The Expressions of TSLP, IL-33, and IL-17A in Monocyte Derived Dendritic Cells from Asthma and COPD Patients are Related to Epithelial–Macrophage Interactions

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Overall Study Design
2.2. Patient Characteristics
2.3. Monocyte Derived Dendritic Cell (moDC) and Monocyte Derived Macrophages (moMφ) Culture
2.4. The Culture of Epithelial Cells in Air Liquid Interface (ALI) and Triple Co-Culture
2.5. RNA Isolation, cDNA Synthesis, and Real Time PCR
2.6. Flow Cytometry Analysis
2.7. Statistical Analysis
3. Results
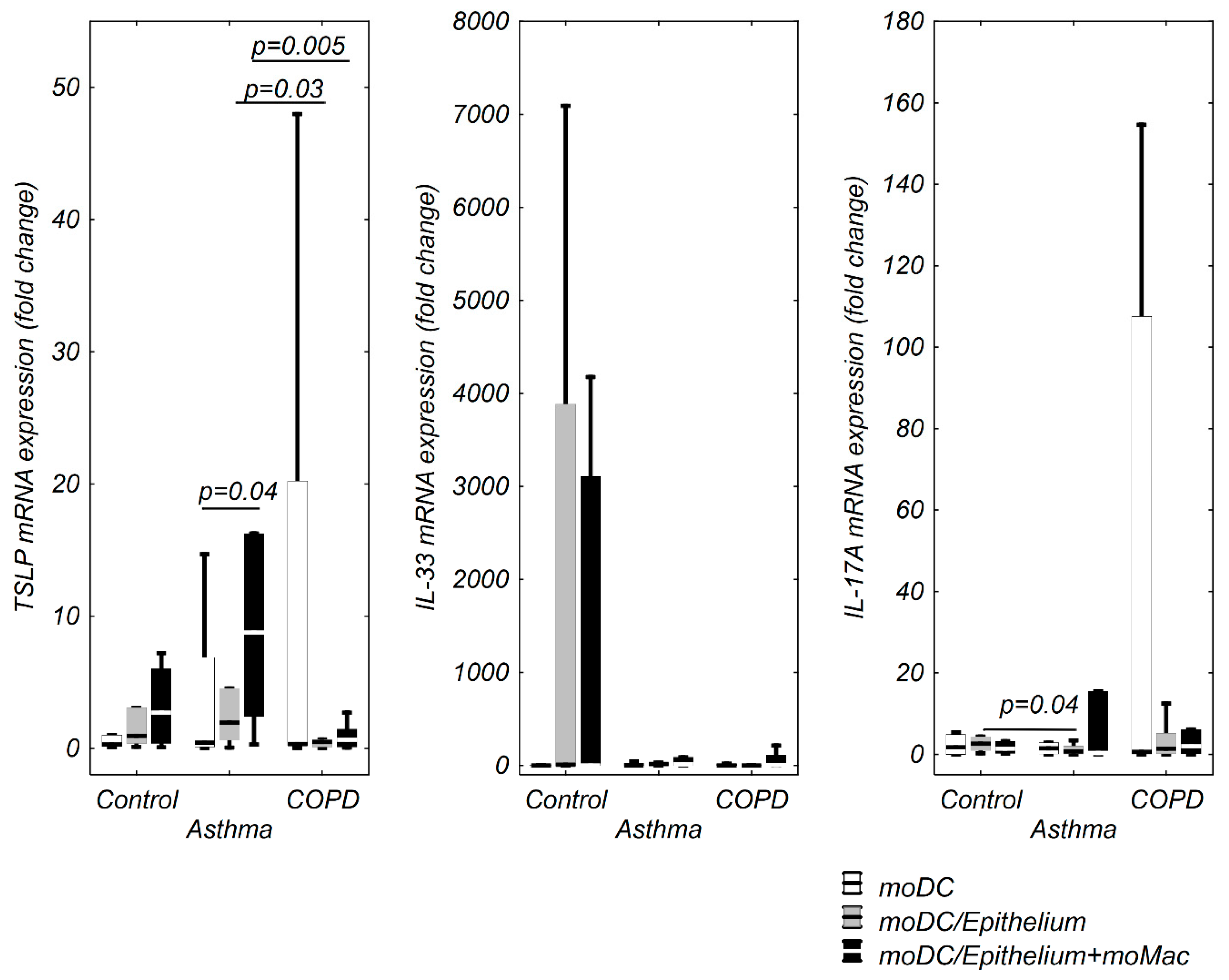
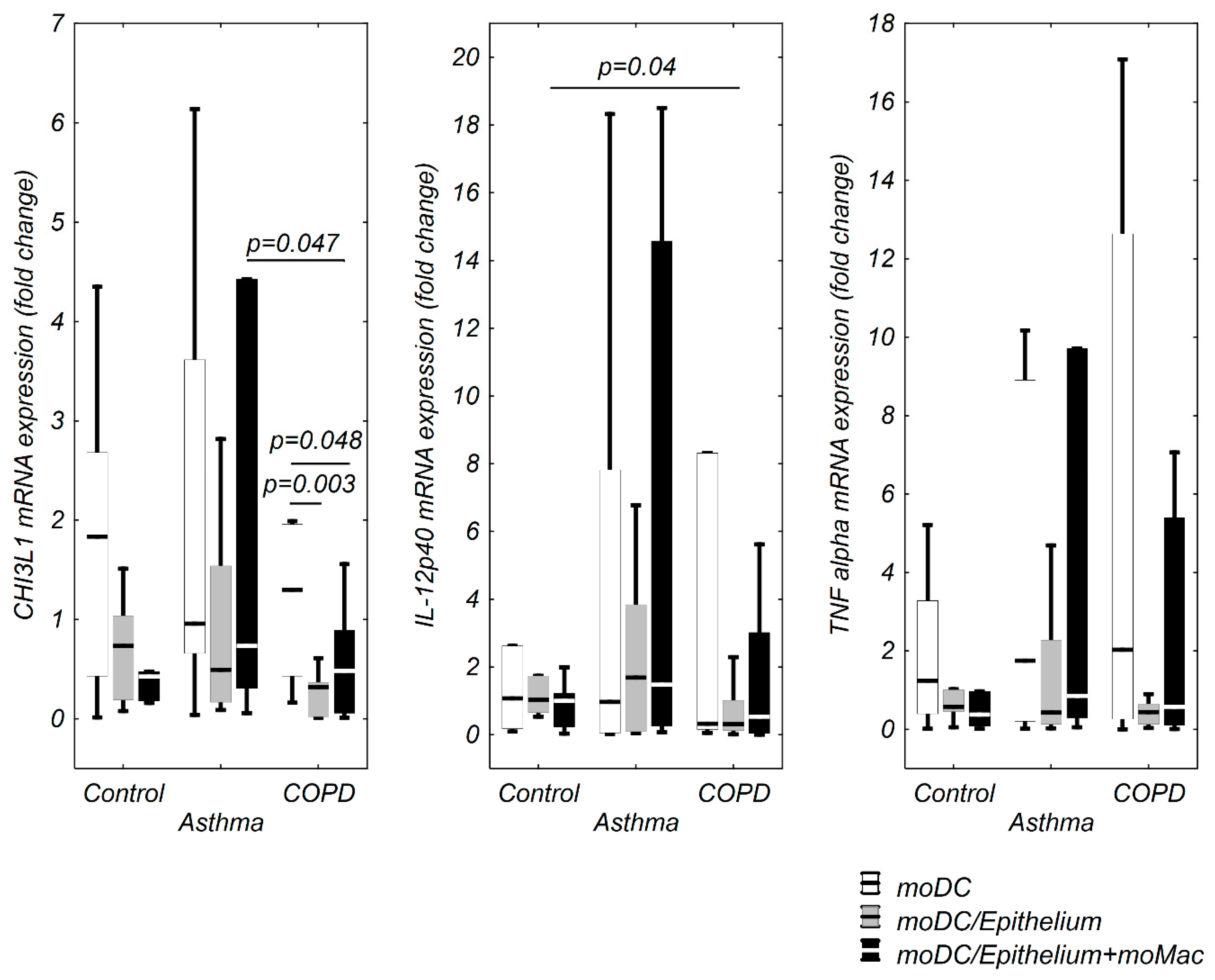
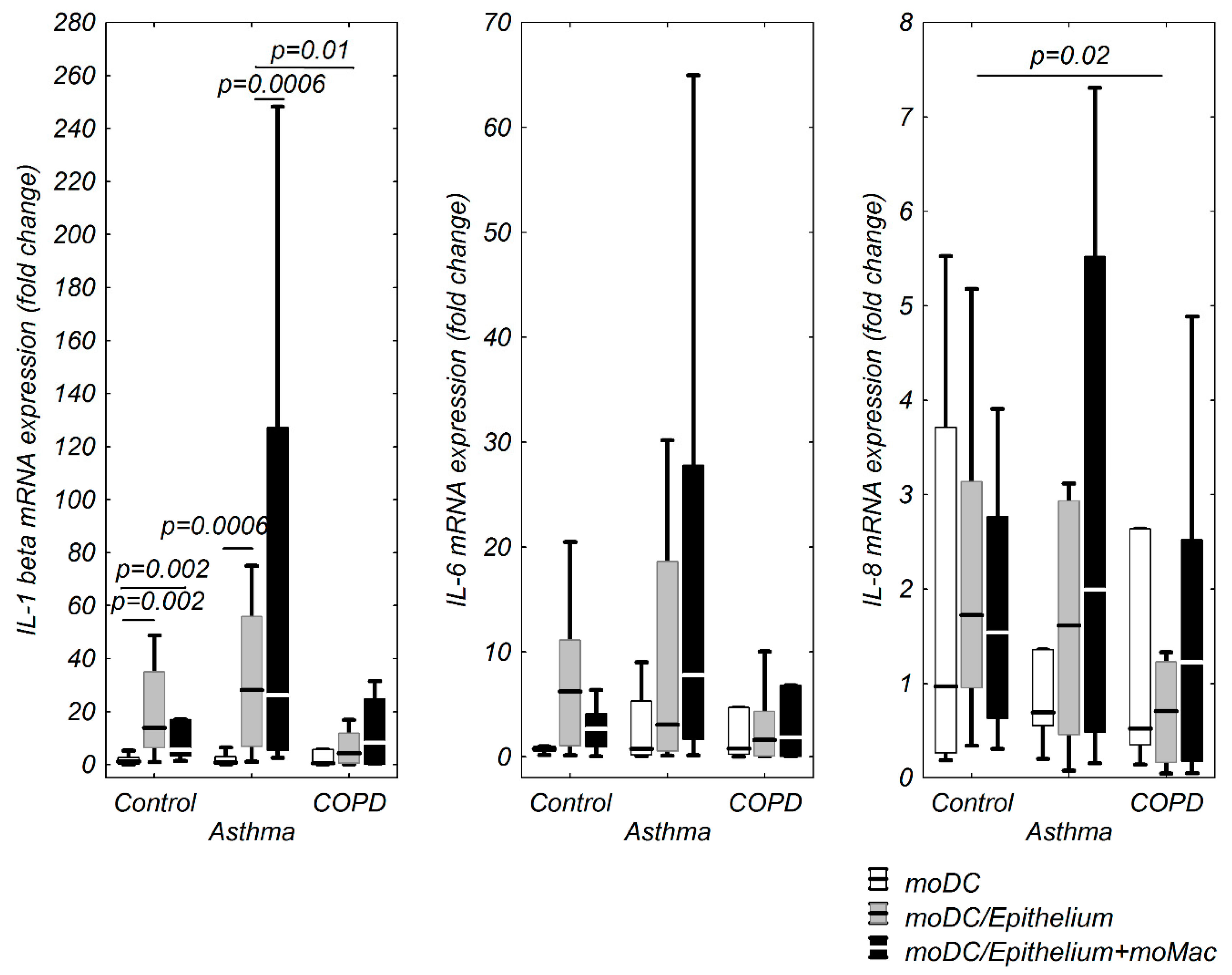
3.1. moDCs TSLP, IL-33, and IL-17A mRNA Expression in Multi Co-Cultures in Control Group
3.2. moDCs TSLP, IL-33, and IL-17A mRNA Expression in Multi Co-Cultures in the Asthma Group
3.3. TSLP, IL-33, and IL-17A mRNA Expression in moDCs in Multi Co-Culture Schemes in the COPD Group
3.4. The Impact of BMI on TSLP, IL-33, and IL-17A mRNA Expression in moDCs
3.5. The Expression of TSLPR, ST2, and IL-17RA on moDCs in Various Co-Cultures from Control, Asthma, and COPD Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kool, M.; Lambrecht, B.N. Dendritic cells in asthma and COPD: Opportunities for drug development. Curr. Opin. Immunol. 2007, 19, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. Dendritic cells and epithelial cells: Linking innate and adaptive immunity in asthma. Nat. Rev. Immunol. 2008, 8, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Whitsett, J.A.; Alenghat, T. Respiratory epithelial cells orchestrate pulmonary innate immunity. Nat. Immunol. 2015, 16, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, D.; Prince, A. Innate immunity in the respiratory epithelium. Am. J. Respir. Cell Mol. Biol. 2011, 45, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Kool, M.; Willart, M.A.M.; Van Nimwegen, M.; Bergen, I.; Pouliot, P.; Virchow, J.C.; Rogers, N.; Osorio, F.; Reis e Sousa, C.; Reis, E.; et al. An unexpected role for uric acid as an inducer of T helper 2 cell immunity to inhaled antigens and inflammatory mediator of allergic asthma. Immunity 2011, 34, 527–540. [Google Scholar] [CrossRef]
- Divekar, R.; Kita, H. Recent advances in epithelium-derived cytokines (IL-33, IL-25, and thymic stromal lymphopoietin) and allergic inflammation. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.G.; Gugilla, A.; Mukherjee, M.; Merim, K.; Irshad, A.; Tang, W.; Kinoshita, T.; Watson, B.; Oliveria, J.-P.; Comeau, M.; et al. Thymic stromal lymphopoietin and IL-33 modulate migration of hematopoietic progenitor cells in patients with allergic asthma. J. Allergy Clin. Immunol. 2015, 135, 1594–1602. [Google Scholar] [CrossRef]
- Walter, M.J.; Kajiwara, N.; Karanja, P.; Castro, M.; Holtzman, M.J. Interleukin 12 p40 production by barrier epithelial cells during airway inflammation. J. Exp. Med. 2001, 193, 339–351. [Google Scholar] [CrossRef]
- Park, J.-A.; Drazen, J.M.; Tschumperlin, D.J. The chitinase-like protein YKL-40 is secreted by airway epithelial cells at base line and in response to compressive mechanical stress. J. Biol. Chem. 2010, 285, 29817–29825. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-J.; Nakagawa, T.; Kitamura, H.; Atsumi, T.; Kamon, H.; Sawa, S.-I.; Kamimura, D.; Ueda, N.; Iwakura, Y.; Ishihara, K.; et al. IL-6 regulates in vivo dendritic cell differentiation through STAT3 activation. J. Immunol. 2004, 173, 3844–3854. [Google Scholar] [CrossRef] [Green Version]
- Elder, M.J.; Webster, S.J.; Williams, D.L.; Gaston, J.S.H.; Goodall, J.C. TSLP production by dendritic cells is modulated by IL-1β and components of the endoplasmic reticulum stress response. Eur. J. Immunol. 2016, 46, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obregon, C.; Rothen-Rutishauser, B.; Gerber, P.; Gehr, P.; Nicod, L.P. Active uptake of dendritic cell-derived exovesicles by epithelial cells induces the release of inflammatory mediators through a TNF-alpha-mediated pathway. Am. J. Pathol. 2009, 175, 696–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, N.; Hanabuchi, S.; Soumelis, V.; Yuan, W.; Ho, S.; de Waal Malefyt, R.; Liu, Y.-J. Human thymic stromal lymphopoietin promotes dendritic cell-mediated CD4+ T cell homeostatic expansion. Nat. Immunol. 2004, 5, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Kitajima M, Lee HC, Nakayama T, Ziegler SF. TSLP enhances the function of helper type 2 cells. Eur. J. Immunol. 2011, 41, 1862–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Wang, Y.-H.; Duramad, O.; Hori, T.; Delespesse, G.J.; Watanabe, N.; Qin, F.X.-F.; Yao, Z.; Cao, W.; Liu, Y.-J. TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J. Exp. Med. 2005, 202, 1213–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, M.J.; Webster, S.J.; Fitzmaurice, T.J.; Shaunak, A.S.D.; Steinmetz, M.; Chee, R.; Mallat, Z.; Cohen, E.S.; Williams, D.L.; Gaston, J.S.H.; et al. Dendritic Cell-Derived TSLP Negatively Regulates HIF-1α and IL-1β During Dectin-1 Signaling. Front. Immunol. 2019, 10, 921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spadoni, I.; Iliev, I.D.; Rossi, G.; Rescigno, M. Dendritic cells produce TSLP that limits the differentiation of Th17 cells, fosters Treg development, and protects against colitis. Mucosal Immunol. 2012, 5, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Besnard, A.-G.; Togbe, D.; Guillou, N.; Erard, F.; Quesniaux, V.; Ryffel, B. IL-33-activated dendritic cells are critical for allergic airway inflammation. Eur. J. Immunol. 2011, 41, 1675–1686. [Google Scholar] [CrossRef]
- Rank, M.A.; Kobayashi, T.; Kozaki, H.; Bartemes, K.R.; Squillace, D.L.; Kita, H. IL-33-activated dendritic cells induce an atypical TH2-type response. J. Allergy Clin. Immunol. 2009, 123, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Lin, J.; Lu, F.; Zhang, X.; Zhang, L.; Gandhi, N.B.; de Paiva, C.S.; Pflugfelder, S.C.; Li, D.-Q. Potential autocrine regulation of interleukin-33/ST2 signaling of dendritic cells in allergic inflammation. Mucosal Immunol. 2013, 6, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Salvatore, G.; Bernoud-Hubac, N.; Bissay, N.; Debard, C.; Daira, P.; Meugnier, E.; Proamer, F.; Hanau, D.; Vidal, H.; Aricò, M.; et al. Human monocyte-derived dendritic cells turn into foamy dendritic cells with IL-17A. J. Lipid Res. 2015, 56, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Strategy for Asthma Management and Prevention. 2018. Available online: www.ginasthma.org (accessed on 30 March 2018).
- Global Strategy for the Diagnosis, Management and Prevention of COPD, Global Initiative for Chronic Obstructive Lung Disease (GOLD). 2018. Available online: http://goldcopd.org/ (accessed on 17 July 2018).
- Paplinska-Goryca, M.; Misiukiewicz-Stepien, P.; Nejman-Gryz, P.; Proboszcz, M.; Mlacki, M.; Gorska, K.; Krenke, R. Epithelial-macrophage-dendritic cell interactions impact alarmins expression in asthma and COPD. Clin. Immunol. 2020, 215, 108421. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; O’Connor, B.; Ratoff, J.; Meng, Q.; Mallett, K.; Cousins, D.; Robinson, D.; Zhang, G.; Zhao, J.; Lee, T.H.; et al. Thymic stromal lymphopoietin expression is increased in asthmatic airways and correlates with expression of Th2-attracting chemokines and disease severity. J. Immunol. 2005, 174, 8183–8190. [Google Scholar] [CrossRef] [PubMed]
- Isaksen, D.E.; Baumann, H.; Zhou, B.; Nivollet, S.; Farr, A.G.; Levin, S.D.; Ziegler, S.F. Uncoupling of proliferation and Stat5 activation in thymic stromal lymphopoietin-mediated signal transduction. J. Immunol. 2002, 168, 3288–3294. [Google Scholar] [CrossRef]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyata, M.; Ohba, T.; Ando, T.; Hatsushika, K.; Suenaga, F.; Shimokawa, N.; Ohnuma, Y.; Katoh, R.; Ogawa, H.; et al. Cigarette smoke extract induces thymic stromal lymphopoietin expression, leading to T(H)2-type immune responses and airway inflammation. J. Allergy Clin. Immunol. 2008, 122, 1208–1214. [Google Scholar] [CrossRef]
- Rate, A.; Bosco, A.; McKenna, K.L.; Holt, P.G.; Upham, J.W. Airway epithelial cells condition dendritic cells to express multiple immune surveillance genes. PLoS ONE 2012, 7, e44941. [Google Scholar] [CrossRef] [Green Version]
- Arellano-Orden, E.; Calero-Acuña, C.; Moreno-Mata, N.; Gómez-Izquierdo, L.; Sánchez-López, V.; López-Ramírez, C.; Tobar, D.; López-Villalobos, J.L.; Gutiérrez, C.; Blanco-Orozco, A.; et al. Cigarette Smoke Decreases the Maturation of Lung Myeloid Dendritic Cells. PLoS ONE 2016, 11, e0152737. [Google Scholar] [CrossRef] [Green Version]
- Vassallo, R.; Kroening, P.R.; Parambil, J.; Kita, H. Nicotine and oxidative cigarette smoke constituents induce immune-modulatory and pro-inflammatory dendritic cell responses. Mol. Immunol. 2008, 45, 3321–3329. [Google Scholar] [CrossRef] [Green Version]
- Givi, M.E.; Peck, M.J.; Boon, L.; Mortaz, E. The role of dendritic cells in the pathogenesis of cigarette smoke-induced emphysema in mice. Eur. J. Pharmacol. 2013, 721, 259–266. [Google Scholar] [CrossRef]
- Freeman, C.M.; Martinez, F.J.; Han, M.K.; Ames, T.M.; Chensue, S.W.; Todt, J.C.; Arenberg, D.A.; Meldrum, C.A.; Getty, C.; McCloskey, L.; et al. Lung dendritic cell expression of maturation molecules increases with worsening chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2009, 180, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzalone, G.; Albano, G.D.; Montalbano, A.M.; Riccobono, L.; Bonanno, A.; Gagliardo, R.; Bucchieri, F.; Marchese, R.; Moscato, M.; Profita, M. IL-17A-associated IKK-α signaling induced TSLP production in epithelial cells of COPD patients. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, H.; Hida, N.; Masuko, H.; Sakamoto, T.; Hizawa, N. Effects of Lung Function-Related Genes and TSLP on COPD Phenotypes. COPD 2020, 17, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Shan, L.; Rahman, M.S.; Unruh, H.; Halayko, A.J.; Gounni, A.S. Constitutive and inducible thymic stromal lymphopoietin expression in human airway smooth muscle cells: Role in chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L375–L382. [Google Scholar] [CrossRef] [PubMed]
- Eiwegger, T.; Akdis, C.A. IL-33 links tissue cells, dendritic cells and Th2 cell development in a mouse model of asthma. Eur. J. Immunol. 2011, 41, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Turnquist, H.R.; Thomson, A.W. IL-33 broadens its repertoire to affect DC. Eur. J. Immunol. 2009, 39, 3292–3295. [Google Scholar] [CrossRef] [PubMed]
- Tjota, M.Y.; Hrusch, C.L.; Blaine, K.M.; Williams, J.W.; Barrett, N.A.; Sperling, A.I. Signaling through FcRγ-associated receptors on dendritic cells drives IL-33-dependent TH2-type responses. J. Allergy Clin. Immunol. 2014, 134, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Schön, M.P.; Erpenbeck, L. The Interleukin-23/Interleukin-17 Axis Links Adaptive and Innate Immunity in Psoriasis. Front. Immunol. 2018, 9, 1323. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.M.; Rubin, C.J.; Khandpur, R.; Wang, J.Y.; Riblett, M.; Yalavarthi, S.; Villanueva, E.C.; Shah, P.; Kaplan, M.J.; Bruce, A.T. Mast cells and neutrophils release IL-17 through extracellular trap formation in psoriasis. J. Immunol. 2011, 187, 490–500. [Google Scholar] [CrossRef] [Green Version]
- Ishigame, H.; Kakuta, S.; Nagai, T.; Kadoki, M.; Nambu, A.; Komiyama, Y.; Fujikado, N.; Tanahashi, Y.; Akitsu, A.; Kotaki, H.; et al. Differential roles of interleukin-17A and -17F in host defense against mucoepithelial bacterial infection and allergic responses. Immunity 2009, 30, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, F.; Von Garnier, C.; Obregon, C.; Rothen-Rutishauser, B.; Gehr, P.; Nicod, L. Role of dendritic cells in the lung: In vitro models, animal models and human studies. Expert Rev. Respir. Med. 2008, 2, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Blom, R.A.M.; Erni, S.T.; Krempaská, K.; Schaerer, O.; Van Dijk, R.M.; Amacker, M.; Moser, C.; Hall, S.R.R.; Von Garnier, C.; Blank, F. A Triple Co-Culture Model of the Human Respiratory Tract to Study Immune-Modulatory Effects of Liposomes and Virosomes. PLoS ONE 2016, 11, e0163539. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Control n = 10 | Asthma n = 11 | COPD n = 11 | Overall p-Value & | Pairwise p-Value * | |||
|---|---|---|---|---|---|---|---|
| Asthma vs. Control | COPD vs. Control | Asthma vs. COPD | |||||
| Age (years) | 30 (25–40) | 55 (43–71) | 65 (60–74.4) | 0.001 | 0.0004 | 0.0001 | 0.084 |
| Gender (F/M) | 8/2 | 6/5 | 7/4 | 0.450 | |||
| BMI (kg/m2) | 21.7 (19.8–23.6) | 27.5 (26.9–29.8) | 28.3 (24.8–29.4) | 0.024 | 0.029 | 0.010 | 0.898 |
| Atopy (n) | 2 | 7 | 3 | 0.103 | |||
| Smoking exposure (pack-years) | 0 (0–1) | 0 (0–0) | 47.4 (30–52.5) | <0.0001 | 0.436 | <0.0001 | 0.0008 |
| FEV1 (% predicted) | 107.5 (104–110) | 79 (75–86) | 58 (48–72) | <0.0001 | <0.0001 | 0.0002 | 0.0066 |
| FEV1/VC (%) | 94.0 (86.0–107) | 81.5 (76–91) | 53 (50–64) | <0.0001 | 0.023 | 0.0001 | 0.0002 |
| FeNO (ppb) | 15.0 (12.6–16.3) | 28.0 (13.5–59.9) | 18.0 (14.6–23.1) | 0.076 | 0.055 | 0.121 | 0.237 |
| ACT (points) | N.A. | 22 (19–24) | N.A. | N.A | N.A. | N.A. | N.A. |
| ICS treatment (n) | N.A. | 6 | 0 | N.A | N.A. | N.A. | N.A. |
| LABA/LAMA treatment (n) | N.A. | 9 | 10 | N.A | N.A | N.A | N.A |
| CAT (points) | N.A. | N.A. | 10 (8–12) | N.A | N.A. | N.A. | N.A. |
| mMRC (points) | N.A. | N.A. | 2 (1–2.5) | N.A | N.A. | N.A. | N.A. |
| Control | moDCs | moDCs/Epithelium | moDCs/Epithelium+moMφs | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TSLP | IL-33 | IL-17A | TSLP | IL-33 | IL-17A | TSLP | IL-33 | IL-17A | ||||||||||
| R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | |
| CHI3L1 | 0.83 | 0.01 | 0.17 | 0.67 | 0.25 | 0.49 | −0.27 | 0.49 | −0.50 | 0.21 | −0.03 | 0.93 | −0.10 | 0.82 | −0.38 | 0.31 | −0.14 | 0.76 |
| IL-12p40 | −0.18 | 0.64 | −0.35 | 0.36 | −0.53 | 0.12 | −0.70 | 0.04 | −0.69 | 0.06 | −0.20 | 0.61 | −0.69 | 0.06 | −0.53 | 0.14 | −0.14 | 0.76 |
| IL-1β | 0.72 | 0.03 | -0.03 | 0.93 | 0.08 | 0.83 | 0.10 | 0.80 | 0.19 | 0.65 | −0.03 | 0.93 | 0.31 | 0.46 | 0.10 | 0.80 | −0.32 | 0.48 |
| IL-6 | −0.17 | 0.67 | 0.28 | 0.46 | 0.58 | 0.08 | 0.57 | 0.11 | 0.40 | 0.32 | 0.62 | 0.08 | 0.69 | 0.06 | 0.18 | 0.64 | 0.18 | 0.70 |
| IL-8 | 0.28 | 0.46 | −0.08 | 0.83 | 0.35 | 0.33 | 0.17 | 0.67 | −0.24 | 0.57 | 0.30 | 0.43 | 0.31 | 0.46 | 0.15 | 0.70 | 0.00 | 1.00 |
| TNF-α | 0.17 | 0.67 | 0.03 | 0.93 | −0.07 | 0.85 | −0.47 | 0.21 | −0.17 | 0.69 | 0.27 | 0.49 | −0.48 | 0.23 | −0.07 | 0.86 | −0.75 | 0.05 |
| TSLP | 0.17 | 0.69 | 0.22 | 0.58 | 0.75 | 0.05 | 0.50 | 0.21 | 0.93 | 0.003 | 0.50 | 0.39 | ||||||
| IL-33 | 0.17 | 0.69 | 0.52 | 0.15 | 0.75 | 0.05 | 0.04 | 0.94 | 0.93 | 0.003 | −0.20 | 0.70 | ||||||
| IL-17A | 0.22 | 0.58 | 0.52 | 0.15 | 0.50 | 0.21 | 0.04 | 0.94 | 0.50 | 0.39 | −0.20 | 0.70 | ||||||
| Asthma | moDCs | moDCs/Epithelium | moDCs/Epithelium+moMφs | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TSLP | IL-33 | IL-17A | TSLP | IL-33 | IL-17A | TSLP | IL-33 | IL-17A | ||||||||||
| R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | |
| CHI3L1 | 0.07 | 0.85 | 0.43 | 0.24 | 0.42 | 0.20 | −0.52 | 0.13 | 0.22 | 0.58 | −0.06 | 0.85 | 0.52 | 0.13 | 0.80 | 0.01 | 0.48 | 0.13 |
| IL-12p40 | 0.65 | 0.04 | 0.33 | 0.38 | 0.72 | 0.01 | 0.48 | 0.16 | −0.13 | 0.73 | 0.54 | 0.09 | 0.77 | 0.01 | 0.57 | 0.11 | 0.70 | 0.02 |
| IL-1β | 0.61 | 0.06 | 0.48 | 0.19 | 0.70 | 0.02 | −0.39 | 0.26 | 0.00 | 1.00 | 0.50 | 0.12 | 0.41 | 0.24 | 0.62 | 0.08 | 0.52 | 0.10 |
| IL-6 | 0.43 | 0.21 | 0.15 | 0.70 | 0.75 | 0.01 | 0.05 | 0.88 | −0.27 | 0.49 | 0.54 | 0.09 | 0.82 | 0.00 | 0.52 | 0.15 | 0.67 | 0.02 |
| IL-8 | −0.12 | 0.75 | 0.55 | 0.12 | 0.40 | 0.22 | −0.20 | 0.58 | −0.03 | 0.93 | 0.50 | 0.12 | 0.42 | 0.23 | 0.72 | 0.03 | 0.57 | 0.07 |
| TNF-α | 0.35 | 0.36 | 0.55 | 0.16 | 0.22 | 0.53 | −0.03 | 0.93 | −0.48 | 0.23 | 0.48 | 0.16 | 0.72 | 0.03 | 0.67 | 0.07 | 0.31 | 0.38 |
| TSLP | 0.31 | 0.46 | 0.38 | 0.28 | −0.13 | 0.73 | 0.32 | 0.37 | 0.43 | 0.29 | 0.25 | 0.49 | ||||||
| IL-33 | 0.31 | 0.46 | 0.37 | 0.33 | −0.13 | 0.73 | −0.23 | 0.55 | 0.43 | 0.29 | 0.68 | 0.04 | ||||||
| IL-17A | 0.38 | 0.28 | 0.37 | 0.33 | 0.32 | 0.37 | −0.23 | 0.55 | 0.25 | 0.49 | 0.68 | 0.04 | ||||||
| COPD | moDCs | moDCs/Epithelium | moDCs/Epithelium+moMφs | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TSLP | IL-33 | IL-17A | TSLP | IL-33 | IL-17A | TSLP | IL-33 | IL-17A | ||||||||||
| R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | R | p-Value | |
| CHI3L1 | 0.53 | 0.10 | 0.62 | 0.04 | 0.63 | 0.04 | 0.67 | 0.03 | 0.10 | 0.82 | 0.61 | 0.05 | 0.70 | 0.03 | 0.55 | 0.16 | 0.43 | 0.19 |
| IL-12p40 | 0.58 | 0.06 | 0.75 | 0.01 | 0.67 | 0.02 | 0.65 | 0.04 | 0.38 | 0.35 | 0.48 | 0.13 | 0.79 | 0.01 | 0.10 | 0.82 | 0.62 | 0.04 |
| IL-1β | 0.66 | 0.03 | 0.75 | 0.01 | 0.75 | 0.01 | 0.52 | 0.13 | −0.10 | 0.82 | 0.68 | 0.02 | 0.87 | 0.001 | 0.19 | 0.65 | 0.54 | 0.09 |
| IL-6 | 0.46 | 0.15 | 0.44 | 0.18 | 0.65 | 0.03 | 0.54 | 0.11 | 0.07 | 0.87 | 0.66 | 0.03 | 0.85 | 0.002 | −0.33 | 0.42 | 0.71 | 0.01 |
| IL-8 | 0.53 | 0.10 | 0.45 | 0.17 | 0.65 | 0.03 | 0.70 | 0.03 | 0.19 | 0.65 | 0.61 | 0.05 | 0.72 | 0.02 | 0.29 | 0.49 | 0.65 | 0.03 |
| TNF-α | 0.65 | 0.03 | 0.54 | 0.09 | 0.73 | 0.01 | 0.88 | 0.002 | 0.11 | 0.82 | 0.61 | 0.06 | 0.88 | 0.001 | −0.33 | 0.42 | 0.51 | 0.11 |
| TSLP | 0.65 | 0.03 | 0.78 | 0.00 | 0.29 | 0.49 | 0.56 | 0.09 | −0.14 | 0.74 | 0.50 | 0.14 | ||||||
| IL-33 | 0.65 | 0.03 | 0.47 | 0.14 | 0.29 | 0.49 | −0.45 | 0.26 | −0.14 | 0.74 | −0.12 | 0.78 | ||||||
| IL-17A | 0.78 | 0.00 | 0.47 | 0.14 | 0.56 | 0.09 | −0.45 | 0.26 | 0.50 | 0.14 | −0.12 | 0.78 | ||||||
| Controls | Asthma | COPD | p-Value ◊ | |
|---|---|---|---|---|
| TSLPR | ||||
| moDC | 10.45 (7.30–12.50) *# | 28.20 (28.20–29.50) * | 21.90 (18.10–22.10) # | 0.009 |
| moDC/epithelium | 9.05 (4.80–13.50) | 36.60 (11.70–46.50) | 12.60 (12.20–17.70) | 0.12 |
| moDC/epithelium/moMφ | 10.10 (6.20–16.70) * | 28.20 (19.00–31.90) * | 18.90 (12.70–33.00) | 0.06 |
| Summed cells | 9.95 (6.20–13.50) *# | 28.20 (16.00–9.30) *^ | 18.10 (12.60–23.50) #^ | 0.001 |
| ST2 | ||||
| moDC | 24.05 (17.30–31.30) | 27.60 (27.00–32.40) | 26.20 (18.60–39.80) | 0.51 |
| moDC/epithelium | 29.30 (18.10–33.80) | 40.90 (31.50–43.80) | 39.90 (20.10–43.40) | 0.39 |
| moDC/epithelium/moMφ | 30.55 (19.60–32.90) | 35.10 (29.70–35.80) | 40.30 (17.80–44.20) | 0.62 |
| Summed cells | 28.30 (18.10–33.60) * | 32.40 (27.00–42.50) * | 39.80 (17.80–44.20) | 0.16 |
| IL17RA | ||||
| moDC | 39.50 (39.50–52.20) | 64.80 (60.40–65.30) | 58.60 (47.00–64.50) | 0.11 |
| moDC/Epithelium | 37.65 (23.30–57.00) | 60.80 (49.60–61.40) | 54.60 (39.80–61.10) | 0.30 |
| moDC/Epithelium/moMφ | 28.60 (23.30–45.10) | 51.60 (47.40–52.70) | 63.80 (26.80–64.80) | 0.15 |
| Summed cells | 37.15 (23.30–52.20) *# | 60.40 (47.40–64.80) * | 58.60 (38.70–64.80) # | 0.008 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paplinska-Goryca, M.; Misiukiewicz-Stepien, P.; Proboszcz, M.; Nejman-Gryz, P.; Gorska, K.; Krenke, R. The Expressions of TSLP, IL-33, and IL-17A in Monocyte Derived Dendritic Cells from Asthma and COPD Patients are Related to Epithelial–Macrophage Interactions. Cells 2020, 9, 1944. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091944
Paplinska-Goryca M, Misiukiewicz-Stepien P, Proboszcz M, Nejman-Gryz P, Gorska K, Krenke R. The Expressions of TSLP, IL-33, and IL-17A in Monocyte Derived Dendritic Cells from Asthma and COPD Patients are Related to Epithelial–Macrophage Interactions. Cells. 2020; 9(9):1944. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091944
Chicago/Turabian StylePaplinska-Goryca, Magdalena, Paulina Misiukiewicz-Stepien, Malgorzata Proboszcz, Patrycja Nejman-Gryz, Katarzyna Gorska, and Rafal Krenke. 2020. "The Expressions of TSLP, IL-33, and IL-17A in Monocyte Derived Dendritic Cells from Asthma and COPD Patients are Related to Epithelial–Macrophage Interactions" Cells 9, no. 9: 1944. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091944