Tissue-Resident Memory T Cells in the Liver—Unique Characteristics of Local Specialists


Division of Gastroenterology, Massachusetts General Hospital and Harvard Medical School, Boston, MA 02114, USA
*
Authors to whom correspondence should be addressed.
Cells 2020, 9(11), 2457; https://0-doi-org.brum.beds.ac.uk/10.3390/cells9112457
Submission received: 8 October 2020
/
Revised: 2 November 2020
/
Accepted: 6 November 2020
/
Published: 11 November 2020
(This article belongs to the Special Issue Tissue-Resident Memory T Cells)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:T cells play an important role to build up an effective immune response and are essential in the eradication of pathogens. To establish a long-lasting protection even after a re-challenge with the same pathogen, some T cells differentiate into memory T cells. Recently, a certain subpopulation of memory T cells at different tissue-sites of infection was detected—tissue-resident memory T cells (TRM cells). These cells can patrol in the tissue in order to encounter their cognate antigen to establish an effective protection against secondary infection. The liver as an immunogenic organ is exposed to a variety of pathogens entering the liver through the systemic blood circulation or via the portal vein from the gut. It could be shown that intrahepatic TRM cells can reside within the liver tissue for several years. Interestingly, hepatic TRM cell differentiation requires a distinct cytokine milieu. In addition, TRM cells express specific surface markers and transcription factors, which allow their identification delimited from their circulating counterparts. It could be demonstrated that liver TRM cells play a particular role in many liver diseases such as hepatitis B and C infection, non-alcoholic fatty liver disease and even play a role in the development of hepatocellular carcinoma and in building long-lasting immune responses after vaccination. A better understanding of intrahepatic TRM cells is critical to understand the pathophysiology of many liver diseases and to identify new potential drug targets for the development of novel treatment strategies.
1. Introduction
T cells play a central role in the immune response against pathogens. CD8+ T cells are highly effective in the eradication of cells infected with pathogens, damaged cells and even cancer cells. CD4+ T cells contribute to and modulate the immune response against pathogens and in the cancer environment. A very important characteristic of the adaptive immune system is to build up a pool of memory T cells, which enables a fast and effective immune response after pathogen re-challenge. These memory T cells patrol in the circulation in order to encounter a known pathogen. In addition to the blood circulation and lymphoid tissues, memory T cells can also be found in non-lymphoid tissues. Within these tissues, they mediate a fast induction of the immune response after the second pathogen challenge and initiate the recruitment of other immune cells by modifying the tissue-specific inflammatory microenvironment. Different scientists were able to detect a persistent tissue-resident memory CD8 and CD4 T cell subpopulation at different tissue sites. These cells can be mainly found in organs that are frequently exposed to pathogens, such as the liver, skin, gut and lung [1]. These resident memory T cells (TRM cells) are known to be important for pathogen surveillance in the respective tissue and have a distinct phenotype in comparison to their circulating counterparts in the blood [2,3]. Interestingly, the tissue itself influences the phenotype of the memory T cell. This review will focus on the development and functional role of CD8+ TRM cells, especially in the liver, and concentrates on the characteristics that make them unique compared to other TRM subpopulations. The liver is an important immunological organ as it is not only affected by many different hepatotropic viruses but is also the first control point of pathogens that enter or re-enter the body through the gastrointestinal tract. Liver tissue contains a diversity of immune cells and interestingly has a special composition of these immune cells in comparison to other tissues or the blood. In order to prevent a systemic infection, the liver plays an important role as a gatekeeper and an effective TRM cell population contributes to effective pathogen clearance.
2. Phenotype and Development of TRM Cells in the Liver
Memory T cell development is an important characteristic of the adaptive immune system and critical for an effective immune response after a second challenge with a pathogen. In order to build systemic protection, circulating and tissue-resident memory T (TRM) cells exist. Based on their expression of cell surface markers and specific functions, circulating memory T cells can be further divided in different subpopulations: central memory T cells (TCM) and effector memory cells (TEM). These memory T cells typically circulate through the body in order to encounter pathogens but were also found in lymphoid tissues. In contrast, TRM cells have the ability to persist within a certain tissue.
2.1. TRM Cell Phenotype
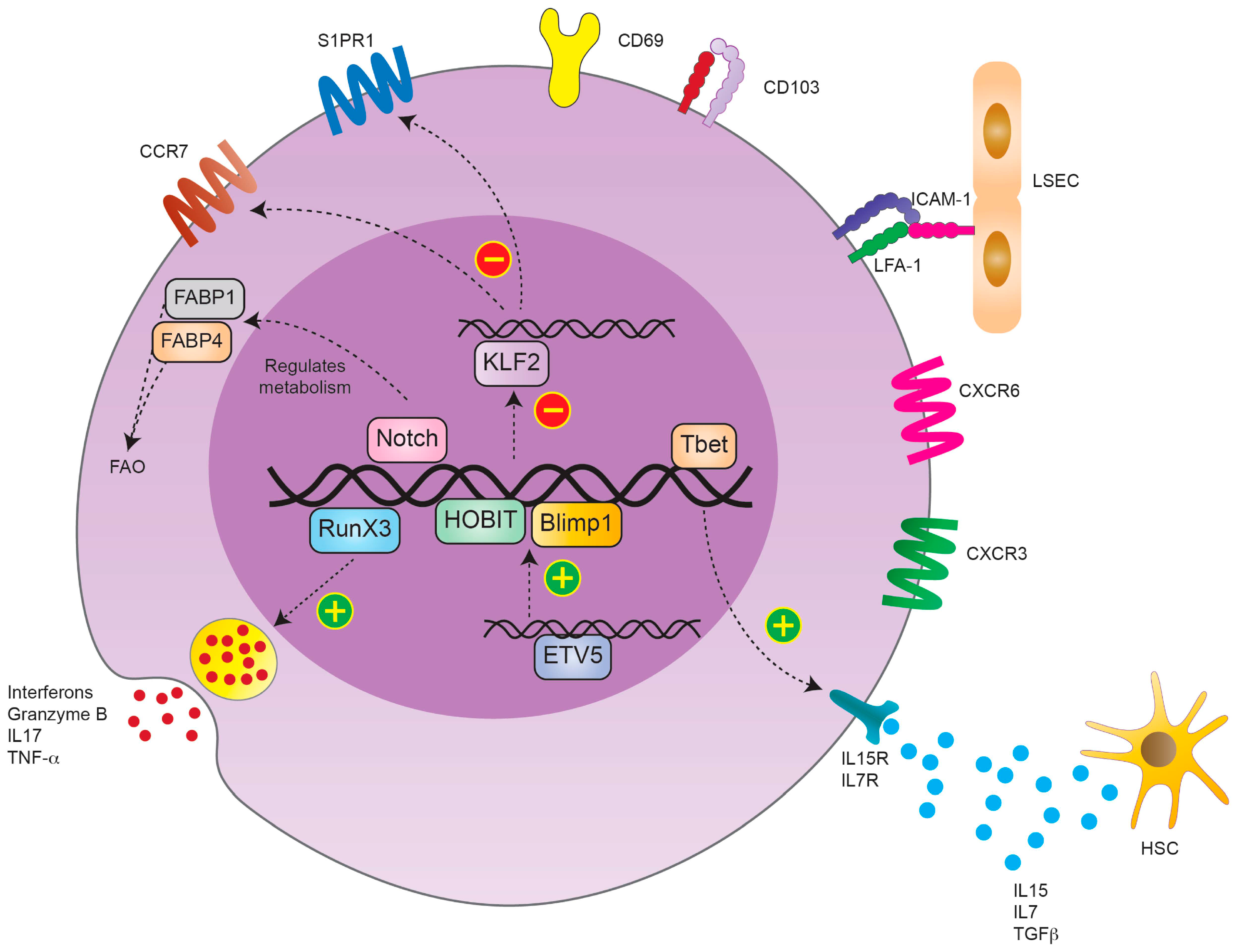
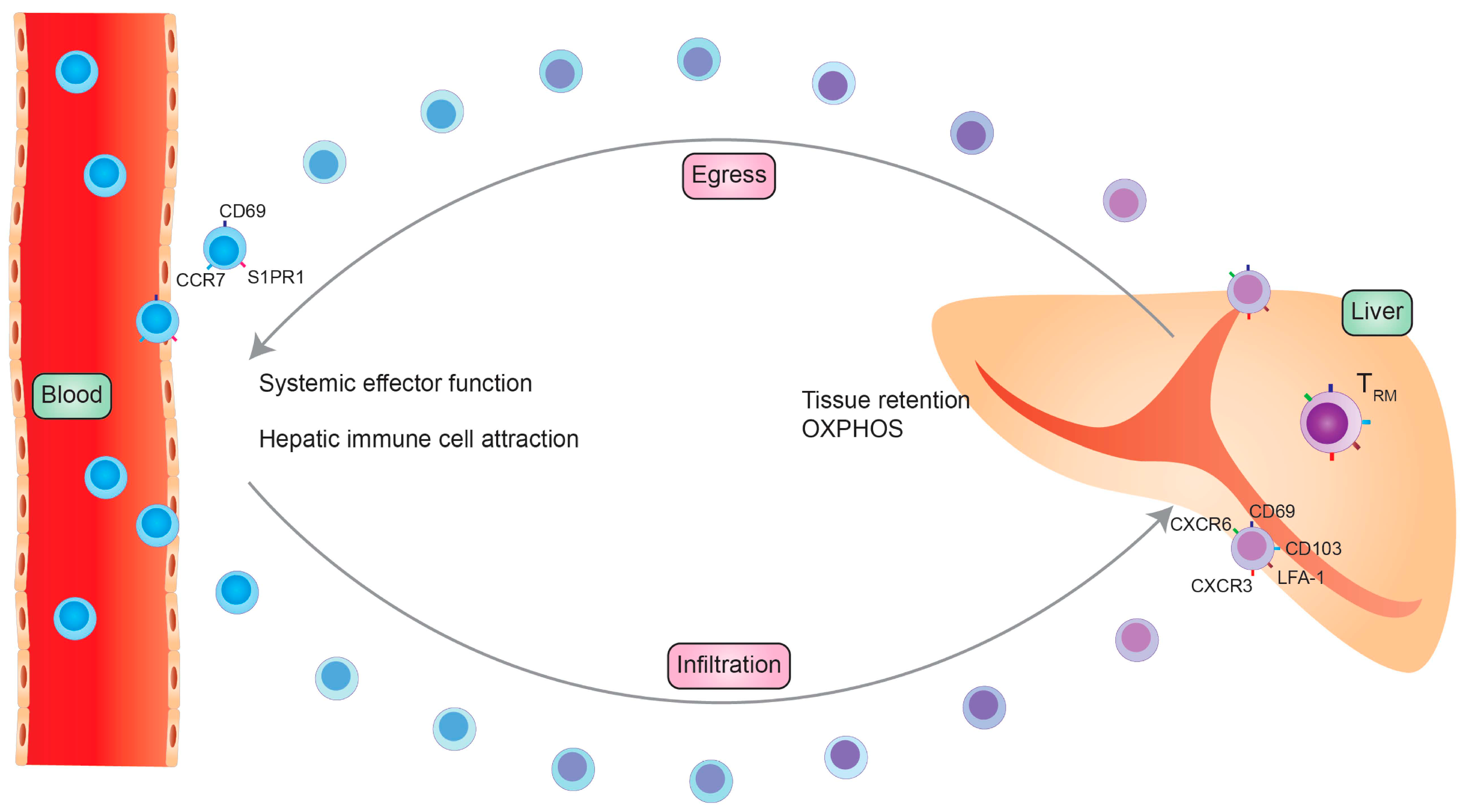
The first detection of TRM cells was possible by analyzing the expression of cell surface markers on a specific memory T cell pool within the tissue. These markers are described to maintain their tissue-specific function. Liver TRM cells usually downregulate the expression of homing receptors such as CCR7 and CD62L. Furthermore, they decrease the expression of tissue egression markers, such as soingosine-1-phosphate 1 (S1PR1), while upregulating CD69. Additionally, liver TRM cells express other adhesions and functional molecules, such as CD103, CD49a and CD44, to enable trafficking within the liver sinusoids and liver tissue [4]. CD69 expression is strongly initiated after T cell activation. Besides being an activation marker, it also mediates T cell homeostasis and cell migration–retention [5]. CD49a is the α1 subunit of the α1β1 integrin, also known as very late antigen-1 (VLA-1). It binds to collagen IV and mediates T cell adhesion. It is therefore important to establish tissue residency [6]. CD44 is another adhesion molecule that is upregulated after T cell receptor (TCR) stimulation and promotes T cell migration [7]. CD103 is the alpha E subunit of the alpha E beta 7 integrin, an adhesion molecule defined as a marker of TRM cells, especially in human tissues [8]. In the liver, on the other hand, CD103 is not expressed in all TRM cells. The CD69+CD103− subpopulation is proposed to be a subpopulation that can recirculate and take on other functions upon pathogen challenge, whereas the CD69+CD103+ are defined to be truly liver resident [9]. In contrast, it could be shown that the majority of CD4+ TRM cells are long living and tissue-resident cells that express high levels of CD69 and show a low CD103 expression [10].
The effector function of TRM requires constant chemokine stimulation. Therefore, liver TRM cells constitutively express the chemokine receptors C-X-C motif chemokine receptor 3 (CXCR3) and C-X-C motif chemokine receptor 6 (CXCR6) on their surface. CXCR3 is an important homing marker and supports the maintenance of TRM cells in the liver tissue. CXCR3 has the ability to bind multiple ligands predominantly secreted by monocytes, endothelial cells and fibroblasts, e.g., chemokines C-X-C ligand 9 (CXCL9), C-X-C ligand 10 (CXCL10) and C-X-C ligand 11 (CXCL11) [11]. CXCR6, on the other hand, is essential for TRM cell development but also supports the maintenance of TRM cells in the liver through binding C-X-C ligand 16 (CXCL16) secreted by liver endothelial cells [12] (Figure 1).
In addition to the surface markers, transcriptional analysis of TRM cells showed that they express a distinct transcription factor profile [13]. TRM cells, including liver TRM cells, upregulate the transcription factors homolog of blimp-1 (HOBIT) and BLIMP1 [14,15]. HOBIT was first described to be upregulated in CD45+ effector T cells after viral infection, e.g., human cytomegaly virus infection (CMV). In TRM cells, HOBIT and BLIMP1 actively downregulate the expression of C-C chemokine receptor 7 (CCR7), Kruppel-like factor 2 (KLF2) and sphingosine-1 phosphate receptor 1 (S1PR1) [16]. CCR7 is highly expressed in naïve cells and TCM. It is the receptor of chemokine ligand 19 (CCL19) and chemokine ligand 21 (CCL21) that convey migration of cells to secondary lymphoid tissues [17]. Opposingly, KLF2 regulates the expression of S1PR1, which directs cells from a specific tissue into the periphery [18]. Therefore, the co-expression of HOBIT and BLIMP1 represses the formation of circulating memory T cells and silences the genes related to cell recirculation into the periphery. In a recent mouse study, Park et al. demonstrated that HOBIT expression in liver TRM cells is mediated by the gene repressor Capicua in collaboration with the ETS variant transcription factor 5 (ETV5), underlining the importance of this transcription factor [19]. In addition to HOBIT and BLIMP1, the transcription factors Runt-related transcription factor 3 (RUNX3), TBX21 (Tbet) and Notch were reported to be upregulated in TRM cells after their development and their expression is essential for a sustained TRM cell population [20]. RUNX3 represses the expression of genes involved in the activation of circulating memory T cells. On the other hand, RUNX3 induces the expression of genes such as integrin subunit alpha E (ITGAE), which encodes for CD103 in TRM cells and mediates the production of granzyme B by TRM cells [20].
Tbet is known to mediate the expression of the IL-15 receptor (IL-15R) in order to establish a long-term lineage stability. The membrane bound transcription factor Notch is predominantly expressed in newly developed TRM cells and responsible for their maintenance through the regulation of their metabolic profile [20] (Figure 1).
2.2. TRM Cell Development
The rather recent identification of TRM cells as a distinct tissue-resident memory T cell subpopulation leads to the question: Which specific factors are involved in their development and maintenance, and which are crucial for their tissue-specific function?
Different models exist to explain the origin and development of memory T cells after a pathogen challenge but the overall development is not fully understood yet [21,22]. It is still unclear whether TRM cells and circulating memory T cells originate from the same precursor cell subset. It is further unclear whether liver TRM cells develop extrahepatic and migrate into the liver or whether they directly differentiate intrahepatic. Adoptive transfer experiments in mice demonstrated that in vitro-activated CD8+ cells can differentiate into TRM cells after transfer into the specific tissue and are not further distinguishable from those generated within the tissue itself [23]. Based on the current literature, which will be discussed in more detail in this review, we presume that both extra- and intrahepatic development contributes to the TRM pool in the liver.
Several factors are known to contribute to T cell development in general and further determine the specificity of a memory T cell and the fate of TRM cells.
2.2.1. Origin of TRM Cells
To answer the question about T cell origin, one important approach is to analyze the TCR repertoire of the cells. In search of the origin of TRM cells different groups analyzed their TCR repertoire and could find that TRM and TEM have an overlapping TCR repertoire, suggesting that these subsets develop from the same progenitor cell [24]. Holz et al. were the first to describe that liver TRM cells also require TCR stimulation and rearrangement upon binding to a specific antigen for their formation [25].
In addition, TCRs differ in their strength of antigen binding, which influences the development of effective CD8 memory T cells. A low affinity TCR stimulation leads to insufficient memory T cell development of cells with a short lifespan and thus leads to impaired secondary immune responses. Furthermore, the strength of TCR binding varies between different memory subsets, e.g., the binding strength has to be especially high for TEM development. However, the specific strength of TCR stimulation for the development of liver TRM cells has not been described yet but the described overlap with the TCR repertoire of TEM cells indicates the potential requirement of high-affinity binding [8].
Another important marker to investigate the origin of TRM cells is the killer cell lectin-like receptor G1 (KLRG1), which is usually upregulated in effector T cells and relatively low expressed in memory precursors of circulating memory T cells.
Mouse studies could demonstrate that approximately 50% of the intrahepatic TRM cell population develops from a KLRG1-expressing precursor whereas the other 50% develop from precursors that only transiently express KLRG1 [26]. These findings indicate that intrahepatic TRM cells develop from two different origins. Cells simultaneously develop extrahepatic from memory precursors and migrate into the liver and another proportion directly develops within the liver from effector T cells. TRM cell tissue dependency might even be less strict as it was recently shown that a certain subset of TRM cells can leave their resident tissue and re-circulate into the blood (Figure 2). These previous TRM cells expresses a similar transcription factor repertoire and surface molecule pattern as their tissue-resident counterparts.
These findings suggest that although TRM cells might be able to reside in a specific tissue for a long period of time, they are able to recirculate if needed [27]. A recently published study by Pallett et al. could nicely demonstrate the longevity of human liver TRM cells. They could demonstrate that, in liver allograft tissue, 2–6% of CD8+ T cells have a donor-derived TRM phenotype and were detectable 11 years post transplantation. Additionally, they showed that recipient CD8+ and CD4+ T cells could develop a TRM phenotype although the recipient-derived TRM cells tended to express fewer liver-resident markers. Furthermore, a small population of donor-derived CD4+ TRM cells was detectable [10]. To conclude, there are strong indications that liver TRM cells can develop from two different origins (Figure 2).
2.2.2. Factors Influencing/Driving TRM Cell Development
Within the tissue, TRM cells can be exposed to a variety of environmental conditions that influence their development, maintenance and function, e.g., nutrient deprivation, hypoxia and the inflammatory cytokine milieu [20].
Certain cytokines are described to promote TRM cells differentiation. One important cytokine is the tumor growth factor β (TGF-β), which induces the expression of CD103 on the cell surface [28]. Furthermore, monocyte-derived IL-10 facilitates the TGF-β release and promotes CD103 expression on TRM cells. Blocking of IL-10 decreased the CD103 expression [29].
In addition, interleukin-7 (IL-7) and IL-15 are involved in TRM development and longevity [25]. The importance of these cytokines is shown by the upregulation of the IL-7 and IL-15 receptors in TRM cells. Thus, it could be shown that IL-15 stimulation is especially essential for hepatic TRM development as IL-15−/− mice were prevented from developing these cells in the liver [25]. Interestingly, the requirement to the cytokine microenvironment differs between CD4 and CD8 TRM cells. CD8 TRM cells require IL-15 and IL-7 stimulation, whereas CD4+ TRM stability only depends on IL-7 signaling [30].
In addition, the pro-inflammatory cytokines type 1 Interferon (IFN) and IL-12 are described to positively influence TRM cell development by inducing the expression of CD103 and CD69 [27].
Another cytokine that functions as an autocrine stimulus on CD8 TRM cells is IL-2. Pallett et al. showed that human liver CD8 TRM cells produce high levels of interleukin-2 (IL-2), which was shown to be critical for intrahepatic TRM cell survival, the ability to function and for their antigen-specific proliferation [11]. Increased IL-2 expression could also be shown in response to oxidative stress induced by stimulation with reactive oxygen species and free radicals, which is a common characteristic of chronic liver diseases such as non-alcoholic steatohepatitis (NASH) [31]. Thus, oxidative stress induction could alter TRM cell function and differentiation.
Comparable to many other T cell subsets, also the metabolic profile influences TRM cell development, maintenance and function. Different T cell subsets preferentially use a certain metabolic pathway to gain their energy. In general, highly proliferating and active cells favor glycolytic pathways, whereas quiescent cells mainly use oxidative phosphorylation and FAO to generate ATP. A key regulator of T cell metabolism, proliferation, activation and survival is the transcription factor mammalian target of rapamycin (mTOR) [32]. mTOR consists of two subunits, mTOR complex 1 (mTORC1) and mTORC2. Both complexes can be activated in T cells by various stimuli like TCR stimulation, cytokine ligation and the presence or absence of certain metabolites. mTOR activation in turn leads to a downregulation of S1P1 and KLF2 in activated T cells.
mTOR can further induce glucose consumption to support T cell proliferation and thereby favors the development and tissue retention of TRM cells [33]. The importance of mTOR signaling in TRM cells could be demonstrated by using rapamycin. Rapamycin is an mTORC1 inhibitor that was shown to reduce TRM formation in mucosal tissues by inhibiting CD103 and CCR9 upregulation in TRM cells and cell migration. The effect of rapamycin was tissue-specific for the gut-associated lymphoid tissue and the exact implications on the liver are still under investigation [34].
More important and better investigated is the impact of fatty acid oxidation (FAO) on TRM cells. It could be demonstrated that TRM cells rely on FAO to gain their energy. In order to take fatty acids up, TRM cells upregulate fatty acid-binding proteins (FABP). TRM cells from different tissues express distinct FABPs with varying fatty acid specificity, depending on the specific tissue of origin [35]. It could be demonstrated that liver TRM cells express mainly FABP1 and FABP4 in a low concentration but do not express FABP5. Thereby, the expression of specific FABPs are upregulated during TRM development and maturation in a tissue-dependent manner to optimize the usage of local fatty acids. Frizzell et al. were able to stress the importance of FAO in a mouse model of LCMV infection. FABP1 deficiency in infected mice was associated with impaired liver TRM cell development [36].
Additionally, liver TRM cells express high levels of P2X purinreceptor 7 (P2RX7). P2RX7 is a sensor for extracellular ATP (eATP) and is involved in various inflammatory processes [37]. Studies suggest that P2RX7 promotes oxidative phosphorylation and FAO and thereby regulates the metabolic function of CD8 TRM cells. P2RX7 deficiency, on the other hand, prevents a stable and durable TRM cell development and reduces the expression of anti-apoptotic molecules. Furthermore, P2RX7 expression was shown to positively influence IL-7 secretion and IL-15 receptor expression and thereby influences TRM cell development [37,38].
Another characteristic of the liver is the presence of hypoxic regions due to its venous blood supply from the portal vein in combination with slow blood flow in the sinusoids. Interestingly, T cells have the ability to adjust the expression of the hypoxia inducible factor 1α (HIF-1α) and HIF-2α as they detect the local oxygen gradient [39]. HIF-1α and HIF-2α are transcription factors that play a crucial role in the cellular response to low oxygen concentration, especially T cell development, metabolism and function.
The finding that an intrahepatic subpopulation of TRM cells can be found that expresses CD69+CD103− and HIF2α suggests that they are predominantly located in hypoxic regions within the liver. Interestingly, this TRM cell subpopulation could not be located in other tissues such as the lung, skin or colon [40]. This finding underlines the importance of tissue-specific adaptions of TRM cells to unique environmental conditions in different organs.
3. The Phenotype and Transcriptional Profile of Liver TRM Cells in Mouse and Man
Blood is the major human sample type for the study of T cell immunology because it is easily available. Human tissue samples, especially liver tissue, can just be obtained from living individuals most often suffering from end-stage liver disease and is limited to invasive tissue sampling through surgical resection, biopsy or more recently fine needle aspirates (FNA). This lack of sample disposability lead to extensive investigations of liver TRM in different rodent models. Various in vivo studies, and not only on their differentiation, maintenance and function but also on their phenotypic and transcriptional features, have been conducted in infection mouse models such as hepatitis B virus (HBV) or malaria.
TRM are phenotypically characterized by their expression of CD69 ± CD103. Transcriptional analysis of these cells in different human tissues, such as skin, lung and liver, revealed that they exhibit a distinct profile, discriminating them from circulating memory T cells. This profile is also mirrored in its key features in mouse TRM cells, making them a suitable model for the investigation of these cells in health, disease and autoimmunity. However, the translational potential of animal models, especially in the course of inflammatory disorders, is still under debate. Although they are important to investigate basic mechanisms, points of criticism are especially that animal models do only partially mimic the complex features and length of chronic injury as they naturally occur in humans. Furthermore, the variety of exposures to different pathogens and environmental influences over the whole disease period hardly can be reflected in any rodent model. Nevertheless, the impact of how much these differences affect the transferability of the basic findings obtained in rodent models, on general and tissue immunity in men, is still unknown. To overcome some doubts, recent studies were performed with so called outbred or dirty mice purchased from pet stores. The commonly used inbred mouse strains are used for basic research due to their comparability based on the removal of genetic variability. Outbred mice in comparison show a higher genetic and phenotypic diversity, which does better parallel or even exceed the human variability. Therefore, these animals have been lately used in different disease models, e.g., when investigating the development and tissue specificity of memory T cells.
It was found that these dirty mice better reflect the human situation especially with regard to homeostatic phenotypical manifestations of TRM cells and may therefore be a conceivable addition and validation to the typically used inbred mouse models. As the microbiome is supposed to have a particular impact on TRM cell development, genetic and phenotypic variability is especially important to investigate this subpopulation. Anyhow, it is yet unclear to what extent these models mirror the human immune reaction in disease [41,42].
3.1. Experimental Models
TRM cells are characterized by their continued presence at a specific tissue site, independent from their circulating counterparts. A variety of experimental animal models was used not only to define the phenotypic manifestations of tissue residency but also to determine the potential of these cells to recirculate into the periphery. Mouse models include parabiosis, the direct in vivo labeling of cells with different antibodies; T cell depletion, the transplantation of specific tissues; and direct profiling.
Parabiotic surgery describes the conjunction of the blood circulation of two different mice. One animal got previously infected to induce the development of TRM cells, whereas the other animal is pathogen naive. In addition, animals are distinguished by their genetic CD45+ cell isoform (CD45.1 and CD45.2). This allows to investigate the circulation and migration potential of T cells and to define the phenotypic and transcriptional characteristics of TRM cells between parabionts. Circulating blood T cells usually attain homeostasis between parabionts within one week. TRM cells on the other hand do not exchange between the donor and recipient and reside in the specific tissue of each animal. This technique made it possible to define and investigate TRM cells as a distinct cell population. However, results obtained in parabiosis have to be critically evaluated as the inflammation caused as a consequence of the surgery itself may recruit circulating TRM cells from the specific tissue sites, blurring the differences between the tissue-resident and circulating TRM cell subsets [43,44,45].
The in vivo labeling of circulating cells with fluorochrome-conjugated or depleting antibodies, such as anti-CD90/Thy-1, is another approach to distinguish circulating from tissue-resident T cells to investigate TRM cells in mice. CD90/Thy-1 is expressed on peripheral T cells, thymocytes and is widely used for the depletion of T lymphocytes. Via intravenous injection of these antibodies all circulating cells get labeled or depleted whereas those residing in a specific tissue niche stay unaffected. This allows a really fast and easy localization and investigation of TRM cells in general, although there are some disadvantages when studying TRM cells of the bone-marrow and in liver sinusoids as the bone-marrow compartment is not targeted by depleting antibodies [46,47]. Especially in the liver, reports suggest that the sinusoids contain a fraction of sinusoidal resident CD8+ T cells, which are affected by the antibody depletion applied to the vasculature but have slightly different features from other liver TRM cells [48,49]. A related technique is the treatment with FTY720.
FTY720 is a sphingosine 1-phpsphate receptor 1 agonist leading to peripheral lymphopenia and preventing the transmigration of TRM cells back into the circulation [50]. This allows the investigation of liver specific TRM cells as they are retained within the tissue while at the same time a systemic lymphopenia is induced.
Additionally, solid organ transplantation is another way to study TRM. Mouse experiments transplanting the intestine or parts of the skin gave interesting insights into the longevity, residency and recirculation potential of TRM [3,51].
Most of the presented techniques are not applicable in humans, making the investigation of the phenotypic and transcriptional appearance of TRM cells difficult. However, few clinical approaches such as T cell depletion therapy and organ transplantation allow us to study TRM cells in human tissues. Anti-CD52 treatment, for example, is commonly used as therapy in T cell lymphoma and leads to the targeted elimination of circulating T cells without affecting tissue-resident CD4+ and CD8+ T cells [51]. A very recent study from Pallett et al. was able to investigate HLA-mismatched liver transplant patients, allowing to discriminate liver-resident (donor) from newly infiltrating (recipient) T cells. Allogenic liver tissues were rapidly infiltrated by recipient T cells that underwent reprogramming to express the TRM markers CD69, CD103 and CXCR3hi, which is important for intrahepatic retention [10]. Interestingly, long-lived liver-resident donor TRM cells could even be detected after more than a decade (2–6% of intrahepatic CD8+ T cells) and it was found that they do not egress into the recipient circulation via the hepatic vein. This long-lasting tissue maintenance could be also shown for TRM in other tissues. In mice, TRM cells are maintained for several months in tissues such as the skin, intestine, lung and brain. Human TRM cells, in contrast, are maintained in their tissue-specific niche for several years [52]. Some TRM subpopulations were even found to be maintained over an entire life span [53,54]. Comparable findings of tissue-specific longevity could be shown for human intrahepatic NK cells while TRM cells in the intestine and lung were described to be maintained in the tissue for at least 1 year after transplantation [55,56,57].
3.2. The Phenotypical and Transcriptional Differences of Liver TRM in Mouse and Man
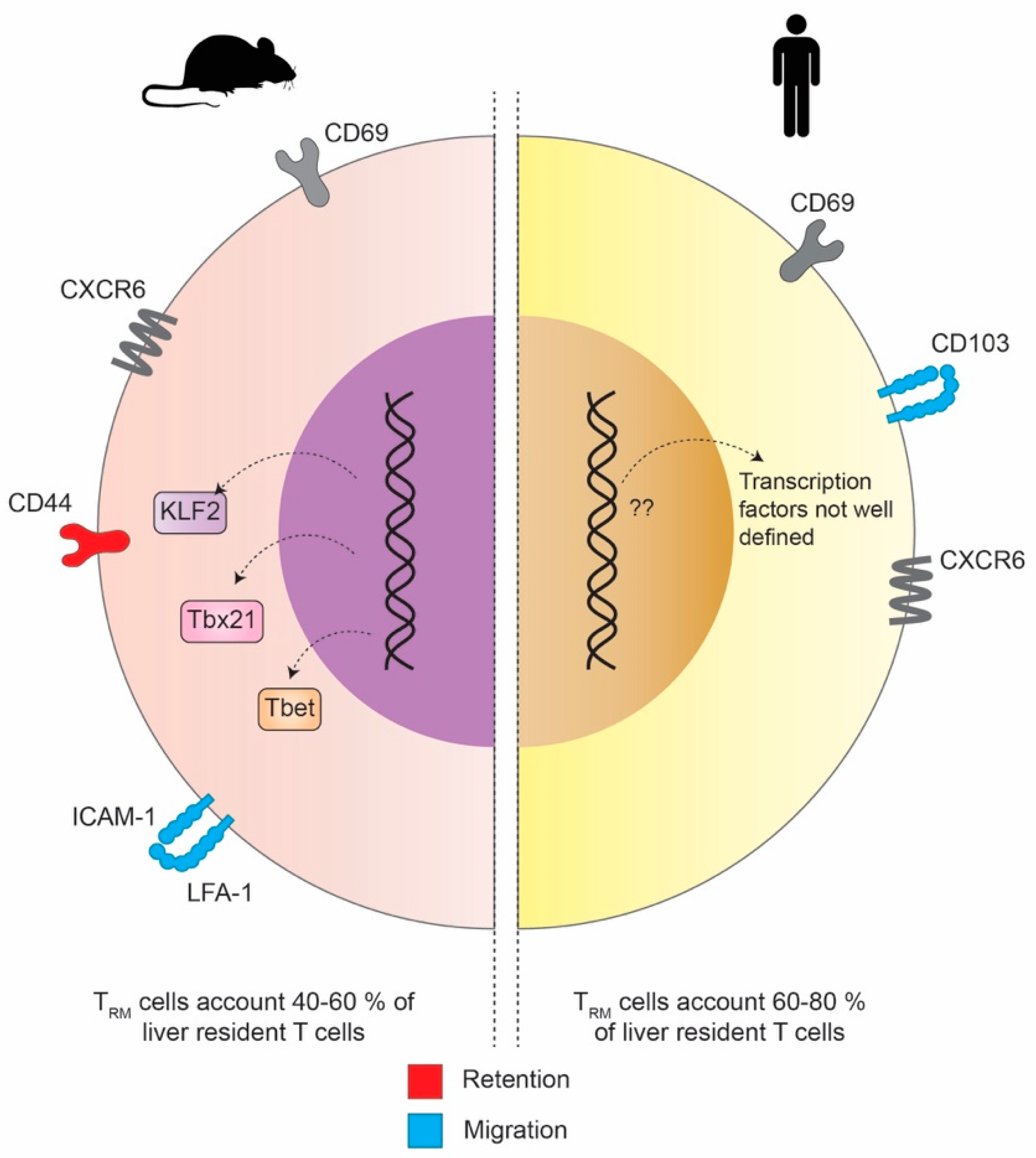
The description of tissue-specific TRM as a distinct T cell subset started about 10–15 years ago. Since then, groups have tried to distinguish these TRM from their circulating counterparts in both humans and mice. The main marker to define these cells is CD69, which was originally described as marker for early T cell activation. However, it is constitutively expressed by almost all liver-resident CD4+ and CD8+ memory T cells across species and serves as a canonical signal for tissue retention [18]. Interestingly, the depletion of CD69 in mice showed an alternative, CD69-independent way of TRM formation.
Animals indeed displayed significantly reduced CD8+ TRM in skin and lung but CD4+ TRM formation was not affected. Given the fact that the liver generally has a larger amount of CD8+ than CD4+ T cells it seems likely that the majority of liver TRM are defined by their expression of CD69.
The most interesting and striking difference between hepatic TRM in the mouse and human is the expression of CD103. Although it is expressed by different subsets of mouse and human CD8+ TRM, e.g., in tissues at barrier sites [58], CD103 is not expressed in intrahepatic CD8+ TRM in mice while human liver CD8+ TRM can be either CD69+/CD103+ or CD69+/CD103− [59]. On the other hand, both mouse and human CD8+ TRM express CXCR6, a chemokine receptor that binds CXCL16. It serves as another important marker promoting the establishment for both human and mouse CD8+ TRM, especially in the skin and the liver (Figure 3) [12,60]. In mice, the transcriptional landscape of liver TRM gets more and more defined and understood. McNamara and colleagues were further able to show in more detail what molecular interactions of adhesion molecules are responsible to retain CD8+ T cells the liver tissue but at the same time allow their special patrolling function within the liver sinusoids. The initial migration of CD8 T cells into the liver was found not to be superficially mediated by the binding of selectins but rather mediated by interactions with platelets that bind to endothelial cells via CD44 [61]. Recent studies proposed that the intracellular adhesion molecule 1 (ICAM-1)/lymphocyte function associated antigen 1 (LFA-1)-axis is required for the retention of naïve and activated CD8+ T cells and NKT cells in the liver upon antigen presentation [62,63]. With intra-vital imaging it could be shown that ICAM-1/LFA-1 interactions are crucial for CD8+ TRM cells to move along the liver sinusoids and within liver tissue. Hence, the depletion of LFA-1 in mice lead to the disability to develop intrahepatic residence. It seems that, in mice, LFA-1 but not CD103 is responsible for the retention of intrahepatic TRM cells and stresses that the distinct expression of different adhesion molecules is crucial for TRM cells to patrol within specific tissues rather than at barrier sites [64].
However, the transcription factors that drive the formation of TRM in humans remain not well defined yet and is a topic of current investigations. In this context, Pallett et al. were able to identify IL-2 as a marker that is highly expressed by long-lived liver-resident CD8+ TRM cells. They could further show that this holds true for either global or virus-specific intrahepatic TRM [11].
When comparing the compartmentalization and tissue niches of TRM in mice and humans, another difference can be found. While in mice the non-circulating liver TRM account for 40–60% of the liver-resident T cells, this amount is significantly higher in humans, where it ranges between 60 and 80%.
4. Liver TRM Cells—In Health, Disease and Vaccination
Memory T cells play a fundamental role in the immune response against a recurring pathogen but are also described to be crucial in cancer immunology and autoimmunity. The overall understanding of TRM cells is still under investigation. A significant characteristic of TRM cells is that they can remain in a distinct non-lymphoid tissue without recirculating into the blood [3,65]. To perform their particular function—to clear recurring pathogens and quickly initiate a confined immune response—the location in the tissue is of special importance. Usually, TRM cells are hardly found intravascular, not even in highly vascularized organs. One important exception is the liver, where one fraction of TRM cells can be found in the vascular space of liver sinusoids [46]. The fenestrated epithelial layer, which is a unique characteristic of the liver, allows a direct interaction of TRM cells with hepatocytes [47,66]. Although TRM cells are found intravascular, the main fraction persists within the liver tissue. In order to fulfill their border patrol function, migration within the tissue is important to encounter the cognate antigen and requires a close interaction with hepatocytes and other liver-resident immune cells. The average speed of the liver TRM migration is relatively fast in comparison to other TRM cells in other non-lymphoid tissues with an average speed of 5–7 µM/min, representing their efficiency and activity. Thus, in comparison to skin-resident TRM cells, liver TRMs are very motile. The average speed of their migration in comparison to non-tissue-resident lymphocytes, on the other hand, is very slow because TRM cells stop repeatedly to establish an immunological synapse. To conclude, the slow kinesis of TRM cells enables the local immune-surveillance and promotes their function [58].
In case of a re-infection, circulating memory T cells contribute to the effective immune response against the pathogen. The re-stimulation in the draining lymphoid organs and migration of these cells into the liver takes several days and mainly promotes TCM cell development. In contrast, circulating TEM cells are recruited into the liver by proinflammatory signals within hours. Thus, the initiation and direct response is created by TRM cells and is essential to avoid pathogen spread and a systemic infection. After encountering their cognate antigen within the tissue, hepatic TRM cells need to rapidly proliferate and provide an effector function in order to resolve the re-infection. After activation, CD8+ TRM cells produce IFN-γ and TNF-α and have the ability to directly lyse target cells. The pro-inflammatory cytokine expression on TRM cells is elevated in comparison to the circulating memory T cells, indicating their efficient effector function at the tissue-site of infection. Additionally, CD8+ TRM cells recruit other immune cells by chemokine production after antigen recognition [67].
They express high levels of CCL3, CCL4 and CXCL1 and can induce the chemokines CXCL9 and CXCL10 even in an IFN-γ-dependent manner [15]. These chemokines contribute to the migration and expansion of neutrophils and monocytes in the liver [68,69]. In addition to the local inflammation, TRM cells can induce a systemic immune response. This could be demonstrated in vaccine studies, which demonstrated a systemic immune response signaled through pro-inflammatory chemokines produced by TRM cells [70]. Schenkel et al. could display that TRM cell quantity increases in the draining lymph nodes after re-infection, which indicates a support of the systemic immune response [71].
In order to efficiently protect against pathogens, liver TRM cells express high IL-2 levels, which allows them to quickly expand after an antigen challenge and maintain their proliferation in homeostatic conditions [11,72]. In comparison to CD8+ TRM cells, the frequency of human CD4+ TRM cells in the liver is relatively low, also potentially due to a reversed CD8/CD4 ratio compared to the blood [73].
Other resident T cells were shown to complement TRM function in the course of pathogen infection. One of the subpopulations are γδ T cells. Usually, the TCR receptor consists of an α and β subunits but a small proportion of T cells express a γδ TCR. These 3–5% of the intrahepatic lymphocytes are called γδ T cells. Although the concrete TCR recognition mechanism is not fully elucidated yet, γδ T cells mostly recognize lipid antigens in the liver, which are presented by hepatocytes. Furthermore, they can sense extracellular stress factors by multiple receptors, e.g., Toll-like receptors (TLR). After activation, γδ T cells produce a broad variety of pro-inflammatory cytokines, such as IFN-γ, TNF-α and IL-17, and chemokines, e.g., RANTES, IP-10 and lymphotactin. Additionally, they express perforin, granzyme and TRAIL and can thereby directly cytolyze infected cells. After a pathogen challenge in the liver, the γδ T cells mainly produce IL-17 [74]. Additionally, the CD8αα γδ T cells exist and are liver-resident cells. CD8αα binds to the class 1 MHC molecule H2-Q10 expressed on hepatocytes and thereby controls their activation and development [75]. In conclusion, γδ T cells are another liver-resident T cell subpopulation that support TRM cell function in the course of pathogen infection.
In addition, natural killer cells (NK cells) and NKT cells contribute to the pro-inflammatory immune response against pathogens and against cancer cells within the liver. NKT cells are a cell population that combines T cell and NK-cell markers and functions. NK cell and NKT cell subsets are enriched in liver sinusoids and complement the immune surveillances function against pathogens and toxins that reach the liver mainly through the portal vein from the gut [76]. Interestingly, liver-resident NK and NKT cells display a higher function and cytotoxicity in comparison to their circulating counterparts [77]. Similar to TRM cells, NK and NKT cells play a role in chronic liver disease, such as liver fibrosis, hepatocellular carcinoma (HCC) and viral hepatitis. In some diseases, they complement TRM cell function, whereas in others they have an opposing function. One challenge is to analyze NKT cells in non-lymphoid tissues. Inhibition of P2RX7 signaling could be a strategy to restore their function after tissue isolation in order to quantify their frequency in healthy and infected liver tissues [78]. By using this strategy, the investigation of the importance and function of the NK-T cells could be improved.
The interaction of the innate and adaptive immune system in various diseases is tremendously important. It could be shown that modern HCC treatment strategies upregulate the Toll-like receptor (TLR) signaling pathways [79]. This general immune activation could be the link to TRM cell development in this treatment strategy by the secretion of cytokines such as IL-10 [29]. In vaccine studies in the lung against influenza viruses, adjuvants, which are TLR agonist, induce an effective polyfunctional T cell immunity. Nevertheless, the induction of CD69 and CD103 expression was similar between different adjuvants [80]. A direct activation and differentiation of TRM cells by TLR activation could not be shown in the liver yet. It is known that TLR activation occurs in the liver subsequent to the leaky gut syndrome in several chronic inflammatory diseases. Therefore, the investigation of alterations in TRM differentiation and function by TLR signaling is a very important topic for future investigations.
4.1. Liver-Resident T Cells in Viral Infection
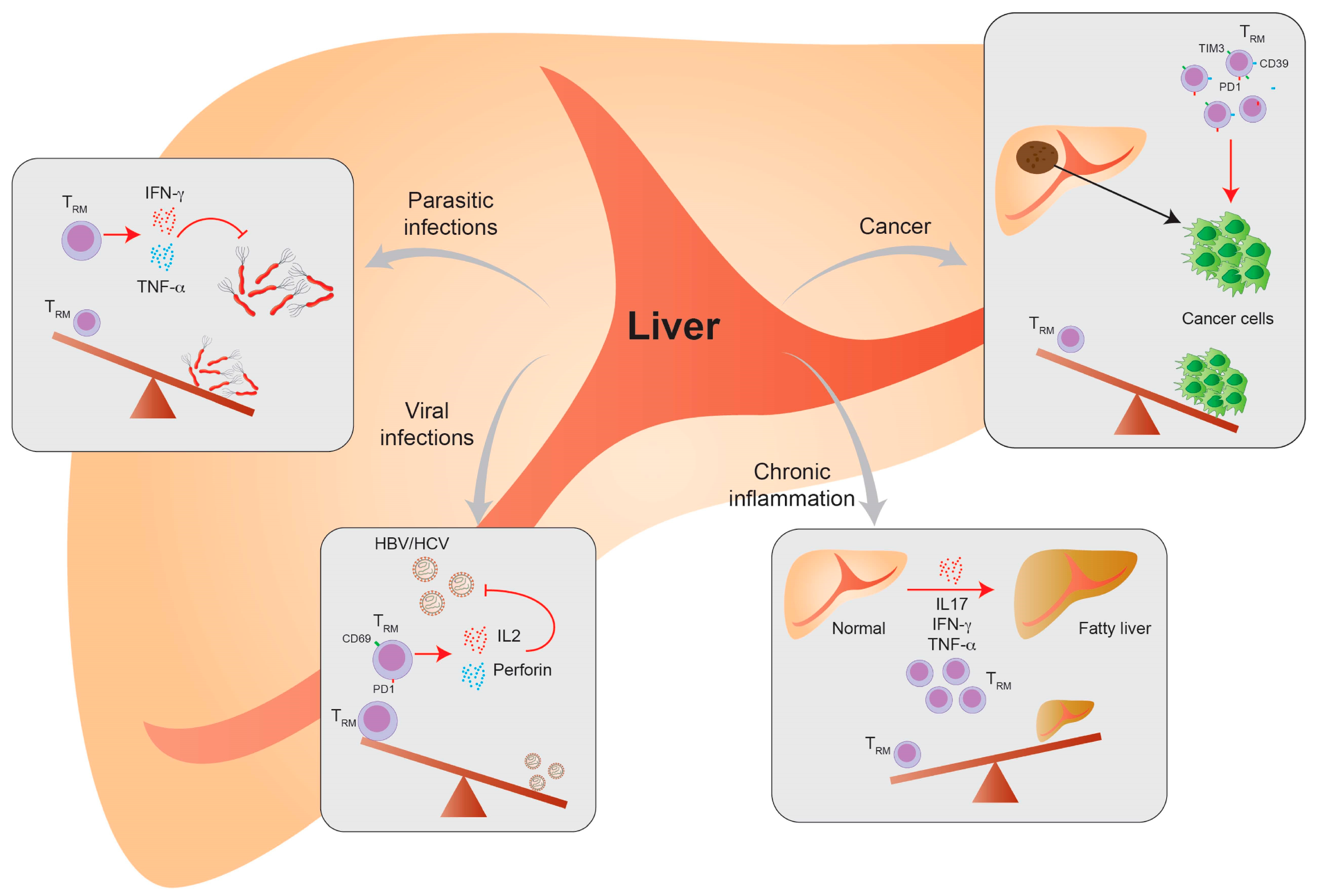
The course of hepatic virus infection can either be acute or progress to a chronic condition. Chronic infection with hepatotropic viruses can cause tissue damage and lead to liver cirrhosis, liver failure and the development of HCC. Chronic viral infection by the hepatitis B (HBV) and hepatitis C (HCV) virus is one of the main causes of HCC development together with ASH and NASH. Liver TRM cells play a major role in promoting a sufficient antiviral response during viral infection and are especially important during chronic infection. CD8+ TRM cells control the viral replication and generate long-lasting viral protection. CD8+ TRM cells can persist in the liver several years after primary infection and can be enriched in chronically infected patients. Depletion of liver CD8+ TRM cells in HCV re-infection mouse models prolonged the virus persistence and prevented effective viral clearance. Furthermore, recovery of liver CD8+ TRM in the same model lead to virus eradication [81]. In human studies, it could be shown that TRM cells are highly increased in chronic HCV-infected patients and these CD69+ cells have a non-naïve and effector memory phenotype. Likewise, these cells express a specific activation and functional phenotype and are important in controlling chronic HCV infection [82].
The importance of virus-specific TRM cells is even more investigated in HBV infection. In chronic HBV infection, a specific liver CD8+ TRM cell subpopulation could be specified. It could be shown that their abundance was negatively correlated with the virus titer. In addition, TRM cells are enriched in HBV patients who reached viral control in comparison to healthy patients—by having the same frequencies of total T cells in the liver [81]. That might indicate that CD8+ TRM cells contribute to a functional cure for HBV infection. A functional cure in HBV means that the continuous transcription of persistent virus DNA—cccDNA—is controlled by the immune system. Furthermore, virus-specific TRM cells were still detectable in spontaneously recovered HBV-infected patients, stressing the role of TRM cells in long-term viral control. In contrast, TRM cells did not differ in chronic HBV-infected patients regarding their liver damage and viral control status, represented by their specific antigen and antibody response [11]. Liver CD8+ TRM cells are very effective in their function, indicated by secretion of pro-inflammatory cytokines. They also express high levels of IL-2 and perforin, showing their ability for effective direct killing, expansion and proliferation. Their high IL-2 expression further contributes to overcome PD-1L-mediated inhibition and exhaustion, mainly in CD69+CD103+ TRM cells [83]. PD-1L expression is upregulated in liver sinusoidal endothelial cells and hepatocytes upon viral infection. TRM cells highly express the exhaustion markers CD39 and PD1 and interact with PD-1L on intrahepatic cells, which can dampen the pro-inflammatory TRM cell response.
Virus-specific T cells are the key cells for an effective antiviral response in HBV infection. It could be shown that approximately 90% of them have a TRM cell-like phenotype (CD69+CD103+ or CD69+CD103−) [11]. By analyzing an allograft of a donor with HBV infection, it could be demonstrated that donor-derived, virus-specific TRM cells persist for a very long time. In addition, the recipient also develops virus-specific TRM cells that were detectable within the donor-derived liver tissue and interestingly also in the blood, albeit in a lower quantity [10]. In HBV-related HCC, CD8+ TRM cell enrichment could be correlated to an improved prognosis [84]. Thus, TRM cell expansion might be a potential therapeutic target in chronic HBV infection and HCC treatment (Figure 2).
Lymphocytic choriomeningitis virus (LCMV) can cause a systemic acute and chronic infection in mice. During LCMV infection, it could be demonstrated that virus-specific TRM cells in the liver could be influenced by other liver-resident immune cells in order to influence effective pathogen clearance. The PD-1/PD-1L interaction between NK and TRM cells negatively influences TRM-dependent cytokine expression in acute and chronic LCMV, and in adenovirus infection. PD-1L inhibition prevented the NK cell-mediated TRM cell inhibition and improved viral clearance [85]. In the course of LCMV infection, γδ T cells contribute to pathogen clearance by producing IFN-γ and TNF-α, migrate and expand at the site of infection [86].
By analyzing CMV-specific TRM cells in human liver allografts, it could interestingly be shown that CMV-specific T cells in humans did not acquire a TRM phenotype in the liver. This phenomenon could be explained by the lack of antigen within the liver [10].
Taken together, the role of TRM cells in viral clearance is not fully elucidated yet.
4.2. Liver-Resident T Cells in Parasite Infection
In addition to viral infections, TRM cells play a key role in the infection with parasites. Not many studies exist investigating the role of TRM cells in this context.
Leishmaniasis is a parasite induced disease and can be transmitted by a certain group of sandflies. The clinical manifestation of the disease can evolve in a cutaneous, mucocutaneous and visceral manner. Different parasite types exist in which Leishmania infantum mainly infects the liver. It can cause acute and resolving hepatic infection, whereas parasite persistence occurs in the spleen. During chronic parasite infection the immune system is not able to fully clear the infection but forms a granuloma to control it. The tissue damage in the liver is relatively limited due to an effective immune response by TRM development. Liver TRM cells are generated and are described to play a protective role during this infection. Thus, it could be shown in different mouse models that circulating memory cells do not provide an effective viral control [87]. Apart from the liver, TRM cells also develop in the skin or other sites of infection. Interestingly, they can also be found far away from the primary site of infection. Liver TRM cells thereby play a special role as their development is especially induced by the Leishmania proteins LirCyp1 and LirSOD and the strategy to induce liver TRM cells is used in vaccine development strategies [87]. The importance of TRM cell induction in vaccine development could be shown in HPV strategies. The vaccine led to an induction of effective resident T cells [88]. An amplification of the TRM cell response by TLR activation could not be demonstrated.
More importantly and well-studied is the role of TRM cells during malaria infection. Malaria is caused by Plasmodium parasites transmitted by Anopheles mosquitos. Plasmodia have a complex life cycle, involving stages in the liver, blood and mosquito. Interestingly, Plasmodium infection promotes TRM cell development in the liver [66]. During Plasmodium infection T cells first get activated in the spleen and form a memory pool that resides in the liver and builds the front line against invading sporozoites. It could be shown in mouse models that circulating memory cells play a role in parasite control but TRM cells are even more protective and modulate the effective immune response. TRM cells mediate cell cytotoxicity and produce pro-inflammatory cytokines, e.g., IFN-γ and TNF-α. TRM cells were shown to patrol in the liver sinusoids to encounter malaria antigens and display a different and more effective migration pattern than other memory T cells. The importance of TRM cells during malaria could be demonstrated in a P. knowlesi infection model, where TRM cell depletion abolished a protective and competent immune response [59].
The development of protective TRM cells in Plasmodium infection is used in vaccine development. Thus, a recently published study by Holz et al. used a glycolipid-peptide vaccination. They were able to induce intrahepatic Plasmodium-specific memory T cells with a TRM phenotype. A second dose of the vaccination even increased the frequency of these cells in the liver. TRM cells were effective in pathogen clearance and the vaccine protected mice from an infection by Plasmodium berghei. The induced TRM cells showed a half-life of 425 days and retained protective function in 90% of the animals up to day 200 [89].
Nevertheless, people who live in areas where malaria is endemic interestingly do not acquire an effective protection against reinfection. Furthermore, the existing vaccines showed insufficient protection and long-term efficiency [90].
To conclude, TRM cells are important during parasite infection and could be helpful in long-term protection in vaccine development (Figure 4).
4.3. Liver-Resident T Cells in Chronic Inflammatory Diseases
Chronic liver inflammation can subsequently lead to organ fibrosis, liver failure and HCC development. One of the main causes of liver transplantation in Western countries is non-alcoholic fatty liver disease (NAFLD)-induced end-stage liver disease. NAFLD is considered as a hepatic manifestation of the metabolic syndrome, hypertension and type-II diabetes. In addition, increasing age and disease-associated genetic variants could be shown to be risk factors [91]. The pathomechanism of the disease involves a complex immune response and is not fully understood yet. The involvement of a disrupted T cell—and a more pronounced pro-inflammatory immune response—could be shown in several studies [92,93]. Studies showed that liver-resident T cells, mainly γδ T cells, are involved in NAFLD disease progression. By the production of pro-inflammatory cytokines, such as IL-17A, IFN-γ and TNF-α, they contribute to the pathogenic immune response in NAFLD [94]. One subpopulation of γδ T cells in NAFLD patients expressed CD69, CXCR3 and CXCR6, which are important TRM cell markers. Additionally, they showed a liver-restricted TCR repertoire that supports their liver residency and origin [86]. The direct involvement of TRM cells with an αβ TCR could not be proven yet but it could be possible that they contribute to the overall pro-inflammatory immune response as they are involved in fibrosis in many other organs, e.g., the lung and kidney. Furthermore, it already was shown that the circulating memory T cell quantity is increased in NAFLD patients [95].
There are some indications that the systemic inflammation in obese patients is associated with an increase of TRM cells in the liver, which consequently induces chronic liver inflammation and may be associated with NAFLD disease progression. It was nicely shown in a study from Conroy et al. that there is an increase in liver TRM cells in patients with esophageal adenocarcinoma (OAC). OAC is connected with obesity and the patient cohort had an average BMI of 25.8, but more importantly, an increase in visceral fat. Interestingly, it could be shown that activated TRM cells were increased in the visceral fat and liver tissue of these patients, indicating a systemic immune activation. The activated TRM cells in the liver and adipose tissue produced pro-inflammatory cytokines, such as IL-1β, IL-12, GM-CSF, IL-6, IL-2, IL-4 and IL-15 [96]. These findings indicate that TRM cells could contribute to the overall pro-inflammatory phenotype in obese patients, which is one of the reasons for chronic inflammatory diseases like NAFLD and cancer development (Figure 4).
Extensive transcription factor and gene set enrichment analysis of lung TRM cells showed that, under inflammatory conditions, several drivers of the T cell effector function were overexpressed in these cells, such as RUNX3, IRF4 and NF-kB [97]. These inflammatory markers are known to be upregulated in chronic liver diseases such as ASH and NASH. There, the upregulation of the NF-kB pathway is well described, playing a major role in disease progression [98]. The exact involvement of NF-kB in TRM differentiation and function is not well studied yet, but it is possible that NF-kB signaling is involved as a crucial integrator within the pro-inflammatory cytokine milieu in the liver.
Recent studies could identify the crucial role of changes in the gut microbiota composition and an impairment of the intestinal barrier function on the regulation of body weight and the body’s fat composition. The translocation of bacterial products from the gut into the liver was further shown to contribute to disease progression in NASH or other chronic inflammatory liver diseases [99]. Inflammasomes are important mediators of the innate immune response that get activated upon recognition of pathogen-associated molecular patterns (PAMPs) in the liver. Activation of the NLR family pyrin domain containing 3 (NLRP3) inflammasome and the subsequent release of IL-1ß and IL-18 is implicated in the pathogenesis of chronic inflammatory liver diseases such as ASH and NAFLD [100]. Studies using NLRP3 knockout mice could show that the ablation of inflammasome activation not only prevents from obesity-induced inflammation in fat deposits and the liver but also directly influences the composition of intrahepatic and fat tissue T cells by increasing the amount of naïve T cell numbers and reducing the numbers of effector and memory T cells [101]. Thus, a direct effect of inflammasome activation on TRM cells is plausible but not shown yet.
4.4. Liver-Resident T Cells in Cancer
The liver is the primary site for HCC development and is often affected by metastasis of other cancers. HCC can be induced by several factors, e.g., chronic viral infection, alcohol consumption or NAFLD. HCC can interestingly develop in fibrotic and non-fibrotic tissue. In general, effector T cells are important to moderate anti-tumor immunology in HCC development. Furthermore, in several solid tumors and HCC, CD103+ TRM cells are enriched and associated with better prognosis and patient outcomes [84]. In contrast, a recently published study by Williams et al. showed that mice with an altered p21 expression have an increased risk of HCC development. The kinase inhibitor p21 promotes cell cycle arrest and has anti-proliferative functions. In this special situation, TRM cells were expanded in mice with HCC and decreased in line with HCC reduction [102]. Another study showed that TRM cells in HCC have an exhausted phenotype, e.g., shown by the expression of PD-1, LAG-3 and TIM3, especially in comparison to TRM cells, which are not in the tumor microenvironment. During HCC progression, the TRM cell frequency decreased, showing their importance in the anti-tumor immune response. Modern tumor therapies include immunotherapies that targets checkpoint inhibition to promote an anti-tumoral immune response. One important target is PD-1. Thus, during anti-tumor immunotherapy, TRMPD1high cells are the most responsive cells to the anti-PD-1 therapy to overcome tumor growth and progression [103]. Furthermore, Ma et al. could demonstrate that TRM cells in a tumor environment express other exhaustion and inhibitory markers, e.g., TIM3 and CTLA-4, but produce pro-inflammatory cytokines such as IFN-γ and TNF-α. Additionally, PD-1 expression in TRM cells within the tumor was correlated to poor disease outcome [104]. Overall, it could be demonstrated that TRM cells and TRM cell function is important for HCC development and anti-tumor therapy (Figure 2).
4.5. Liver-Resident T Cells in Transplantation
Solid organ transplantation can be the last therapeutic option in several end-stage organ diseases, such as acute liver failure and cirrhosis. It could be demonstrated that donor-derived TRM cells are detectable in the allografts and that their abundancy could be correlated with organ survival and reduced rejection. Although, organ rejection was associated with increased donor-derived TRM cell abundancy in areas of tissue damage [105,106,107]. A recently published study by Pallett et al. nicely demonstrated that long-lived TRM cells are detectable in liver allografts. Additionally, it could be shown that these cells can migrate into the draining lymph nodes, while downregulating some TRM-specific markers like CXCR6; however, they were not measurable intravascular. Furthermore, recipient-derived TRM cells were also abundant in the liver allografts. Interestingly, the same study could demonstrate that donor-derived TRM cells from an HBV-infected donor were still detectable in the liver allograft years after transplantation. Likewise, recipient-derived virus-specific T cells were detectable in the blood and liver. The latter had also a TRM-like phenotype. To conclude, this study could generate an important overview of the longevity, plasticity and phenotype of TRM cells in liver transplantation [10]. Further studies are needed to investigate the concrete role of liver TRM cells in organ rejection and function.
5. Conclusions
Liver inflammation can be caused by many different diseases but for little of them the importance of TRM cells remains to be investigated. Thus, it is possible that TRM cells play a role in liver autoimmune diseases as these are typically associated with an unbalanced T cell response. TRM cell involvement is already described in multiple sclerosis or type 1 diabetes and are best understood in autoimmune diseases of the skin. In these diseases, TRM cells were shown to be increased and correlated with disease severity. In addition, TRM cell abundancy was associated with pro-inflammatory cytokine expression [108]. A similar mechanism could be possible for autoimmune diseases in liver tissue.
Some studies indicate an involvement of TRM cell development in protective vaccination, but little is known about TRM cell development after vaccination in the liver. The importance of liver TRM cells is only described for parasite infections, as mentioned before. In other organs, TRM development upon vaccination could already be demonstrated. For instance, the commonly used influenza vaccine induces the development of long-lived TRM cell subpopulations independently of the use of neutralizing antibodies or the depletion of circulating memory T cells in mouse experiments. TRM cells were thereby sufficient to protect against reinfection [109].
The function and role of liver TRM cells is a relatively new research field with many unanswered questions. Although there are many interesting new studies, further investigations are urgently needed to analyze the role and importance of these cells in different diseases.
Funding
This research was funded by the German Research Foundation (DFG) (HKD: DR 1161/1-1, LMB: BA 7175/1-1).
Acknowledgments
We want to thank Georg M. Lauer for his continuous support, fruitful discussions and proofreading this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ariotti, S.; Haanen, J.B.; Schumacher, T.N. Behavior and function of tissue-resident memory T cells. Adv. Immunol. 2012, 114, 203–216. [Google Scholar] [PubMed]
- Sallusto, F.; Lenig, D.; Forster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Masopust, D.; Choo, D.; Vezys, V.; Wherry, E.J.; Duraiswamy, J.; Akondy, R.; Wang, J.; Casey, K.A.; Barber, D.L.; Kawamura, K.S.; et al. Dynamic T cell migration program provides resident memory within intestinal epithelium. J. Exp. Med. 2010, 207, 553–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, C. The Roles of Liver-Resident Lymphocytes in Liver Diseases. Front. Immunol. 2019, 10, 1582. [Google Scholar] [CrossRef]
- Mackay, L.K.; Braun, A.; Macleod, B.L.; Collins, N.; Tebartz, C.; Bedoui, S.; Carbone, F.R.; Gebhardt, T. Cutting edge: CD69 interference with sphingosine-1-phosphate receptor function regulates peripheral T cell retention. J. Immunol. 2015, 194, 2059–2063. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.I.; Brolin, R.E.; Ebert, E.C. Integrin alpha1beta1 (VLA-1) mediates adhesion of activated intraepithelial lymphocytes to collagen. Immunology 1999, 97, 679–685. [Google Scholar] [CrossRef]
- Baaten, B.J.; Tinoco, R.; Chen, A.T.; Bradley, L.M. Regulation of Antigen-Experienced T Cells: Lessons from the Quintessential Memory Marker CD44. Front. Immunol. 2012, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Iwasaki, A. Tissue-resident memory T cells. Immunol. Rev. 2013, 255, 165–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, L.K.; Wynne-Jones, E.; Freestone, D.; Pellicci, D.G.; Mielke, L.A.; Newman, D.M.; Braun, A.; Masson, F.; Kallies, A.; Belz, G.T.; et al. T-box Transcription Factors Combine with the Cytokines TGF-beta and IL-15 to Control Tissue-Resident Memory T Cell Fate. Immunity 2015, 43, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallett, L.J.; Burton, A.R.; Amin, O.E.; Rodriguez-Tajes, S.; Patel, A.A.; Zakeri, N.; Jeffery-Smith, A.; Swadling, L.; Schmidt, N.M.; Baiges, A.; et al. Longevity and replenishment of human liver-resident memory T cells and mononuclear phagocytes. J. Exp. Med. 2020, 217, e20200050. [Google Scholar] [CrossRef] [PubMed]
- Pallett, L.J.; Davies, J.; Colbeck, E.J.; Robertson, F.; Hansi, N.; Easom, N.J.W.; Burton, A.R.; Stegmann, K.A.; Schurich, A.; Swadling, L.; et al. IL-2(high) tissue-resident T cells in the human liver: Sentinels for hepatotropic infection. J. Exp. Med. 2017, 214, 1567–1580. [Google Scholar] [CrossRef] [PubMed]
- Tse, S.W.; Radtke, A.J.; Espinosa, D.A.; Cockburn, I.A.; Zavala, F. The chemokine receptor CXCR6 is required for the maintenance of liver memory CD8⁺ T cells specific for infectious pathogens. J. Infect. Dis. 2014, 210, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.J.; Goldrath, A.W. Transcriptional programming of tissue-resident memory CD8(+) T cells. Curr. Opin. Immunol. 2018, 51, 162–169. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, C.; Zhang, N. Tissue-Specific Control of Tissue-Resident Memory T Cells. Crit Rev. Immunol. 2018, 38, 79–103. [Google Scholar] [CrossRef] [PubMed]
- Mackay, L.K.; Minnich, M.; Kragten, N.A.; Liao, Y.; Nota, B.; Seillet, C.; Zaid, A.; Man, K.; Preston, S.; Freestone, D.; et al. Hobit and Blimp1 instruct a universal transcriptional program of tissue residency in lymphocytes. Science 2016, 352, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Bird, L. Hunker down with HOBIT and BLIMP1. Nat. Rev. Immunol. 2016, 16, 338–339. [Google Scholar] [CrossRef]
- Förster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef]
- Skon, C.N.; Lee, J.Y.; Anderson, K.G.; Masopust, D.; Hogquist, K.A.; Jameson, S.C. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+ T cells. Nat. Immunol. 2013, 14, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, J.; Kim, E.; Lee, Y. The Capicua/ETS Translocation Variant 5 Axis Regulates Liver-Resident Memory CD8(+) T-Cell Development and the Pathogenesis of Liver Injury. Hepatology 2019, 70, 358–371. [Google Scholar] [CrossRef]
- Behr, F.M.; Chuwonpad, A.; Stark, R.; van Gisbergen, K. Armed and Ready: Transcriptional Regulation of Tissue-Resident Memory CD8 T Cells. Front. Immunol. 2018, 9, 1770. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, R.; Bevan, M.J.; Reiner, S.L.; Fearon, D.T. The precursors of memory: Models and controversies. Nat. Rev. Immunol. 2009, 9, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Raphael, I.; Joern, R.R.; Forsthuber, T.G. Memory CD4(+) T Cells in Immunity and Autoimmune Diseases. Cells 2020, 9, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, L.K.; Rahimpour, A.; Ma, J.Z.; Collins, N.; Stock, A.T.; Hafon, M.L.; Vega-Ramos, J.; Lauzurica, P.; Mueller, S.N.; Stefanovic, T.; et al. The developmental pathway for CD103(+)CD8+ tissue-resident memory T cells of skin. Nat. Immunol. 2013, 14, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Gaide, O.; Emerson, R.O.; Jiang, X.; Gulati, N.; Nizza, S.; Desmarais, C.; Robins, H.; Krueger, J.G.; Clark, R.A.; Kupper, T.S. Common clonal origin of central and resident memory T cells following skin immunization. Nat. Med. 2015, 21, 647–653. [Google Scholar] [CrossRef]
- Holz, L.E.; Prier, J.E.; Freestone, D.; Steiner, T.M.; English, K.; Johnson, D.N.; Mollard, V.; Cozijnsen, A.; Davey, G.M.; Godfrey, D.I.; et al. CD8(+) T Cell Activation Leads to Constitutive Formation of Liver Tissue-Resident Memory T Cells that Seed a Large and Flexible Niche in the Liver. Cell Rep. 2018, 25, 68–79.e4. [Google Scholar] [CrossRef] [Green Version]
- Herndler-Brandstetter, D.; Ishigame, H.; Shinnakasu, R.; Plajer, V.; Stecher, C.; Zhao, J.; Lietzenmayer, M.; Kroehling, L.; Takumi, A.; Kometani, K.; et al. KLRG1(+) Effector CD8(+) T Cells Lose KLRG1, Differentiate into All Memory T Cell Lineages, and Convey Enhanced Protective Immunity. Immunity 2018, 48, 716–729.e8. [Google Scholar] [CrossRef] [Green Version]
- Bergsbaken, T.; Bevan, M.J.; Fink, P.J. Local Inflammatory Cues Regulate Differentiation and Persistence of CD8(+) Tissue-Resident Memory T Cells. Cell Rep. 2017, 19, 114–124. [Google Scholar] [CrossRef]
- Mueller, S.N.; Mackay, L.K. Tissue-resident memory T cells: Local specialists in immune defence. Nat. Rev. Immunol. 2016, 16, 79–89. [Google Scholar] [CrossRef]
- Thompson, E.A.; Darrah, P.A.; Foulds, K.E.; Hoffer, E.; Caffrey-Carr, A.; Norenstedt, S.; Perbeck, L.; Seder, R.A.; Kedl, R.M.; Lore, K. Monocytes Acquire the Ability to Prime Tissue-Resident T Cells via IL-10-Mediated TGF-beta Release. Cell Rep. 2019, 28, 1127–1135.e4. [Google Scholar] [CrossRef] [Green Version]
- Steinbach, K.; Vincenti, I.; Merkler, D. Resident-Memory T Cells in Tissue-Restricted Immune Responses: For Better or Worse? Front. Immunol. 2018, 9, 2827. [Google Scholar] [CrossRef] [Green Version]
- Swadling, L.; Pallett, L.J.; Diniz, M.O.; Baker, J.M.; Amin, O.E.; Stegmann, K.A.; Burton, A.R.; Schmidt, N.M.; Jeffery-Smith, A.; Zakeri, N.; et al. Human Liver Memory CD8(+) T Cells Use Autophagy for Tissue Residence. Cell Rep. 2020, 30, 687–698.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlay, D.K.; Rosenzweig, E.; Sinclair, L.V.; Feijoo-Carnero, C.; Hukelmann, J.L.; Rolf, J.; Panteleyev, A.A.; Okkenhaug, K.; Cantrell, D.A. PDK1 regulation of mTOR and hypoxia-inducible factor 1 integrate metabolism and migration of CD8+ T cells. J. Exp. Med. 2012, 209, 2441–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, L.V.; Finlay, D.; Feijoo, C.; Cornish, G.H.; Gray, A.; Ager, A.; Okkenhaug, K.; Hagenbeek, T.J.; Spits, H.; Cantrell, D.A. Phosphatidylinositol-3-OH kinase and nutrient-sensing mTOR pathways control T lymphocyte trafficking. Nat. Immunol. 2008, 9, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sowell, R.T.; Rogozinska, M.; Nelson, C.E.; Vezys, V.; Marzo, A.L. Cutting edge: Generation of effector cells that localize to mucosal tissues and form resident memory CD8 T cells is controlled by mTOR. J. Immunol. 2014, 193, 2067–2071. [Google Scholar] [CrossRef]
- Pan, Y.; Tian, T.; Park, C.O.; Lofftus, S.Y.; Mei, S.; Liu, X.; Luo, C.; O’Malley, J.T.; Gehad, A.; Teague, J.E.; et al. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism. Nature 2017, 543, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frizzell, H.; Fonseca, R.; Christo, S.N.; Evrard, M.; Cruz-Gomez, S.; Zanluqui, N.G.; von Scheidt, B.; Freestone, D.; Park, S.L.; McWilliam, H.E.G.; et al. Organ-specific isoform selection of fatty acid-binding proteins in tissue-resident lymphocytes. Sci. Immunol. 2020, 5, eaay9283. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Wesselink, T.H.; Behr, F.M.; Kragten, N.A.M.; Arens, R.; Koch-Nolte, F.; van Gisbergen, K.; van Lier, R.A.W. TRM maintenance is regulated by tissue damage via P2RX7. Sci. Immunol. 2018, 3, eaau1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges da Silva, H.; Beura, L.K.; Wang, H.; Hanse, E.A.; Gore, R.; Scott, M.C.; Walsh, D.A.; Block, K.E.; Fonseca, R.; Yan, Y.; et al. The purinergic receptor P2RX7 directs metabolic fitness of long-lived memory CD8(+) T cells. Nature 2018, 559, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Doedens, A.L.; Phan, A.T.; Stradner, M.H.; Fujimoto, J.K.; Nguyen, J.V.; Yang, E.; Johnson, R.S.; Goldrath, A.W. Hypoxia-inducible factors enhance the effector responses of CD8(+) T cells to persistent antigen. Nat. Immunol. 2013, 14, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Han, J.W.; Choi, Y.J.; Rha, M.S.; Koh, J.Y.; Kim, K.H.; Kim, C.G.; Lee, Y.J.; Kim, A.R.; Park, J.; et al. Functions of human liver CD69(+)CD103(−)CD8(+) T cells depend on HIF-2alpha activity in healthy and pathologic livers. J. Hepatol. 2020, 72, 1170–1181. [Google Scholar] [CrossRef]
- Beura, L.K.; Hamilton, S.E.; Bi, K.; Schenkel, J.M.; Odumade, O.A.; Casey, K.A.; Thompson, E.A.; Fraser, K.A.; Rosato, P.C.; Filali-Mouhim, A.; et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Reese, T.A.; Bi, K.; Kambal, A.; Filali-Mouhim, A.; Beura, L.K.; Burger, M.C.; Pulendran, B.; Sekaly, R.P.; Jameson, S.C.; Masopust, D.; et al. Sequential Infection with Common Pathogens Promotes Human-like Immune Gene Expression and Altered Vaccine Response. Cell Host Microbe 2016, 19, 713–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klonowski, K.D.; Williams, K.J.; Marzo, A.L.; Blair, D.A.; Lingenheld, E.G.; Lefrancois, L. Dynamics of blood-borne CD8 memory T cell migration in vivo. Immunity 2004, 20, 551–562. [Google Scholar] [CrossRef] [Green Version]
- Steinert, E.M.; Schenkel, J.M.; Fraser, K.A.; Beura, L.K.; Manlove, L.S.; Igyarto, B.Z.; Southern, P.J.; Masopust, D. Quantifying Memory CD8 T Cells Reveals Regionalization of Immunosurveillance. Cell 2015, 161, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Takamura, S.; Yagi, H.; Hakata, Y.; Motozono, C.; McMaster, S.R.; Masumoto, T.; Fujisawa, M.; Chikaishi, T.; Komeda, J.; Itoh, J.; et al. Specific niches for lung-resident memory CD8+ T cells at the site of tissue regeneration enable CD69-independent maintenance. J. Exp. Med. 2016, 213, 3057–3073. [Google Scholar] [CrossRef]
- Anderson, K.G.; Mayer-Barber, K.; Sung, H.; Beura, L.; James, B.R.; Taylor, J.J.; Qunaj, L.; Griffith, T.S.; Vezys, V.; Barber, D.L.; et al. Intravascular staining for discrimination of vascular and tissue leukocytes. Nat. Protoc. 2014, 9, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.G.; Sung, H.; Skon, C.N.; Lefrancois, L.; Deisinger, A.; Vezys, V.; Masopust, D. Cutting edge: Intravascular staining redefines lung CD8 T cell responses. J. Immunol. 2012, 189, 2702–2706. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.N.; Gebhardt, T.; Carbone, F.R.; Heath, W.R. Memory T cell subsets, migration patterns, and tissue residence. Annu. Rev. Immunol. 2013, 31, 137–161. [Google Scholar] [CrossRef]
- Geissmann, F.; Cameron, T.O.; Sidobre, S.; Manlongat, N.; Kronenberg, M.; Briskin, M.J.; Dustin, M.L.; Littman, D.R. Intravascular immune surveillance by CXCR6+ NKT cells patrolling liver sinusoids. PLoS Biol. 2005, 3, e113. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, M.; Brinkmann, V.; Zerwes, H.G. FTY720 preferentially depletes naive T cells from peripheral and lymphoid organs. Int. Immunopharmacol. 2006, 6, 1902–1910. [Google Scholar] [CrossRef]
- Clark, R.A.; Watanabe, R.; Teague, J.E.; Schlapbach, C.; Tawa, M.C.; Adams, N.; Dorosario, A.A.; Chaney, K.S.; Cutler, C.S.; Leboeuf, N.R.; et al. Skin effector memory T cells do not recirculate and provide immune protection in alemtuzumab-treated CTCL patients. Sci. Transl. Med. 2012, 4, 117ra7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thome, J.J.; Yudanin, N.; Ohmura, Y.; Kubota, M.; Grinshpun, B.; Sathaliyawala, T.; Kato, T.; Lerner, H.; Shen, Y.; Farber, D.L. Spatial map of human T cell compartmentalization and maintenance over decades of life. Cell 2014, 159, 814–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senda, T.; Dogra, P.; Granot, T.; Furuhashi, K.; Snyder, M.E.; Carpenter, D.J.; Szabo, P.A.; Thapa, P.; Miron, M.; Farber, D.L. Microanatomical dissection of human intestinal T-cell immunity reveals site-specific changes in gut-associated lymphoid tissues over life. Mucosal Immunol. 2019, 12, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Ruiz, D.; Ng, W.Y.; Holz, L.E.; Ma, J.Z.; Zaid, A.; Wong, Y.C.; Lau, L.S.; Mollard, V.; Cozijnsen, A.; Collins, N.; et al. Liver-Resident Memory CD8(+) T Cells Form a Front-Line Defense against Malaria Liver-Stage Infection. Immunity 2016, 45, 889–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuff, A.O.; Robertson, F.P.; Stegmann, K.A.; Pallett, L.J.; Maini, M.K.; Davidson, B.R.; Male, V. Eomeshi NK Cells in Human Liver Are Long-Lived and Do Not Recirculate but Can Be Replenished from the Circulation. J. Immunol. 2016, 197, 4283–4291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, M.E.; Farber, D.L. Human lung tissue resident memory T cells in health and disease. Curr. Opin. Immunol. 2019, 59, 101–108. [Google Scholar] [CrossRef]
- Bartolome-Casado, R.; Landsverk, O.J.B.; Chauhan, S.K.; Richter, L.; Phung, D.; Greiff, V.; Risnes, L.F.; Yao, Y.; Neumann, R.S.; Yaqub, S.; et al. Resident memory CD8 T cells persist for years in human small intestine. J. Exp. Med. 2019, 216, 2412–2426. [Google Scholar] [CrossRef]
- Gebhardt, T.; Whitney, P.G.; Zaid, A.; Mackay, L.K.; Brooks, A.G.; Heath, W.R.; Carbone, F.R.; Mueller, S.N. Different patterns of peripheral migration by memory CD4+ and CD8+ T cells. Nature 2011, 477, 216–219. [Google Scholar] [CrossRef]
- Fernandez-Ruiz, D.; Ng, W.Y.; Holz, L.E.; Ma, J.Z.; Zaid, A.; Wong, Y.C.; Lau, L.S.; Mollard, V.; Cozijnsen, A.; Collins, N.; et al. Liver-Resident Memory CD8(+) T Cells Form a Front-Line Defense against Malaria Liver-Stage Infection. Immunity 2019, 51, 780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.V.; Ma, W.; Miron, M.; Granot, T.; Guyer, R.S.; Carpenter, D.J.; Senda, T.; Sun, X.; Ho, S.H.; Lerner, H.; et al. Human Tissue-Resident Memory T Cells Are Defined by Core Transcriptional and Functional Signatures in Lymphoid and Mucosal Sites. Cell Rep. 2017, 20, 2921–2934. [Google Scholar] [CrossRef] [Green Version]
- Guidotti, L.G.; Inverso, D.; Sironi, L.; Di Lucia, P.; Fioravanti, J.; Ganzer, L.; Fiocchi, A.; Vacca, M.; Aiolfi, R.; Sammicheli, S.; et al. Immunosurveillance of the liver by intravascular effector CD8(+) T cells. Cell 2015, 161, 486–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolino, P.; Schrage, A.; Bowen, D.G.; Klugewitz, K.; Ghani, S.; Eulenburg, K.; Holz, L.; Hogg, N.; McCaughan, G.W.; Hamann, A. Early intrahepatic antigen-specific retention of naive CD8+ T cells is predominantly ICAM-1/LFA-1 dependent in mice. Hepatology 2005, 42, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- John, B.; Crispe, I.N. Passive and active mechanisms trap activated CD8+ T cells in the liver. J. Immunol. 2004, 172, 5222–5229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, H.A.; Cai, Y.; Wagle, M.V.; Sontani, Y.; Roots, C.M.; Miosge, L.A.; O’Connor, J.H.; Sutton, H.J.; Ganusov, V.V.; Heath, W.R.; et al. Up-regulation of LFA-1 allows liver-resident memory T cells to patrol and remain in the hepatic sinusoids. Sci. Immunol. 2017, 2, eaaj1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebhardt, T.; Wakim, L.M.; Eidsmo, L.; Reading, P.C.; Heath, W.R.; Carbone, F.R. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat. Immunol. 2009, 10, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Tse, S.W.; Cockburn, I.A.; Zhang, H.; Scott, A.L.; Zavala, F. Unique transcriptional profile of liver-resident memory CD8+ T cells induced by immunization with malaria sporozoites. Genes Immun 2013, 14, 302–309. [Google Scholar] [CrossRef]
- Chou, C.; Li, M.O. Tissue-Resident Lymphocytes across Innate and Adaptive Lineages. Front. Immunol. 2018, 9, 2104. [Google Scholar] [CrossRef] [Green Version]
- Reichel, C.A.; Puhr-Westerheide, D.; Zuchtriegel, G.; Uhl, B.; Berberich, N.; Zahler, S.; Wymann, M.P.; Luckow, B.; Krombach, F. C-C motif chemokine CCL3 and canonical neutrophil attractants promote neutrophil extravasation through common and distinct mechanisms. Blood 2012, 120, 880–890. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 2011, 11, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Kadoki, M.; Patil, A.; Thaiss, C.C.; Brooks, D.J.; Pandey, S.; Deep, D.; Alvarez, D.; von Andrian, U.H.; Wagers, A.J.; Nakai, K.; et al. Organism-Level Analysis of Vaccination Reveals Networks of Protection across Tissues. Cell 2017, 171, 398–413.e21. [Google Scholar] [CrossRef] [Green Version]
- Schenkel, J.M.; Fraser, K.A.; Masopust, D. Cutting edge: Resident memory CD8 T cells occupy frontline niches in secondary lymphoid organs. J. Immunol. 2014, 192, 2961–2964. [Google Scholar] [CrossRef] [PubMed]
- Feau, S.; Arens, R.; Togher, S.; Schoenberger, S.P. Autocrine IL-2 is required for secondary population expansion of CD8(+) memory T cells. Nat. Immunol. 2011, 12, 908–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crispe, I.N. Immune tolerance in liver disease. Hepatology 2014, 60, 2109–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Hao, X.; Chen, Y.; Bai, L.; Gao, X.; Lian, Z.; Wei, H.; Sun, R.; Tian, Z. The microbiota maintain homeostasis of liver-resident gammadeltaT-17 cells in a lipid antigen/CD1d-dependent manner. Nat. Commun. 2017, 7, 13839. [Google Scholar] [CrossRef] [PubMed]
- Goodall, K.J.; Nguyen, A.; Matsumoto, A.; McMullen, J.R.; Eckle, S.B.; Bertolino, P.; Sullivan, L.C.; Andrews, D.M. Multiple receptors converge on H2-Q10 to regulate NK and gammadeltaT-cell development. Immunol. Cell Biol. 2019, 97, 326–339. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Radaeva, S.; Park, O. Liver natural killer and natural killer T cells: Immunobiology and emerging roles in liver diseases. J. Leukoc. Biol. 2009, 86, 513–528. [Google Scholar] [CrossRef]
- Vermijlen, D.; Luo, D.; Froelich, C.J.; Medema, J.P.; Kummer, J.A.; Willems, E.; Braet, F.; Wisse, E. Hepatic natural killer cells exclusively kill splenic/blood natural killer-resistant tumor cells by the perforin/granzyme pathway. J. Leukoc. Biol. 2002, 72, 668–676. [Google Scholar]
- Borges da Silva, H.; Wang, H.; Qian, L.J.; Hogquist, K.A.; Jameson, S.C. ARTC2.2/P2RX7 Signaling during Cell Isolation Distorts Function and Quantification of Tissue-Resident CD8(+) T Cell and Invariant NKT Subsets. J. Immunol. 2019, 202, 2153–2163. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, R.; Kanda, T.; Fujisawa, M.; Matsumoto, N.; Masuzaki, R.; Ogawa, M.; Matsuoka, S.; Kuroda, K.; Moriyama, M. Different Mechanisms of Action of Regorafenib and Lenvatinib on Toll-Like Receptor-Signaling Pathways in Human Hepatoma Cell Lines. Int. J. Mol. Sci. 2020, 21, 3349. [Google Scholar] [CrossRef]
- Marinaik, C.B.; Kingstad-Bakke, B.; Lee, W.; Hatta, M.; Sonsalla, M.; Larsen, A.; Neldner, B.; Gasper, D.J.; Kedl, R.M.; Kawaoka, Y.; et al. Programming Multifaceted Pulmonary T Cell Immunity by Combination Adjuvants. Cell Rep. Med. 2020, 1, 100095. [Google Scholar] [CrossRef]
- Shoukry, N.H.; Grakoui, A.; Houghton, M.; Chien, D.Y.; Ghrayeb, J.; Reimann, K.A.; Walker, C.M. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J. Exp. Med. 2003, 197, 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Stelma, F.; de Niet, A.; Sinnige, M.J.; van Dort, K.A.; van Gisbergen, K.; Verheij, J.; van Leeuwen, E.M.M.; Kootstra, N.A.; Reesink, H.W. Human intrahepatic CD69 + CD8+ T cells have a tissue resident memory T cell phenotype with reduced cytolytic capacity. Sci. Rep. 2017, 7, 6172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolte, F.J.; Rehermann, B. Tissue-resident T cells in hepatitis B: A new target for cure? J. Exp. Med. 2017, 214, 1564–1566. [Google Scholar] [CrossRef]
- Lim, C.J.; Lee, Y.H.; Pan, L.; Lai, L.; Chua, C.; Wasser, M.; Lim, T.K.H.; Yeong, J.; Toh, H.C.; Lee, S.Y.; et al. Multidimensional analyses reveal distinct immune microenvironment in hepatitis B virus-related hepatocellular carcinoma. Gut 2019, 68, 916–927. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Peng, H.; Li, K.; Qu, K.; Wang, B.; Wu, Y.; Ye, L.; Dong, Z.; Wei, H.; Sun, R.; et al. Liver-Resident NK Cells Control Antiviral Activity of Hepatic T Cells via the PD-1-PD-L1 Axis. Immunity 2019, 50, 403–417.e4. [Google Scholar] [CrossRef] [Green Version]
- Hunter, S.; Willcox, C.R.; Davey, M.S.; Kasatskaya, S.A.; Jeffery, H.C.; Chudakov, D.M.; Oo, Y.H.; Willcox, B.E. Human liver infiltrating gammadelta T cells are composed of clonally expanded circulating and tissue-resident populations. J. Hepatol 2018, 69, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.; Claro, M.; Alexandre-Pires, G.; Santos-Mateus, D.; Martins, C.; Valerio-Bolas, A.; Rafael-Fernandes, M.; Pereira, M.A.; Pereira da Fonseca, I.; Tomas, A.M.; et al. Leishmania infantum antigens modulate memory cell subsets of liver resident T lymphocyte. Immunobiology 2017, 222, 409–422. [Google Scholar] [CrossRef]
- Cuburu, N.; Kim, R.; Guittard, G.C.; Thompson, C.D.; Day, P.M.; Hamm, D.E.; Pang, Y.S.; Graham, B.S.; Lowy, D.R.; Schiller, J.T. A Prime-Pull-Amplify Vaccination Strategy to Maximize Induction of Circulating and Genital-Resident Intraepithelial CD8(+) Memory T Cells. J. Immunol. 2019, 202, 1250–1264. [Google Scholar] [CrossRef] [Green Version]
- Holz, L.E.; Chua, Y.C.; de Menezes, M.N.; Anderson, R.J.; Draper, S.L.; Compton, B.J.; Chan, S.T.S.; Mathew, J.; Li, J.; Kedzierski, L.; et al. Glycolipid-peptide vaccination induces liver-resident memory CD8(+) T cells that protect against rodent malaria. Sci. Immunol. 2020, 5, eaaz8035. [Google Scholar] [CrossRef]
- Tran, T.M.; Li, S.; Doumbo, S.; Doumtabe, D.; Huang, C.Y.; Dia, S.; Bathily, A.; Sangala, J.; Kone, Y.; Traore, A.; et al. An intensive longitudinal cohort study of Malian children and adults reveals no evidence of acquired immunity to Plasmodium falciparum infection. Clin. Infect. Dis. 2013, 57, 40–47. [Google Scholar] [CrossRef]
- Drescher, H.K.; Weiskirchen, S.; Weiskirchen, R. Current Status in Testing for Nonalcoholic Fatty Liver Disease (NAFLD) and Nonalcoholic Steatohepatitis (NASH). Cells 2019, 8, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diedrich, T.; Kummer, S.; Galante, A.; Drolz, A.; Schlicker, V.; Lohse, A.W.; Kluwe, J.; Eberhard, J.M.; Schulze Zur Wiesch, J. Characterization of the immune cell landscape of patients with NAFLD. PLoS ONE 2020, 15, e0230307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Herck, M.A.; Weyler, J.; Kwanten, W.J.; Dirinck, E.L.; De Winter, B.Y.; Francque, S.M.; Vonghia, L. The Differential Roles of T Cells in Non-alcoholic Fatty Liver Disease and Obesity. Front. Immunol. 2019, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harley, I.T.; Stankiewicz, T.E.; Giles, D.A.; Softic, S.; Flick, L.M.; Cappelletti, M.; Sheridan, R.; Xanthakos, S.A.; Steinbrecher, K.A.; Sartor, R.B.; et al. IL-17 signaling accelerates the progression of nonalcoholic fatty liver disease in mice. Hepatology 2014, 59, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Inzaugarat, M.E.; Ferreyra Solari, N.E.; Billordo, L.A.; Abecasis, R.; Gadano, A.C.; Chernavsky, A.C. Altered phenotype and functionality of circulating immune cells characterize adult patients with nonalcoholic steatohepatitis. J. Clin. Immunol. 2011, 31, 1120–1130. [Google Scholar] [CrossRef]
- Conroy, M.J.; Galvin, K.C.; Doyle, S.L.; Kavanagh, M.E.; Mongan, A.M.; Cannon, A.; Moore, G.Y.; Reynolds, J.V.; Lysaght, J. Parallel Profiles of Inflammatory and Effector Memory T Cells in Visceral Fat and Liver of Obesity-Associated Cancer Patients. Inflammation 2016, 39, 1729–1736. [Google Scholar] [CrossRef]
- Hombrink, P.; Helbig, C.; Backer, R.A.; Piet, B.; Oja, A.E.; Stark, R.; Brasser, G.; Jongejan, A.; Jonkers, R.E.; Nota, B.; et al. Programs for the persistence, vigilance and control of human CD8(+) lung-resident memory T cells. Nat. Immunol. 2016, 17, 1467–1478. [Google Scholar] [CrossRef]
- Farrell, G.C.; van Rooyen, D.; Gan, L.; Chitturi, S. NASH is an Inflammatory Disorder: Pathogenic, Prognostic and Therapeutic Implications. Gut Liver 2012, 6, 149–171. [Google Scholar] [CrossRef] [Green Version]
- Baothman, O.A.; Zamzami, M.A.; Taher, I.; Abubaker, J.; Abu-Farha, M. The role of Gut Microbiota in the development of obesity and Diabetes. Lipids Health Dis. 2016, 15, 108. [Google Scholar] [CrossRef] [Green Version]
- Wree, A.; Broderick, L.; Canbay, A.; Hoffman, H.M.; Feldstein, A.E. From NAFLD to NASH to cirrhosis-new insights into disease mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 627–636. [Google Scholar] [CrossRef]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.; Liu, X.; Zhang, Y.; Reske, J.; Bahal, D.; Gohl, T.G.; Hollern, D.; Ensink, E.; Kiupel, M.; Luo, R.; et al. NCOA5 deficiency promotes a unique liver protumorigenic microenvironment through p21(WAF1/CIP1) overexpression, which is reversed by metformin. Oncogene 2020, 39, 3821–3836. [Google Scholar] [CrossRef] [PubMed]
- Chew, V.; Lai, L.; Pan, L.; Lim, C.J.; Li, J.; Ong, R.; Chua, C.; Leong, J.Y.; Lim, K.H.; Toh, H.C.; et al. Delineation of an immunosuppressive gradient in hepatocellular carcinoma using high-dimensional proteomic and transcriptomic analyses. Proc. Natl. Acad. Sci. USA 2017, 114, E5900–E5909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Zheng, B.; Goswami, S.; Meng, L.; Zhang, D.; Cao, C.; Li, T.; Zhu, F.; Ma, L.; Zhang, Z.; et al. PD1(Hi) CD8(+) T cells correlate with exhausted signature and poor clinical outcome in hepatocellular carcinoma. J. Immunother. Cancer 2019, 7, 331. [Google Scholar] [CrossRef]
- Zuber, J.; Shonts, B.; Lau, S.P.; Obradovic, A.; Fu, J.; Yang, S.; Lambert, M.; Coley, S.; Weiner, J.; Thome, J.; et al. Bidirectional intragraft alloreactivity drives the repopulation of human intestinal allografts and correlates with clinical outcome. Sci. Immunol. 2016, 1, eaah3732. [Google Scholar] [CrossRef] [Green Version]
- Snyder, M.E.; Finlayson, M.O.; Connors, T.J.; Dogra, P.; Senda, T.; Bush, E.; Carpenter, D.; Marboe, C.; Benvenuto, L.; Shah, L.; et al. Generation and persistence of human tissue-resident memory T cells in lung transplantation. Sci. Immunol. 2019, 4, eaav5581. [Google Scholar] [CrossRef]
- Lian, C.G.; Bueno, E.M.; Granter, S.R.; Laga, A.C.; Saavedra, A.P.; Lin, W.M.; Susa, J.S.; Zhan, Q.; Chandraker, A.K.; Tullius, S.G.; et al. Biomarker evaluation of face transplant rejection: Association of donor T cells with target cell injury. Mod. Pathol. 2014, 27, 788–799. [Google Scholar] [CrossRef] [Green Version]
- Park, C.O.; Kupper, T.S. The emerging role of resident memory T cells in protective immunity and inflammatory disease. Nat. Med. 2015, 21, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Zens, K.D.; Chen, J.K.; Farber, D.L. Vaccine-generated lung tissue-resident memory T cells provide heterosubtypic protection to influenza infection. JCI Insight 2016, 1. [Google Scholar] [CrossRef]
Figure 1.
To convey hepatic infiltration, memory T cells express high levels of liver-specific homing markers such as CD103, LFA-1, CXCR6 or CXCR3. Within the liver, TRM cells are exposed to a variety of environmental conditions that directly influence their maintenance, function and development; for example, nutrient deprivation and fatty acid oxidation, hypoxia and different inflammatory conditions. Besides the expression of IL-15, IL-7 and TGF-β, which directly effects hepatic stellate cells (HSCs), intrahepatic TRM cells are also able to secret pro-inflammatory molecules such as Interferon, Granzyme B, IL-17 and TNF-α. Liver-resident TRM cells have the ability to egress back into the bloodstream by upregulating CCR7 and S1PR1.
Figure 1.
To convey hepatic infiltration, memory T cells express high levels of liver-specific homing markers such as CD103, LFA-1, CXCR6 or CXCR3. Within the liver, TRM cells are exposed to a variety of environmental conditions that directly influence their maintenance, function and development; for example, nutrient deprivation and fatty acid oxidation, hypoxia and different inflammatory conditions. Besides the expression of IL-15, IL-7 and TGF-β, which directly effects hepatic stellate cells (HSCs), intrahepatic TRM cells are also able to secret pro-inflammatory molecules such as Interferon, Granzyme B, IL-17 and TNF-α. Liver-resident TRM cells have the ability to egress back into the bloodstream by upregulating CCR7 and S1PR1.

Figure 2.
Upon stimulation, TRM cells can recirculate into the blood by inducing the expression of higher levels of CD69, CCR7 and S1PR1. In the blood they can have systemic effector functions and convey hepatic immune cell attraction. For the infiltration into the liver, memory T cells express high levels of the hepatic homing markers LFA-1, CD103, CXCR6, CXCR3 and CD69. In the liver they downregulate the expression of these homing receptors and differentiate into liver-resident TRM cells.
Figure 2.
Upon stimulation, TRM cells can recirculate into the blood by inducing the expression of higher levels of CD69, CCR7 and S1PR1. In the blood they can have systemic effector functions and convey hepatic immune cell attraction. For the infiltration into the liver, memory T cells express high levels of the hepatic homing markers LFA-1, CD103, CXCR6, CXCR3 and CD69. In the liver they downregulate the expression of these homing receptors and differentiate into liver-resident TRM cells.

Figure 3.
TRM cells can be detected in mice and humans. In both species, TRM cells express CD69 and CXCR3. However, in humans, one population of TRM cells expresses CD103+, while CD103 surface expression is not found in mice TRM cells. On the other hand, mice TRM cells express CD44, which mediates their liver retention and ICAM-1/LFA-1 responsible for TRM cell migration within the liver. The frequency of TRM cells within the resident T cell compartment varies between mice and humans. Some studies have described that mice TRM cells express the transcription factors Tbet, Tbx21 and KLF2, but the transcription factors upregulated by human TRM cells are not defined yet.
Figure 3.
TRM cells can be detected in mice and humans. In both species, TRM cells express CD69 and CXCR3. However, in humans, one population of TRM cells expresses CD103+, while CD103 surface expression is not found in mice TRM cells. On the other hand, mice TRM cells express CD44, which mediates their liver retention and ICAM-1/LFA-1 responsible for TRM cell migration within the liver. The frequency of TRM cells within the resident T cell compartment varies between mice and humans. Some studies have described that mice TRM cells express the transcription factors Tbet, Tbx21 and KLF2, but the transcription factors upregulated by human TRM cells are not defined yet.

Figure 4.
TRM cells play a central role during intrahepatic inflammation. In parasitic infections, TRM cells mediate cytotoxicity by the production of pro-inflammatory cytokines such as IFN-γ and TNF-α. During hepatitis virus infection, virus-specific TRM cells could be negatively correlated with virus titers by secreting high levels of IL-2 and perforin. Patients who attained viral control showed enriched intrahepatic TRM cell numbers. In sterile chronic inflammation of the liver, such as alcoholic or nonalcoholic steatohepatitis (ASH and NASH) TRM cells were associated with the overall pro-inflammatory cytokine milieu in the liver. Their expression of IL-17, IFN-γ and TNF-α contributes to disease progression. During the subsequent development of hepatocellular carcinoma (HCC), the CD103+ TRM cells showed opposing effects. Patients with enriched numbers of these cells showed a better prognosis by mediating the anti-tumoral immune response.
Figure 4.
TRM cells play a central role during intrahepatic inflammation. In parasitic infections, TRM cells mediate cytotoxicity by the production of pro-inflammatory cytokines such as IFN-γ and TNF-α. During hepatitis virus infection, virus-specific TRM cells could be negatively correlated with virus titers by secreting high levels of IL-2 and perforin. Patients who attained viral control showed enriched intrahepatic TRM cell numbers. In sterile chronic inflammation of the liver, such as alcoholic or nonalcoholic steatohepatitis (ASH and NASH) TRM cells were associated with the overall pro-inflammatory cytokine milieu in the liver. Their expression of IL-17, IFN-γ and TNF-α contributes to disease progression. During the subsequent development of hepatocellular carcinoma (HCC), the CD103+ TRM cells showed opposing effects. Patients with enriched numbers of these cells showed a better prognosis by mediating the anti-tumoral immune response.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bartsch, L.M.; Damasio, M.P.S.; Subudhi, S.; Drescher, H.K. Tissue-Resident Memory T Cells in the Liver—Unique Characteristics of Local Specialists. Cells 2020, 9, 2457. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9112457
AMA Style
Bartsch LM, Damasio MPS, Subudhi S, Drescher HK. Tissue-Resident Memory T Cells in the Liver—Unique Characteristics of Local Specialists. Cells. 2020; 9(11):2457. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9112457
Chicago/Turabian StyleBartsch, Lea M., Marcos P. S. Damasio, Sonu Subudhi, and Hannah K. Drescher. 2020. "Tissue-Resident Memory T Cells in the Liver—Unique Characteristics of Local Specialists" Cells 9, no. 11: 2457. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9112457
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.