Investigating the Role of Methylation in Silencing of VDR Gene Expression in Normal Cells during Hematopoiesis and in Their Leukemic Counterparts



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Murine Blood Cells
2.2. Isolation of Human Hematopoietic Stem and Progenitor Cells
2.3. Cell Lines and Acute Myeloid Leukemia Cells
2.4. cDNA Synthesis and Real-Time Polymerase Chain Reaction (PCR)
- murine Gapdh forward 5′AACTTTGGCATTGTGGAAGG3′, reverse 5′ ACACATTGGGGGTAGGAACA3′;
- murine Vdr forward 5′CACCTGGCTGATCTTGTCAGT 3′, reverse 5′ CTGGTCATCAGAGGTGAGGTC 3′;
- murine Cyp24a1 forward 5′CACGGTAGGCTGCTGAGATT 3′, reverse 5′ CCAGTCTTCGCAGTTGTCC 3′;
- human GAPDH forward 5′-CATGAGAAGTATGACAACAGCCT-30, reverse 5′-AGTCCTTCCACGATACCAAAGT-3′;
- human VDR forward 5′-CCTTCACCATGGACGACATG-30, reverse 5′-CGGCTTTGGTCACGTCACT-3′;
- human VDR1C forward 5′-GGGTCTGAAGTGTCTGTGAGA-3′, reverse 5′-GAAGTGCTGGCC GCCATTG-3′;
- human CYP24A1 forward 5′-CTCATGCTAAATACCCAGGTG-30, reverse 5′-TCGCTGGCAAAACGCGATGGG-3′;
2.5. Analysis of Methylation in Promoter Regions of Murine and Human VDR
- murine Vdr forward outer and inner: 5′-GAGAAATTTATTTGAGGTTTTTTATT-3′;
- murine Vdr reverse outer: 5′-CCAACCACAATACAACACAAAC-3′;
- murine Vdr reverse inner: 5′-TAATTCTACCCAATCTACTATAAAC-3′;
- human VDR1A forward outer 5′-GTTGGGTTGTTTTTGTTTGTTAAA -3′, reverse outer 5′-TCAAACCTCAATACCCCTTAATATC-3′;
- human VDR1C forward outer 5′-GGGATTAAAGTTTTTGGAAAGAGTT-3′, reverse outer 5′-CACCTACCTAAAAAAACAAAAAACAA-3′;
- human VDR1A_1 forward inner 5′-TGGGTTGTTTTTGTTTGTTAAAAG-3′, reverse inner 5′-CCCTATCCTAAAACCCCCTTTC-3′;
- human VDR1A_2 forward inner 5′-GAAAGGGGGTTTTAGGATAGGG-3′, reverse inner 5′-TACCCC TTAATATCCCAACCTC-3′;
- human VDR1C forward inner 5′-GTTTATTTTTTTAGAGATTGGGG-3′, reverse inner 5′-ATCTCACAAACACTTCAAAC-3′;
2.6. Statistical Analysis
3. Results
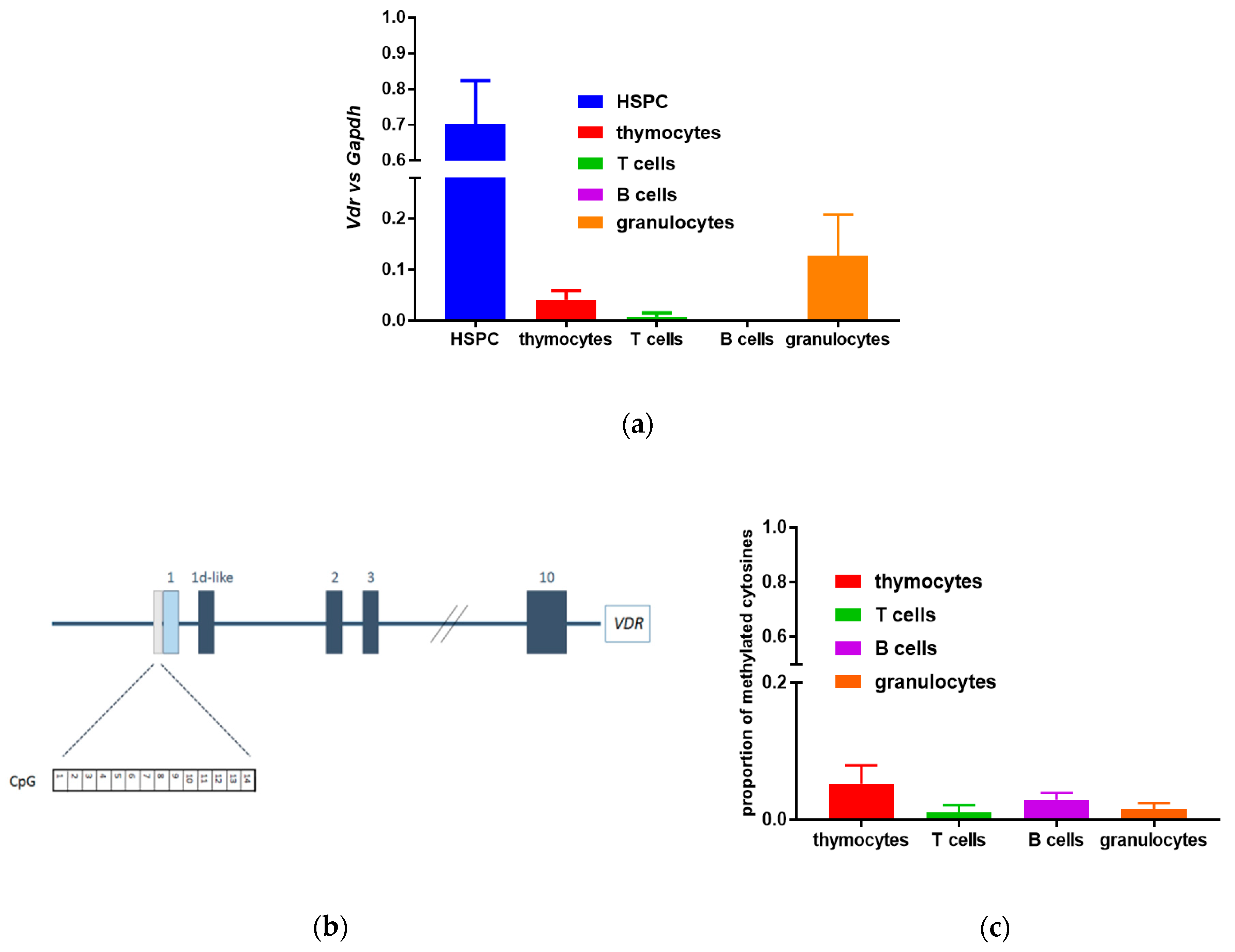
3.1. Lack of Methylation in the Promoter Region of the Murine Vdr Gene in Murine Blood Cells
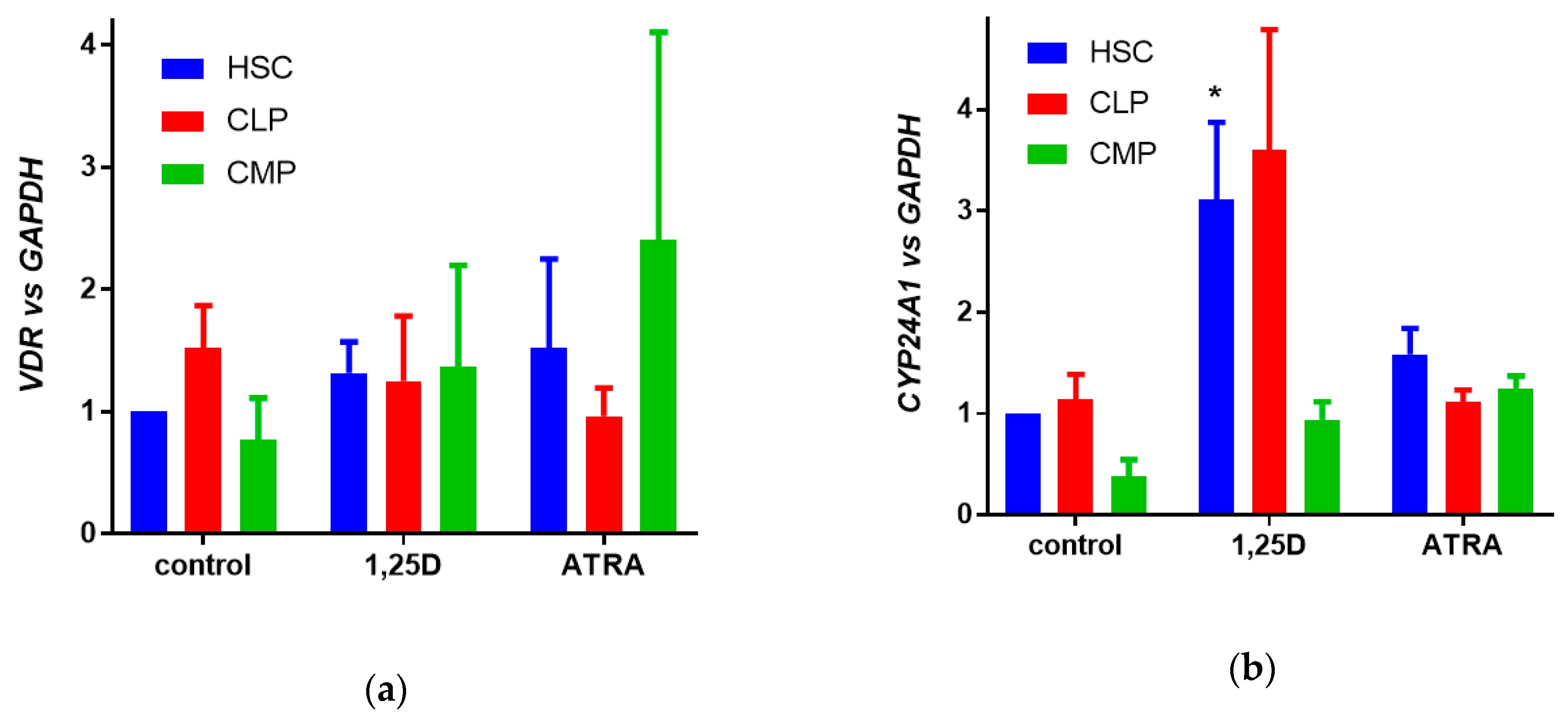
3.2. Expression of VDR Gene and Activity of VDR Protein in Human Hematopoietic Stem and Progenitor Cells
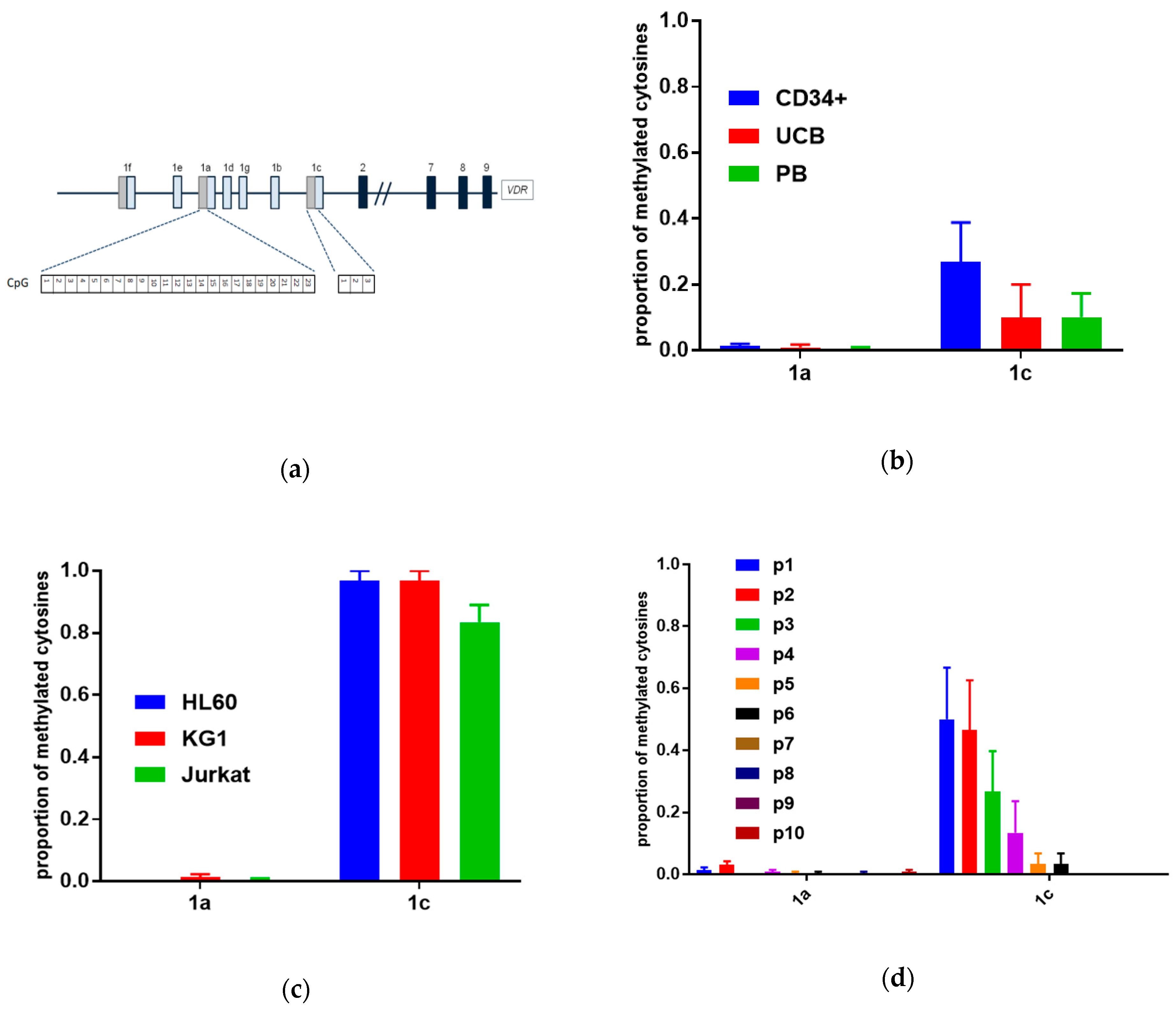
3.3. Methylation in Promoter Regions of Human VDR Gene in Human Blood Cells
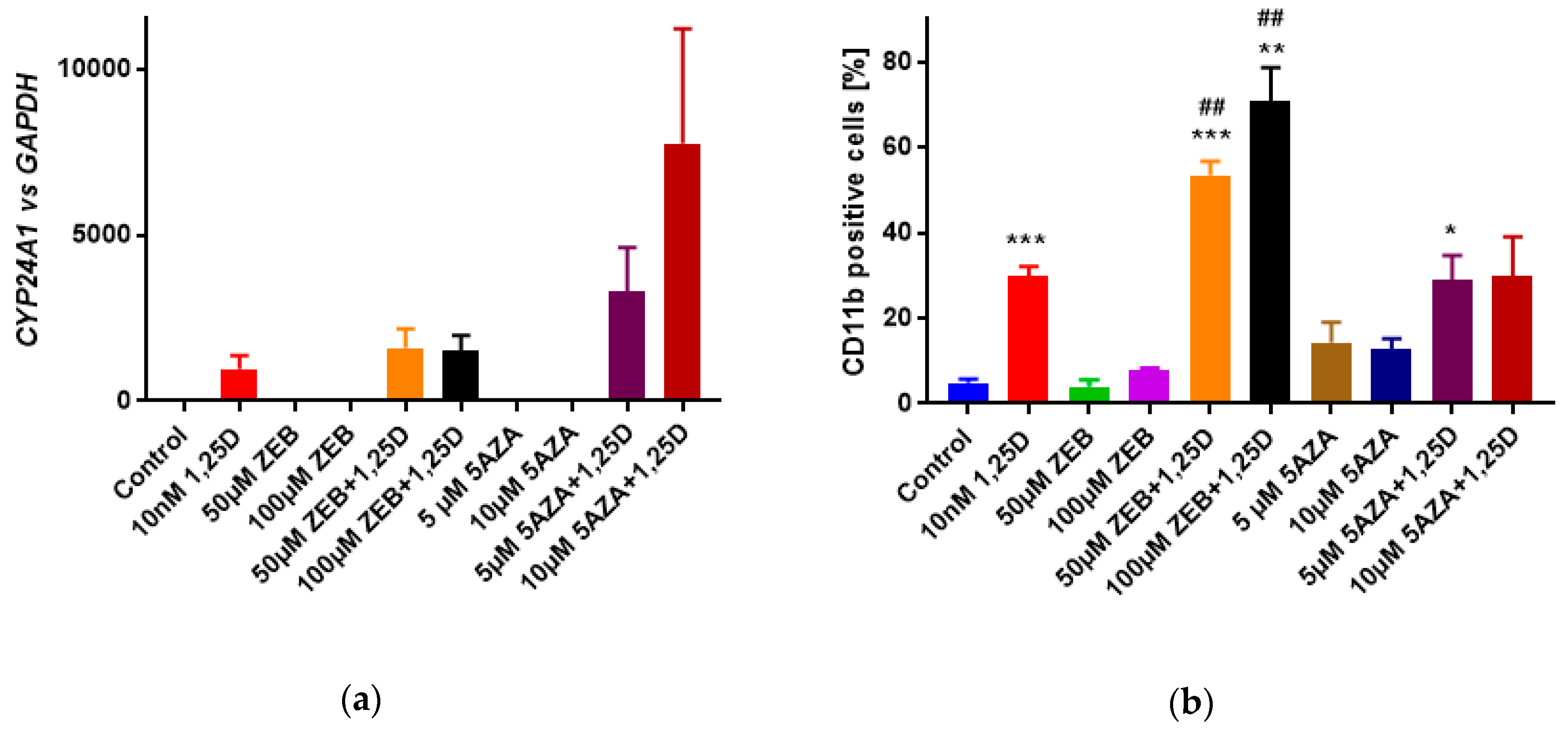
3.4. Influence of Hypomethylating Agents on 1,25D-Induced Differentiation of HL60 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aranda, A.; Pascual, A. Nuclear hormone receptors and gene expression. Physiol. Rev. 2001, 81, 1269–1304. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef] [PubMed]
- Holick, M. Vitamin D and bone health. J. Nutr. 1996, 126, 1159S–1164S. [Google Scholar] [CrossRef] [PubMed]
- Makishima, M.; Lu, T.; Xie, W.; Whitfield, G.; Domoto, H.; Evans, R.; Haussler, M.; Mangelsdorf, D. Vitamin D receptor as an intestinal bile acid sensor. Science 2002, 296, 1313–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollitz, E.; Zhang, G.; Hawkins, M.; Whitfield, G.; Reif, D.; Kullman, S. Evolutionary and functional diversification of the vitamin D receptor-lithocholic acid partnership. PLoS ONE 2016, 11, e0168278. [Google Scholar] [CrossRef] [Green Version]
- van Etten, E.; Mathieu, C. Immunoregulation by 1,25-dihydroxyvitamin D3: Basic concepts. J. Steroid Biochem. Mol. Biol. 2005, 97, 93–101. [Google Scholar] [CrossRef]
- Brown, G.; Mooney, C.J.; Alberti-Servera, L.; Muenchow, L.v.; Toellner, K.-M.; Ceredig, R.; Rolink, A. Versatility of stem and progenitor cells and the instructive actions of cytokines on hematopoiesis. Crit. Rev. Clin. Lab. Sci. 2015, 52, 168–179. [Google Scholar] [PubMed]
- Janik, S.; Nowak, U.; Łaszkiewicz, A.; Satyr, A.; Majkowski, M.; Marchwicka, A.; Śnieżewski, Ł.; Berkowska, K.; Gabryś, M.; Cebrat, M.; et al. Diverse regulation of vitamin D receptor gene expression by 1,25-dihydroxyvitamin D and ATRA in murine and human blood cells at early stages of their differentiation. Int. J. Mol. Sci. 2017, 18, 1323. [Google Scholar] [CrossRef] [Green Version]
- Cortes, M.; Chen, M.; Stachura, D.; Liu, S.; Kwan, W.; Wright, F.; Vo, L.; Theodore, L.; Esain, V.; Frost, I.; et al. Developmental vitamin D availability impacts hematopoietic stem cell production. Cell Rep. 2016, 17, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Grande, A.; Montanari, M.; Tagliafico, E.; Manfredini, R.; Zanocco Marani, T.; Siena, M.; Tenedini, E.; Gallinelli, A.; Ferrari, S. Physiological levels of 1alpha, 25 dihydroxyvitamin D3 induce the monocytic commitment of CD34+ hematopoietic progenitors. J. Leukoc. Biol. 2002, 71, 641–651. [Google Scholar]
- Takahashi, T.; Suzuki, A.; Ichiba, S.; Okuno, Y.; Sugiyama, H.; Imura, H.; Nakamura, K.; Iho, S.; Hoshino, T. Effect of 1,25(OH)2D3 on normal human CFU-GM: Target cells of the agent in the suppression of colony formation and induction of macrophage colonies. Int. J. Hematol. 1991, 54, 57–63. [Google Scholar]
- Studzinski, G.; Harrison, J.; Wang, X.; Sarkar, S.; Kalia, V.; Danilenko, M. Vitamin D control of hematopoietic cell differentiation and leukemia. J. Cell. Biochem. 2015, 116, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Bird, A. DNA methylation landscapes provocative inside from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Beckett, E.; Jones, P.; Veysey, M.; Duesing, K.; Martin, C.; Furst, J.; Yates, Z.; Jablonski, N.; Chaplin, G.; Lucock, M. VDR gene methylation as a molecular adaption to light exposure: Historic, recent and genetic influences. Am. J. Hum. Biol. 2017, 29, e23010. [Google Scholar] [CrossRef] [PubMed]
- Crofts, L.; Hancock, M.; Morrison, N.; Eisman, J. Multiple promoters direct the tissue-specific expression of novel N-terminal variant human vitamin D receptor gene transcripts. Proc. Natl. Acad. Sci. USA 1998, 95, 10529–10534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halsall, J.; Osborne, J.; Hutchinson, P.; Pringle, J. In silico analysis of the 5’ region of the vitamin D receptor gene: Functional implications of evolutionary conservation. J. Steroid Biochem. Mol. Biol. 2007, 103, 352–356. [Google Scholar] [CrossRef]
- Wang, X.; Shook, J.; Edinger, M.; Warner, N.; Bush-Donovan, C. Multiparametric immunophenotyping of human hematopoietic stem cells and progenitor cells by flow cytometry. In BD Biosciences Application Note; BD Biosciences: San Jose, CA, USA, 2012. [Google Scholar]
- Baurska, H.; Kiełbiński, M.; Biecek, P.; Haus, O.; Jaźwiec, B.; Kutner, A.; Marcinkowska, E. Monocytic differentiation induced by side-chain modified analogs of vitamin D in ex vivo cells from patients with acute myeloid leukemia. Leuk. Res. 2014, 38, 638–647. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, L.; Dahiya, R. Methprimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef] [Green Version]
- Cebrat, M.; Cebula, A.; Laszkiewicz, A.; Kasztura, M.; Miazek, A.; Kisielow, P. Mechanism of lymphocyte-specific inactivation of RAG-2 intragenic promoter of NWC: Implications for epigenetic control of RAG locus. Mol. Immunol. 2008, 45, 2297–2306. [Google Scholar] [CrossRef]
- Ayuso, T.; Aznar, P.; Soriano, L.; Olaskoaga, A.; Roldan, M.; Otano, M.; Ajuria, I.; Soriano, G.; Lacruz, F.; Mendioroz, M. Vitamin D receptor gene is epigenetically altered and transcriptionally up-regulated in multiple sclerosis. PLoS ONE 2017, 12, e0174726. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.; Catovsky, D.; Daniel, M.; Flandrin, G.; Galton, D.; Gralnick, H.; Sultan, C. Proposals for the classification of the acute leukaemias. French-American-British (FAB) co-operative group. Br. J. Haematol. 1976, 33, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Etzioni, A.; Hochberg, Z.; Pollak, S.; Meshulam, T.; Zakut, V.; Tzehoval, E.; Keisari, Y.; Aviram, I.; Spirer, Z.; Benderly, A.; et al. Defective leukocyte fungicidal activity in end-organ resistance to 1,25-dihydroxyvitamin D. Pediatr. Res. 1989, 25, 276–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchwicka, A.; Cebrat, M.; Łaszkiewicz, A.; Śnieżewski, Ł.; Brown, G.; Marcinkowska, E. Regulation of vitamin D receptor expression by retinoic acid receptor alpha in acute myeloid leukemia cells. J. Steroid Biochem. Mol. Biol. 2016, 159, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchwicka, A.; Corcoran, A.; Berkowska, K.; Marcinkowska, E. Restored expression of vitamin D receptor and sensitivity to 1,25-dihydroxyvitamin D3 in response to disrupted fusion FOP2-FGFR1 gene in acute myeloid leukemia cells. Cell Biosci. 2016, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Munker, R.; Norman, A.; Koeffler, H. Vitamin D compounds. Effect on clonal proliferation and differentiation of human myeloid cells. J. Clin. Investig. 1986, 78, 424–430. [Google Scholar] [CrossRef]
- Gocek, E.; Kielbinski, M.; Baurska, H.; Haus, O.; Kutner, A.; Marcinkowska, E. Different susceptibilities to 1,25-dihydroxyvitamin D3-induced differentiation of AML cells carrying various mutations. Leuk. Res. 2010, 34, 649–657. [Google Scholar] [CrossRef]
- Harrison, J.; Bershadskiy, A. Clinical experience using vitamin D and analogs in the treatment of myelodysplasia and acute myeloid leukemia: A review of the literature. Leuk. Res. Treat. 2012, 2012, 125814. [Google Scholar] [CrossRef]
- Lowenberg, B. Acute myeloid leukemia: The challenge of capturing disease variety. Hematol. Am. Soc. Hematol. Educ. Program 2008, 2008, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Marik, R.; Fackler, M.; Gabrielson, E.; Zeiger, M.; Sukumar, S.; Stearns, V.; Umbricht, C. DNA methylation-related vitamin D receptor insensitivity in breast cancer. Cancer Biol. Ther. 2010, 10, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Paubelle, E.; Zylbersztejn, F.; Maciel, T.; Carvalho, C.; Mupo, A.; Cheok, M.; Lieben, L.; Sujobert, P.; Decroocq, J.; Yokoyama, A.; et al. Vitamin D receptor controls cell stemness in acute myeloid leukemia and in normal bone marrow. Cell Rep. 2020, 30, 739–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granja, J.; Klemm, S.; McGinnis, L.; Kathiria, A.; Mezger, A.; Corces, M.; Parks, B.; Gars, E.; Liedtke, M.; Zheng, G.; et al. Single-cell multiomic analysis identifies regulatory programs in mixed-phenotype acute leukemia. Nat. Biotechnol. 2019, 37, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Höbaus, J.; Fetahu, I.; Khorchide, M.; Manhardt, T.; Kallay, E. Epigenetic regulation of the 1,25-dihydroxyvitamin D3 24-hydroxylase (CYP24A1) in colon cancer cells. J. Steroid Biochem. Mol. Biol. 2013, 136, 296–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Age | Sex | Type 1 | Prognosis Based on Cytogenetics | Methylation of 1c Promoter 2 |
|---|---|---|---|---|---|
| p1 | 54 | M | M5b | intermediate | +++ |
| p2 | 60 | F | M1 | poor | +++ |
| p3 | 79 | M | M5b | poor | ++ |
| p4 | 42 | M | M2 | good | ++ |
| p5 | 54 | F | M2 | good | + |
| p6 | 50 | F | M2 | intermediate | + |
| p7 | 75 | F | M1 | poor | - |
| p8 | 23 | M | M2 | good | - |
| p9 | 79 | M | M1 | poor | - |
| p10 | 74 | F | M5b | intermediate | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, U.; Janik, S.; Marchwicka, A.; Łaszkiewicz, A.; Jakuszak, A.; Cebrat, M.; Marcinkowska, E. Investigating the Role of Methylation in Silencing of VDR Gene Expression in Normal Cells during Hematopoiesis and in Their Leukemic Counterparts. Cells 2020, 9, 1991. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091991
Nowak U, Janik S, Marchwicka A, Łaszkiewicz A, Jakuszak A, Cebrat M, Marcinkowska E. Investigating the Role of Methylation in Silencing of VDR Gene Expression in Normal Cells during Hematopoiesis and in Their Leukemic Counterparts. Cells. 2020; 9(9):1991. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091991
Chicago/Turabian StyleNowak, Urszula, Sylwia Janik, Aleksandra Marchwicka, Agnieszka Łaszkiewicz, Agnieszka Jakuszak, Małgorzata Cebrat, and Ewa Marcinkowska. 2020. "Investigating the Role of Methylation in Silencing of VDR Gene Expression in Normal Cells during Hematopoiesis and in Their Leukemic Counterparts" Cells 9, no. 9: 1991. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9091991