The Intricate Role of p53 in Adipocyte Differentiation and Function


1
Department of Internal Medicine, Seoul National University Bundang Hospital, Seongnam 13620, Korea
2
Herbal Medicine Resources Research Center, Korea Institute of Oriental Medicine, Naju-si 58245, Korea
*
Author to whom correspondence should be addressed.
Cells 2020, 9(12), 2621; https://0-doi-org.brum.beds.ac.uk/10.3390/cells9122621
Submission received: 2 November 2020
/
Revised: 2 December 2020
/
Accepted: 3 December 2020
/
Published: 7 December 2020
(This article belongs to the Special Issue p53 Regulation Mechanisms)
Abstract
:For more than three decades, numerous studies have demonstrated the function of p53 in cell cycle, cellular senescence, autophagy, apoptosis, and metabolism. Among diverse functions, the essential role of p53 is to maintain cellular homeostatic response to stress by regulating proliferation and apoptosis. Recently, adipocytes have been studied with increasing intensity owing to the increased prevalence of metabolic diseases posing a serious public health concern and because metabolic dysfunction can directly induce tumorigenesis. The prevalence of metabolic diseases has steadily increased worldwide, and a growing interest in these diseases has led to the focus on the role of p53 in metabolism and adipocyte differentiation with or without metabolic stress. However, our collective understanding of the direct role of p53 in adipocyte differentiation and function remains insufficient. Therefore, this review focuses on the newly discovered roles of p53 in adipocyte differentiation and function.
{kind=link}
{kind=link}
1. Introduction
The multifunctional protein p53 (encoded by TP53 in humans and Trp53 in mice, hereafter termed p53) is a key regulator of cell proliferation and apoptotic response to exogenous or endogenous cellular stresses [1]. Under various cellular stresses, p53 protects normal cells against tumorigenesis in response to oncogene activation, dysregulation of the cell cycle, ribosomal stress, oxidative stress, DNA damage, and hypoxia [2]. p53 is involved in several intricate signaling pathways including those that regulate gene expression, protein stability, protein interaction, and post-transcriptional modifications [3]. In response to cellular stress, p53 contributes to various cellular physiological functions. Its concentration or phosphorylation status directly regulates essential physiological functions. It also regulates metabolic homeostasis, mitochondrial function, and antioxidant defense mechanisms [4,5,6,7]. Under mild or no stress, p53 controls cellular physiological activities by regulating the energy status of the cell and availability of nutrients, growth factors, and hormones [4,6,8,9]. It also regulates antioxidant gene functions [4,10,11]. Because of these multifunctional roles of p53, although p53 knockout mice are viable, a significant number of p53 knockout mice die during embryogenesis because of developmental problems including defects in embryo implantation, the neuronal system, and serious multiple congenital abnormalities [12,13]. However, the role of p53 in specific research areas such as aging is still unclear. For example, p53 levels were found to be increased in a pro- and anti-aging mouse model [14].
In the absence of cellular stresses, mouse double minute 2 (MDM2), an E3 ubiquitin ligase, regulates p53 levels via proteasomal degradation pathways [15]. When cells are exposed to stress, ataxia–telangiectasia-mutated kinase phosphorylates p53, which forms the p53-MDM2 complex [16]. When the p53 pathway is activated, the total level of p53 and its binding activity rapidly increases after phosphorylation. Activated p53 further activates several genes whose products induce cell cycle arrest, apoptosis, DNA repair, cellular senescence, and metabolic reprogramming [17,18]. p53 activity should be tightly regulated to allow normal cell growth and development because its activation prevents cell growth and proliferation. Interestingly, several studies have suggested that p53 regulates the balance between cellular development and differentiation. Schmid et al. demonstrated that p53 is highly expressed in midgestation mouse embryos and its level decreases after organogenesis [19]. However, the contribution of p53 in differentiation or development in a mouse model is still debatable [20].
Adipose tissue plays an important role in the regulation of systemic energy balance and glucose homeostasis. For example, adipose tissues regulate energy storage, lipid metabolism, glucose homeostasis, and energy expenditure [21]. The epidemic of metabolic disorders has presented a serious public health concern worldwide. Hence, adipocyte differentiation needs to be thoroughly studied. Three types of adipose tissues are found in mammals including humans: white adipose tissue (WAT), beige (or brite) adipose tissue, and brown adipose tissue (BAT) [22]. Although all adipose tissues are derived from mesenchymal progenitor cells, specific proteins or transcriptional factors regulate adipocyte lineage during adipogenesis in vivo and in vitro [23,24,25]. On embryonic day 14.5 and 15.5 post-coitus, BATs develop from early muscle progenitor cells that express myogenic transcription factors (Myf5) and BATs located around the central organs in mouse model [23]. BATs are rich in mitochondria and express uncoupling protein 1 (UCP1). They use substrates such as fatty acids and glucose to generate heat in response to various stimuli such as cold. Because of the function of non-shivering thermogenesis, BATs are considered a novel therapeutic target for treating metabolic diseases such as obesity.
On the other hand, the main function of WATs is energy storage in the triglyceride form. WATs are generally located subcutaneously throughout the body and constitute as much as 20–30% of the body weighty of normal humans. WAT progenitor cells are differently developed according to location; subcutaneous white adipose tissues develop before birth but visceral white adipose tissues develop after birth [26,27]. However, none of the WATs have the same characteristics. Specific WATs are incredibly dynamic and respond to stimuli including internal or external signaling and metabolic demands. Subcutaneous WATs are more prone to browning than visceral adipose tissues, and they can be activated upon exposure to cold temperatures or β-adrenergic stimulation because subcutaneous WATs are initiated from different lineage progenitor cells compared to visceral adipose tissues [28]. These browning adipocytes that appear in WATs are called “beige” adipocytes, which are induced by the browning stimulus leading to phenotypical similarity with the classical BAT such as abundance of respiratory mitochondria, multilocular small lipid droplets, and increased expression of Ucp1. WAT adipose tissues increase in cell number and size after weaning [29]. The mature white adipocyte is filled with a single lipid droplet containing a few mitochondria. Recent studies have demonstrated that p53 levels in WATs are increased in diet-induced or genetic obesity mouse models [30,31]. Hence, determining the role of p53 in the regulation of adipogenesis and cellular metabolism could reveal an important association between obesity and cancer.
In this review, we discuss the important role of p53 and highlight the key molecules that are regulated by p53 during adipogenesis. We also discuss the role of p53 in regulating metabolism in adipose tissues. Finally, we identify important outstanding questions and issues pertaining to the role of p53 in adipogenesis that remain to be addressed.
2. Role of P53 and White Adipocyte Differentiation and Lipid Metabolism
p53 is a regulator of cell survival and proliferation in response to multiple stress signals against abnormal cell growth and tumorigenesis. Apart from its role as a tumor suppressor, p53 has been involved in the regulation of differentiation of mesenchymal cells that can suppress white adipocyte differentiation [32,33,34]. White adipocytes store and accumulate lipids throughout their differentiation cycle. Adipocytes are derived from multipotent mesenchymal stem cells (MSCs), which are first committed to the adipogenic lineage and then transformed into preadipocytes. In secondary terminal differentiation, preadipocytes become mature adipocytes. The process of adipocyte differentiation involves three defined stages. The first step is the commitment of MSCs to the adipocyte lineage. The second step is mitotic clonal expansion (MCE) involving DNA replication and duplication of cells. During the early phase of differentiation, cell-cycle-arrested cells re-enter the cell cycle and undergo one or two rounds of the cell cycle, which is regarded as MCE. Given the association of cell cycle regulation with adipogenesis, the inhibitory effects of p53 in white adipocyte differentiation might occur via regulation of expression of several genes including p21 [35]. For example, MEFs lacking either p53 or p21 undergo spontaneous adipogenesis, and mice lacking p21 and p27 show adipose tissue hyperplasia [36,37]. The inhibitory effect of p53 on adipocyte differentiation relies on its transcriptional activity. Inhibition of adipocyte differentiation might result from forced expression of p53 during mitotic clonal expansion. Thus, the possible mechanism linking clonal expansion to adipogenesis is the capability of p53 to suppress adipogenesis and is dependent on its DNA binding ability, demonstrated by the failure to inhibit MEF adipogenesis by p53 with a mutation in the DNA-binding domain [38,39]. The third step is terminal differentiation, which involves transcriptional factors such as the CCAAT/enhancer-binding proteins (C/EBPs) family and peroxisome proliferator-activated receptor–γ (PPARγ), and a significant expression of lipogenic genes such as adiponectin and fatty acid-binding protein 4 (Fabp4, also called aP2) [40,41,42]. Huang et al. demonstrated that p53 partially suppressed preadipocyte differentiation and adipogenesis in the process of regulating adipogenic gene expression and Akt signaling [34]. Therefore, p53 protein is involved in white adipogenesis and plays a specific role in the regulation of adipogenesis. Although the role of p53 as a tumor suppressor has been clearly established, the transcriptional activity of p53 during adipogenic differentiation has not been adequately studied.
Adipogenesis is controlled by several transcriptional factors including C/EBPs and PPARγ. C/EBPβ and C/EBPδ are expressed at an early stage of adipocyte differentiation and they induce the expression of C/EBPα and PPARγ [40,41,42]. In murine preadipocyte cell lines such as 3T3-L1, nutlin-3a-mediated p53 protein accumulation, and ectopic p53 overexpression cause a decrease in the expression of Pparγ and its coactivator PPARγ coactivator1 alpha (Ppargc1a or Pgc-1α) [32,34,43]. Knockdown of p53 in differentiated 3T3-L1 cells increases Pgc-1α gene expression. Furthermore, a few studies have reported enhanced accumulation of lipid droplets with an increase in Pparγ and aP2 protein levels [32,43]. The mesenchymal stem cell line C3H10T1/2 can differentiate into white adipocytes upon induction of adipogenesis. Compared to 3T3-L1 cells, the C3H10T1/2 cell line has been used to investigate early events of adipogenesis [44,45]. Knockdown of p53 in C3H10T1/2 cells was enhanced by the upregulation of adipogenic genes such as Pparγ, C/ebpα, aP2, and adiponectin [32]. Furthermore, knockdown of p53 by specific shRNA enhanced the adipogenic capacity in both mouse and human cell lines, in which the levels of adipogenic gene markers such as PPARγ, aP2, and Adipoq increased [32,33]. In addition to cell lines, mouse embryonic fibroblasts (MEFs) isolated from p53 knockout mice were differentiated into adipocytes. The MEFs obtained from p53 knockout mice accumulated more lipid droplets and showed increased expression of adipogenic markers such as Pparγ and C/ebpα compared with wild-type MEFs [32,33]. In addition, nutlin is known to induce stabilization and activation of p53 [46]. Nutlin-3a-mediated p53 increment led the downregulation of Pparγ expression in wild-type MEFs [33]. In line with the inhibitory role of p53 in adipocyte differentiation, the expression of p53 was downregulated in primary differentiated cells derived from the stromal vascular fraction of inguinal WAT (iWAT) obtained from p53 knockout mice and human adipocyte-derived stem cells with knockdown of p53 [32,47]. Moreover, coactivator-associated arginine methyltransferase 1 (Carm1) has been shown to enhance adipogenesis by activating PPARγ in 3T3-L1 cells [43]. Thus, CARM1 has also been suggested as a possible modulator of the anti-adipogenic effects of p53. Furthermore, p53 activation following nutlin-3a treatment in 3T3-L1 cells reduced adipocyte development and downregulated Carm1 and reduced its protein expression (Figure 1). Importantly, overexpression of CARM1 prevented the inhibition of adipocyte differentiation after nutlin-3a treatment [48,49]. Another p53 target protein is twist family BHLH transcription factor 2 (Twist2). Twist2 is an essential protein that regulates adipocyte differentiation in marrow-resident MSCs. Loss of p53 induces the exhaustion of reactive oxygen species (ROS) in mitochondria and downregulates the expression of Twist2. These phenomena block the adipogenic differentiation and lead to osteogenic differentiation in MSCs. These results indicate that p53 is important not only for adipogenesis but also to maintain MSC integrity [50]. Lastly, the amount of Mdm2 is also important. Mdm2 is a major negative regulator of p53 and is highly expressed in 3T3-L1 preadipocytes [51]. However, Mdm2-p53 double knockout mouse embryonic fibroblast is not able to differentiate into mature adipocyte, and Mdm2 is known to regulate adipocyte differentiation in a p53-independent manner [52,53,54]. Therefore, as a negative regulator of p53 in 3T3-L1 adipogenesis, the role of Mdm2 needs to be carefully considered.
The canonical pathway of adipocyte lipolysis is catalyzed by three main lipases [53,54]. The first lipase is called patatin-like phospholipase domain containing-2 (PNPLA2)/adipocyte triglyceride lipase (ATGL), which is the rate-limiting enzyme for triacylglycerol (TAG) hydrolysis. The cholesterol esterase called “hormone-sensitive lipase” can utilize multiple substrates such as TAG, DAG, and retinyl. The third lipase is monoacylglycerol lipase (MGL). These three lipases function together to degrade TAGs sequentially, resulting in the release of three types of fatty acids (FAs) and glycerol. Additionally, adipocyte lipolysis involves multifunctional enzymes that catalyze reactions in both directions, resulting in the re-esterification and recycling of FAs [55,56]. The role of p53 in white adipocytes extends beyond the regulation of adipogenesis. p53-associated lipolysis could control the regulation of genes involved in lipid droplet accumulation [57]. In human adipocytes, overexpression of p53 reduces lipid droplet accumulation. Furthermore, inhibition of lipolysis by acipimox and induction of lipolysis by isoproterenol showed that p53 expression and lipolysis were closely regulated in mouse WATs [58]. Furthermore, Andreas et al. showed that DNA damaged induced transcript 4 (Ddit4), which is upregulated by fasting in white adipose tissue, is a p53 target gene. Ddit4 is induced by p53 activation and its ectopic expression augments lipolysis in differentiated C3H10T1/2 adipocytes [59]. Most recently, Wang et al. demonstrated that ATGL promoter has a conserved p53 binding site that promotes p53-induced ATGL promoter activity and transcription. Therefore, p53 induces lipolysis by directly activating ATGL transcription [60].
Recent studies have suggested that activation of p53 under abnormal metabolic conditions in adipose tissues aggravates obesity. Kung et al. demonstrated that mice harboring the proline-to-arginine 72 (P72R) variant of p53 who were administered a high-fat diet developed severe obesity and metabolic dysfunction than mice expressing a proline 72 variant [61]. In line with these results, p53 inhibition is known to prevent weight gain against a high-fat diet in mouse model [62]. Altogether, these results indicate that p53 regulates lipid metabolism in adipocytes. Taken together, these studies have suggested that p53 plays an important role in white adipocyte differentiation and function.
3. Role of P53 in Brown Adipocyte Differentiation and Thermogenesis
BATs are located around the central organs and are essential for whole-body energy homeostasis, particularly energy expenditure [63,64]. Hence, BATs have been suggested to be a novel therapeutic target for the treatment of diabetes and obesity [22]. Because BATs are important for cold-induced, nonshivering thermogenesis in newborn mouse pups the generation of BATs begins in the mid/late postnatal stage and undergoes rapid development [23,65]. Although BATs and WATs have different developmental origins and physiological functions, they share many common adipogenic developments including a conserved PPARγ and C/EBPα-driven transcriptional regulation of adipogenesis. Several recent studies have revealed that p53 plays a specific role in brown adipogenesis both in vitro and in vivo. We discuss these roles of p53 in brown adipogenesis in the subsequent sections.
A recent study performed by Molchadsky et al. demonstrated that p53 is a positive regulatory factor that maintains proper brown adipocyte maturation in vitro and prevents the development of obesity because of a high-fat diet in the mouse model [32]. Loss of p53 in the PR-domain contains 16 (PRDM16)- overexpressing myogenic C2 cells reduced the expression of general adipogenic genes and functional brown marker genes such as aP2, cell death-inducing DFFA-like effector a (Cidea), and ELOVL fatty acid elongase 3 (Elovl3). In addition, oil red O staining results support the assumption that p53 is required for the normal development of brown adipocytes [32]. Consistent with these results, the depletion of p53 downregulates PRDM16 expression. PRDM16 is a key transcriptional regulator in determining the brown adipocyte lineage and their subsequent development [66,67]. PRDM16 consists of a PR/SET domain at the N-terminal, two zine-finger domains containing seven or three C2H2 zinc finger domains, a proximal regulatory region, a repression domain, and an acidic activation domain in the C-terminal [68]. Because PRDM16 can bind directly to a specific DNA sequence via two sets of zinc finger domains or form a protein-protein complex with other adipogenic regulators, it regulates transcriptional activities in BAT development. PRDM16 knockout has been shown to induce a significant reduction in BAT development in a mouse model [69]. Moreover, PRDM16 regulates not only differentiation but also functional genetic expression in brown adipocytes. The expression of PRDM16 controls the transcription of essential brown adipocyte functional genes such as Elovl3, Cidea, Pgc-1α, and PPARα [70,71,72]. Cidea and Pgc-1α are important transcriptional regulators of brown adipocyte thermogenesis, which are involved in the regulation of Ucp1 transcriptional activity [73,74]. Overexpression of PRDM16 could upregulate the expression of genes whose products are involved in the mitochondrial oxidative pathway in mature brown adipocytes [69]. In vivo studies have shown that depletion of p53 induces abnormal BAT differentiation and is unprotected against diet-induced obesity [32]. This phenomenon may be caused by the loss of p53 could not suppress the expression of PPARγ in WATs and induction of energy expenditure in BATs. Consistent with these findings, Gan et al. demonstrated that Forkhead box C2 (Foxc2) suppressed differentiation and enhanced proliferation in preadipocytes [75]. Foxc2 is a forkhead/winged-helix transcription factor family protein. It plays an important role in cell growth, proliferation, differentiation, apoptosis, longevity, and energy metabolism [76,77]. Gan et al. showed that feeding a high-fat diet upregulated the expression of Foxc2 in mouse preadipocytes, which increased the expression of cyclin E and inhibited that of p53 and p27 in preadipocytes (Figure 1).
On the other hand, Hallenborg et al. showed that depletion of p53 in mice protected them against high-fat diet-induced obesity [47]. Loss of p53 increased the transcriptional activity of Pgc-1α and Pgc-1β and upregulated the expression of Ucp1 in MEF cells derived from p53 mutant mice. Importantly, they also showed that BATs from the p53 mutant mice were phenotypically different than the regular BAT, and that p53 mutant-expressing brown adipocytes more closely resemble beige adipocytes. Consistent with these results, whole p53 KO mice fed a high-fat diet show less body weight gain than WT littermates; however, conditional genetic mutation of p53 in BAT did not affect body weight gain or thermogenic activity [78]. Although this interesting study showed controversial phenotypes in mouse models fed a high-fat diet compared to other studies, these results suggested that p53 plays a critical role in brown adipocyte differentiation.
4. Role of P53 in Beige Adipocytes and Browning
p53 has been known to be involved in the aging process of WATs [79]. The total level of p53 has been found to be increased in the WATs of aged mice, and increased p53 level induces the expression of p21 in WATs, which further increases the release of proinflammatory cytokines and causes insulin resistance [79]. Furthermore, a recent study demonstrated that p53 regulates the browning of iWAT [80]. Elevated p53 suppressed the mitochondrial increase during browning in iWATs. In addition, they showed that transient depletion of p53 in the adipose tissue-specific p53 conditional knockout mouse model or using a pharmacological inhibitor (Pifithrin-α) restored cold-induced iWAT browning and whole-body energy metabolism in aged mice [80]. It is also possible that p53 regulates PRDM16 expression via interaction with PRDM16 or binding to the PRDM16 promoter [32,47]. However, the definitive role of p53 in beige adipogenesis and browning will require further study.
5. Conclusions
Over the past four decades, the roles of p53 have been extensively studied because the mutation or inhibition of p53 initiates tumorigenesis [1,14,81,82,83]. p53 has been found to be regulated by various stress signaling pathways and controls the cellular homeostasis response to stress [1,2]. p53 is the control tower of both intra- and extracellular stresses that activate various p53 pathways, which affects multidirectional physiological functions in various tissues. Recently, interesting studies using mouse models have demonstrated that p53 not only responds to cellular stress but is also involved in tissue or organ differentiation [81]. Recent advancements in determining the role of p53 in developmental biology have suggested its fundamental role in controlling cellular differentiation and why we need to improve our understanding of the different roles of p53. The newly identified biological roles of p53 in the regulation of adipogenesis and metabolic function have enabled researchers to study important and previously unexplored aspects of p53 function. For example, how do cancer cells affect metabolism or body fat mass in cancer patients? Depending on the cancer type, 30–80% of cancer patients develop cancer-associated cachexia (CAC) in the last stage of malignancy. Cancer cachexia leads to atrophy of adipose tissue and/or metabolic dysfunction, which accelerates the progression of cancer. Cancer cells secrete many inflammation mediators including nuclear factor kappa-light-chain-enhancer of activated B cells, tumor necrosis factor-a, interleukin-1, and interleukin-6 that have been shown to increase the metabolic expenditure through the activation of thermogenesis, inhibit differentiation of muscle and adipose tissues, and cause loss of appetite [84,85,86]. These inflammatory mediators can suppress the transcriptional activity of p53, and abnormal regulation of p53 could affect the initiation or progression of CAC [87]. A better understanding of p53 in regulating adipose tissue development and function will lead to the identification of novel therapeutic targets for CAC.
Heretofore, p53 role in adipogenesis, especially white adipocytes, is a negative regulator of adipogenesis. However, the role of p53 in negative regulation of adipogenesis under exact molecular upstream and downstream networks remains unknown. Generally, p53 is considered a stress response protein. WATs have been considered to be simple energy storage tissues containing lipid droplet organelles but they also have endocrine functions and secrete several factors such as adipokines [21]. The association between adipose tissues and p53 has become increasingly evident in the last few years. For example, excessive calorie intake could enhance the generation of reactive ROS in the adipose tissues and cause DNA damage, which could activate p53 [58,79]. Exogenous stress such as cardiac pressure can activate p53 in adipose tissue. Activation of the sympathetic nervous system upregulates adipose tissue p53 through increasing lipolysis, and the inhibition of p53 attenuates adipose tissue inflammation [58]. However, the upstream signals that activate p53 to induce stress signals remain unclear. The question remains as to whether p53 activation is stress-induced or depends on the tissue environment? Also, the environment in which p53 regulates lipid metabolism needs to be determined. Although several in vivo and in vitro studies have demonstrated the role of p53 in white adipocyte differentiation and function, the direct role of p53 in WAT formation needs to be determined. Since mutation of p53 results in the reduction of cell proliferation or growth, further investigation is needed to distinguish the direct role of p53 in the cellular proliferation and differentiation of adipocytes.
One of the major unsolved questions is the direct role of p53 in brown adipogenesis. Previously performed studies have reported contrasting results regarding the role of p53 in brown adipogenesis. Inhibition of p53 suppressed brown adipogenesis in Prdm16-overexpressed myogenic C2 cells in vitro [32]. On the other hand, p53 depletion enhanced the expression of brown adipogenic genes in MEFs and primary adipocytes derived from p53 knockout mice [47]. Similarly, the same mouse model showed different phenotypes in the high-fat diet experiment. These results have caused a lack of clarity in understanding the direct role of p53. Why did the depletion of p53 lead to the development of different phenotypes? The first reason may be the difference in cellular characteristics. Although these cells were derived from the same progenitor cells, different cellular characteristics may affect the response of p53 or cellular development. A pioneering study has suggested that brown adipocytes arise from the same progenitor cells as the skeletal muscle that expresses the transcription factor Myf5 [69]. Furthermore, this study demonstrated that brown adipocytes share many features with skeletal myocytes, and that loss of PRDM16 induces muscle differentiation in brown adipocyte progenitors. Supporting this hypothesis, loss of Ewing’s sarcoma breakpoint region 1 blocks maturation of brown adipocytes and induces myogenic differentiation in brown preadipocytes [23]. Physiologically, skeletal myocytes are rich in mitochondria and are specialized in lipid catabolism rather than lipid storage similar to brown adipocytes. Although skeletal myocytes share many features with brown adipocytes, they exhibit different characteristics. For example, they can express Pgc-1α; however, myocytes do not express Ucp1. These differences induce a different response to the expression of adipogenic genes. The second reason is the unique characteristic of p53, which has specific roles dependent on tissue or development stage. Omar Al-Massadi et al. showed that pharmacological or genetic mutation of p53 in BAT affects thermogenic programs and body weight in adult male mice, but not in embryos [78]. Additionally, they showed that pharmacological or genetic stimulation of p53 in the BAT of obese mice protects them from obesity and induces thermogenic activity. The third reason may be the original role of p53. p53 regulates endogenous or exogenous cellular stress. Hallenborg et al. suggested that housing temperatures, housing styles (group or individual), or differences in the gut microbiota could affect these contradictory results [47]. However, it is notable that normal housing temperatures could not suppress Ucp1 expression in p53-deficient mice [47]. These differences may be due to differences in behavior and mouse housing styles. Locomotion activities (horizontal, total distance traveled, number of movements, stereotypy counts) were significantly increased in neuronal-specific conditional p53 KO mice compared to wild-type mice [88]. In addition, individually housed mice had high nest scores, low body weight, and increased sucrose and food consumption [89]. These multiple factors may affect the contradictory phenotypes of p53 mice. However, further studies are needed to clarify these contradictory results.
The other mechanism by which p53 regulates cellular differentiation involves its chromatin-binding activity or interaction with specific protein partners. As described above, p53 affects adipogenesis dependent cell types in vitro. These results suggest that promoter remodeling following p53 binding to chromatin occurs at different sites in different adipogenic stages or binding with different protein partners in each cell. BATs and WATs share many essential adipogenic regulatory transcriptional factors including PPARγ and CEBPα. If p53 regulates promoter remodeling in the same way, there is no different response to adipogenic development regardless of the cell type. p53 interacts with chromatin response to diverse signaling and regulates transcription of specific target genes with or without co-binding factors [90,91,92]. Supporting this possibility, Chip analysis results showed that p53 binds to PRDM16 promoter regions in primary brown adipocytes [32]. In addition, global protein-protein interaction analysis by proteomics screening showed that the p53-Prdm16 or p53-Pgc-1a protein complex regulates adipocyte differentiation or function [47,93].
The role of p53 in tumorigenesis has been extensively studied because the activation of p53 kills cancer cells [94]. Many researchers have screened to find p53 modulator from herbal medicines to single molecules. However, due to multi-functional roles of p53 in diverse tissues, keeping a homeostatic balance of p53 is most important. Recently, studies have suggested that herbal medicines or single compounds from natural products might modulate p53 [95]. Nontoxic herbal medicines from plants, fruits, insects, and animals can provide novel p53 modulators. In this short review, we highlighted the roles of p53 in WAT and BAT adipogenesis and function. It is now important to carefully study the role of p53 in adipocytes. In the case of metabolic dysfunction, essential tissues are exposed to chronic cellular stress, and abnormal adipogenesis or metabolic dysfunction resulting from activation of p53 can cause more severe chronic stress. Considering the unique function of BATs, research on the role of p53 in these tissues could provide a novel therapeutic approach for treating metabolic disease. Although there are still many unanswered questions and contrasting results, this review highlights and explains the important role of p53 in adipocytes.
Author Contributions
Conceptualization, Y.K.L. and J.H.P.; visualization, Y.S.C.; writing—original draft, Y.K.L. and J.H.P.; Writing—review and editing, Y.K.L., Y.S.C., J.H.L., J.M.C. and J.H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant on the Development of Sustainable Application for Standard Herbal Resources from the Korea Institute of Oriental Medicine (KSN2012320) and Basic Science Research Program (NRF-2019R1F1A1050505) through the National Research Foundation of Korea (NRF).
Acknowledgments
We thank the members of Herbal Medicine Resource Center.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Finlay, C.A.; Hinds, P.W.; Levine, A.J. The p53 proto-oncogene can act as a suppressor of transformation. Cell 1989, 57, 1083–1093. [Google Scholar] [CrossRef]
- Horn, H.F.; Vousden, K.H. Coping with stress: Multiple ways to activate p53. Oncogene 2007, 26, 1306–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashcroft, M.; Kubbutat, M.H.; Vousden, K.H. Regulation of p53 function and stability by phosphorylation. Mol. Cell. Biol. 1999, 19, 1751–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olovnikov, I.A.; Kravchenko, J.E.; Chumakov, P.M. Homeostatic functions of the p53 tumor suppressor: Regulation of energy metabolism and antioxidant defense. Semin. Cancer Biol. 2009, 19, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.G.; Plas, D.R.; Kubek, S.; Buzzai, M.; Mu, J.; Xu, Y.; Birnbaum, M.J.; Thompson, C.B. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol. Cell 2005, 18, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M.; Razmara, M.; Nguyen, D.; Donahue, R.J.; Wubah, J.A.; Knudsen, T.B. Altered expression of mitochondrial 16S ribosomal RNA in p53-deficient mouse embryos revealed by differential display. Biochim. Biophys. Acta 1998, 1403, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Sung, H.J.; Park, J.Y.; Matoba, S.; Hwang, P.M. A pivotal role for p53: Balancing aerobic respiration and glycolysis. J. Bioenerg. Biomembr. 2007, 39, 243–246. [Google Scholar] [CrossRef]
- Buzzai, M.; Jones, R.G.; Amaravadi, R.K.; Lum, J.J.; DeBerardinis, R.J.; Zhao, F.; Viollet, B.; Thompson, C.B. Systemic treatment with the antidiabetic drug metformin selectively impairs p53-deficient tumor cell growth. Cancer Res. 2007, 67, 6745–6752. [Google Scholar] [CrossRef] [Green Version]
- Buckbinder, L.; Talbott, R.; Velasco-Miguel, S.; Takenaka, I.; Faha, B.; Seizinger, B.R.; Kley, N. Induction of the growth inhibitor IGF-binding protein 3 by p53. Nature 1995, 377, 646–649. [Google Scholar] [CrossRef]
- Polyak, K.; Xia, Y.; Zweier, J.L.; Kinzler, K.W.; Vogelstein, B. A model for p53-induced apoptosis. Nature 1997, 389, 300–305. [Google Scholar] [CrossRef]
- Yoon, K.A.; Nakamura, Y.; Arakawa, H. Identification of ALDH4 as a p53-inducible gene and its protective role in cellular stresses. J. Hum. Genet. 2004, 49, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Feng, Z.; Teresky, A.K.; Levine, A.J. p53 regulates maternal reproduction through LIF. Nature 2007, 450, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.F.; Kaufman, M.H.; Harrison, D.J.; Clarke, A.R. High-frequency developmental abnormalities in p53-deficient mice. Curr. Biol. 1995, 5, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Matheu, A.; Maraver, A.; Serrano, M. The Arf/p53 pathway in cancer and aging. Cancer Res. 2008, 68, 6031–6034. [Google Scholar] [CrossRef] [Green Version]
- Haupt, Y.; Maya, R.; Kazaz, A.; Oren, M. Mdm2 promotes the rapid degradation of p53. Nature 1997, 387, 296–299. [Google Scholar] [CrossRef]
- Lavin, M.F.; Gueven, N. The complexity of p53 stabilization and activation. Cell Death Differ. 2006, 13, 941–950. [Google Scholar] [CrossRef]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Soga, T. Cancer metabolism: Key players in metabolic reprogramming. Cancer Sci. 2013, 104, 275–281. [Google Scholar] [CrossRef]
- Schmid, P.; Lorenz, A.; Hameister, H.; Montenarh, M. Expression of p53 during mouse embryogenesis. Development 1991, 113, 857–865. [Google Scholar]
- Tedeschi, A.; Di Giovanni, S. The non-apoptotic role of p53 in neuronal biology: Enlightening the dark side of the moon. EMBO Rep. 2009, 10, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Cohen, P.; Spiegelman, B.M. Adaptive thermogenesis in adipocytes: Is beige the new brown? Genes Dev. 2013, 27, 234–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Kang, H.J.; Kang, S.I.; Lee, J.E.; Hur, J.; Ge, K.; Mueller, E.; Li, H.; Lee, B.C.; Lee, S.B. A multifunctional protein, EWS, is essential for early brown fat lineage determination. Dev. Cell 2013, 26, 393–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Hur, W.; Lee, S.B. Intricate Transcriptional Networks of Classical Brown and Beige Fat Cells. Front. Endocrinol. (Lausanne) 2015, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Seale, P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 2016, 17, 691–702. [Google Scholar] [CrossRef]
- Han, J.; Lee, J.E.; Jin, J.; Lim, J.S.; Oh, N.; Kim, K.; Chang, S.I.; Shibuya, M.; Kim, H.; Koh, G.Y. The spatiotemporal development of adipose tissue. Development 2011, 138, 5027–5037. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Chau, Y.Y.; Bandiera, R.; Serrels, A.; Martinez-Estrada, O.M.; Qing, W.; Lee, M.; Slight, J.; Thornburn, A.; Berry, R.; McHaffie, S.; et al. Visceral and subcutaneous fat have different origins and evidence supports a mesothelial source. Nat. Cell Biol. 2014, 16, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Di Girolamo, M.; Fine, J.B.; Tagra, K.; Rossmanith, R. Qualitative regional differences in adipose tissue growth and cellularity in male Wistar rats fed ad libitum. Am. J. Physiol. 1998, 274, R1460–R1467. [Google Scholar] [CrossRef]
- Liu, S.; Kim, T.H.; Franklin, D.A.; Zhang, Y. Protection against High-Fat-Diet-Induced Obesity in MDM2(C305F) Mice Due to Reduced p53 Activity and Enhanced Energy Expenditure. Cell Rep. 2017, 18, 1005–1018. [Google Scholar] [CrossRef]
- Yahagi, N.; Shimano, H.; Matsuzaka, T.; Najima, Y.; Sekiya, M.; Nakagawa, Y.; Ide, T.; Tomita, S.; Okazaki, H.; Tamura, Y.; et al. p53 Activation in adipocytes of obese mice. J. Biol. Chem. 2003, 278, 25395–25400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molchadsky, A.; Ezra, O.; Amendola, P.G.; Krantz, D.; Kogan-Sakin, I.; Buganim, Y.; Rivlin, N.; Goldfinger, N.; Folgiero, V.; Falcioni, R.; et al. p53 is required for brown adipogenic differentiation and has a protective role against diet-induced obesity. Cell Death Differ. 2013, 20, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Molchadsky, A.; Shats, I.; Goldfinger, N.; Pevsner-Fischer, M.; Olson, M.; Rinon, A.; Tzahor, E.; Lozano, G.; Zipori, D.; Sarig, R.; et al. p53 plays a role in mesenchymal differentiation programs, in a cell fate dependent manner. PLoS ONE 2008, 3, e3707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Liu, M.; Du, X.; Zhang, R.; Xue, Y.; Zhang, Y.; Zhu, W.; Li, D.; Zhao, A.; Liu, Y. Role of p53 in preadipocyte differentiation. Cell Biol. Int. 2014, 38, 1384–1393. [Google Scholar] [CrossRef]
- Chang, E.; Kim, C.Y. Natural Products and Obesity: A Focus on the Regulation of Mitotic Clonal Expansion during Adipogenesis. Molecules 2019, 24, 1157. [Google Scholar] [CrossRef] [Green Version]
- Naaz, A.; Holsberger, D.R.; Iwamoto, G.A.; Nelson, A.; Kiyokawa, H.; Cooke, P.S. Loss of cyclin-dependent kinase inhibitors produces adipocyte hyperplasia and obesity. FASEB J. 2004, 18, 1925–1927. [Google Scholar] [CrossRef]
- Inoue, N.; Yahagi, N.; Yamamoto, T.; Ishikawa, M.; Watanabe, K.; Matsuzaka, T.; Nakagawa, Y.; Takeuchi, Y.; Kobayashi, K.; Takahashi, A.; et al. Cyclin-dependent kinase inhibitor, p21WAF1/CIP1, is involved in adipocyte differentiation and hypertrophy, linking to obesity, and insulin resistance. J. Biol. Chem. 2008, 283, 21220–21229. [Google Scholar] [CrossRef] [Green Version]
- Hallenborg, P.; Petersen, R.K.; Feddersen, S.; Sundekilde, U.; Hansen, J.B.; Blagoev, B.; Madsen, L.; Kristiansen, K. PPARgamma ligand production is tightly linked to clonal expansion during initiation of adipocyte differentiation. J. Lipid Res. 2014, 55, 2491–2500. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.Q.; Gronborg, M.; Huang, H.; Kim, J.W.; Otto, T.C.; Pandey, A.; Lane, M.D. Sequential phosphorylation of CCAAT enhancer-binding protein beta by MAPK and glycogen synthase kinase 3beta is required for adipogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 9766–9771. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Evan, D.; Rosen, C.J.W.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar]
- Rosen, E.D.; Spiegelman, B.M. What We Talk About When We Talk About Fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okita, N.; Ishikawa, N.; Mizunoe, Y.; Oku, M.; Nagai, W.; Suzuki, Y.; Matsushima, S.; Mikami, K.; Okado, H.; Sasaki, T.; et al. Inhibitory effect of p53 on mitochondrial content and function during adipogenesis. Biochem. Biophys. Res. Commun. 2014, 446, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yang, X.; Zhao, Q.; Li, Z.; Fu, F.; Zhang, H.; Zheng, M.; Zhang, S. Molecular Mechanism of Stem Cell Differentiation into Adipocytes and Adipocyte Differentiation of Malignant Tumor. Stem Cells Int. 2020, 2020, 8892300. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular Mechanisms of Adipogenesis: The Anti-adipogenic Role of AMP-Activated Protein Kinase. Front. Mol. Biosci. 2020, 7, 76. [Google Scholar] [CrossRef]
- Vassilev, L.T.; Vu, B.T.; Graves, B.; Carvajal, D.; Podlaski, F.; Filipovic, Z.; Kong, N.; Kammlott, U.; Lukacs, C.; Klein, C.; et al. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science 2004, 303, 844–848. [Google Scholar] [CrossRef] [Green Version]
- Hallenborg, P.; Fjaere, E.; Liaset, B.; Petersen, R.K.; Murano, I.; Sonne, S.B.; Falkerslev, M.; Winther, S.; Jensen, B.A.; Ma, T.; et al. p53 regulates expression of uncoupling protein 1 through binding and repression of PPARgamma coactivator-1alpha. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E116–E128. [Google Scholar] [CrossRef] [Green Version]
- Yadav, N.; Cheng, D.; Richard, S.; Morel, M.; Iyer, V.R.; Aldaz, C.M.; Bedford, M.T. CARM1 promotes adipocyte differentiation by coactivating PPARgamma. EMBO Rep. 2008, 9, 193–198. [Google Scholar] [CrossRef]
- Behera, A.K.; Bhattacharya, A.; Vasudevan, M.; Kundu, T.K. p53 mediated regulation of coactivator associated arginine methyltransferase 1 (CARM1) expression is critical for suppression of adipogenesis. FEBS J. 2018, 285, 1730–1744. [Google Scholar] [CrossRef] [Green Version]
- Boregowda, S.V.; Krishnappa, V.; Strivelli, J.; Haga, C.L.; Booker, C.N.; Phinney, D.G. Basal p53 expression is indispensable for mesenchymal stem cell integrity. Cell Death Differ. 2018, 25, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Berberich, S.J.; Litteral, V.; Mayo, L.D.; Tabesh, D.; Morris, D. mdm-2 gene amplification in 3T3-L1 preadipocytes. Differentiation 1999, 64, 205–212. [Google Scholar] [CrossRef]
- Liu, Z.; Jin, L.; Yang, J.K.; Wang, B.; Wu, K.K.L.; Hallenborg, P.; Xu, A.; Cheng, K.K.Y. The Dysfunctional MDM2-p53 Axis in Adipocytes Contributes to Aging-Related Metabolic Complications by Induction of Lipodystrophy. Diabetes 2018, 67, 2397–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallenborg, P.; Siersbaek, M.; Barrio-Hernandez, I.; Nielsen, R.; Kristiansen, K.; Mandrup, S.; Grontved, L.; Blagoev, B. MDM2 facilitates adipocyte differentiation through CRTC-mediated activation of STAT3. Cell Death Dis. 2016, 7, e2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallenborg, P.; Feddersen, S.; Francoz, S.; Murano, I.; Sundekilde, U.; Petersen, R.K.; Akimov, V.; Olson, M.V.; Lozano, G.; Cinti, S.; et al. Mdm2 controls CREB-dependent transactivation and initiation of adipocyte differentiation. Cell Death Differ. 2012, 19, 1381–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmel, A.R.; Sztalryd, C. The Perilipins: Major Cytosolic Lipid Droplet-Associated Proteins and Their Roles in Cellular Lipid Storage, Mobilization, and Systemic Homeostasis. Annu. Rev. Nutr. 2016, 36, 471–509. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Sohn, J.H.; Han, J.S.; Park, Y.J.; Jeon, Y.G.; Ji, Y.; Dalen, K.T.; Sztalryd, C.; Kimmel, A.R.; Kim, J.B. Perilipin 3 Deficiency Stimulates Thermogenic Beige Adipocytes Through PPARalpha Activation. Diabetes 2018, 67, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, B.; Nerstedt, A.; Smith, U. Reduced subcutaneous adipogenesis in human hypertrophic obesity is linked to senescent precursor cells. Nat. Commun. 2019, 10, 2757. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Yoshida, Y.; Katsuno, T.; Tateno, K.; Okada, S.; Moriya, J.; Yokoyama, M.; Nojima, A.; Ito, T.; Zechner, R.; et al. p53-induced adipose tissue inflammation is critically involved in the development of insulin resistance in heart failure. Cell Metab. 2012, 15, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Schupp, M.; Chen, F.; Briggs, E.R.; Rao, S.; Pelzmann, H.J.; Pessentheiner, A.R.; Bogner-Strauss, J.G.; Lazar, M.A.; Baldwin, D.; Prokesch, A. Metabolite and transcriptome analysis during fasting suggest a role for the p53-Ddit4 axis in major metabolic tissues. BMC Genom. 2013, 14, 758. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xu, M.; Peng, Y.; Naren, Q.; Xu, Y.; Wang, X.; Yang, G.; Shi, X.; Li, X. Triptolide enhances lipolysis of adipocytes by enhancing ATGL transcription via upregulation of p53. Phytother. Res. 2020. [Google Scholar] [CrossRef]
- Kung, C.P.; Leu, J.I.; Basu, S.; Khaku, S.; Anokye-Danso, F.; Liu, Q.; George, D.L.; Ahima, R.S.; Murphy, M.E. The P72R Polymorphism of p53 Predisposes to Obesity and Metabolic Dysfunction. Cell Rep. 2016, 14, 2413–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derdak, Z.; Villegas, K.A.; Harb, R.; Wu, A.M.; Sousa, A.; Wands, J.R. Inhibition of p53 attenuates steatosis and liver injury in a mouse model of non-alcoholic fatty liver disease. J. Hepatol. 2013, 58, 785–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handschin, C.; Spiegelman, B.M. Peroxisome proliferator-activated receptor gamma coactivator 1 coactivators, energy homeostasis, and metabolism. Endocr. Rev. 2006, 27, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S. The adipose organ. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 9–15. [Google Scholar] [CrossRef]
- Hirning, U.; Schmid, P.; Schulz, W.A.; Kozak, L.P.; Hameister, H. In developing brown adipose tissue c-myc protooncogene expression is restricted to early differentiation stages. Cell Differ. Dev. 1989, 27, 243–248. [Google Scholar] [CrossRef]
- Seale, P.; Kajimura, S.; Yang, W.; Chin, S.; Rohas, L.M.; Uldry, M.; Tavernier, G.; Langin, D.; Spiegelman, B.M. Transcriptional control of brown fat determination by PRDM16. Cell Metab. 2007, 6, 38–54. [Google Scholar] [CrossRef] [Green Version]
- Harms, M.J.; Ishibashi, J.; Wang, W.; Lim, H.W.; Goyama, S.; Sato, T.; Kurokawa, M.; Won, K.J.; Seale, P. Prdm16 is required for the maintenance of brown adipocyte identity and function in adult mice. Cell Metab. 2014, 19, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, J.; Seale, P. Functions of Prdm16 in thermogenic fat cells. Temperature (Austin) 2015, 2, 65–72. [Google Scholar] [CrossRef]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scime, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Westerberg, R.; Mansson, J.E.; Golozoubova, V.; Shabalina, I.G.; Backlund, E.C.; Tvrdik, P.; Retterstol, K.; Capecchi, M.R.; Jacobsson, A. ELOVL3 is an important component for early onset of lipid recruitment in brown adipose tissue. J. Biol. Chem. 2006, 281, 4958–4968. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Yon Toh, S.; Chen, Z.; Guo, K.; Ng, C.P.; Ponniah, S.; Lin, S.C.; Hong, W.; Li, P. Cidea-deficient mice have lean phenotype and are resistant to obesity. Nat. Genet. 2003, 35, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Barbera, M.J.; Schluter, A.; Pedraza, N.; Iglesias, R.; Villarroya, F.; Giralt, M. Peroxisome proliferator-activated receptor alpha activates transcription of the brown fat uncoupling protein-1 gene. A link between regulation of the thermogenic and lipid oxidation pathways in the brown fat cell. J. Biol. Chem. 2001, 276, 1486–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jash, S.; Banerjee, S.; Lee, M.J.; Farmer, S.R.; Puri, V. CIDEA Transcriptionally Regulates UCP1 for Britening and Thermogenesis in Human Fat Cells. iScience 2019, 20, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Fernand, V.E.; Henagan, T.M.; Shin, J.; Huypens, P.; Newman, S.; Gettys, T.W.; Chang, J.S. Regulation of Brown and White Adipocyte Transcriptome by the Transcriptional Coactivator NT-PGC-1alpha. PLoS ONE 2016, 11, e0159990. [Google Scholar] [CrossRef]
- Gan, L.; Liu, Z.; Jin, W.; Zhou, Z.; Sun, C. Foxc2 enhances proliferation and inhibits apoptosis through activating Akt/mTORC1 signaling pathway in mouse preadipocytes. J. Lipid Res. 2015, 56, 1471–1480. [Google Scholar] [CrossRef] [Green Version]
- Hader, C.; Marlier, A.; Cantley, L. Mesenchymal-epithelial transition in epithelial response to injury: The role of Foxc2. Oncogene 2010, 29, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Cederberg, A.; Gronning, L.M.; Ahren, B.; Tasken, K.; Carlsson, P.; Enerback, S. FOXC2 is a winged helix gene that counteracts obesity, hypertriglyceridemia, and diet-induced insulin resistance. Cell 2001, 106, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Al-Massadi, O.; Porteiro, B.; Kuhlow, D.; Köhler, M.; Gonzalez-Rellan, M.J.; Garcia-Lavandeira, M.; Díaz-Rodríguez, E.; Quiñones, M.; Senra, A.; Alvarez, C.V.; et al. Pharmacological and Genetic Manipulation of p53 in Brown Fat at Adult But Not Embryonic Stages Regulates Thermogenesis and Body Weight in Male Mice. Endocrinology 2016, 157, 2735–2749. [Google Scholar] [CrossRef] [Green Version]
- Minamino, T.; Orimo, M.; Shimizu, I.; Kunieda, T.; Yokoyama, M.; Ito, T.; Nojima, A.; Nabetani, A.; Oike, Y.; Matsubara, H.; et al. A crucial role for adipose tissue p53 in the regulation of insulin resistance. Nat. Med. 2009, 15, 1082–1087. [Google Scholar] [CrossRef]
- Fu, W.; Liu, Y.; Sun, C.; Yin, H. Transient p53 inhibition sensitizes aged white adipose tissue for beige adipocyte recruitment by blocking mitophagy. FASEB J. 2019, 33, 844–856. [Google Scholar] [CrossRef]
- Lozano, G. Mouse models of p53 functions. Cold Spring Harb. Perspect. Biol. 2010, 2, a001115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, A.M.; Dong, Z. Post-translational modification of p53 in tumorigenesis. Nat. Rev. Cancer 2004, 4, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Stiewe, T. The p53 family in differentiation and tumorigenesis. Nat. Rev. Cancer 2007, 7, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Pfitzenmaier, J.; Vessella, R.; Higano, C.S.; Noteboom, J.L.; Wallace, D., Jr.; Corey, E. Elevation of cytokine levels in cachectic patients with prostate carcinoma. Cancer 2003, 97, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Shen, S.; Verma, I.M. NF-kappaB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petruzzelli, M.; Wagner, E.F. Mechanisms of metabolic dysfunction in cancer-associated cachexia. Genes Dev. 2016, 30, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Gudkov, A.V.; Gurova, K.V.; Komarova, E.A. Inflammation and p53: A Tale of Two Stresses. Genes Cancer 2011, 2, 503–516. [Google Scholar] [CrossRef]
- Filichia, E.; Shen, H.; Zhou, X.; Qi, X.; Jin, K.; Greig, N.; Hoffer, B.; Luo, Y. Forebrain neuronal specific ablation of p53 gene provides protection in a cortical ischemic stroke model. Neuroscience 2015, 295, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jirkof, P.; Bratcher, N.; Medina, L.; Strasburg, D.; Ebert, P.; Gaskill, B.N. The effect of group size, age and handling frequency on inter-male aggression in CD 1 mice. Sci. Rep. 2020, 10, 2253. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, J.M.; Emerson, B.M. Transcriptional regulation by p53 through intrinsic DNA/chromatin binding and site-directed cofactor recruitment. Mol. Cell 2001, 8, 57–69. [Google Scholar] [CrossRef]
- Wei, C.L.; Wu, Q.; Vega, V.B.; Chiu, K.P.; Ng, P.; Zhang, T.; Shahab, A.; Yong, H.C.; Fu, Y.; Weng, Z.; et al. A global map of p53 transcription-factor binding sites in the human genome. Cell 2006, 124, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, K.D.; Galbraith, M.D.; Andrysik, Z.; Espinosa, J.M. Mechanisms of transcriptional regulation by p53. Cell Death Differ. 2018, 25, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajimura, S.; Seale, P.; Kubota, K.; Lunsford, E.; Frangioni, J.V.; Gygi, S.P.; Spiegelman, B.M. Initiation of myoblast to brown fat switch by a PRDM16-C/EBP-beta transcriptional complex. Nature 2009, 460, 1154–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Sun, Y. Targeting p53 for Novel Anticancer Therapy. Transl. Oncol. 2010, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.J.; Li, X.; Hunt, C.; Wang, W.; Wang, H.; Zhang, R. Natural products targeting the p53-MDM2 pathway and mutant p53: Recent advances and implications in cancer medicine. Genes Dis. 2018, 5, 204–219. [Google Scholar] [CrossRef]
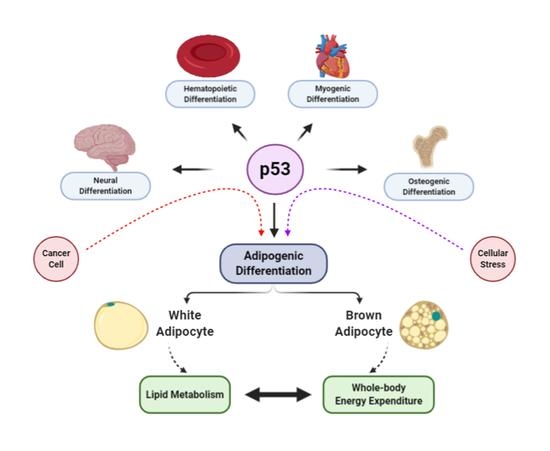
Figure 1.
Proposed hypothetical model of the transcriptional role of p53 in white and brown adipogenesis.
Figure 1.
Proposed hypothetical model of the transcriptional role of p53 in white and brown adipogenesis.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, Y.K.; Chung, Y.S.; Lee, J.H.; Chun, J.M.; Park, J.H. The Intricate Role of p53 in Adipocyte Differentiation and Function. Cells 2020, 9, 2621. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9122621
AMA Style
Lee YK, Chung YS, Lee JH, Chun JM, Park JH. The Intricate Role of p53 in Adipocyte Differentiation and Function. Cells. 2020; 9(12):2621. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9122621
Chicago/Turabian StyleLee, Yun Kyung, Yu Seong Chung, Ji Hye Lee, Jin Mi Chun, and Jun Hong Park. 2020. "The Intricate Role of p53 in Adipocyte Differentiation and Function" Cells 9, no. 12: 2621. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9122621
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.