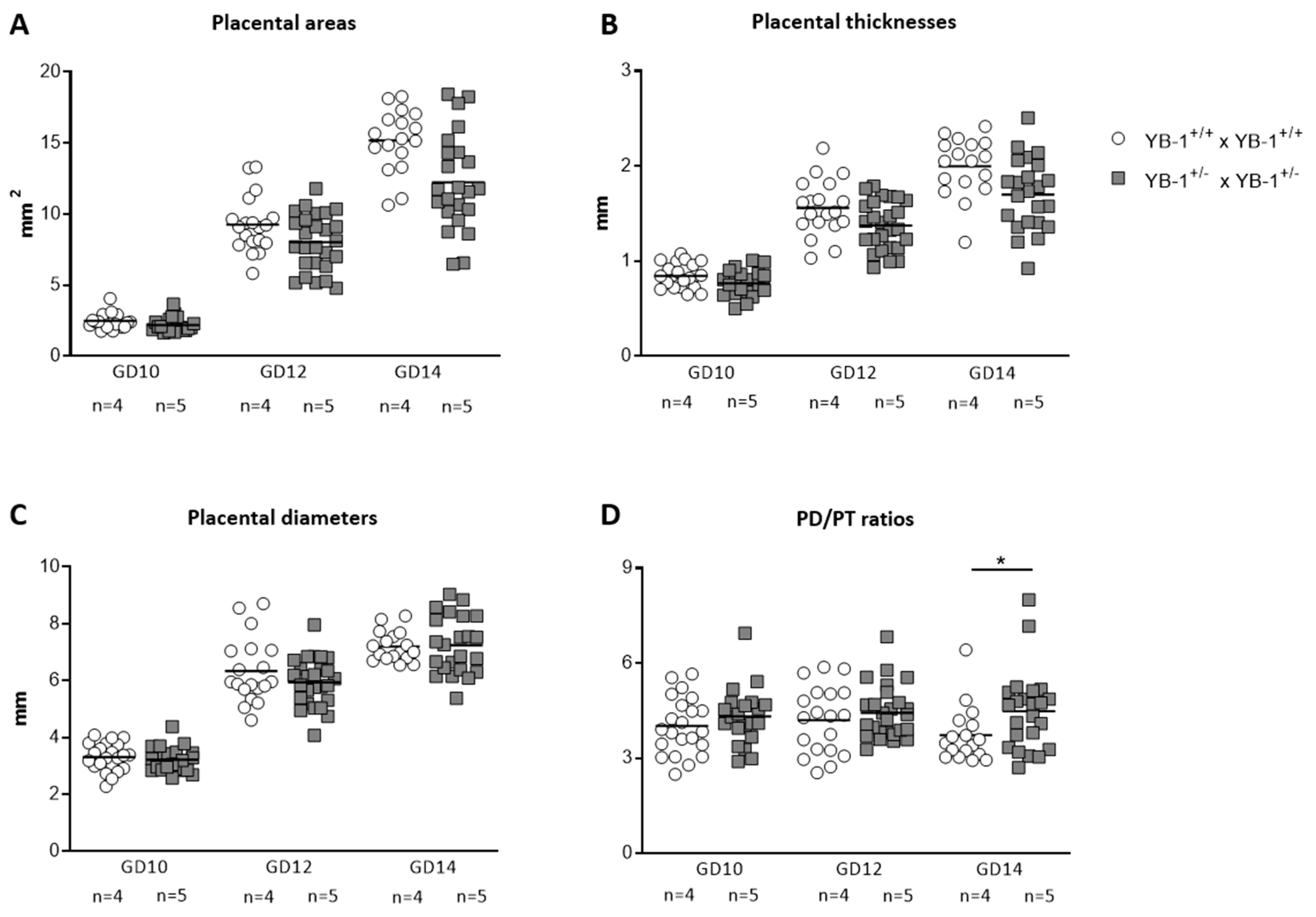
2.3. High-Frequency Ultrasound Measurement
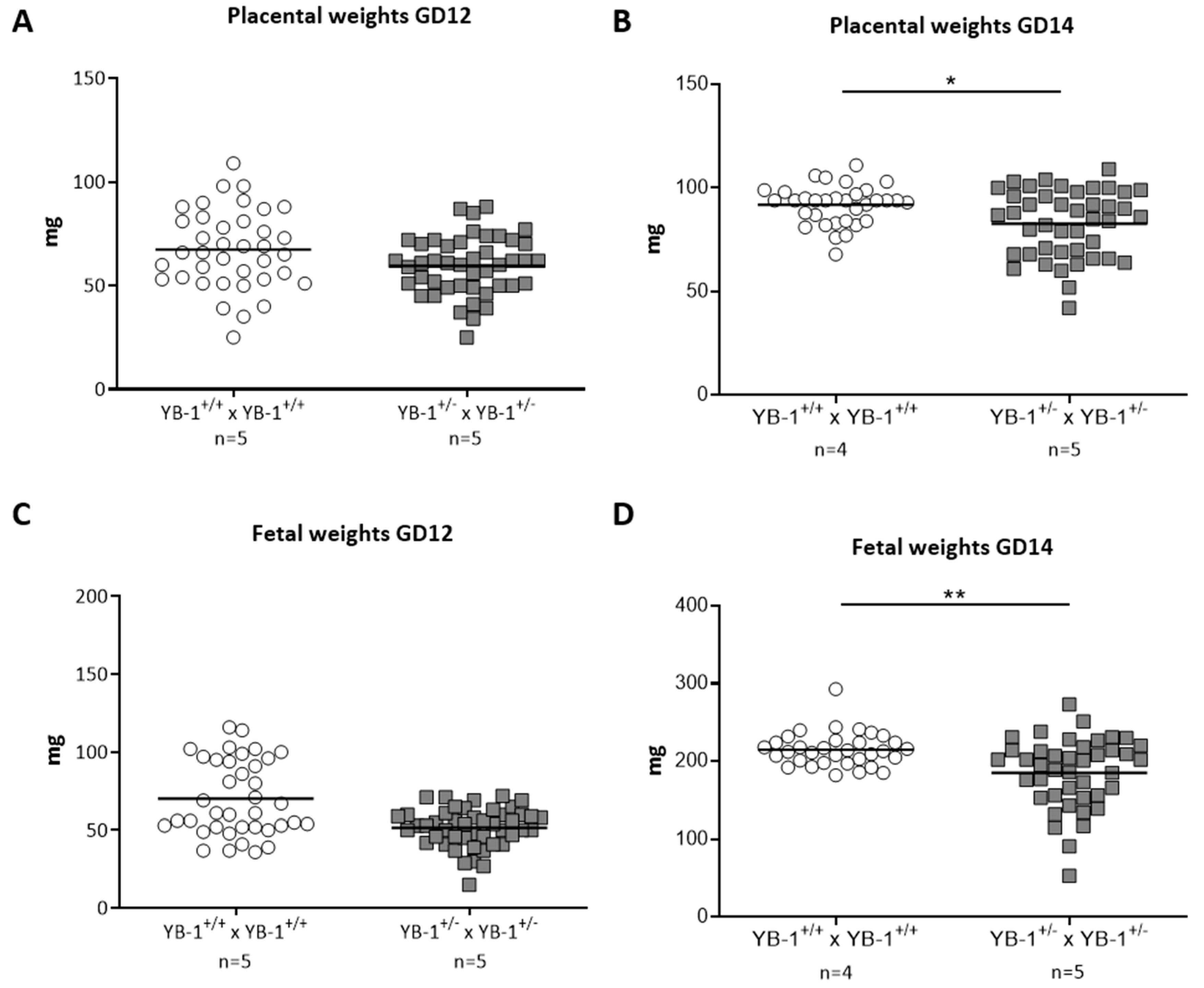
2.4. Measurement of Fetal and Placental Weights
2.5. Histology
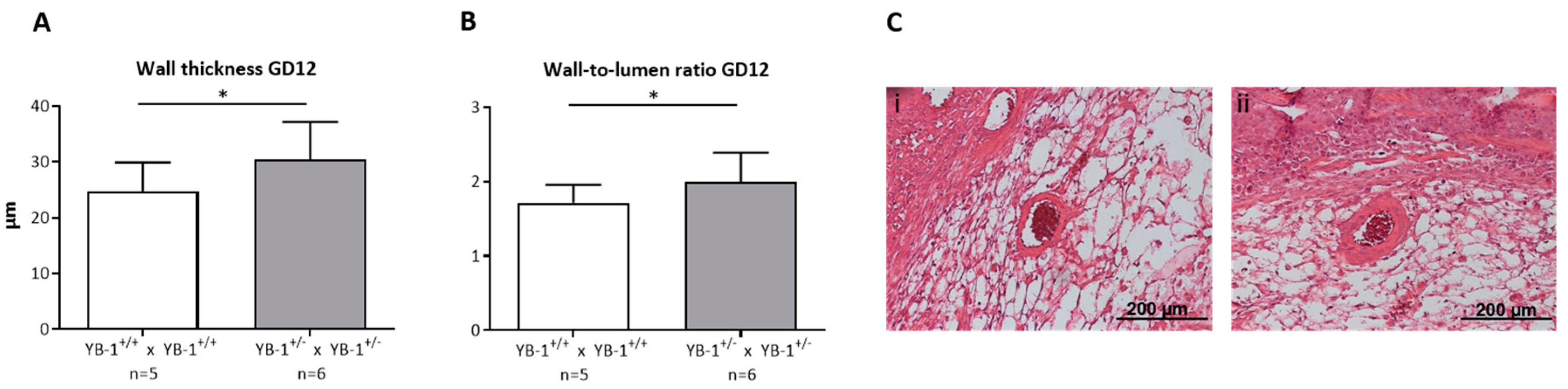
2.6. Spiral Artery Analysis
2.7. Statistics
3. Results
3.1. YB-1 Deficiency Negatively Affected Implantation and Fetal Growth
3.2. YB-1 Deficiency Altered Fetal and Placental Parameters
3.3. Placental YB-1 Deficiency Resulted in Impaired Remodeling of Uterine Spiral Arteries without Affecting Uterine Artery Flow Parameters
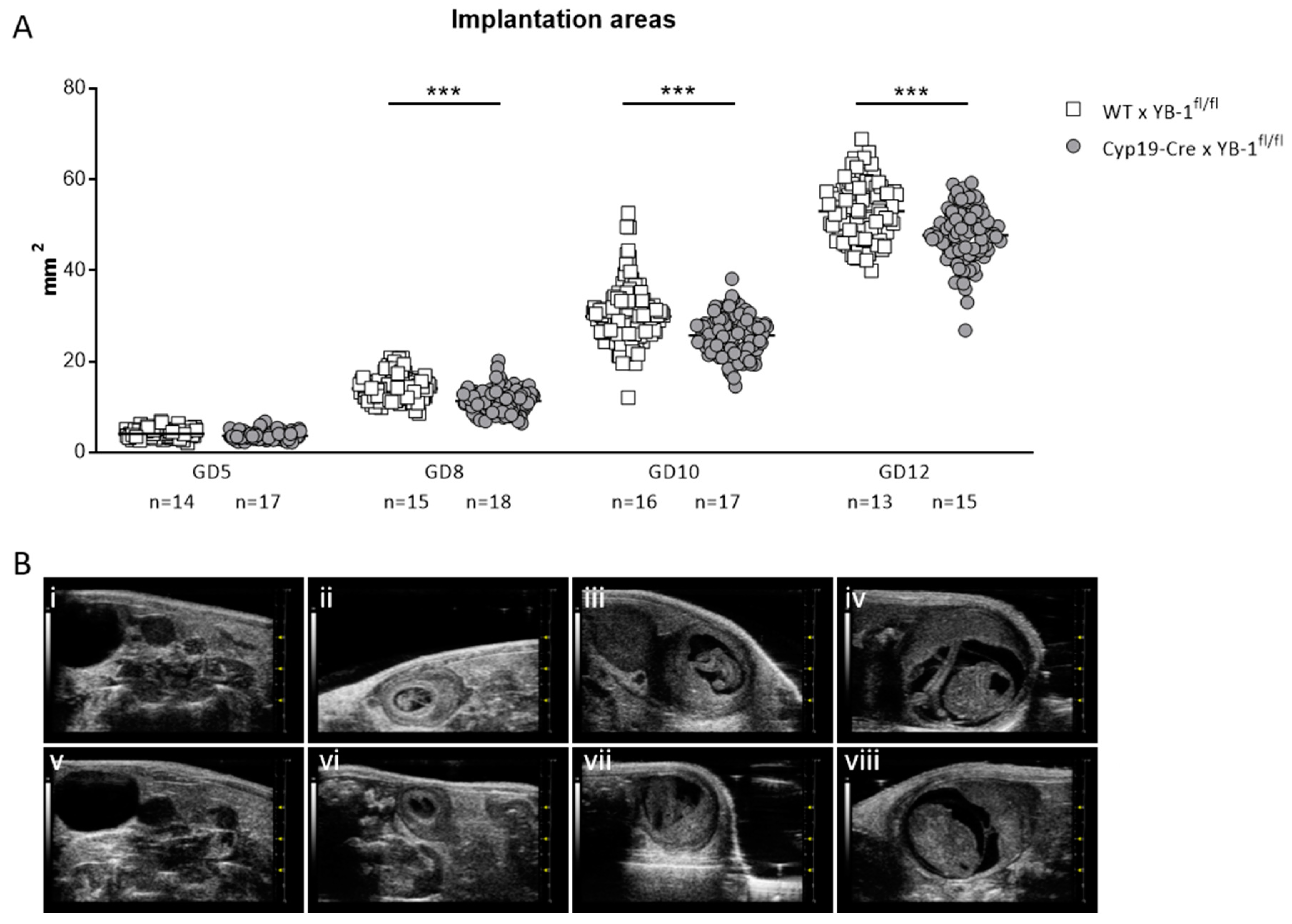
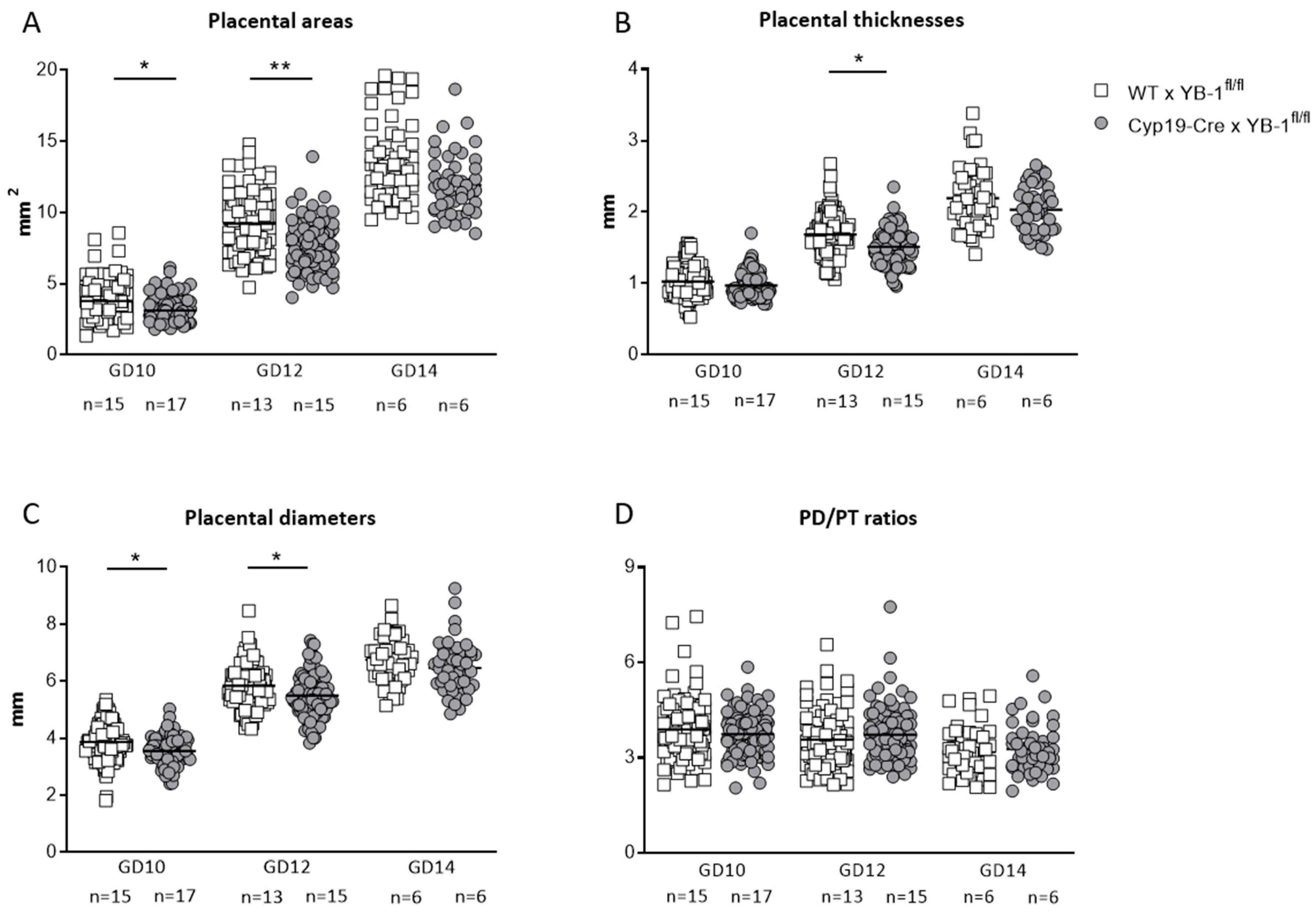
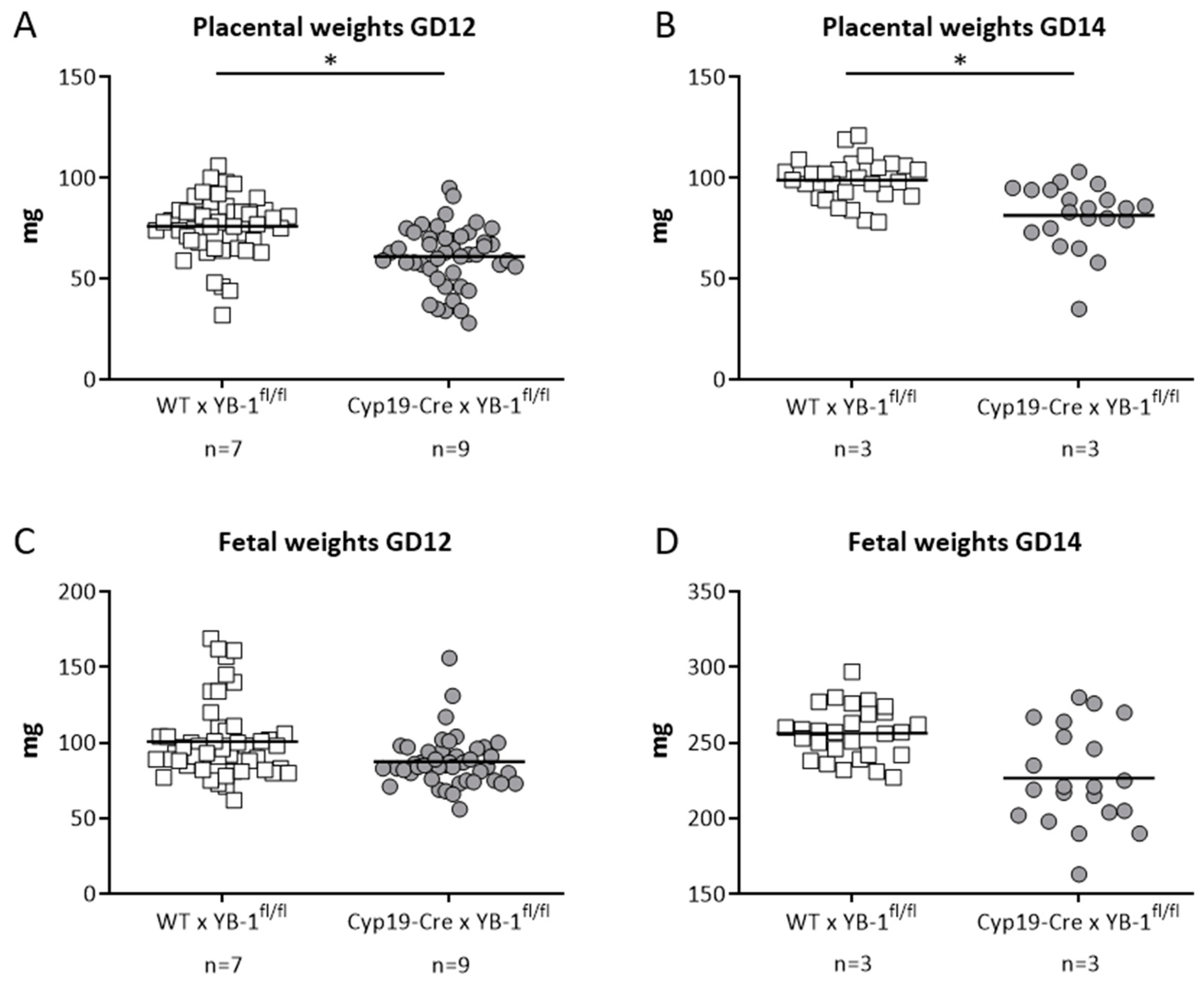
3.4. Trophoblast-Specific YB-1 Deficiency Negatively Affected Fetal and Placental Parameters with a Phenotype of Decreased Placental Weight
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindquist, J.A.; Mertens, P.R. Cold shock proteins: From cellular mechanisms to pathophysiology and disease. Cell Commun. Signal 2018, 16, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frye, B.C.; Halfter, S.; Djudjaj, S.; Muehlenberg, P.; Weber, S.; Raffetseder, U.; En-Nia, A.; Knott, H.; Baron, J.M.; Dooley, S.; et al. Y-box protein-1 is actively secreted through a non-classical pathway and acts as an extracellular mitogen. EMBO Rep. 2009, 10, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Guarino, A.M.; Troiano, A.; Pizzo, E.; Bosso, A.; Vivo, M.; Pinto, G.; Amoresano, A.; Pollice, A.; La Mantia, G.; Calabro, V. Oxidative Stress Causes Enhanced Secretion of YB-1 Protein that Restrains Proliferation of Receiving Cells. Genes 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, E.J.; Kim, Y.J. What is fetal programming? A lifetime health is under the control of in utero health. Obstet. Gynecol. Sci. 2017, 60, 506–519. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Budkina, K.S.; Zlobin, N.E.; Kononova, S.V.; Ovchinnikov, L.P.; Babakov, A.V. Cold Shock Domain Proteins: Structure and Interaction with Nucleic Acids. Biochemistry (Moscow) 2020, 85, S1–S19. [Google Scholar] [CrossRef]
- Shibahara, K.; Uchiumi, T.; Fukuda, T.; Kura, S.; Tominaga, Y.; Maehara, Y.; Kohno, K.; Nakabeppu, Y.; Tsuzuki, T.; Kuwano, M. Targeted disruption of one allele of the Y-box binding protein-1 (YB-1) gene in mouse embryonic stem cells and increased sensitivity to cisplatin and mitomycin C. Cancer Sci. 2004, 95, 348–353. [Google Scholar] [CrossRef]
- Uchiumi, T.; Fotovati, A.; Sasaguri, T.; Shibahara, K.; Shimada, T.; Fukuda, T.; Nakamura, T.; Izumi, H.; Tsuzuki, T.; Kuwano, M.; et al. YB-1 is important for an early stage embryonic development: Neural tube formation and cell proliferation. J. Biol. Chem. 2006, 281, 40440–40449. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.H.; Books, J.T.; Ley, T.J. YB-1 is important for late-stage embryonic development, optimal cellular stress responses, and the prevention of premature senescence. Mol. Cell Biol. 2005, 25, 4625–4637. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.H.; Books, J.T.; Ley, T.J. Cold shock domain family members YB-1 and MSY4 share essential functions during murine embryogenesis. Mol. Cell Biol. 2006, 26, 8410–8417. [Google Scholar] [CrossRef] [Green Version]
- Basaki, Y.; Taguchi, K.; Izumi, H.; Murakami, Y.; Kubo, T.; Hosoi, F.; Watari, K.; Nakano, K.; Kawaguchi, H.; Ohno, S.; et al. Y-box binding protein-1 (YB-1) promotes cell cycle progression through CDC6-dependent pathway in human cancer cells. Eur. J. Cancer 2010, 46, 954–965. [Google Scholar] [CrossRef] [PubMed]
- Maurya, P.K.; Mishra, A.; Yadav, B.S.; Singh, S.; Kumar, P.; Chaudhary, A.; Srivastava, S.; Murugesan, S.N.; Mani, A. Role of Y Box Protein-1 in cancer: As potential biomarker and novel therapeutic target. J. Cancer 2017, 8, 1900–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astanehe, A.; Finkbeiner, M.R.; Hojabrpour, P.; To, K.; Fotovati, A.; Shadeo, A.; Stratford, A.L.; Lam, W.L.; Berquin, I.M.; Duronio, V.; et al. The transcriptional induction of PIK3CA in tumor cells is dependent on the oncoprotein Y-box binding protein-1. Oncogene 2009, 28, 2406–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.P.; Shyamasundar, S.; Gunaratne, J.; Scully, O.J.; Matsumoto, K.; Bay, B.H. YBX1 gene silencing inhibits migratory and invasive potential via CORO1C in breast cancer in vitro. BMC Cancer 2017, 17, 201. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, M.; Rancso, C.; Stuhmer, T.; Eckstein, N.; Andrulis, M.; Gerecke, C.; Lorentz, H.; Royer, H.D.; Bargou, R.C. The Y-box binding protein YB-1 is associated with progressive disease and mediates survival and drug resistance in multiple myeloma. Blood 2008, 111, 3714–3722. [Google Scholar] [CrossRef]
- Lee, C.; Dhillon, J.; Wang, M.Y.; Gao, Y.; Hu, K.; Park, E.; Astanehe, A.; Hung, M.C.; Eirew, P.; Eaves, C.J.; et al. Targeting YB-1 in HER-2 overexpressing breast cancer cells induces apoptosis via the mTOR/STAT3 pathway and suppresses tumor growth in mice. Cancer Res. 2008, 68, 8661–8666. [Google Scholar] [CrossRef] [Green Version]
- Mertens, P.R.; Alfonso-Jaume, M.A.; Steinmann, K.; Lovett, D.H. A synergistic interaction of transcription factors AP2 and YB-1 regulates gelatinase A enhancer-dependent transcription. J. Biol. Chem. 1998, 273, 32957–32965. [Google Scholar] [CrossRef] [Green Version]
- Lovett, D.H.; Cheng, S.; Cape, L.; Pollock, A.S.; Mertens, P.R. YB-1 alters MT1-MMP trafficking and stimulates MCF-7 breast tumor invasion and metastasis. Biochem. Biophys. Res. Commun. 2010, 398, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Ding, R.; Liu, X.M.; Xiang, Y.Q.; Zhang, Y.; Zhang, J.Y.; Guo, F.; Liu, Z.W.; Fan, J.X. Altered Matrix Metalloproteinases Expression in Placenta from Patients with Gestational Diabetes Mellitus. Chin. Med. J. 2018, 131, 1255–1258. [Google Scholar] [CrossRef]
- Hiden, U.; Ghaffari-Tabrizi, N.; Gauster, M.; Tam-Amersdorfer, C.; Cetin, I.; Dieber-Rotheneder, M.; Lang, U.; Desoye, G. Membrane-type matrix metalloproteinase 1 regulates trophoblast functions and is reduced in fetal growth restriction. Am. J. Pathol. 2013, 182, 1563–1571. [Google Scholar] [CrossRef]
- Meyer, N.; Woidacki, K.; Knofler, M.; Meinhardt, G.; Nowak, D.; Velicky, P.; Pollheimer, J.; Zenclussen, A.C. Chymase-producing cells of the innate immune system are required for decidual vascular remodeling and fetal growth. Sci. Rep. 2017, 7, 45106. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.; Schuler, T.; Zenclussen, A.C. Simultaneous Ablation of Uterine Natural Killer Cells and Uterine Mast Cells in Mice Leads to Poor Vascularization and Abnormal Doppler Measurements That Compromise Fetal Well-being. Front Immunol. 2017, 8, 1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, N.; Schuler, T.; Zenclussen, A.C. High Frequency Ultrasound for the Analysis of Fetal and Placental Development In Vivo. J. Vis. Exp. 2018, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitley, G.S.; Cartwright, J.E. Cellular and molecular regulation of spiral artery remodelling: Lessons from the cardiovascular field. Placenta 2010, 31, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.R.; Bettencourt, M.; Alho, I.; Costa, A.L.; Sousa, A.R.; Mansinho, A.; Abreu, C.; Pulido, C.; Macedo, D.; Vendrell, I.; et al. Serum YB-1 (Y-box binding protein 1) as a biomarker of bone disease progression in patients with breast cancer and bone metastases. J. Bone Oncol. 2017, 6, 16–21. [Google Scholar] [CrossRef]
- Johnson, T.G.; Schelch, K.; Mehta, S.; Burgess, A.; Reid, G. Why Be One Protein When You Can Affect Many? The Multiple Roles of YB-1 in Lung Cancer and Mesothelioma. Front Cell Dev. Biol. 2019, 7, 221. [Google Scholar] [CrossRef] [Green Version]
- Lasham, A.; Print, C.G.; Woolley, A.G.; Dunn, S.E.; Braithwaite, A.W. YB-1: Oncoprotein, prognostic marker and therapeutic target? Biochem. J. 2013, 449, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Hanssen, L.; Alidousty, C.; Djudjaj, S.; Frye, B.C.; Rauen, T.; Boor, P.; Mertens, P.R.; van Roeyen, C.R.; Tacke, F.; Heymann, F.; et al. YB-1 is an early and central mediator of bacterial and sterile inflammation in vivo. J. Immunol. 2013, 191, 2604–2613. [Google Scholar] [CrossRef]
- Raffetseder, U.; Liehn, E.A.; Weber, C.; Mertens, P.R. Role of cold shock Y-box protein-1 in inflammation, atherosclerosis and organ transplant rejection. Eur. J. Cell Biol. 2012, 91, 567–575. [Google Scholar] [CrossRef]
- Wang, J.; Djudjaj, S.; Gibbert, L.; Lennartz, V.; Breitkopf, D.M.; Rauen, T.; Hermert, D.; Martin, I.V.; Boor, P.; Braun, G.S.; et al. YB-1 orchestrates onset and resolution of renal inflammation via IL10 gene regulation. J. Cell Mol. Med. 2017, 21, 3494–3505. [Google Scholar] [CrossRef] [Green Version]
- Almasry, S.M.; Elfayomy, A.K. Morphometric analysis of terminal villi and gross morphological changes in the placentae of term idiopathic intrauterine growth restriction. Tissue Cell 2012, 44, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Egbor, M.; Ansari, T.; Morris, N.; Green, C.J.; Sibbons, P.D. Pre-eclampsia and fetal growth restriction: How morphometrically different is the placenta? Placenta 2006, 27, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, T.M.; Manwani, R.; Ohadike, C.; Wijesekara, J.; Baker, P.N. The placenta in pre-eclampsia and intrauterine growth restriction: Studies on exchange surface areas, diffusion distances and villous membrane diffusive conductances. Placenta 2007, 28, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Salafia, C.M.; Zhang, J.; Charles, A.K.; Bresnahan, M.; Shrout, P.; Sun, W.; Maas, E.M. Placental characteristics and birthweight. Paediatr. Perinat. Epidemiol. 2008, 22, 229–239. [Google Scholar] [CrossRef]
- Salavati, N.; Sovio, U.; Mayo, R.P.; Charnock-Jones, D.S.; Smith, G.C. The relationship between human placental morphometry and ultrasonic measurements of utero-placental blood flow and fetal growth. Placenta 2016, 38, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.; Zenclussen, A.C. Immune Cells in the Uterine Remodeling: Are They the Target of Endocrine Disrupting Chemicals? Front Immunol. 2020, 11, 246. [Google Scholar] [CrossRef]
- Wenzel, P.L.; Leone, G. Expression of Cre recombinase in early diploid trophoblast cells of the mouse placenta. Genesis 2007, 45, 129–134. [Google Scholar] [CrossRef]
- Lu, J.; Wu, W.; Xin, Q.; Zhou, C.; Wang, J.; Ni, Z.; Liu, D.; Xu, Y.; Yu, Y.; Yang, N.; et al. Spatiotemporal coordination of trophoblast and allantoic Rbpj signaling directs normal placental morphogenesis. Cell Death Dis. 2019, 10, 438. [Google Scholar] [CrossRef]
- Ouseph, M.M.; Li, J.; Chen, H.Z.; Pecot, T.; Wenzel, P.; Thompson, J.C.; Comstock, G.; Chokshi, V.; Byrne, M.; Forde, B.; et al. Atypical E2F repressors and activators coordinate placental development. Dev. Cell 2012, 22, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Shawber, C.J.; Brown-Grant, D.A.; Wu, T.; Kitajewski, J.K.; Douglas, N.C. Dominant-negative inhibition of canonical Notch signaling in trophoblast cells does not disrupt placenta formation. Biol. Open 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.L.; Wu, L.; de Bruin, A.; Chong, J.L.; Chen, W.Y.; Dureska, G.; Sites, E.; Pan, T.; Sharma, A.; Huang, K.; et al. Rb is critical in a mammalian tissue stem cell population. Genes Dev. 2007, 21, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.L.; Hsiao, E.Y.; Yan, Z.; Mazmanian, S.K.; Patterson, P.H. The placental interleukin-6 signaling controls fetal brain development and behavior. Brain Behav. Immun. 2017, 62, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauen, T.; Raffetseder, U.; Frye, B.C.; Djudjaj, S.; Muhlenberg, P.J.; Eitner, F.; Lendahl, U.; Bernhagen, J.; Dooley, S.; Mertens, P.R. YB-1 acts as a ligand for Notch-3 receptors and modulates receptor activation. J. Biol. Chem. 2009, 284, 26928–26940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.-X.; Zhuang, X.; Huang, T.-T.; Feng, R.; Lin, J.-H. Effects of Notch2 and Notch3 on Cell Proliferation and Apoptosis of Trophoblast Cell Lines. Int. J. Med. Sci. 2015, 12, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.-X.; Wu, Z.-M.; Liu, W.; Lin, J.-H. Notch2 and Notch3 suppress the proliferation and mediate invasion of trophoblast cell lines. Biol. Open 2017, 6, 1123–1129. [Google Scholar] [CrossRef] [Green Version]
- West, R.C.; Bouma, G.J.; Winger, Q.A. Shifting perspectives from “oncogenic” to oncofetal proteins; how these factors drive placental development. Reprod. Biol. Endocrinol. 2018, 16, 101. [Google Scholar] [CrossRef]
- Kosnopfel, C.; Sinnberg, T.; Schittek, B. Y-box binding protein 1--a prognostic marker and target in tumour therapy. Eur. J. Cell Biol. 2014, 93, 61–70. [Google Scholar] [CrossRef]
- DeLoia, J.A.; Burlingame, J.M.; Krasnow, J.S. Differential expression of G1 cyclins during human placentogenesis. Placenta 1997, 18, 9–16. [Google Scholar] [CrossRef]
- Fock, V.; Plessl, K.; Fuchs, R.; Dekan, S.; Milla, S.K.; Haider, S.; Fiala, C.; Knofler, M.; Pollheimer, J. Trophoblast subtype-specific EGFR/ERBB4 expression correlates with cell cycle progression and hyperplasia in complete hydatidiform moles. Hum. Reprod. 2015, 30, 789–799. [Google Scholar] [CrossRef]
- Ferretti, C.; Bruni, L.; Dangles-Marie, V.; Pecking, A.P.; Bellet, D. Molecular circuits shared by placental and cancer cells, and their implications in the proliferative, invasive and migratory capacities of trophoblasts. Hum. Reprod. Update 2007, 13, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhao, T.; Duan, E. Matrix metalloproteinases (MMPs) and trophoblast invasion. Chin. Sci. Bulletin 2005, 50, 1169–1173. [Google Scholar] [CrossRef]
- Gopal, S.K.; Greening, D.W.; Mathias, R.A.; Ji, H.; Rai, A.; Chen, M.; Zhu, H.-J.; Simpson, R.J. YBX1/YB-1 induces partial EMT and tumourigenicity through secretion of angiogenic factors into the extracellular microenvironment. Oncotarget 2015, 6, 13718–13730. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | YB-1+/+ × YB-1+/+ | YB-1+/− × YB-1+/− |
|---|---|---|
| Number of implantations | 10 (3–12) | 10 (9–11) |
| Number of abortions | 2 (0–3) | 1 (0–3) |
| Abortion rate (%) | 18.13 (0.00–100) | 10.56 (10–27.27) |
| Measurement | WT × YB-1fl/fl | Cyp19-Cre × YB-1fl/fl |
|---|---|---|
| Number of implantations | 11 (1–13) | 9 (2–14) |
| Number of abortions | 0 (0–3) | 1 (0–3) |
| Abortion rate (%) | 0.00 (0.00–23.08) | 11.11 (0.00–50.00) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, N.; Schumacher, A.; Coenen, U.; Woidacki, K.; Schmidt, H.; Lindquist, J.A.; Mertens, P.R.; Zenclussen, A.C. Y-Box Binding Protein 1 Expression in Trophoblast Cells Promotes Fetal and Placental Development. Cells 2020, 9, 1942. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091942
Meyer N, Schumacher A, Coenen U, Woidacki K, Schmidt H, Lindquist JA, Mertens PR, Zenclussen AC. Y-Box Binding Protein 1 Expression in Trophoblast Cells Promotes Fetal and Placental Development. Cells. 2020; 9(9):1942. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091942
Chicago/Turabian StyleMeyer, Nicole, Anne Schumacher, Urs Coenen, Katja Woidacki, Hannah Schmidt, Jonathan A. Lindquist, Peter R. Mertens, and Ana C. Zenclussen. 2020. "Y-Box Binding Protein 1 Expression in Trophoblast Cells Promotes Fetal and Placental Development" Cells 9, no. 9: 1942. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091942