
Spatial and Temporal Variability of the Floral Scent Emitted by Barlia robertiana (Loisel.) Greuter, a Mediterranean Food-Deceptive Orchid

 , ,
, ,
Abstract
:1. Introduction
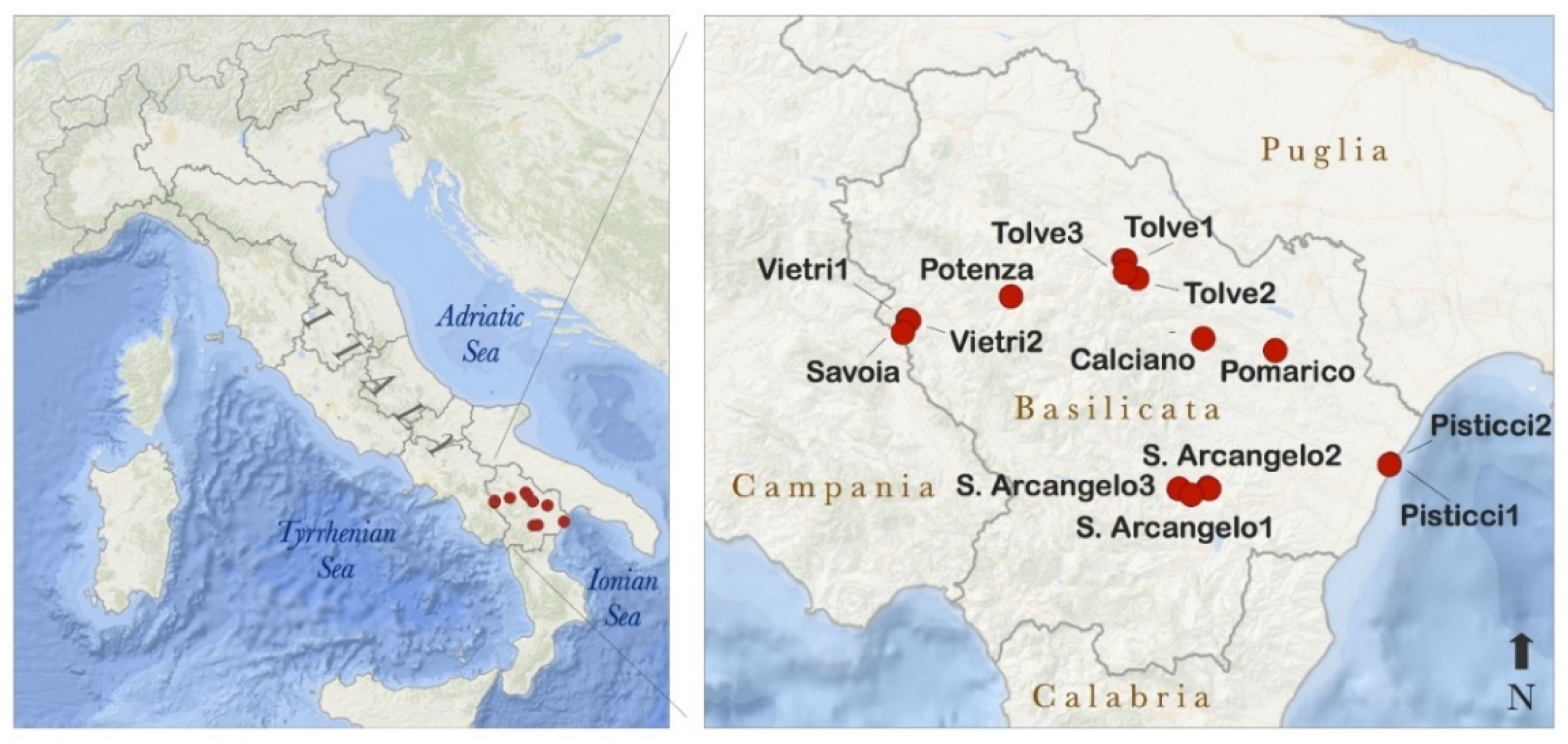
2. Experimental Section
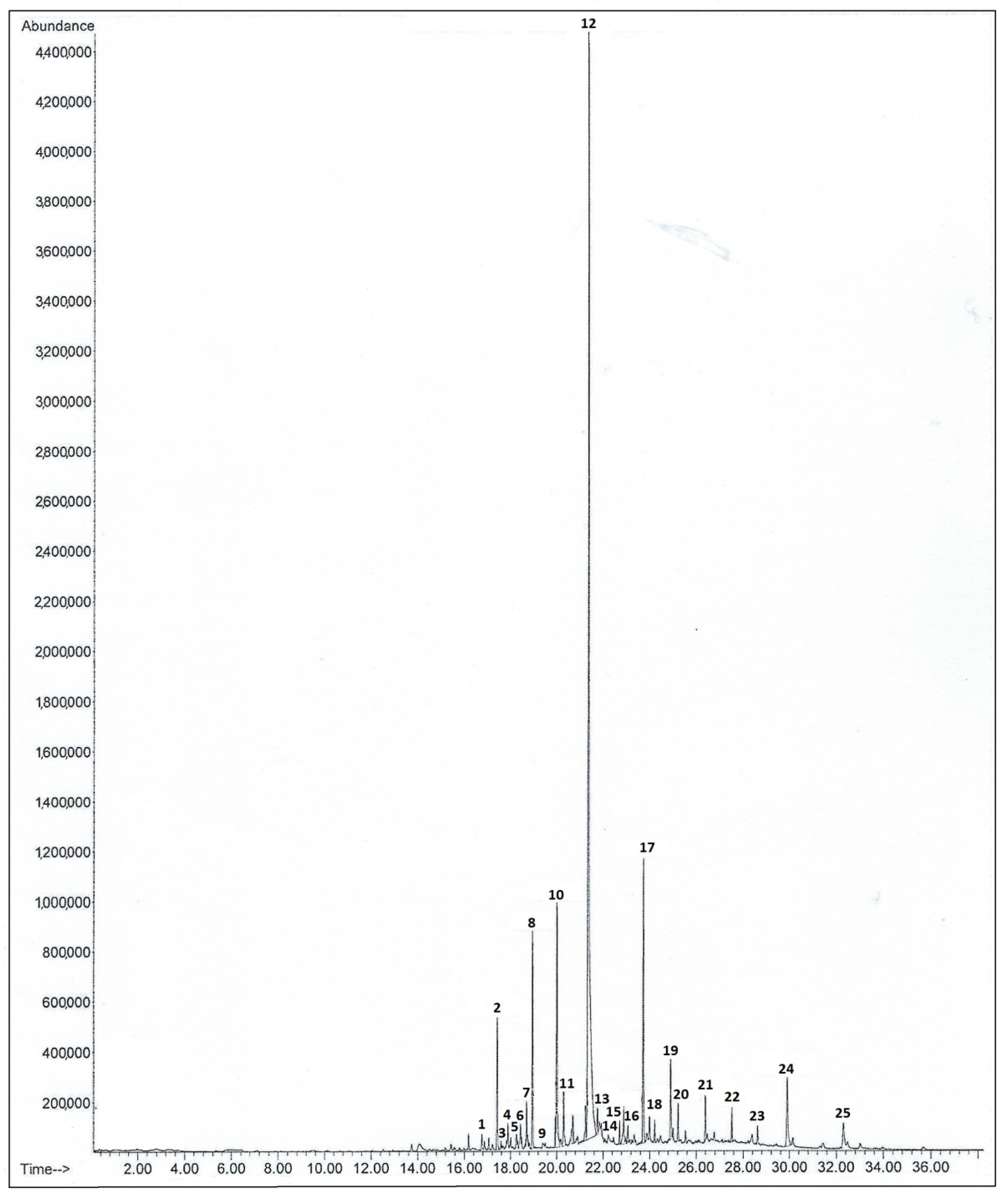
3. Results
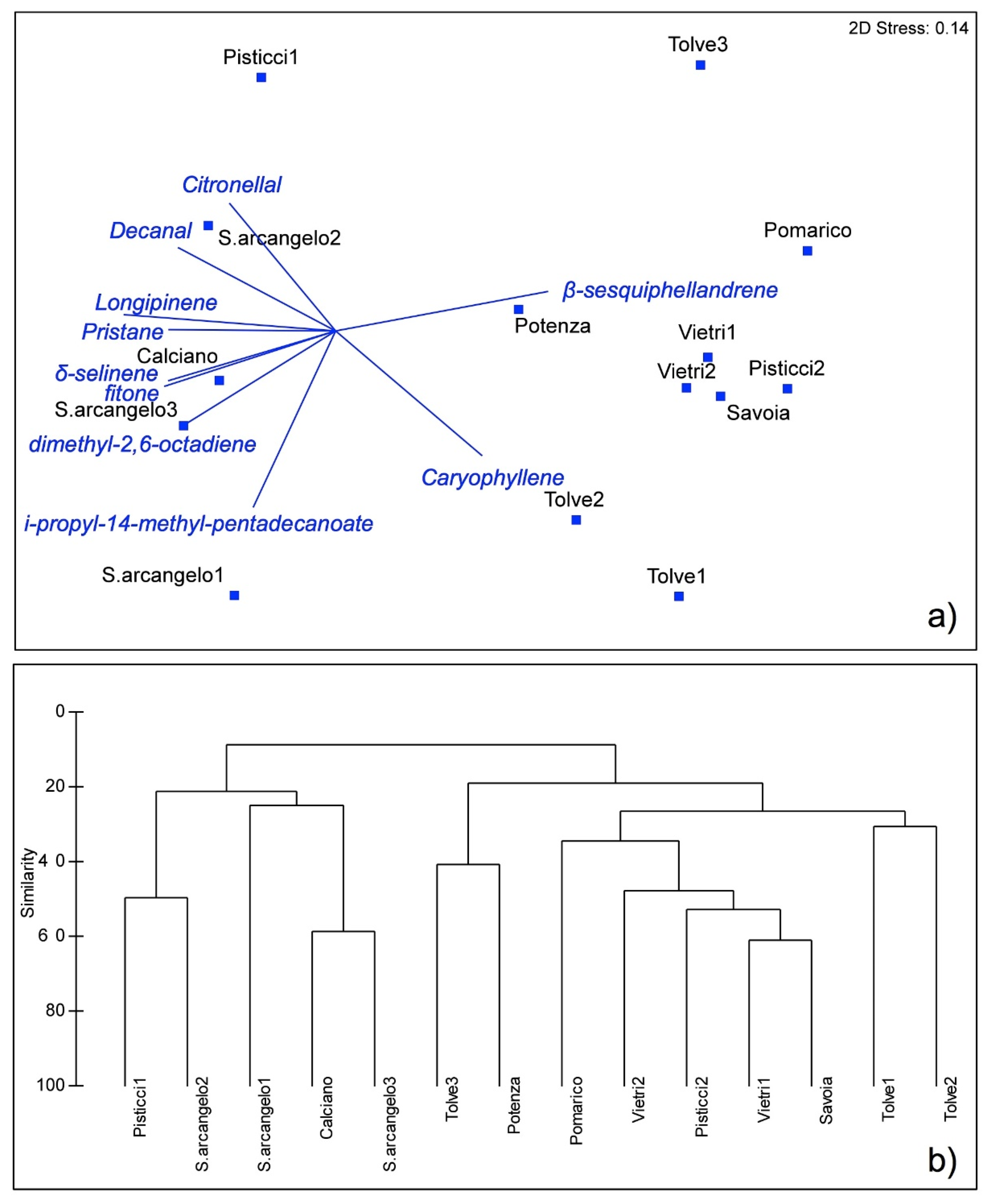
3.1. Spatial Variability
3.2. Temporal Variability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | r.t. [min] | KI | Adsorption Time (h) | ||
|---|---|---|---|---|---|
| 5 | 12 | 24 | |||
| Area [%] ± 0.03 | |||||
| D-limonene | 9.86 | 1022 | 1.35 | 1.04 | 0.93 |
| nonanal | 11.36 | 1103 | 2.87 | ||
| decanal | 13.29 | 1203 | 3.35 | ||
| 4,6,6-trimethylbicyclo[3.1.1]ept-3-en-2-one | 13.46 | 1212 | 6.28 | 2.01 | 1.54 |
| methyl (S)-(-)-citronellate | 14.24 | 1258 | 3.44 | 2.64 | 2.68 |
| α/β-caryophyllene | 17.12 | 1474 | 60.36 | 35.16 | 35.32 |
| 1,3-di-isopropylnaphthalene | 17.13 | 1668 | 1.71 | ||
| trans-α-bergamotene | 17.28 | 1433 | 1.04 | 0.41 | |
| 6,10-dimethyl-5,9-undecadien-2-one | 17.51 | 1453 | 10.92 | ||
| (E),(Z),α/β-3-metylene-7,11-dimethyl-1,6,10-dodecatriene | 17.54 | 1459 | 2.20 | ||
| β-farnesene | 17.60 | 1463 | 25.60 | 20.71 | |
| humulene | 17.66 | 1467 | 1.08 | ||
| pentadecane | 18.17 | 1499 | 3.66 | 5.17 | 3.98 |
| β-bisabolene | 18.42 | 1515 | 7.03 | 7.56 | 5.60 |
| diethyltoluamide | 19.55 | 1571 | 0.97 | ||
| hexadecane | 19.65 | 1600 | 1.30 | 4.71 | |
| (E),(Z)-8-heptadecene | 20.80 | 1676 | 1.42 | ||
| 2,6-diisopropylnaphthalene | 21.23 | 1695 | 1.71 | ||
| heptadecane | 21.75 | 1700 | 2.01 | 0.41 | |
| 2,3-dihydrofarnesyl acetate | 22.60 | 1805 | 2.92 | ||
| isopropyl myristate | 22.78 | 1827 | 5.16 | 25.83 | 7.15 |
| (E),(Z)-5,9,13-trimethyl-4,8,12-tetradecatrienal | 23.03 | 1840 | 1.19 | 1.58 | |
| 7-acetyl-6-ethyl-1,1,4,4-tetramethyltetraline | 23.33 | 1843 | 0.74 | 0.73 | |
| galaxolide | 23.35 | 1850 | 0.74 | 1.46 | |
| 4-octadecyl morfoline | 23.60 | 1880 | 1.80 | 0.97 | |
| methyl hexadecanoate | 24.05 | 1927 | 0.77 | ||
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C. On the Various Contrivances by Which British and Foreign Orchids Are Fertilised by Insects, and on the Good Effects of Intercrossing; Murray, J., Ed.; University of Chicago Press: London, UK, 1862. [Google Scholar]
- Vereecken, N.J.; Dafni, A.; Cozzolino, S. Pollination Syndromes in Mediterranean Orchids—Implications for Speciation, Taxonomy and Conservation. Bot. Rev. 2010, 76, 220–240. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, J.D. Mechanisms and evolution of food-deceptive pollination systems in orchids. Lindleyana 1986, 1, 108–113. [Google Scholar]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219. [Google Scholar] [CrossRef]
- Johnson, S.D.; Schiestl, F.P. Floral Mimicry, 1st ed.; Oxford University Press: Oxford, NY, USA, 2016. [Google Scholar]
- Kunze, J.; Gumbert, A. The combined effect of color and odor on flower choice behavior of bumble bees in flower mimicry systems. Behav. Ecol. 2001, 12, 447–456. [Google Scholar] [CrossRef]
- Galizia, C.G.; Kunze, J.; Gumbert, A.; Borg-Karlson, A.-K.; Sachse, S.; Markl, C.; Menzel, R. Relationship of visual and olfactory signal parameters in a food-deceptive flower mimicry system. Behav. Ecol. 2005, 16, 159–168. [Google Scholar] [CrossRef]
- Johnson, S. Batesian mimicry in the non-rewarding orchid Disa pulchra, and its consequences for pollinator behaviour. Biol. J. Linn. Soc. 2000, 71, 119–132. [Google Scholar] [CrossRef]
- D’Auria, M.; Lorenz, R.; Mecca, M.; Racioppi, R.; Antonio Romano, V. Aroma components of Cephalanthera orchids. Nat. Prod. Res. 2021, 35, 174–177. [Google Scholar] [CrossRef]
- Heinrich, B. Bee flowers: A hypothesis on flower variety and blooming times. Evolution 1975, 29, 325–334. [Google Scholar] [CrossRef]
- Johnson, S.D.; Peter, C.I.; Nilsson, L.A.; Ågren, J. Pollination success in a deceptive orchid is enhanced by co-occuring rewarding magnet plants. Ecology 2003, 84, 2919–2927. [Google Scholar] [CrossRef] [Green Version]
- Dötterl, S.; Vereecken, N.J. The chemical ecology and evolution of bee–flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Delforge, P. Orchidées d’Europe, d’Afrique du Nord et du Proche-Orient: La Bible des Orchidophiles, Plus de 600 Espèces et de Nombreuses Variétés et Illustrées; Guide Delachaux; 4e éd.; Revue et augmentée; Delachaux et Niestlé: Paris, France, 2016; ISBN 978-2-603-02407-2. [Google Scholar]
- Baumann, H.; Künkele, S.; Lorenz, R. Orchideen Europas: Mit angrenzenden Gebieten; Ulmer-Naturführer; Eugen Ulmer KG: Stuttgart, Germany, 2006; ISBN 978-3-8001-4162-3. [Google Scholar]
- Wartmann, B. Orchideen als “Neophyten” in der Schweiz? AGEO Orchis 2020, 1, 9–13. [Google Scholar]
- Vögtlin, J. Himantoglossum robertianum (Loisel.) Delforge am Isteiner Klotz. Berichte Bot. Arbeitsgemeinschaft Südwestdtsch. 2008, 5, 128. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Sramkó, G.; Molnár, A.V.; Hawkins, J.A.; Bateman, R.M. Molecular phylogenetics and evolution of the Eurasiatic orchid genus Himantoglossum sl. Ann. Bot. 2014, 114, 1609–1626. [Google Scholar] [CrossRef]
- Colombo, P.; Giardina, S.; Perrone, A. Studio morfoanatomico di Himantoglossum robertianum ed H. hircinum (Orchidaceae) della Sicilia. J. Eur. Orch. 2009, 41, 359–388. [Google Scholar]
- Bazzicalupo, M.; Burlando, B.; Denaro, M.; Barreca, D.; Trombetta, D.; Smeriglio, A.; Cornara, L. Polyphenol Characterization and Skin-Preserving Properties of Hydroalcoholic Flower Extract from Himantoglossum robertianum (Orchidaceae). Plants 2019, 8, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Emerico, S.; Galasso, I.; Pignone, D.; Scrugli, A. Localization of rDNA loci by Fluorescent In Situ Hybridization in some wild orchids from Italy (Orchidaceae). Caryologia 2001, 54, 31–36. [Google Scholar] [CrossRef]
- Doneddu, M. Osservazioni sull’impollinazione di Barlia robertiana ad opera dei coleotteri Tropinota squalida e Oxythyrea funesta (Cetoniidae) in Sardegna. GIROS Orch. Spont. Eur. 2015, 58, 262–265. [Google Scholar]
- Smithson, A.; Gigord, L.D.B. Are there fitness advantages in being a rewardless orchid? Reward supplementation experiments with Barlia robertiana. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 1435–1441. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Rosa, E.J. Estudi de la Biologia Reproductiva de L’orquídia Gegant (Barlia robertiana) a L’illa de Mallorca. Ph.D. Thesis, Universitat del les Illes Baleares, Palma, Spain, 2014. [Google Scholar]
- Rosati, L.; Romano, V.A.; Cerone, L.; Fascetti, S.; Potenza, G.; Bazzato, E.; Cillo, D.; Mecca, M.; Racioppi, R.; D’Auria, M.; et al. Pollination features and floral volatiles of Gymnospermium scipetarum (Berberidaceae). J. Plant Res. 2019, 132, 49–56. [Google Scholar] [CrossRef]
- Rivas-Martınez, S.; Rivas-Saenz, S.; Penas-Merino, A. Worldwide Bioclimatic classification system. Glob. Geobot. 2011, 1, 1–638. [Google Scholar]
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate map of Sardinia (Italy). J. Maps 2015, 11, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Farris, E.; Filigheddu, R.; Mameli, G.; Falanga, V.; Vanetti, I.; Rosati, L.; Binelli, G. Is population genetic structure of vascular plants shaped more by ecological or geographic factors? A study case on the Mediterranean endemic Centaurea filiformis (Asteraceae). Plant Biol. 2018, 20, 936–947. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, M.; Fascetti, S.; Racioppi, R.; Romano, V.A.; Rosati, L. Orchids from Basilicata: The Scent. In Orchids Phytochemistry, Biology and Horticulture; Merillon, J.-M., Kodja, H., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–22. ISBN 978-3-030-11257-8. [Google Scholar]
- Gallego, E.; Gelabert, A.; Roca, F.J.; Perales, J.F.; Guardino, X. Identification of volatile organic compounds (VOC) emitted from three European orchid species with different pollination strategies: Two deceptive orchids (Himantoglossum robertianum and Ophrys apifera) and a rewarding (Gymnadenia conopsea). J. Biodivers. Environ. Sci. 2012, 2, 18–29. [Google Scholar]
- McBrien, H.L.; Millar, J.G.; Rice, R.E.; McElfresh, J.S.; Cullen, E.; Zalom, F.G. Sex Attractant Pheromone of the Red-Shouldered Stink Bug Thyanta pallidovirens: A Pheromone Blend with Multiple Redundant Components. J. Chem. Ecol. 2002, 28, 1797–1818. [Google Scholar] [CrossRef] [PubMed]
- Bakthavatsalam, N. Semiochemicals. In Ecofriendly Pest Management for Food Security; Academic Press: Cambridge, MA, USA, 2016; pp. 563–611. [Google Scholar]
- Raguso, R.A. Start making scents: The challenge of integrating chemistry into pollination ecology. Entomol. Exp. Appl. 2008, 128, 196–207. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Johnson, S.D. Pollinator-mediated evolution of floral signals. Trends Ecol. Evol. 2013, 28, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Smithson, A.; Macnair, M.R. Negative frequency-dependent selection by pollinators on artificial flowera without rewards. Evolution 1997, 51, 715–723. [Google Scholar] [CrossRef]
- Moya, S.; Ackerman, J.D. Variation in the floral fragrance of Epidendrum ciliare (Orchidaceae). Nord. J. Bot. 1993, 13, 41–47. [Google Scholar] [CrossRef]





| Sample | Pisticci 1 | Pisticci 2 | Calciano | S. Arcangelo 1 | S. Arcangelo 2 | S. Arcangelo 3 | Tolve 1 | Tolve 2 | Tolve 3 | Pomarico | Vietri 1 | Vietri 2 | Potenza | Savoia | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | r.t. [min] | Area [%] ± 0.03 | |||||||||||||
| Ethyl dodecanoate | 19.93 | 0.33 | 0.67 | 1.75 | 0.33 | 0.82 | 1.17 | 8.29 | 3.76 | 2.63 | 1.10 | 2.60 | |||
| β-Bisabolene | 18.43 | 2.13 | 0.96 | 4.06 | 1.05 | 2.13 | 4.61 | 8.98 | 3.17 | 2.00 | 5.32 | ||||
| Hexadecane | 20.16 | 0.14 | 0.53 | 0.96 | 0.88 | 3.09 | 0.60 | 4.01 | 1.80 | 1.01 | 3.36 | ||||
| δ-Selinene | 18.65 | 0.98 | 7.04 | 9.06 | 17.59 | 6.49 | 1.65 | 2.49 | 1.16 | 1.54 | |||||
| β-Sesquiphellandrene | 19.35 | 29.06 | 0.91 | 2.94 | 8.39 | 17.59 | 43.08 | 15.14 | 8.97 | 25.05 | |||||
| Caryophyllene | 17.46 | 8.51 | 24.63 | 10.11 | 3.68 | 3.07 | 5.21 | 6.75 | 17.96 | ||||||
| cis-α-Bergamotene | 17.58 | 0.42 | 0.53 | 1.73 | 0.77 | 0.77 | 14.04 | 4.15 | 1.14 | ||||||
| Heptadecane | 21.29 | 0.74 | 7.41 | 1.43 | 0.73 | 2.94 | 2.24 | 1.49 | 2.11 | ||||||
| α-Pinene | 7.79 | 0.49 | 3.02 | 0.46 | 0.48 | 0.55 | 0.92 | 7.87 | |||||||
| D-limonene | 9.80 | 1.34 | 0.34 | 0.10 | 1.65 | 1.49 | 0.46 | 3.21 | 0.11 | ||||||
| Citronellol | 14.36 | 0.57 | 1.34 | 2.53 | 0.5 | 1.77 | 17.96 | 1.53 | |||||||
| Methyl citronellate | 15.03 | 0.30 | 0.75 | 0.9 | 0.31 | 3.01 | 3.61 | 1.85 | |||||||
| Octadecane | 22.85 | 0.23 | 1.30 | 0.21 | 1.24 | 0.66 | 0.31 | 1.25 | |||||||
| Verbenone | 13.74 | 45.22 | 31.48 | 1.12 | 3.81 | 0.57 | 1.32 | ||||||||
| Tetradecane | 17.02 | 0.53 | 0.96 | 1.73 | 3.09 | 1.39 | 2.44 | ||||||||
| α-Zingiberene | 17.09 | 6.88 | 0.98 | 2.63 | 17.14 | 0.94 | 3.59 | ||||||||
| Z-β-Farnesene | 17.54 | 0.22 | 0.42 | 0.40 | 1.49 | 0.41 | 0.49 | ||||||||
| Pristane | 21.68 | 35.57 | 9.61 | 58.90 | 1.56 | 1.13 | 22.57 | ||||||||
| i-propyl 14-methyl-pentadecanoate | 26.35 | 3.74 | 1.44 | 1.44 | 12.26 | 1.54 | 0.57 | ||||||||
| Longipinene | 18.40 | 0.72 | 3.86 | 1.77 | 4.40 | 1.32 | |||||||||
| Ethyl tetradecanoate | 22.58 | 15.96 | 12.40 | 2.57 | 1.00 | 0.92 | |||||||||
| Dihydrofarnesol | 22.7 | 0.09 | 0.52 | 1.01 | 0.82 | 22.23 | |||||||||
| Nonadecane | 24.22 | 0.39 | 2.06 | 0.65 | 0.90 | 0.24 | |||||||||
| Methyl hexadecanoate | 25.08 | 0.22 | 1.15 | 0.57 | 0.78 | 0.31 | |||||||||
| β-Myrcene | 8.99 | 0.36 | 0.37 | 0.23 | 3.26 | ||||||||||
| Tridecane | 15.12 | 1.25 | 2.35 | 1.29 | 1.63 | ||||||||||
| E-β-Farnesene | 17.96 | 0.14 | 0.87 | 1.97 | 7.10 | ||||||||||
| Humulene | 18.08 | 0.11 | 0.46 | 0.44 | 0.22 | ||||||||||
| 2,6-Bis(1,1-dimethylethyl)-2,5-cyclohexadiene-1,4-dione | 18.11 | 0.19 | 0.64 | 0.37 | 0.63 | ||||||||||
| β-Curcumene | 18.13 | 0.45 | 1.53 | 2.43 | 1.56 | ||||||||||
| α-Farnesene | 18.60 | 0.25 | 0.47 | 16.97 | 0.89 | ||||||||||
| Nerolidol | 19.73 | 0.33 | 1.69 | 0.57 | 0.86 | ||||||||||
| Fitone | 23.98 | 0.16 | 0.28 | 1.32 | 1.12 | ||||||||||
| β-Pinene | 8.72 | 0.18 | 1.13 | 0.39 | |||||||||||
| p-Menth-8-en-1-ol | 12.26 | 1.35 | 0.83 | 21.68 | |||||||||||
| Decanal | 13.72 | 0.88 | 2.12 | 0.25 | |||||||||||
| 2,6-dimethyl-2,6-Octadiene | 16.72 | 0.42 | 1.16 | 0.32 | |||||||||||
| Pentadecane | 18.49 | 0.12 | 0.45 | 0.79 | |||||||||||
| Farnesol | 21.85 | 1.70 | 36.63 | 19.36 | |||||||||||
| Tetradecanoic acid | 22.5 | 0.30 | 0.73 | 0.16 | |||||||||||
| Eicosane | 25.39 | 1.36 | 0.61 | 0.64 | |||||||||||
| cis-p-menthan-1-ol | 10.93 | 0.46 | 0.77 | ||||||||||||
| Linalool | 11.43 | 0.46 | 0.24 | ||||||||||||
| Citronellal | 13.07 | 0.31 | 0.42 | ||||||||||||
| α-Terpineol | 13.16 | 0.54 | 5.39 | ||||||||||||
| Citronellyl formate | 14.52 | 2.42 | 0.33 | ||||||||||||
| Bornyl acetate | 14.98 | 1.71 | 1.00 | ||||||||||||
| Geranyl acetone | 17.52 | 0.21 | 0.37 | ||||||||||||
| α-Curcumene | 18.28 | 0.63 | 0.23 | ||||||||||||
| Tetradecanal | 21.23 | 0.42 | 2.24 | ||||||||||||
| Farnesal | 22.04 | 5.81 | 0.49 | ||||||||||||
| 2,6-diisopropylnaphthalene | 22.22 | 0.48 | 1.00 | ||||||||||||
| 2,3-Dihydrofarnesyl acetate | 22.76 | 0.5 | 0.86 | ||||||||||||
| Pentadecanal | 23.02 | 0.21 | 0.27 | ||||||||||||
| 4-methyl-3-penten-2-one | 5.33 | 0.19 | |||||||||||||
| β-Phellandrene | 9.65 | 4.51 | |||||||||||||
| γ-Terpinene | 10.66 | 5.24 | |||||||||||||
| Geraniol | 14.04 | 0.53 | |||||||||||||
| D-carvone | 14.49 | 0.35 | |||||||||||||
| Citral | 14.74 | 1.1 | |||||||||||||
| Citronellyl acid | 15.56 | 1.53 | |||||||||||||
| Citronellyl propionate | 16.1 | 1.02 | |||||||||||||
| Phytan | 22.98 | 0.33 | |||||||||||||
| Hexahydrofarnesyl acetone | 23.21 | 0.29 | |||||||||||||
| Farnesyl acetate | 23.26 | 0.46 | |||||||||||||
| Isopropyl myristate | 23.35 | 0.47 | |||||||||||||
| Methyl 9-octadecenoate | 26.4 | 0.28 | |||||||||||||
| Heneicosane | 27.05 | 0.37 | |||||||||||||
| Isopropyl linoleate | 27.43 | 0.09 | |||||||||||||
| Number of compounds | 28 | 20 | 24 | 17 | 21 | 17 | 19 | 20 | 21 | 16 | 14 | 21 | 19 | 14 | |
| Flowering Phase | Sample 1 | Sample 2 | Sample 1 | Sample 2 | |
|---|---|---|---|---|---|
| Early | Late | ||||
| Compound | r.t. [min] | Area % | |||
| α-Pinene | 7.83 | 0.24 | 0.61 | ||
| β-Myrcene | 9.03 | 0.29 | 0.52 | ||
| D-Limonene | 9.84 | 0.54 | 1.00 | 0.66 | 2.01 |
| Sylvestrene | 11.25 | 0.54 | |||
| Citronellal | 12.29 | 1.50 | 0.61 | ||
| α-Terpineol | 13.17 | 0.78 | |||
| Verbenone | 13.45 | 0.70 | 0.81 | 0.54 | 0.23 |
| Carvone | 14.07 | 0.42 | 0.35 | ||
| Citronellol | 14.14 | 42.54 | 15.66 | ||
| Methyl (S)-citronellate | 14.24 | 1.28 | 1.06 | 1.23 | |
| 4-Carene | 15.84 | 2.63 | |||
| α-Terpineol acetate | 15.85 | 13.83 | |||
| (E,Z)-2,6-Dimethyl-2,6-octadiene | 15.87 | 0.38 | |||
| Dihydro-b-ionone | 16.20 | 0.42 | |||
| 2-Methylene-4,8,8-trimethyl-4-vinylbicyclo [5.2.0]nonane | 16.70 | 2.52 | |||
| Caryophyllene | 17.12 | 34.56 | 3.34 | 32.35 | |
| α-Bergamotene | 17.29 | 1.10 | 0.50 | 2.74 | |
| (Z)-β-Farnesene | 17.47 | 1.50 | 0.57 | 1.99 | |
| trans-Geranylacetone | 17.50 | 0.73 | |||
| (E)-β-Farnesene | 17.60 | 5.29 | |||
| 4-Methyltetradecane | 17.65 | 30.68 | |||
| α-longipinene | 17.97 | 0.36 | |||
| Pentadecane | 18.17 | 4.27 | 3.78 | ||
| δ-Selinene | 18.30 | 0.32 | |||
| β-Bisabolene | 18.42 | 4.85 | 3.82 | 6.53 | 6.98 |
| β-Sesquiphellandrene | 19.35 | 24.95 | 44.13 | ||
| Elemene | 19.54 | 1.14 | |||
| Hexadecane | 19.65 | 0.98 | 0.37 | ||
| 8-Heptadecene | 20.80 | 1.47 | 1.60 | ||
| 2,3-Dihydrofarnesol | 21.08 | 34.80 | 11.28 | 17.15 | 1.02 |
| 2,6-Diisopropylnaphthalene | 21.23 | 0.98 | |||
| Octadecane | 22.41 | 0.98 | 1.12 | ||
| (E)-5-Octadecene | 22.55 | 1.47 | |||
| Isopropyl miristate | 22.78 | 0.89 | 0.90 | 0.83 | 0.23 |
| Phytone | 22.90 | 1.09 | |||
| 4-Octadecylmorfoline | 23.60 | 1.40 | |||
| Nonadecane | 23.70 | 0.37 | |||
| Methyl hexadecanoate | 24.05 | 0.42 | |||
| (E,Z)-5,9,13-Trimethyl-4,8,12-tetradecatrienal | 24.32 | 1.61 | |||
| trans-Geranylgeraniol | 26.50 | 1.61 | |||
| Number of compounds | 22 | 18 | 15 | 18 | |
| Calciano | Calciano | Potenza | Potenza | S. Arcangelo 2 | S. Arcangelo 2 | ||
|---|---|---|---|---|---|---|---|
| Year of Sampling | 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | |
| Compound | r.t. [min] | Area (%) | |||||
| α-Pinene | 7.83 | 3.02 | 10.63 | 6.43 | 7.87 | 0.46 | 0.24 |
| β-Pinene | 8.74 | 0.42 | 0.35 | 0.39 | |||
| Sulcatone | 8.91 | 0.19 | |||||
| β-Myrcene | 9.03 | 0.65 | 0.33 | 0.37 | 0.29 | ||
| β-Phellandrene | 9.65 | 0.06 | |||||
| 2-Ethyl-1-hexanol | 9.83 | 0.18 | |||||
| D-limonene | 9.84 | 0.10 | 1.31 | 0.95 | 0.11 | 1.65 | 1.00 |
| Eucaliptol | 9.88 | 0.35 | 0.35 | ||||
| α-Terpinolene | 11.05 | 32.45 | |||||
| trans-Sabinene idrate | 11.13 | 1.46 | |||||
| 6-Methyl-2-pyridinecarboxyaldehyde | 11.19 | 0.36 | |||||
| Undecane | 11.3 | 1.03 | |||||
| Linalool | 11.43 | 0.24 | |||||
| Nonanal | 11.45 | 1.35 | |||||
| Rose oxide | 11.64 | 0.59 | |||||
| 2-Phenylethanol | 11.67 | 0.42 | |||||
| p-Menth-8-en-1-ol | 12.26 | 1.66 | 0.83 | ||||
| Citronellal | 12.29 | 1,12 | 0.4 | 0.42 | 1.50 | ||
| α-Terpineol | 13.17 | 0.45 | 0.17 | 0.78 | |||
| 1-Butoxy-2-ethylhexane | 13.23 | 0.18 | |||||
| Verbenone | 13.45 | 4.72 | 55.18 | 31.48 | 0.81 | ||
| Decanal | 13.72 | 2.12 | |||||
| Carvone | 14.07 | 0.42 | |||||
| Citronellol | 14.14 | 1.53 | 1.34 | 42.54 | |||
| cis-Octahydro-3a-methyl-2H-inden-2-one | 14.23 | 1.80 | |||||
| Methyl (S)-citronellate | 14.24 | 1.80 | 1.06 | ||||
| D-carvone | 14.49 | 7.92 | |||||
| Citronellyl formate | 14.52 | 0.33 | |||||
| (E)-cinnamaldehyde | 14.59 | 8.25 | |||||
| Methyl citronellate | 15.03 | 0.30 | |||||
| Tridecane | 15.12 | 1.55 | |||||
| Citronellyl acid | 15.56 | 0.72 | |||||
| Cinnamyl alcohol | 15.80 | 0.15 | |||||
| 4-Carene | 15.84 | 1.53 | |||||
| (E,Z)-2,6-Dimethyl-2,6-octadiene | 15.87 | 0.38 | |||||
| 2,6-dimethyl-2,6-octadiene | 16.72 | 0.42 | |||||
| Tetradecane | 17.02 | 0.96 | |||||
| α-zingiberene | 17.09 | 2.63 | 0.97 | 0.94 | |||
| Caryophyllene | 17.12 | 1.74 | 0.32 | 6.75 | 3.34 | ||
| α-Bergamotene | 17.29 | 0.64 | 0.50 | ||||
| (Z)-β-Farnesene | 17.47 | 1.77 | 0.49 | 0.42 | 1.50 | ||
| Trans-Geranyl acetone | 17.50 | 0.37 | |||||
| Geranyl acetone | 17.52 | 0.21 | 0.09 | 0.37 | |||
| cis-α-bergamotene | 17.58 | 1.73 | 0.67 | 0.77 | |||
| (E)-β-Farnesene | 17.60 | 14.22 | 5.29 | ||||
| 2,6-bis(1,1-dimethylethyl)-2,5-cyclohexadiene-1,4-dione | 18.11 | 0.19 | 0.63 | 0.64 | |||
| β-curcumene | 18.13 | 1.53 | 0.31 | 1.56 | |||
| Longipinene | 18.40 | 3.86 | 0.34 | 4.40 | |||
| β-Bisabolene | 18.42 | 5.22 | 2.00 | 3.82 | |||
| Pentadecane | 18.49 | 0.79 | |||||
| δ-selinene | 18.65 | 7.04 | 1.37 | 17.59 | |||
| β-sesquiphellandrene | 19.35 | 0.91 | 1.98 | 8.97 | 24.95 | ||
| Hexadecane | 19.65 | 0.35 | 1.01 | ||||
| Nerolidol | 19.73 | 0.33 | 0.57 | ||||
| Ethyl dodecanoate | 19.93 | 0.67 | 1.10 | 0.33 | |||
| 2,3-Dihydrofarnesol | 21.08 | 11.28 | |||||
| Heptadecane | 21.29 | 7.41 | |||||
| Pristane | 21.68 | 35.57 | 22.57 | 9.61 | |||
| Farnesol | 21.85 | 19.36 | |||||
| Farnesal | 22.04 | 0.54 | |||||
| Tetradecanoic acid | 22.5 | 0.30 | |||||
| Ethyl tetradecanoate | 22.58 | 12.40 | 1.23 | ||||
| Dihydrofarnesol | 22.7 | 0.52 | |||||
| 2,3-Dihydrofarnesyl acetate | 22.76 | 0.86 | |||||
| Isopropyl miristate | 22.78 | 0.90 | |||||
| Octadecane | 22.85 | 0.31 | |||||
| pentadecanal | 23.02 | 0.27 | |||||
| Hexahydrofarnesyl acetone | 23.21 | 0.29 | |||||
| Fitone | 23.98 | 0.28 | |||||
| methyl hexadecanoate | 25.08 | 0.22 | |||||
| Eicosane | 25.39 | 1.36 | |||||
| i-propyl 14-methyl-pentadecanoate | 26.35 | 0.57 | |||||
| methyl 9-octadecenoate | 26.4 | 0.28 | |||||
| Number of compounds | 24 | 22 | 31 | 19 | 21 | 18 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, V.A.; Rosati, L.; Fascetti, S.; Cittadini, A.M.R.; Racioppi, R.; Lorenz, R.; D’Auria, M. Spatial and Temporal Variability of the Floral Scent Emitted by Barlia robertiana (Loisel.) Greuter, a Mediterranean Food-Deceptive Orchid. Compounds 2022, 2, 37-53. https://0-doi-org.brum.beds.ac.uk/10.3390/compounds2010004
Romano VA, Rosati L, Fascetti S, Cittadini AMR, Racioppi R, Lorenz R, D’Auria M. Spatial and Temporal Variability of the Floral Scent Emitted by Barlia robertiana (Loisel.) Greuter, a Mediterranean Food-Deceptive Orchid. Compounds. 2022; 2(1):37-53. https://0-doi-org.brum.beds.ac.uk/10.3390/compounds2010004
Chicago/Turabian StyleRomano, Vito Antonio, Leonardo Rosati, Simonetta Fascetti, Anna Maria Roberta Cittadini, Rocco Racioppi, Richard Lorenz, and Maurizio D’Auria. 2022. "Spatial and Temporal Variability of the Floral Scent Emitted by Barlia robertiana (Loisel.) Greuter, a Mediterranean Food-Deceptive Orchid" Compounds 2, no. 1: 37-53. https://0-doi-org.brum.beds.ac.uk/10.3390/compounds2010004