
Influence of TGFBR2, TGFB3, DNMT1, and DNMT3A Knockdowns on CTGF, TGFBR2, and DNMT3A in Neonatal and Adult Human Dermal Fibroblasts Cell Lines

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Transfection
2.3. Western Blot
2.4. Protein Specificity Test
2.5. Endogenous In Vitro Protein Level
2.6. Statistical Analysis
3. Results
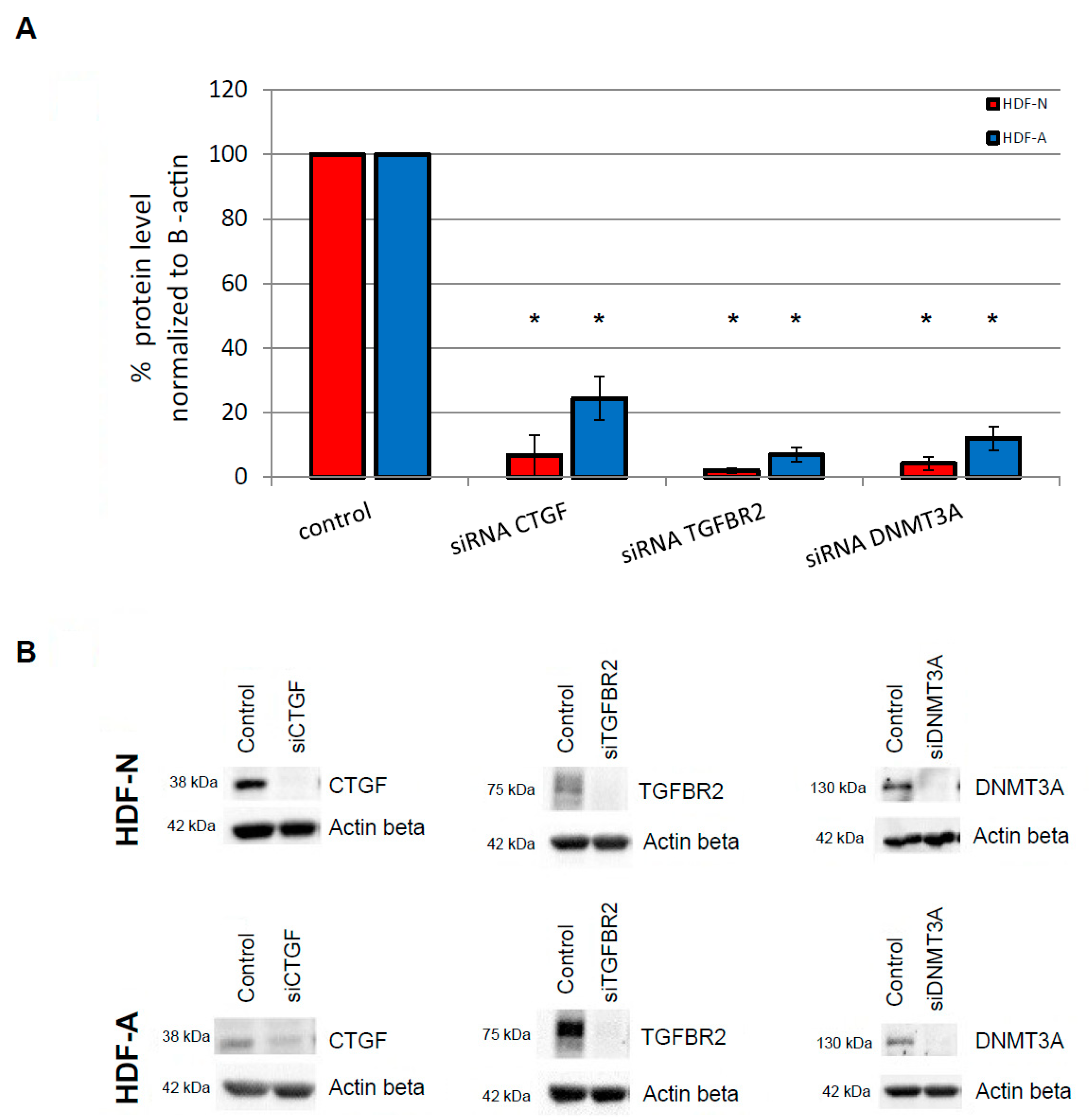
3.1. Protein Specificity Test
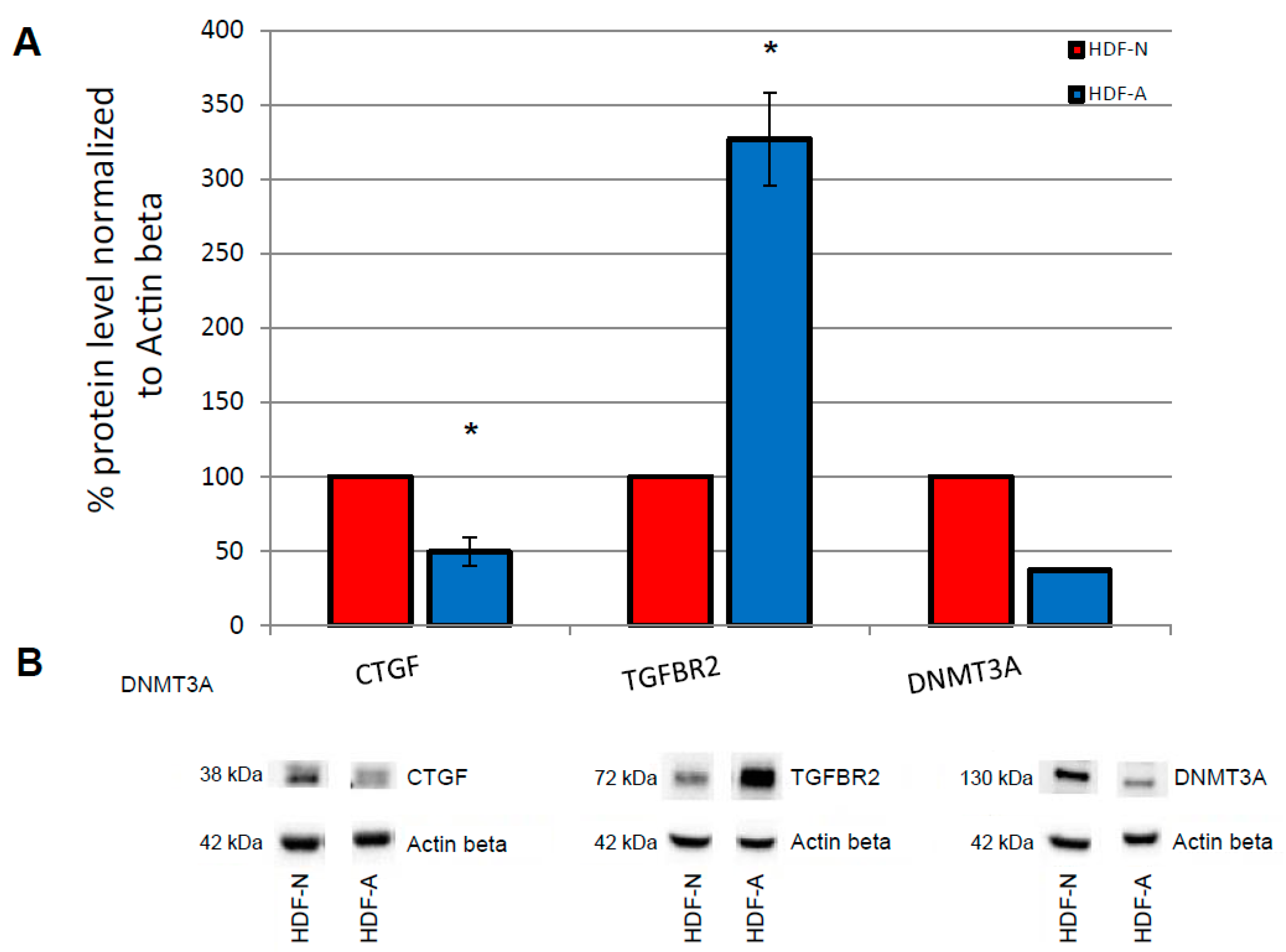
3.2. Endogenous In Vitro Levels of Proteins Associated with the TGF-β Pathway and DNA Methyltransferases
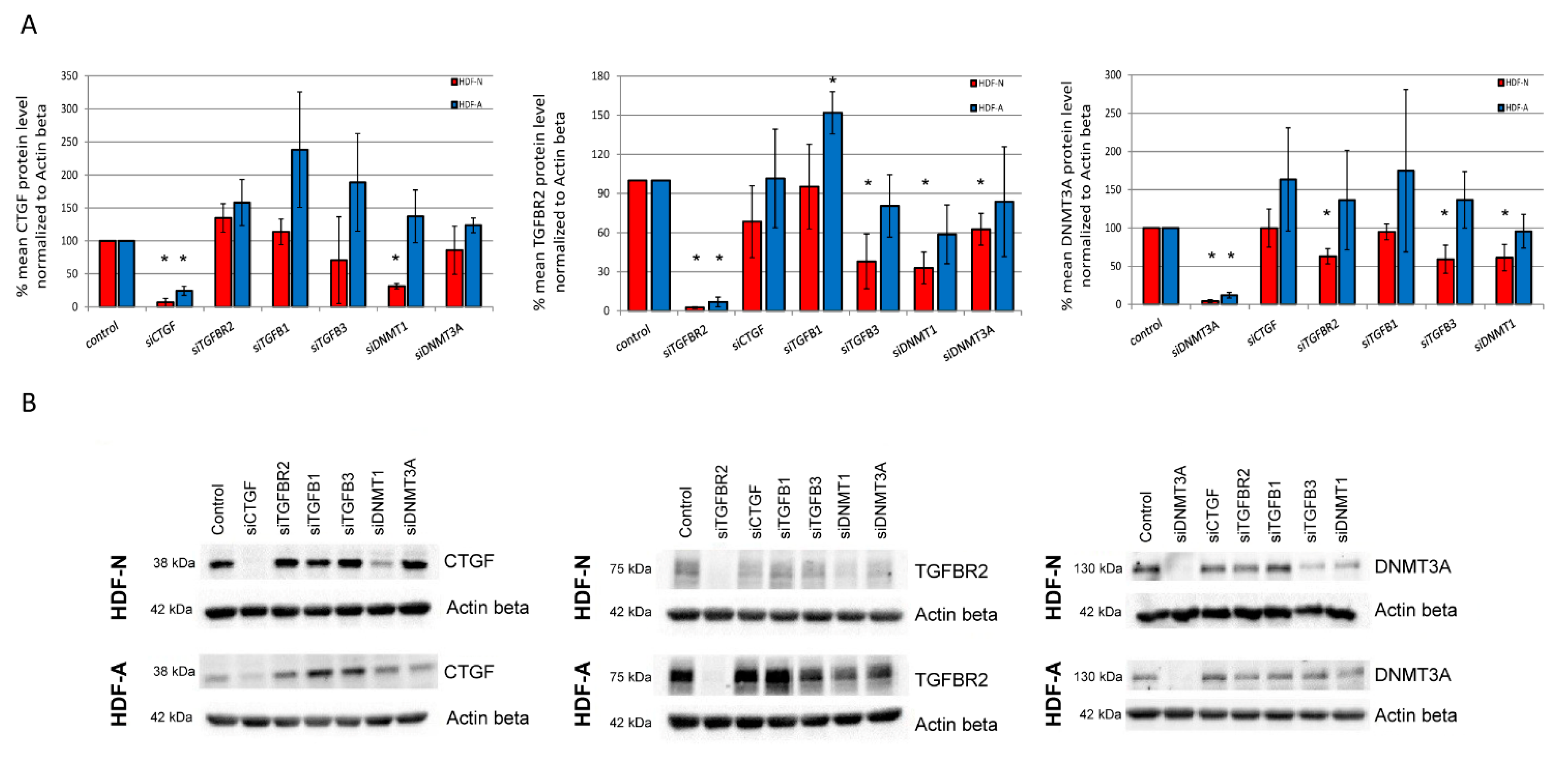
3.3. The In Vitro Effect of Analyzed Genes on the CTGF, TGFBR2, and DNMT3A Protein Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matsui, M.S.; Pelle, E.; Dong, K.; Pernodet, N. Biological Rhythms in the Skin. Int. J. Mol. Sci. 2016, 17, 801. [Google Scholar] [CrossRef]
- Plikus, M.V.; Van Spyk, E.N.; Pham, K.; Geyfman, M.; Kumar, V.; Takahashi, J.S.; Andersen, B. The Circadian Clock in Skin: Implications for Adult Stem Cells, Tissue Regeneration, Cancer, Aging, and Immunity. J. Biol. Rhythm. 2015, 30, 163–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandu, C.; Liu, T.; Malan, A.; Challet, E.; Pévet, P.; Felder-Schmittbuhl, M.-P. Circadian Clocks in Rat Skin and Dermal Fibroblasts: Differential Effects of Aging, Temperature and Melatonin. Cell Mol. Life Sci. 2015, 72, 2237–2248. [Google Scholar] [CrossRef] [PubMed]
- Stunova, A.; Vistejnova, L. Dermal Fibroblasts-A Heterogeneous Population with Regulatory Function in Wound Healing. Cytokine Growth Factor Rev. 2018, 39, 137–150. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, T.; Fisher, G.J. Role of Age-Associated Alterations of the Dermal Extracellular Matrix Microenvironment in Human Skin Aging: A Mini-Review. Gerontology 2015, 61, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krejčí, E.; Kodet, O.; Szabo, P.; Borský, J.; Smetana, K.; Grim, M.; Dvořánková, B. In Vitro Differences of Neonatal and Later Postnatal Keratinocytes and Dermal Fibroblasts. Physiol. Res. 2015, 64, 561–569. [Google Scholar] [CrossRef]
- Mateu, R.; Živicová, V.; Krejčí, E.D.; Grim, M.; Strnad, H.; Vlček, Č.; Kolář, M.; Lacina, L.; Gál, P.; Borský, J.; et al. Functional Differences between Neonatal and Adult Fibroblasts and Keratinocytes: Donor Age Affects Epithelial-Mesenchymal Crosstalk in Vitro. Int. J. Mol. Med. 2016, 38, 1063–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, K.J.; Blobe, G.C. Role of Transforming Growth Factor-Beta Superfamily Signaling Pathways in Human Disease. Biochim. Biophys. Acta 2008, 1782, 197–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakyari, M.; Farrokhi, A.; Maharlooei, M.K.; Ghahary, A. Critical Role of Transforming Growth Factor Beta in Different Phases of Wound Healing. Adv. Wound Care 2013, 2, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihn, H. Pathogenesis of Fibrosis: Role of TGF-Beta and CTGF. Curr. Opin. Rheumatol. 2002, 14, 681–685. [Google Scholar] [CrossRef]
- Abreu, J.G.; Ketpura, N.I.; Reversade, B.; De Robertis, E.M. Connective-Tissue Growth Factor (CTGF) Modulates Cell Signalling by BMP and TGF-β. Nat. Cell Biol. 2002, 4, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurkowska, R.Z.; Jurkowski, T.P.; Jeltsch, A. Structure and Function of Mammalian DNA Methyltransferases. Chembiochem 2011, 12, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Attisano, L. The TGFbeta Superfamily Signaling Pathway. Wiley Interdiscip Rev. Dev. Biol. 2013, 2, 47–63. [Google Scholar] [CrossRef]
- Hata, A.; Chen, Y.-G. TGF-β Signaling from Receptors to Smads. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, V. TGF-β Signaling in Cancer. J. Cell. Biochem. 2016, 117, 1279–1287. [Google Scholar] [CrossRef]
- Tatler, A.L.; Jenkins, G. TGF-β Activation and Lung Fibrosis. Proc. Am. Thorac. Soc. 2012, 9, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.S.; Poulsen, E.T.; Lukassen, M.V.; Shern, C.C.; Mogensen, E.H.; Weberskov, C.E.; De Dionisio, L.; Schauser, L.; Moore, T.C.B.; Otzen, D.E.; et al. Biochemical Mechanisms of Aggregation in TGFBI-Linked Corneal Dystrophies. Prog. Retin. Eye Res. 2020, 100843. [Google Scholar] [CrossRef]
- Kabza, M.; Karolak, J.A.; Rydzanicz, M.; Szcześniak, M.W.; Nowak, D.M.; Ginter-Matuszewska, B.; Polakowski, P.; Ploski, R.; Szaflik, J.P.; Gajecka, M. Collagen Synthesis Disruption and Downregulation of Core Elements of TGF-β, Hippo, and Wnt Pathways in Keratoconus Corneas. Eur. J. Hum. Genet. 2017, 25, 582–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarsini, S.; McKay, T.B.; Sarker-Nag, A.; Karamichos, D. Keratoconus in Vitro and the Key Players of the TGF-β Pathway. Mol. Vis. 2015, 21, 577–588. [Google Scholar] [PubMed]
- Evans, C.J.; Davidson, A.E.; Carnt, N.; López, K.E.R.; Veli, N.; Thaung, C.M.; Tuft, S.J.; Hardcastle, A.J. Genotype-Phenotype Correlation for TGFBI Corneal Dystrophies Identifies p.(G623D) as a Novel Cause of Epithelial Basement Membrane Dystrophy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5407–5414. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, I.E.; Eickelberg, O. The Impact of TGF-β on Lung Fibrosis: From Targeting to Biomarkers. Proc. Am. Thorac. Soc. 2012, 9, 111–116. [Google Scholar] [CrossRef]
- Quan, T.; He, T.; Kang, S.; Voorhees, J.J.; Fisher, G.J. Connective Tissue Growth Factor: Expression in Human Skin in Vivo and Inhibition by Ultraviolet Irradiation. J. Investig. Dermatol. 2002, 118, 402–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, T.; Shao, Y.; He, T.; Voorhees, J.J.; Fisher, G.J. Reduced Expression of Connective Tissue Growth Factor (CTGF/CCN2) Mediates Collagen Loss in Chronologically Aged Human Skin. J. Investig. Dermatol. 2010, 130, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Živicová, V.; Lacina, L.; Mateu, R.; Smetana, K.; Kavková, R.; Krejčí, E.D.; Grim, M.; Kvasilová, A.; Borský, J.; Strnad, H.; et al. Analysis of Dermal Fibroblasts Isolated from Neonatal and Child Cleft Lip and Adult Skin: Developmental Implications on Reconstructive Surgery. Int. J. Mol. Med. 2017, 40, 1323–1334. [Google Scholar] [CrossRef] [Green Version]
- Gardner, H.; Strehlow, D.; Bradley, L.; Widom, R.; Farina, A.; de Fougerolles, A.; Peyman, J.; Koteliansky, V.; Korn, J.H. Global Expression Analysis of the Fibroblast Transcriptional Response to TGFbeta. Clin. Exp. Rheumatol. 2004, 22, S47–S57. [Google Scholar]
- Oliver, N.; Sternlicht, M.; Gerritsen, K.; Goldschmeding, R. Could Aging Human Skin Use a Connective Tissue Growth Factor Boost to Increase Collagen Content? J. Investig. Derm. 2010, 130, 338–341. [Google Scholar] [CrossRef] [Green Version]
- Rolfe, K.J.; Richardson, J.; Vigor, C.; Irvine, L.M.; Grobbelaar, A.O.; Linge, C. A Role for TGF-Beta1-Induced Cellular Responses during Wound Healing of the Non-Scarring Early Human Fetus? J. Investig. Dermatol. 2007, 127, 2656–2667. [Google Scholar] [CrossRef]
- Quan, T.; He, T.; Shao, Y.; Lin, L.; Kang, S.; Voorhees, J.J.; Fisher, G.J. Elevated Cysteine-Rich 61 Mediates Aberrant Collagen Homeostasis in Chronologically Aged and Photoaged Human Skin. Am. J. Pathol. 2006, 169, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokhortchouk, E.; Defossez, P.-A. The Cell Biology of DNA Methylation in Mammals. Biochim. Biophys. Acta 2008, 1783, 2167–2173. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Chen, Z.; Huang, R.; Yao, Y.; Ma, G. Transforming Growth Factor Β1 Induces the Expression of Collagen Type I by DNA Methylation in Cardiac Fibroblasts. PLoS ONE 2013, 8, e60335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, H.B.; Scruggs, A.M.; Huang, S.K. Transforming Growth Factor-Β1 Increases DNA Methyltransferase 1 and 3a Expression through Distinct Post-Transcriptional Mechanisms in Lung Fibroblasts. J. Biol. Chem. 2016, 291, 19287–19298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dees, C.; Pötter, S.; Zhang, Y.; Bergmann, C.; Zhou, X.; Luber, M.; Wohlfahrt, T.; Karouzakis, E.; Ramming, A.; Gelse, K.; et al. TGF-β-Induced Epigenetic Deregulation of SOCS3 Facilitates STAT3 Signaling to Promote Fibrosis. J. Clin. Investig. 2020, 130, 2347–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, A.L.; Bartz, S.R.; Schelter, J.; Kobayashi, S.V.; Burchard, J.; Mao, M.; Li, B.; Cavet, G.; Linsley, P.S. Expression Profiling Reveals Off-Target Gene Regulation by RNAi. Nat. Biotechnol. 2003, 21, 635–637. [Google Scholar] [CrossRef]
- Scacheri, P.C.; Rozenblatt-Rosen, O.; Caplen, N.J.; Wolfsberg, T.G.; Umayam, L.; Lee, J.C.; Hughes, C.M.; Shanmugam, K.S.; Bhattacharjee, A.; Meyerson, M.; et al. Short Interfering RNAs Can Induce Unexpected and Divergent Changes in the Levels of Untargeted Proteins in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2004, 101, 1892–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Analyzed Protein | Specific Gene Targeted siRNA | ||||||
|---|---|---|---|---|---|---|---|
| siCTGF | siTGFBR2 | siTGFB1 | siTGFB3 | siDNMT1 | siDNMT3A | ||
| CTGF | HDF-N | ↓ p = 0.001 | N/E | N/E | N/E | ↓ p = 0.001 | N/E |
| HDF-A | ↓ p = 0.003 | N/E | N/E | N/E | N/E | N/E | |
| TGFBR2 | HDF-N | N/E | ↓ p < 0.001 | N/E | ↓ p = 0.053 | ↓ p = 0.015 | ↓ p = 0.053 |
| HDF-A | N/E | ↓ p < 0.001 | ↑ p = 0.050 | N/E | N/E | N/E | |
| DNMT3A | HDF-N | N/E | ↓ p = 0.011 | N/E | ↓ p = 0.050 | ↓ p = 0.050 | ↓ p < 0.001 |
| HDF-A | N/E | N/E | N/E | N/E | N/E | ↓ p = 0.007 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomela, K.; Karolak, J.A.; Ginter-Matuszewska, B.; Kabza, M.; Gajecka, M. Influence of TGFBR2, TGFB3, DNMT1, and DNMT3A Knockdowns on CTGF, TGFBR2, and DNMT3A in Neonatal and Adult Human Dermal Fibroblasts Cell Lines. Curr. Issues Mol. Biol. 2021, 43, 276-285. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010023
Tomela K, Karolak JA, Ginter-Matuszewska B, Kabza M, Gajecka M. Influence of TGFBR2, TGFB3, DNMT1, and DNMT3A Knockdowns on CTGF, TGFBR2, and DNMT3A in Neonatal and Adult Human Dermal Fibroblasts Cell Lines. Current Issues in Molecular Biology. 2021; 43(1):276-285. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010023
Chicago/Turabian StyleTomela, Katarzyna, Justyna A. Karolak, Barbara Ginter-Matuszewska, Michal Kabza, and Marzena Gajecka. 2021. "Influence of TGFBR2, TGFB3, DNMT1, and DNMT3A Knockdowns on CTGF, TGFBR2, and DNMT3A in Neonatal and Adult Human Dermal Fibroblasts Cell Lines" Current Issues in Molecular Biology 43, no. 1: 276-285. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43010023