LPS-Induced Endotoxemia Evokes Epigenetic Alterations in Mitochondrial DNA That Impacts Inflammatory Response

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Oversight
2.2. Volunteer Recruitment and Cell Culture
2.3. Enzyme-Linked Immunosorbent Assay
2.4. Isolation of Mitochondria
2.5. Western Blot
2.6. Methylation Analysis
2.7. Quantitative Polymerase Chain Reaction
2.8. Statistical Analysis
3. Results
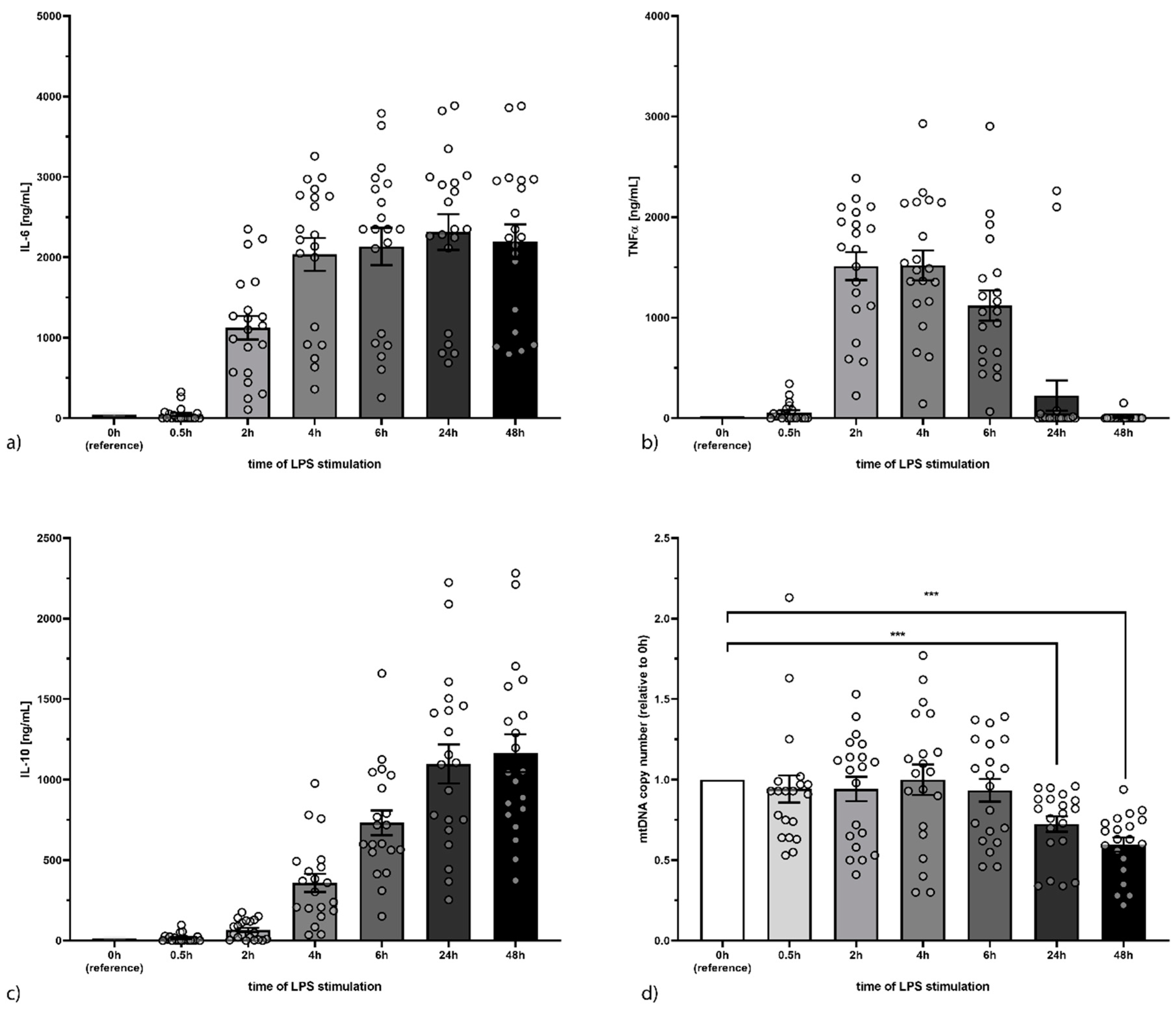
3.1. Characterisation of the Immune Response
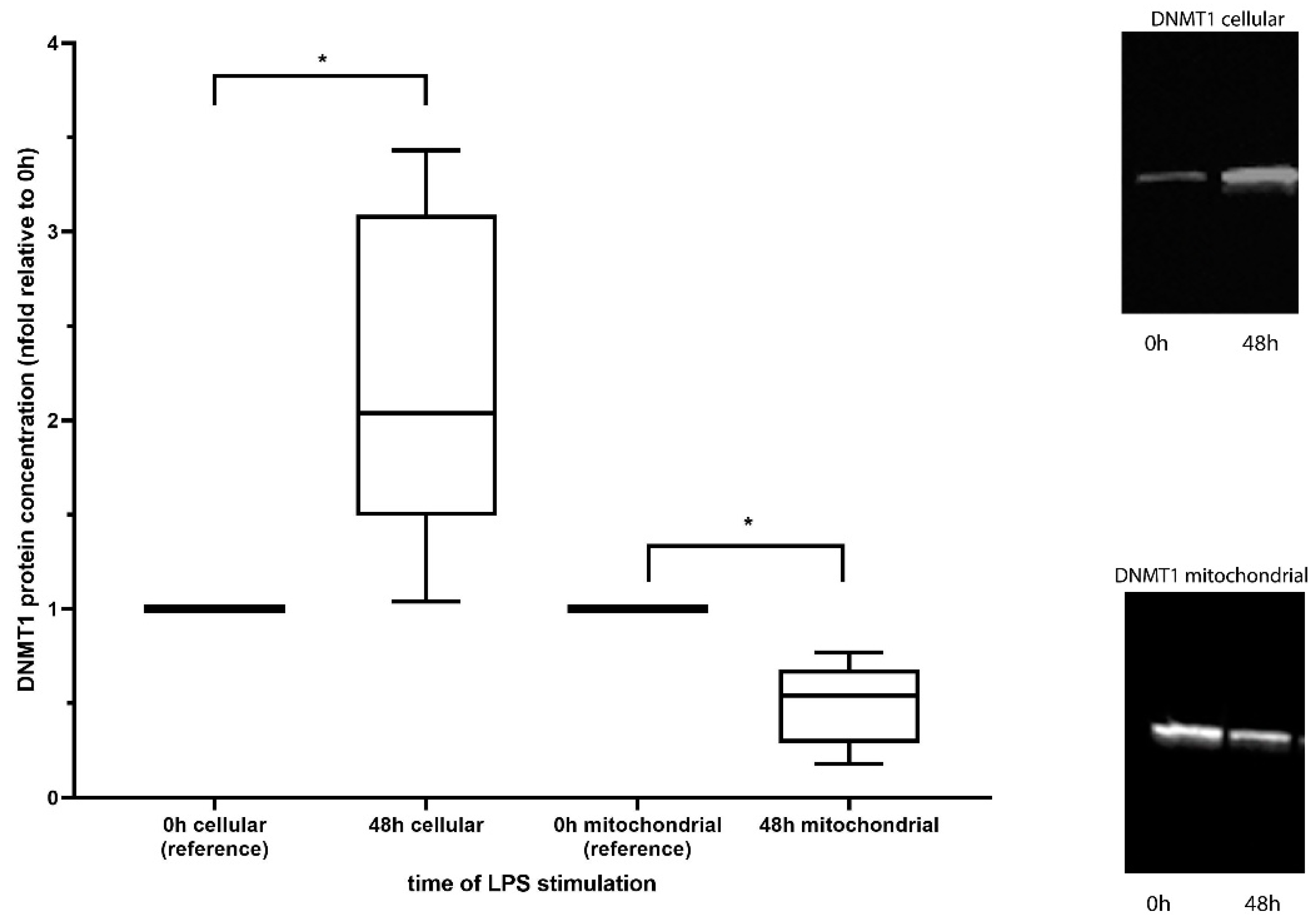
3.2. Mitochondrial Concentration of DNA Methyltransferase 1
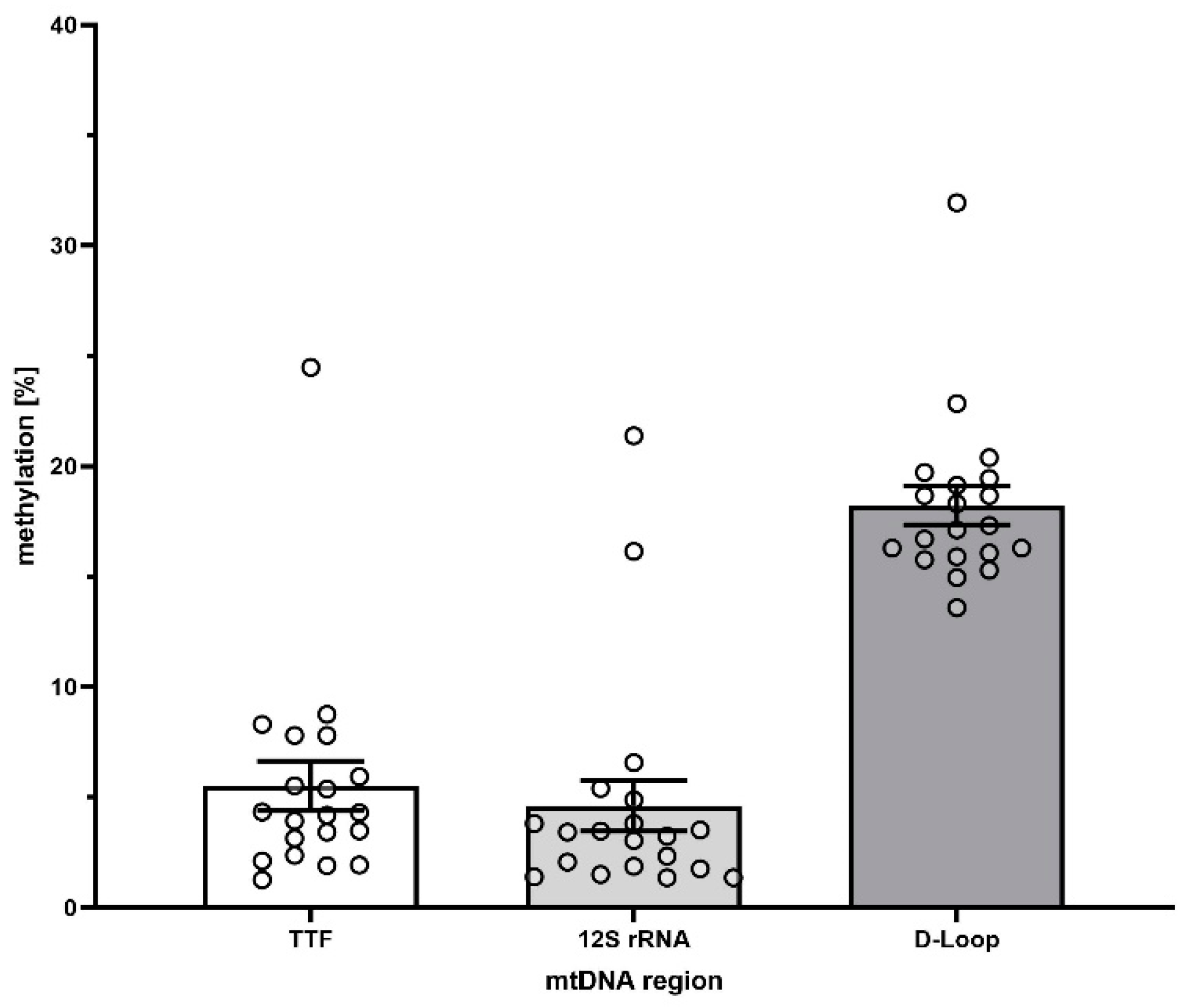
3.3. Identification of Methylated Regions in the Mitochondrial Genome
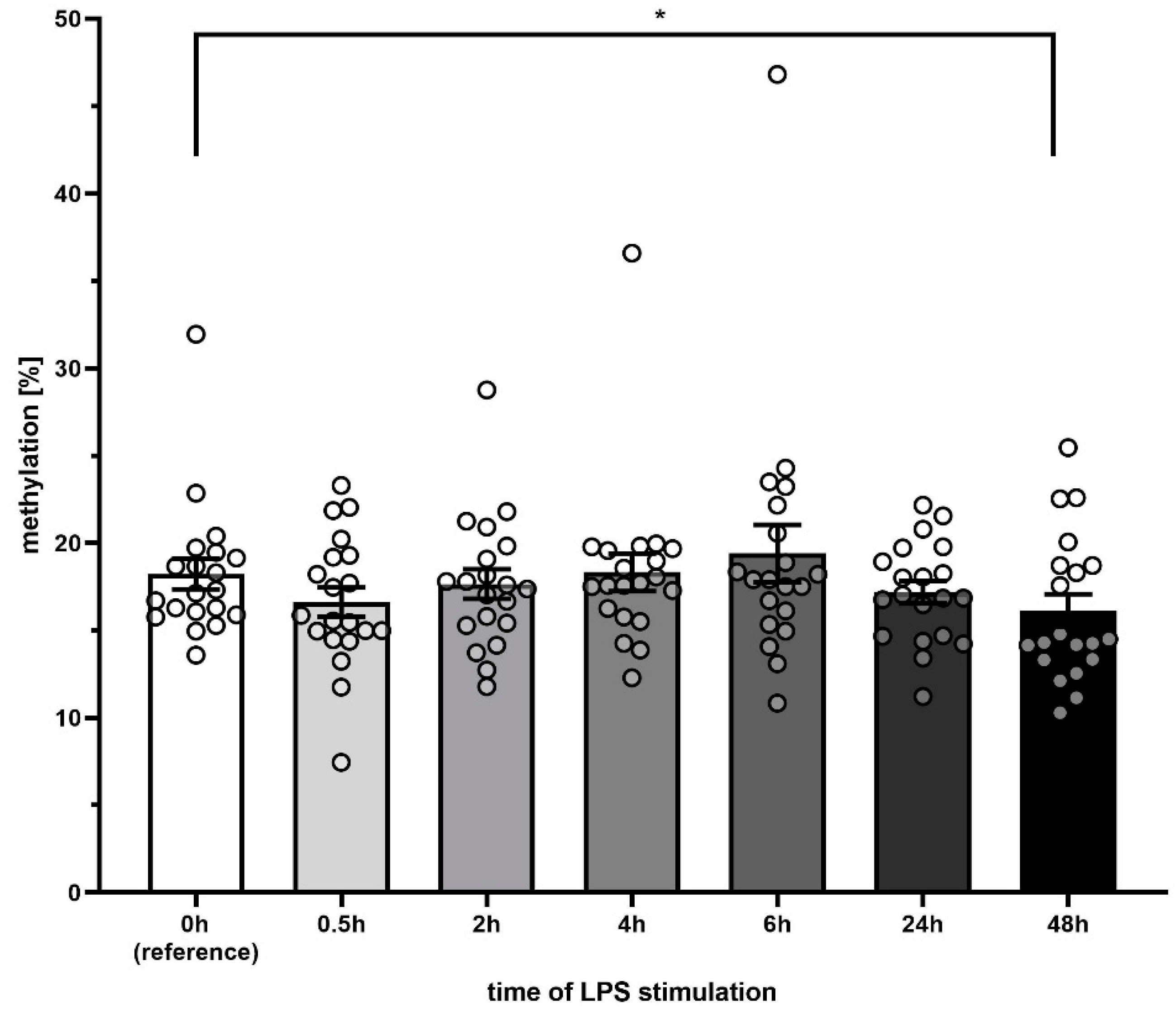
3.4. D-Loop DNA Methylation after LPS Stimulation
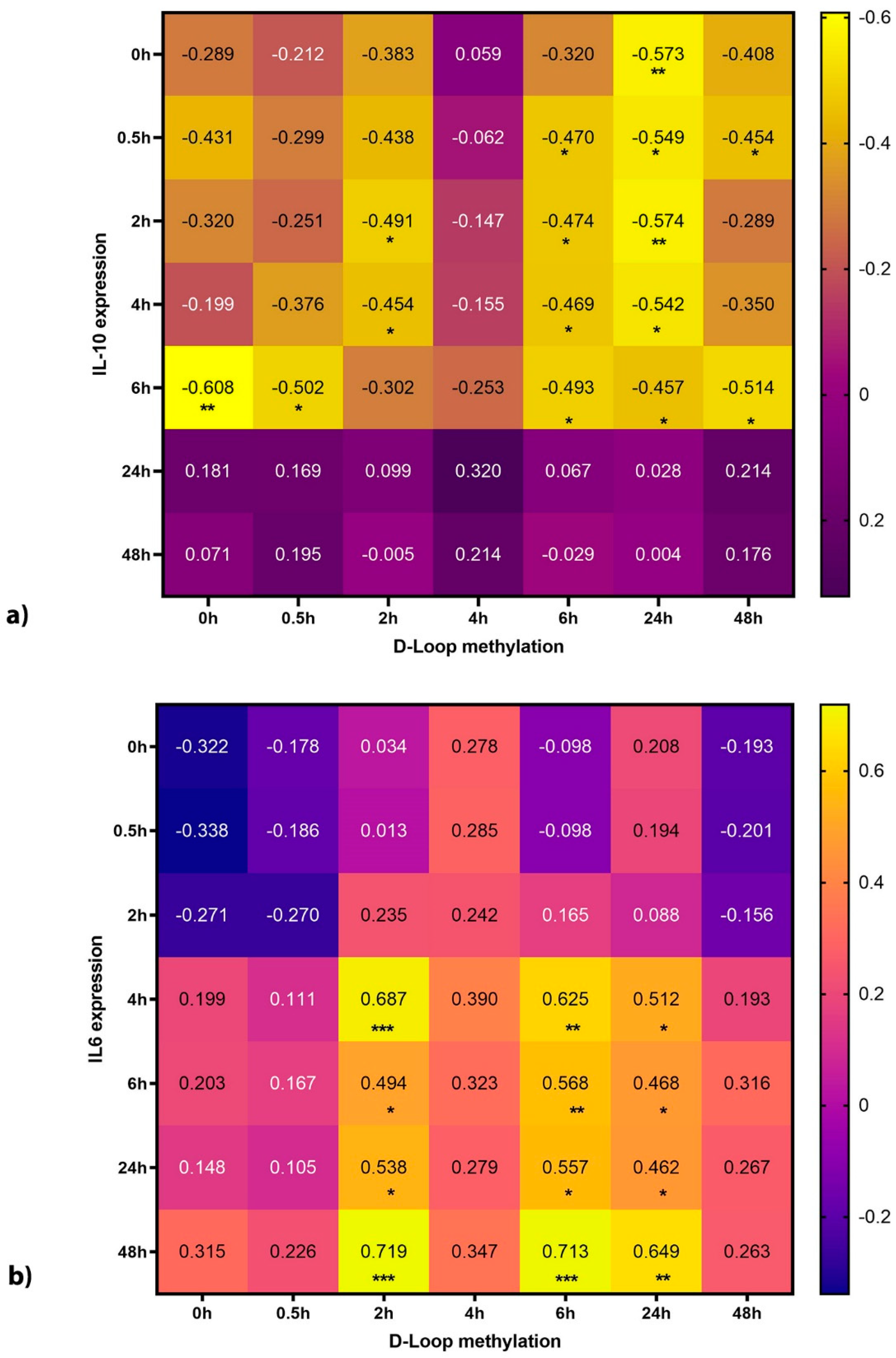
3.5. Mitochondrial DNA Methylation Correlates with Cytokine Production
4. Discussion
4.1. DNMT1 Abundance Is Reduced in Mitochondria in an Endotoxin Cell Culture Model
4.2. Methylation of mtDNA Is only Marginally Reduced upon LPS Treatment
4.3. D-Loop Methylation Levels Correlate Strongly with Cytokine Release
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A. Sepsis and septic shock. Lancet 2018, 392, 75–87. [Google Scholar] [CrossRef]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Matzinger, P. The danger model: A renewed sense of self. Science 2002, 296, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, L.V.; Hajizadeh, S.; Holme, E.; Jonsson, I.-M.; Tarkowski, A. Endogenously oxidized mitochondrial DNA induces in vivo and in vitro inflammatory responses. J. Leukoc. Biol. 2004, 75, 995–1000. [Google Scholar] [CrossRef]
- Zhang, Q.; Raoof, M.; Chen, Y.; Sumi, Y.; Sursal, T.; Junger, W.; Brohi, K.; Itagaki, K.; Hauser, C.J. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nat. Cell Biol. 2010, 464, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Schafer, S.T.; Franken, L.; Adamzik, M.; Schumak, B.; Scherag, A.; Engler, A.; Schönborn, N.; Walden, J.; Koch, S.; Baba, H.A. Mitochondrial DNA: An Endogenous Trigger for Immune Paralysis. Anesthesiology 2016, 124, 923–933. [Google Scholar] [CrossRef]
- Stacey, K.J.; Young, G.R.; Clark, F.; Sester, D.P.; Roberts, T.L.; Naik, S.; Sweet, M.J.; Hume, D.A. The molecular basis for the lack of immunostimulatory activity of vertebrate DNA. J. Immunol. 2003, 170, 3614–3620. [Google Scholar] [CrossRef] [Green Version]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar] [CrossRef]
- Ma, F.; Li, Z.; Cao, J.; Kong, X.; Gonga, G. A TGFBR2/SMAD2/DNMT1/miR-145 negative regulatory loop is responsible for LPS-induced sepsis. Biomed. Pharmacother. 2019, 112, 108626. [Google Scholar] [CrossRef]
- Dakhlallah, D.A.; Wisler, J.; Gencheva, M.; Brown, C.; Leatherman, E.R.; Singh, K.; Brundage, K.; Karsies, T.; Dakhlallah, A.; Witwer, K.W.; et al. Circulating extracellular vesicle content reveals de novo DNA methyltransferase expression as a molecular method to predict septic shock. J. Extracell. Vesicles 2019, 8, 1669881. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.A.; Liang, G. Rethinking how DNA methylation patterns are maintained. Nat. Rev. Genet. 2009, 10, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Shock, L.S.; Thakkar, P.V.; Peterson, E.J.; Moran, R.G.; Taylor, S.M. DNA methyltransferase 1, cytosine methylation, and cytosine hydroxymethylation in mammalian mitochondria. Proc. Natl. Acad. Sci. USA 2011, 108, 3630–3635. [Google Scholar] [CrossRef] [Green Version]
- Argan, C.; Lusty, C.J.; Shore, G.C. Membrane and cytosolic components affecting transport of the precursor for ornithine carbamyltransferase into mitochond J. Biol. Chem. 1983, 258, 6667–6670. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.-Z.; Zhu, J.-J.; Yang, F.-F.; Zhang, Y.-P.; Xie, S.-A.; Liu, Y.-F.; Yao, W.-J.; Pang, W.; Han, L.-L.; Kong, W.; et al. DNA methyltransferase 1 and Kruppel-like factor 4 axis regulates macrophage inflammation and atherosclerosis. J. Mol. Cell. Cardiol. 2019, 128, 11–24. [Google Scholar] [CrossRef]
- Rahmel, T.; Marko, B.; Nowak, H.; Bergmann, L.; Thon, P.; Rump, K.; Kreimendahl, S.; Rassow, J.; Peters, J.; Singer, M.; et al. Mitochondrial dysfunction in sepsis is associated with diminished intramitochondrial Transcription Factor A despite increased cellular expression. Sci. Rep. 2020. under review. [Google Scholar]
- Quiles, J.M.; Gustafsson, A.B. Mitochondrial Quality Control and Cellular Proteostasis: Two Sides of the Same Coin. Front. Physiol. 2020, 11, 515. [Google Scholar] [CrossRef]
- Iwanami, N.; Takeshita, K.; Lawir, D.-F.; Suetake, I.; Tajima, S.; Sikora, K.; Trancoso, I.; O’meara, C.; Siamishi, I.; Takahama, Y.; et al. Epigenetic Protection of Vertebrate Lymphoid Progenitor Cells by Dnmt1. iScience 2020, 23, 101260. [Google Scholar] [CrossRef]
- Liu, B.; Du, Q.; Chen, L.; Fu, G.; Li, S.; Fu, L.; Zhang, X.; Ma, C.; Bin, C. CpG methylation patterns of human mitochondrial DNA. Sci. Rep. 2016, 6, 23421. [Google Scholar] [CrossRef] [Green Version]
- Reis, R.J.S.; Goldstein, S. Mitochondrial DNA in mortal and immortal human cells. Genome number, integrity, and methylation. J. Biol. Chem. 1983, 258, 9078–9085. [Google Scholar]
- Shih, C.-C.; Liao, M.-H.; Hsiao, T.-S.; Hii, H.-P.; Shen, C.-H.; Chen, S.-J.; Ka, S.-M.; Chang, Y.-L.; Wu, C.-C. Procainamide Inhibits DNA Methylation and Alleviates Multiple Organ Dysfunction in Rats with Endotoxic Shock. PLoS ONE 2016, 11, e0163690. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, W.; Wang, K.; He, S.; Chen, Y. Predictive value of circulating plasma mitochondrial DNA for Sepsis in the emergency department: Observational study based on the Sepsis-3 definition. BMC Emerg. Med. 2020, 20, 25. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, K.; Kox, M.; Scheffer, G.J.; Pickkers, P. Plasma Nuclear and Mitochondrial DNA Levels, and Markers of Inflammation, Shock, and Organ Damage in Patients with Septic Shock. Shock 2016, 45, 607–612. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mtDNA Region | Forward Primer | Reverse Primer |
|---|---|---|
| D-Loop | CTCGTCCCCATGGATGACCC | TGAAGTAGGAACCAGATGTCGGA |
| TTF | CACCCAAGAACAGGGTTTGT | TGGCCATGGGTATGTTGTTAAG |
| 12S-rRNA | GGTCACACGATTAACCCAAGT | TGTTAAAGCCACTTTCGTAGTCTAT |
| mtND1 | CACCCAAGAACAGGGTTTGT | TGGCCATGGGTATGTTGTTAA |
| 18S-rRNA | TAGAGGGACAAGTGGCGTTC | CGCTGAGCCAGTCAGTGT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koos, B.; Moderegger, E.L.; Rump, K.; Nowak, H.; Willemsen, K.; Holtkamp, C.; Thon, P.; Adamzik, M.; Rahmel, T. LPS-Induced Endotoxemia Evokes Epigenetic Alterations in Mitochondrial DNA That Impacts Inflammatory Response. Cells 2020, 9, 2282. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9102282
Koos B, Moderegger EL, Rump K, Nowak H, Willemsen K, Holtkamp C, Thon P, Adamzik M, Rahmel T. LPS-Induced Endotoxemia Evokes Epigenetic Alterations in Mitochondrial DNA That Impacts Inflammatory Response. Cells. 2020; 9(10):2282. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9102282
Chicago/Turabian StyleKoos, Björn, Eva Lotta Moderegger, Katharina Rump, Hartmuth Nowak, Katrin Willemsen, Caroline Holtkamp, Patrick Thon, Michael Adamzik, and Tim Rahmel. 2020. "LPS-Induced Endotoxemia Evokes Epigenetic Alterations in Mitochondrial DNA That Impacts Inflammatory Response" Cells 9, no. 10: 2282. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9102282