

Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts

1
Division of Animal Science, College of Agriculture, Tamagawa University, 6-1-1 Tamagawa-gakuen, Tokyo 194-9610, Japan
2
Biosystems & Biofunctions Research Center, Tamagawa University Research Institute, 6-1-1 Tamagawa-gakuen, Tokyo 194-9610, Japan
3
MAC Gifu Research Institute, MicroAlgae Corporation, 4-15 Akebono-cho, Gifu 500-8148, Japan
*
Author to whom correspondence should be addressed.
Cosmetics 2023, 10(1), 30; https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10010030
Submission received: 11 January 2023
/
Revised: 1 February 2023
/
Accepted: 7 February 2023
/
Published: 9 February 2023
(This article belongs to the Special Issue Cosmetics in the Age of Green Technologies)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Nostoc verrucosum, an edible microalgal species, forms colonies in streams. Here, we investigated the antimelanogenic and anti-oxidative effects of N. verrucosum extracts. We collected N. verrucosum from Toyama Prefecture, Japan, and successfully cultured it in indoor cultivation systems. Aqueous, methanol, and hexane extracts of N. verrucosum were prepared for various experiments. To elucidate the antimelanogenic effects of N. verrucosum, we performed tyrosinase assay, melanin content assay, western blotting, and real-time quantitative reverse-transcription polymerase chain reaction (qRT-PCR). Anti-oxidative effects were evaluated using the 2,2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid (ABTS) assay. The methanol and hexane extracts significantly inhibited melanin synthesis in B16F1 melanoma cells. Western blotting showed that 12.5 and 25.0 µg/mL N. verrucosum hexane extract suppressed tyrosinase activity. The qRT-PCR analysis revealed that N. verrucosum hexane extract inhibited α-melanocyte stimulating hormone-enhanced tyrosinase, tyrosinase-related protein (TRP)-1, TRP-2, and melanocortin 1 receptor (MC1R) mRNA expression. On the contrary, N. verrucosum hexane extract did not alter microphthalmia-associated transcription factor (Mitf) transcription. The ABTS assay showed that all extracts had radical scavenging activities, and the IC50 values of the aqueous and methanol extracts were 294.6 and 172.8 µg/mL, respectively. Our findings demonstrate that N. verrucosum is a suitable candidate for the development of antimelanogenic agents, cosmetics, or functional food ingredients.
Keywords:
Nostoc verrucosum (ashitsuki); microalga; melanin synthesis; melanoma; tyrosinase; antioxidant; MC1R; Mitf1. Introduction
Nostoc verrucosum, “ashitsuki” in Japanese, is a cyanobacterium that forms macroscopic colonies comprising trichomes of cells and an extracellular matrix in shallow streams [1]. The eighth century Japanese waka poetry anthology “Man-yo-shu” describes that N. verrucosum was consumed as an edible alga in ancient Japan. Nostoc commune is also consumed as food in Indonesia, China, Peru, and Japan [2]. Although N. commune is highly tolerant to frost, high temperatures, and extreme desiccation, its colonies always occur in streams [3]. Oku et al. reported that the ethanolic extract of N. verrucosum inhibited the growth of bacteria owing to the presence of n-1 fatty acids, demonstrating its antibacterial activity [1]. Korteerakul et al. reported that N. commune aqueous solution has anti-oxidative and antiglycative properties [4].
Exposure to solar ultraviolet radiation results in skin pigmentation. The overproduction of melanin in the skin leads to hyperpigmentation disorders, such as freckles, melasma, and solar lentigo, as well as melanoma [5]. Melanin biosynthesis involves three key enzymes that catalyze several reactions. For example, tyrosinase catalyzes two different reactions: the hydroxylation of tyrosine to 3,4-dihydroxyphenylalanine (DOPA) and the oxidation of DOPA to DOPAquinone [6]. The tyrosinase-related proteins TRP-1 and TRP-2 produce other types of eumelanin [7,8].
Melanin production is triggered by an elevation in the intracellular cAMP level initiated by α-melanocyte-stimulating hormone (α-MSH), isobutylmethylxanthine (IBMX), forskolin, and adrenocorticotropic hormone. The increased cAMP levels lead to the activation of protein kinase A and cAMP-response element binding protein (CREB). CREB binds to the promoter of the microphthalmia-associated transcription factor (Mitf) gene, and Mitf activates tyrosinase (TRP-1 and TRP-2) gene transcription [9].
In our previous study, we found that the aqueous extract of the coccolithophore Pleurochrysis carterae inhibits melanin synthesis by downregulating Mitf expression [10]. However, there is a lack of information on the application prospects of microalgae in cosmetic, food additive, and fine chemical industries. Moreover, there are only a few reports on the biological activities of N. verrucosum. In this study, wild colonies of N. verrucosum were collected and used to prepare aqueous, methanol, and hexane extracts (hereinafter referred to as AE, ME, and HE, respectively). Subsequently, the effect of N. verrucosum extracts on melanin synthesis was investigated.
2. Materials and Methods
2.1. Preparation of N. verrucosum Extracts
2.1.1. Collection of N. verrucosum Colonies
We collected N. verrucosum from Nanto, Toyama Prefecture (Japan), and named the strain MAC1606TGR (Figure 1). After isolation, N. verrucosum was incubated in a KNO3-free BG11 medium at room temperature under a fluorescent light for 10 d (Figure 2). Thereafter, the cells were collected and freeze-dried.
2.1.2. Preparation of N. verrucosum HE
To prepare the HE, N. verrucosum dried powder (10 g) was dissolved in 2 L of hexane, stirred for 1 h at room temperature, and passed through GA100 Glass Fiber Filters (Advantec, Tokyo, Japan). The filtrate was then centrifuged (10,000 rpm, 10 min) and dried in a vacuum; the resulting vacuum-dried product was used as HE.
2.1.3. Preparation of N. verrucosum ME
To prepare N. verrucosum ME, the hexane extraction residue was added to 2 L of methanol and stirred for 1 h at room temperature. The mixture was then centrifuged (10,000 rpm, 10 min), filtered, and dried in a vacuum.
2.1.4. Preparation of N. verrucosum AE
To prepare AE, the methanol extraction residue was added to 2 L of water and stirred for 1 h at 90–95 °C. The AE of N. verrucosum was then filtered and freeze-dried.
The HE and ME were dissolved in dimethyl sulfoxide, whereas the aqueous extract was dissolved in distilled water; the three extracts were then stored at −20 °C until use.
2.2. Cell Culture
B16F1 melanoma cells (Riken BioResource Center, Ibaraki, Japan) were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Sigma, St. Louis, MO, USA) supplemented with 5% fetal bovine serum and penicillin/streptomycin (Life Technologies, Carlsbad, CA, USA) at 37 °C in a humidified atmosphere containing 5% CO2.
2.3. Assay of 2,2’-Azino-bis (3-ethylbenzothiazoline-6-sulfonic Acid (ABTS) Radical Scavenging Activity
The assay of ABTS radical scavenging activity was performed as described by Re et al. [11]. To determine the radical scavenging activity, 180 µL of ABTS radical solution was mixed with 20 µL of N. verrucosum extracts and incubated at room temperature in the dark for 30 min. The absorbance of the samples was measured at 732 nm, and ABTS radical scavenging activity was calculated as follows:
ABTS radical scavenging activity (%) = (1 − Asample/Acontrol) × 100.
2.4. Assay of Tyrosinase Activity
To determine the effects of N. verrucosum extracts on tyrosinase activity, we performed a cell-free assay using mushroom tyrosinase as reported previously [12].
2.5. Assay of Cell Proliferation
To evaluate the effects of N. verrucosum extracts on cell proliferation, we performed the trypan blue exclusion test. In this test, the cells were treated with N. verrucosum extracts in the presence of α-MSH (10 nM) for 3 d. After treatment, the cells were rinsed once with phosphate-buffered saline (PBS) and resuspended in trypsin/EDTA (Life Technologies, Carlsbad, CA). The number of viable cells was determined using a Fuchs-Rosenthal cytometer under a light microscope.
2.6. Determination of Melanin Content
Melanin content was measured as described previously [13]. B16F1 melanoma cells were treated with N. verrucosum extracts for 3 d in the presence of α-MSH (10 nM). After treatment, the cells were collected and lysed by boiling in 2 M NaOH. Melanin content was spectrophotometrically determined at an absorbance wavelength of 405 nm.
2.7. DOPA Staining
DOPA staining was performed as described previously [13]. B16F1 melanoma cells were treated with N. verrucosum extracts for 3 d in the presence of α-MSH (10 nM). After treatment, the cells were collected and lysed in 0.1 M sodium phosphate buffer (pH 6.8) containing 1% Triton X-100, 1 mM phenylmethylsulfonyl fluoride, 10 µg/mL aprotinin, and 10 µg leupeptin, without mercaptoethanol, or by heating. Following electrophoresis, SDS-PAGE gels were stained with 0.5 mg/mL L-DOPA (Sigma) solution for 2 h at 37 °C. Gel images were obtained using WSE-6100 LuminoGraph 1 (ATTO Corporation, Tokyo, Japan).
2.8. Western Blotting
To elucidate the effect of N. verrucosum extracts on tyrosinase and Mitf levels, we performed western blotting analysis. Nostoc verrucosum HE-treated cells were lysed using RIPA lysis buffer, and whole-cell lysates were separated via SDS-PAGE; the resolved proteins were then transferred onto PVDF membranes (Merck Millipore, Burlington, MA, USA). After blocking with 5% skimmed milk in PBS containing 0.1% Tween 20, the membranes were probed overnight with tyrosinase and Mitf antibodies (Santa Cruz Biotechnology, TX, USA) at 4 °C and further incubated with horseradish peroxidase-conjugated secondary antibodies. The bound antibodies were detected via chemiluminescence using an ImmunoStar zeta (Fujifilm Wako Pure Chemical, Osaka, Japan) following the manufacturer’s instructions.
2.9. Quantitative Reverse-Transcription Polymerase Chain Reaction
To investigate the effects of N. verrucosum extracts on melanin synthesis-related gene expression, we performed real-time quantitative reverse-transcription polymerase chain reaction (qRT-PCR). B16F1 melanoma cells were treated with N. verrucosum HE in DMSO for the indicated period. After treatment, qRT-PCR was performed as described in our previous report [14].
2.10. Statistical Analysis
The data were analyzed with GraphPad Prism 9 software (GraphPad Software, Inc., La Jolla, CA). All numerical data are presented as the mean ± SEM of experiments done in at least triplicate. Results with p < 0.05 and p <0.01 were considered statistically significant.
3. Results
3.1. Antioxidant Activity
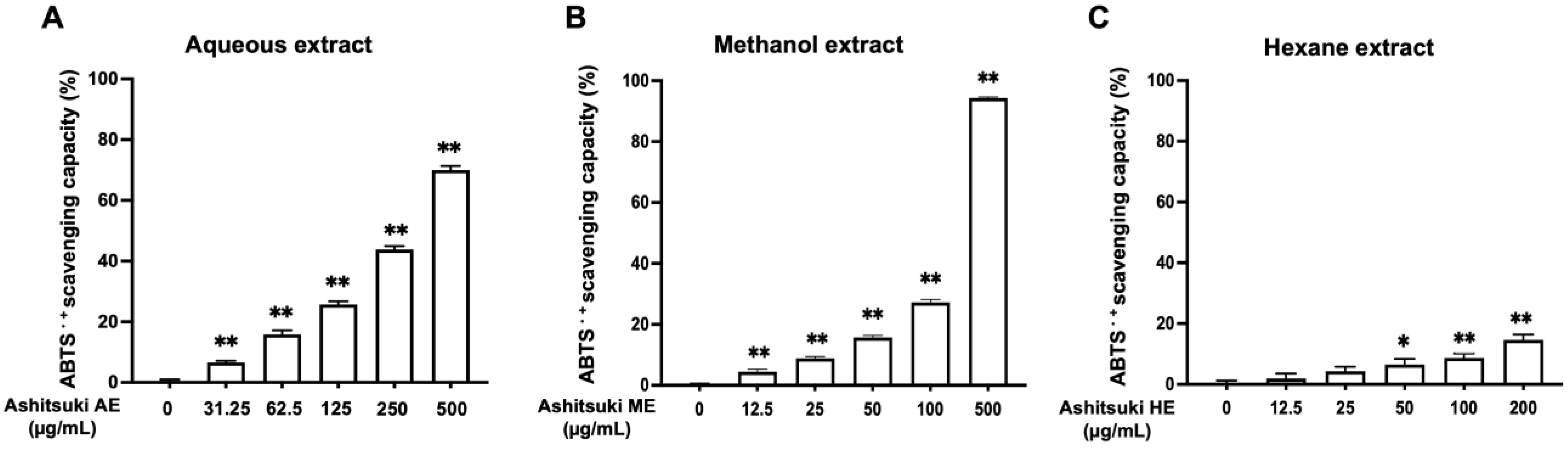
First, we evaluated the anti-oxidant activity of N. verrucosum extracts using the assay of ABTS radical scavenging activity. All extracts exhibited anti-oxidant activity, particularly the AE and ME (Figure 3). The HE did not show radical scavenging activity up to 25.0 µg/mL. The IC50 values of the AE and ME were 294.6 and 172.8 µg/mL, respectively.
3.2. Mushroom Tyrosinase Activity
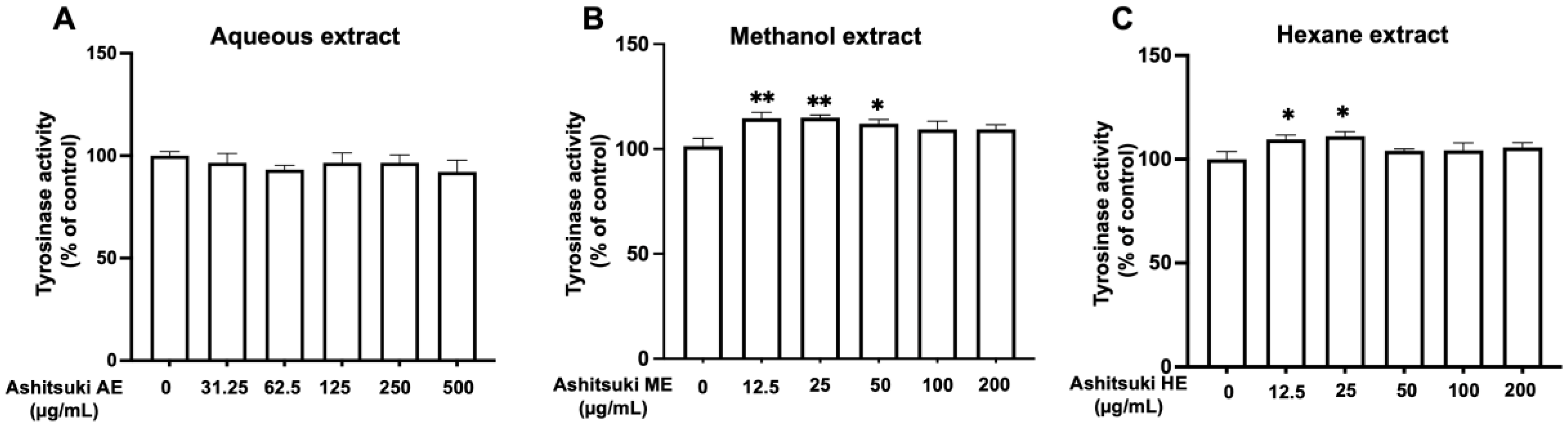
To clarify the mechanism underlying the antimelanogenic effect of N. verrucosum extracts, we performed a tyrosinase activity assay using mushroom tyrosinase. Nostoc verrucosum extracts did not inhibit mushroom tyrosinase activity (Figure 4). Unexpectedly, the ME and AE slightly enhanced mushroom tyrosinase activity at 12.5 and 25.0 µg/mL.
3.3. Cell Proliferation
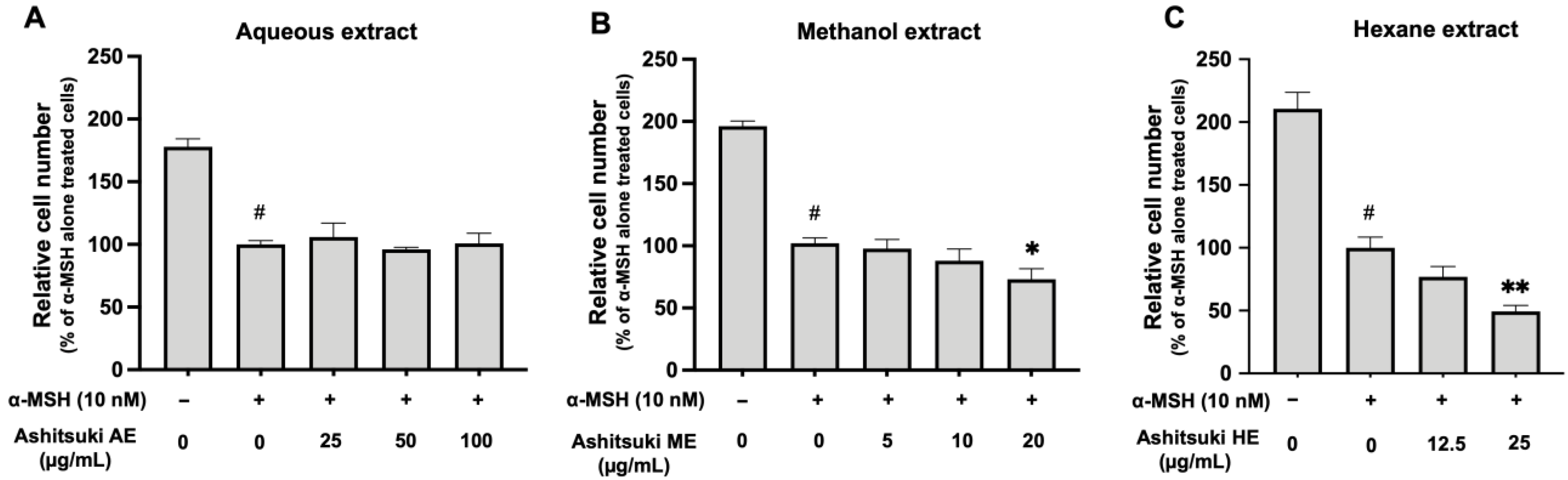
The effects of N. verrucosum extracts on cell proliferation were assessed (Figure 5), and the number of α-MSH-treated cells was lower than that in the non-stimulated control. cAMP inducers, such as α-MSH and IBMX, inhibit cell proliferation [14]. Cell proliferation was not affected by N. verrucosum AE (Figure 5A). The ME inhibited cell proliferation at 20 µg/mL (73% of α-MSH treated control) (Figure 5B). The HE also inhibited cell proliferation at 25 µg/mL (49.4% of α-MSH treated control) (Figure 5C). These findings revealed that the lipophilic compounds in N. verrucosum might inhibit the growth of malignant melanoma cells. Additionally, none of the extracts induced cell death in the concentration range used in this study (data not shown).
3.4. Melanin Production and Intracellular Tyrosinase Activity
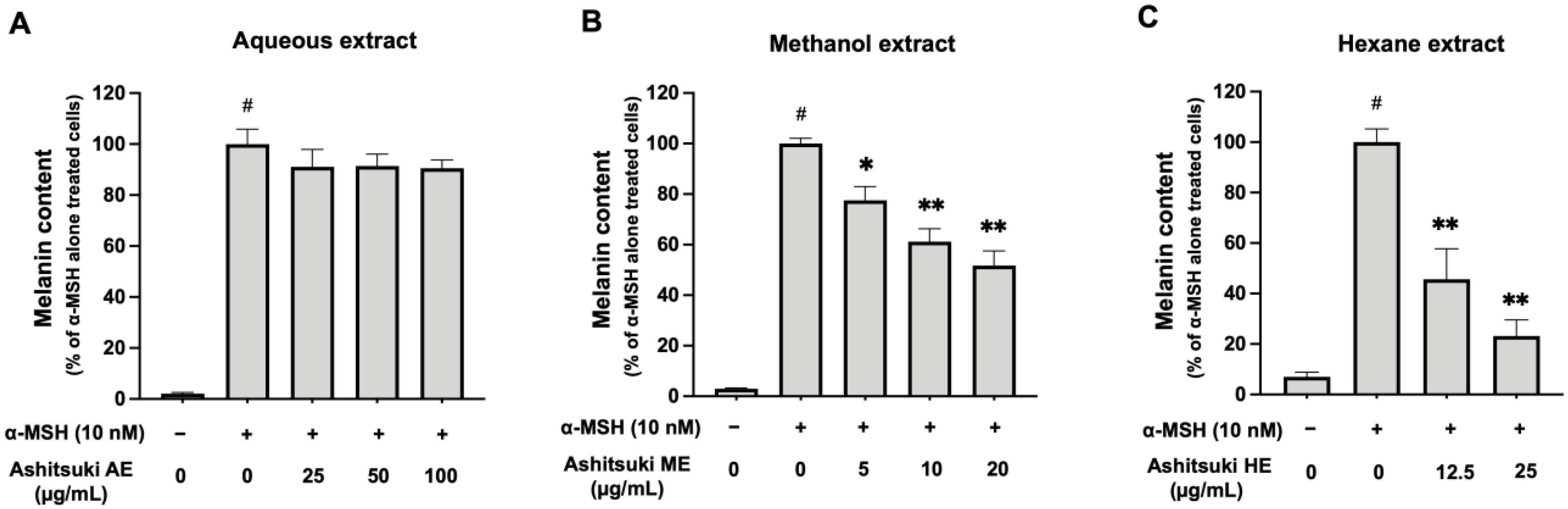
To confirm the effect of N. verrucosum extracts on melanin synthesis, we determined the melanin content. In the presence of α-MSH, the melanin content increased (Figure 6A). However, N. verrucosum AE did not significantly decrease the melanin content. In contrast, the ME inhibited melanin synthesis in B16F1 melanoma cells (approximately 61% at 10 µg/mL and 52% at 20 µg/mL) (Figure 6B). Moreover, the HE inhibited melanin synthesis (approximately 46% at 12.5 µg/mL and 23% at 25.0 µg/mL) (Figure 6C).
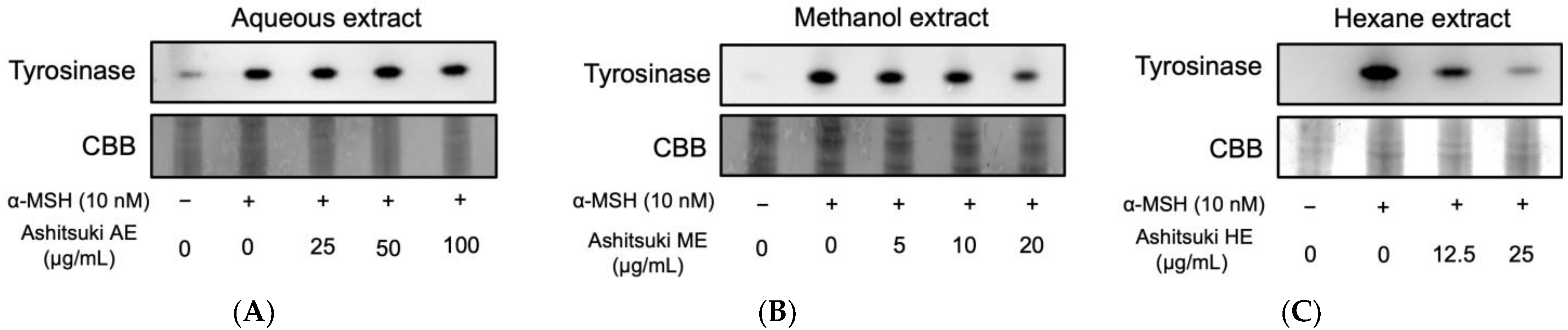
Intracellular tyrosinase activity was assessed using DOPA staining. The ME inhibited the tyrosinase activity in B16F1 melanoma cells (Figure 7). The HE also inhibited tyrosinase activity in a dose-dependent manner. However, the AE-treated cells did not exhibit any change in intracellular tyrosinase activity. These results imply that the ME and HE inhibited melanin synthesis by inhibiting melanogenic protein expression and gene transcription. Therefore, we attempted to elucidate the antimelanogenic mechanism of N. verrucosum HE.
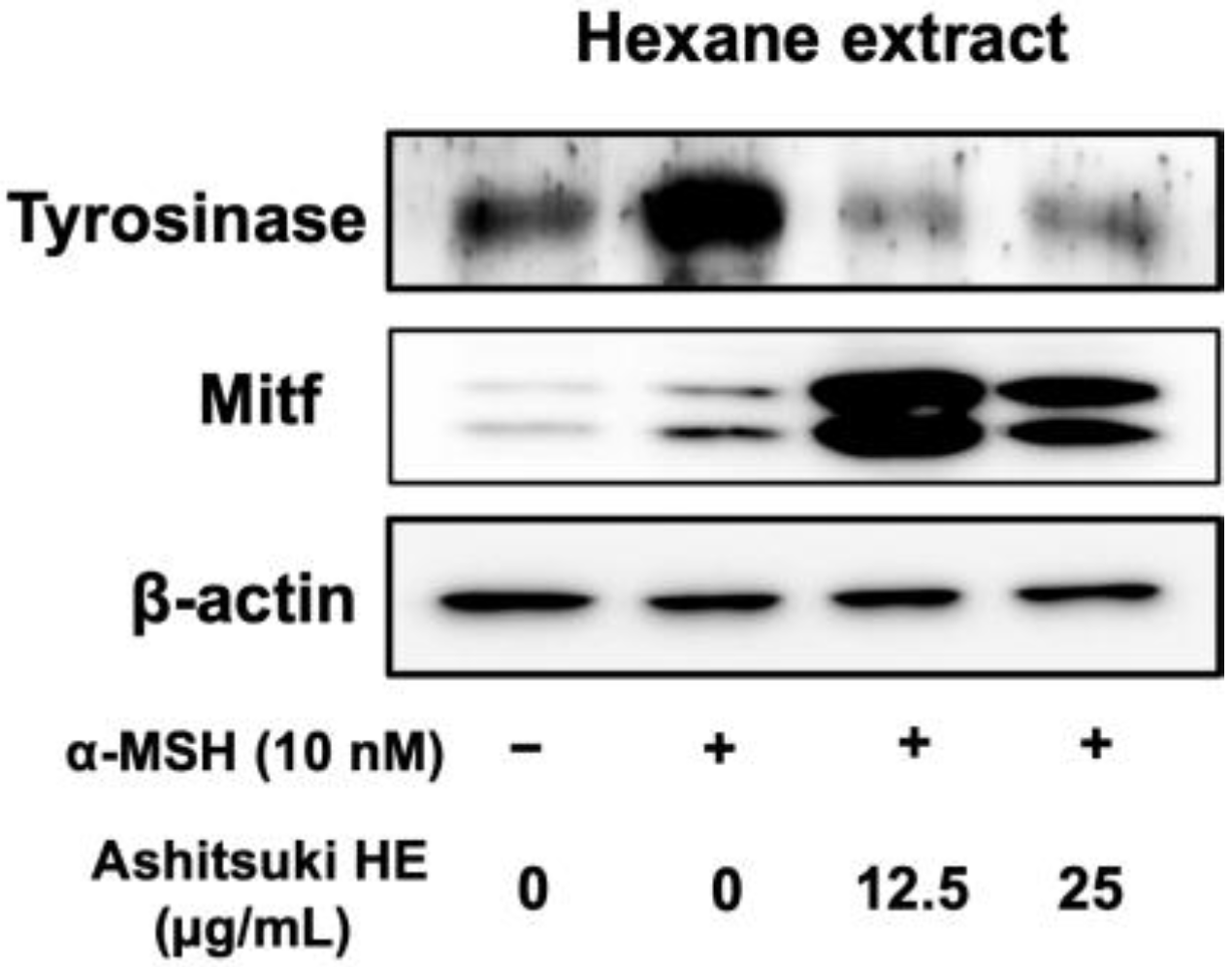
3.5. Effect of N. verrucosum HE on Tyrosinase and Mitf Protein Levels
Based on the results of DOPA staining, N. verrucosum extracts were found to inhibit tyrosinase expression. Therefore, we performed western blotting to determine tyrosinase protein levels. The HE decreased the tyrosinase protein levels (Figure 8). Unexpectedly, the Mitf protein level was upregulated by the N. verrucosum extract treatment.
3.6. Influence of N. verrucosum HE on Melanogenic Genes Transcription
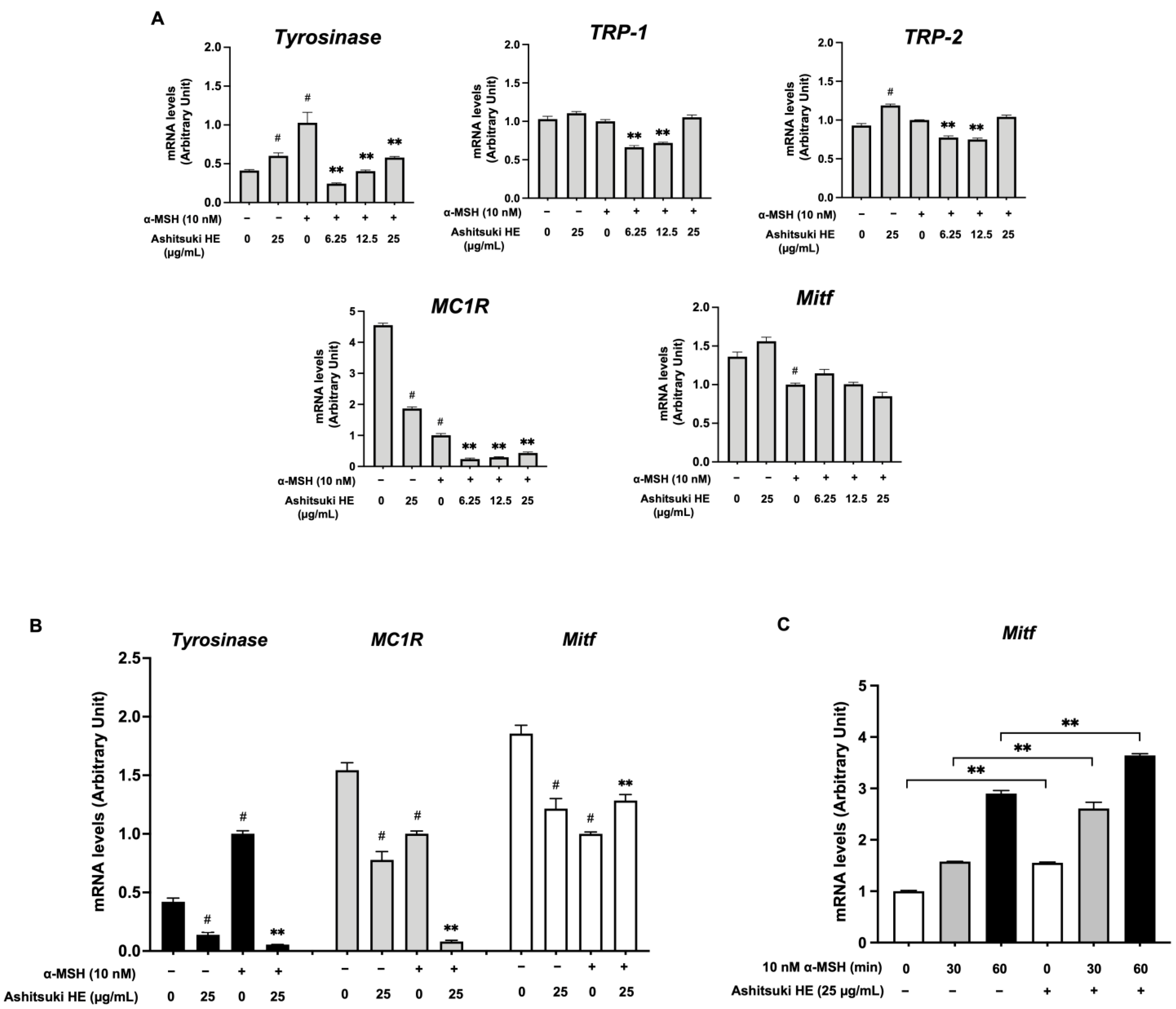
Real-time RT-PCR analysis was performed to elucidate the effect of N. verrucosum on melanogenic gene transcription. Figure 9A shows the mRNA levels of genes in B16F1 melanoma cells after 72 h of treatment with the HE. In the absence of α-MSH, the tyrosinase mRNA level was slightly increased after N. verrucosum extract treatment. In contrast, the MC1R mRNA level was significantly decreased after treatment (approximately 40% of the untreated control). In the presence of α-MSH, the HE suppressed the α-MSH-induced increase in tyrosinase mRNA level. The TRP-1 and TRP-2 mRNA levels were suppressed by treatment with N. verrucosum extract at 6.25 and 12.5 µg/mL. The MC1R mRNA level was decreased in cells treated with α-MSH alone compared with that in the non-treated control. The MC1R mRNA levels in N. verrucosum extract and α-MSH co-treated cells were reduced (approximately 24% at 6.25 µg/mL and 30% at 12.5 µg/mL of the α-MSH-alone-treated control). Unexpectedly, the Mitf mRNA level did not show a significant difference after 3 d of treatment. The transcription levels of genes in the cells treated with N. verrucosum extract for 24 h were also assessed (Figure 9B); the tyrosinase and MC1R mRNA levels significantly decreased the mRNA level after N. verrucosum treatment, regardless of the presence of α-MSH. In the absence of α-MSH, Mitf mRNA transcription was inhibited after N. verrucosum extract treatment; conversely, the mRNA level increased after N. verrucosum extract and α-MSH co-treatment. Additionally, we assessed the Mitf mRNA levels after a short period of α-MSH treatment in the presence of N. verrucosum extract. As shown in Figure 9C, the upregulation of Mitf mRNA expression started shortly after α-MSH stimulation. Compared with that in N. verrucosum-pretreated groups, the Mitf mRNA level did not decrease, but rather increased.
4. Discussion
In this study, we focused on assessing the anti-oxidative and antimelanogenic effects of N. verrucosum extracts. First, we isolated wild N. verrucosum from a stream and cultured it in artificial cultivation systems. Unlike N. commune, N. verrucosum is sensitive to desiccation [3]. Additionally, there is a possibility that the components of N. verrucosum vary between habitats and aquatic environments. Therefore, it is important to develop a robust in vitro culture system for a reliable supply of N. verrucosum.
To evaluate the radical scavenging activity of N. verrucosum extracts, we performed an ABTS assay. The results indicated that all of the extracts possessed radical scavenging capacities. Plant-derived extracts exert both radical scavenging activity and tyrosinase inhibitory activity [15,16,17]. Therefore, we also assessed the effect of N. verrucosum extracts on mushroom tyrosinase activity. Although N. verrucosum has a radical scavenging capacity, we did not observe any inhibitory effect of N. verrucosum on the monophenolase activity of mushroom tyrosinase.
Subsequently, we performed cell culture-based assays. First, we assessed the cell proliferation rate in the presence of N. verrucosum extracts. The AE did not inhibit cell proliferation, whereas ME and HE inhibited the rate of cell growth. The melanin content assay showed that the ME and HE significantly inhibited melanin synthesis in B16F1 melanoma cells. Additionally, the ME and HE decreased the level of the active form of tyrosinase in melanoma cells. These findings suggest that N. verrucosum extracts inhibit melanin synthesis via the downregulation of tyrosinase expression.
To clarify the antimelanogenic effect of N. verrucosum HE, we performed western blotting to determine the protein levels of tyrosinase and Mitf. The results indicated that N. verrucosum extracts significantly inhibited the α-MSH-induced increase in the tyrosinase protein level. In contrast, Mitf expression was upregulated by α-MSH and cotreatment with α-MSH and HE. Hence, we performed real-time RT-PCR to assess the transcription of the melanogenic genes. Tyrosinase mRNA expression was significantly inhibited by N. verrucosum HE treatment. However, The Mitf mRNA level was enhanced within a shorter duration of α-MSH stimulation. Interestingly, MC1R mRNA expression was strongly inhibited in the presence of N. verrucosum HE. These results indicate that N. verrucosum HE primarily inhibits tyrosinase transcription, subsequently downregulating melanin synthesis. MC1R, which is activated by α-MSH, is a receptor expressed on melanocytes and melanoma cells that triggers the melanin synthesis pathway [18]. Hasegawa et al. reported that Elephantopus mollis extract inhibits melanin synthesis via the downregulation of MC1R and Mitf expression, and the subsequent decrease in their mRNA levels [19]. Although N. verrucosum strongly inhibited MC1R transcription, there was no significant change in Mitf mRNA levels. This inconsistency may be attributed to the duration of the treatment. Importantly, downregulating MC1R expression leads to the inhibition of melanoma cell migration [20,21]. Moreover, we performed qRT-PCR analysis to elucidate the effect of N. verrucosum HE on matrix metalloproteinase 1 (MMP-1) transcription using human keratinocyte HaCaT. MMP-1 is involved in collagen degradation, and its enhancement by exposure to ultraviolet light results in the breakdown of type I procollagen level [22,23]. MMP-1 also acts as a metastasis promoter [24,25,26]. Our findings revealed that N. verrucosum HE strongly inhibits MMP-1 mRNA expression (approximately 7% of untreated control) (data not shown). These findings suggest that N. verrucosum can prevent cancer metastasis.
With regard to extracts of other microalgae, Lee et al. reported that Chlamydomonas reinhardtii ethanolic extract can inhibit melanin synthesis via the downregulation of phosphorylated CREB, which leads to Mitf transcription [27]. However, C. reinhardtii did not inhibit tyrosinase activity directly. Kim et al. also investigated the functional effects of Nannochloropsis sp. on skin and reported that it had protective effects [28]. The authors indicated that Nannochloropsis sp. inhibits mushroom tyrosinase activity and exerts an anti-oxidative effect. Moreover, Nannochloropsis sp. suppressed the MMP-1 mRNA level. Additionally, Oliva et al. demonstrated that Botryococcus terribilis ethanolic extract upregulates the expression of hair growth-related genes in human dermal papilla cells [29]. These reports suggest that microalgae have potential to be developed as therapeutic agents or functional food ingredients.
5. Conclusions
In conclusion, our results indicate that N. verrucosum HE may inhibit melanin synthesis by inhibiting tyrosinase gene transcription. We also demonstrated that N. verrucosum possesses anti-oxidant activity. However, the HE did not show strong anti-oxidant activity at the concentration that we used in the melanin content assay. These findings show that the anti-oxidant effect of N. verrucosum HE does not significantly contribute to antimelanogenesis. Additionally, we established a steady cultivation system for N. verrucosum.
However, we believe that additional research on improving the system is required to enhance the beneficial effects of N. verrucosum and further identify compounds with antimelanogenic and anti-oxidative properties.
Author Contributions
Conceptualization, K.S. and H.T.; formal analysis, K.S. and Y.H.; investigation, K.S. and Y.H.; developed the indoor cultivation systems for N. verrucosum, Y.Y., S.S. and H.T.; preparation of N. verrucosum extracts, Y.Y., S.S. and H.T.; writing—original draft preparation, K.S.; writing—review and editing, K.S.; project administration, K.S. and H.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to Professor Keiko Ohashi-Kaneko of Tamagawa University for her technical advice in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Oku, N.; Yonejima, K.; Sugawa, T.; Igarashi, Y. Identification of the n-1 fatty acid as an antibacterial constituent from the edible freshwater cyanobacterium Nostoc verrucosum. Biosci. Biotechnol. Biochem. 2014, 78, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Sakamoto, K.; Nazifi, E.; Tsuji, C.; Asano, T.; Nishiuchi, T.; Matsuo, S.; Ishihara, K.; Kanesaki, Y.; Yoshikawa, H.; Sakamoto, T. Characterization of mycosporine-like amino acids in the cyanobacterium Nostoc verrucosum. J. Gen. Appl. Microbiol. 2018, 64, 201–211. [Google Scholar] [CrossRef]
- Korteerakul, C.; Honda, M.; Ngoennet, S.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Antioxidative and antiglycative properties of mycosporine-like amino acids—Aontaining aqueous extracts derived from edible terrestrial cyanobacteria. J. Nutr. Sci. Vitaminol. 2020, 66, 339–346. [Google Scholar] [CrossRef]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and physiological implications of melanic pigments. Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef]
- Matoba, Y.; Kumagai, T.; Yamamoto, A.; Yoshitsu, H.; Sugiyama, M. Crystallographic evidence that the dinuclear copper center of tyrosinase is flexible during catalysis. J. Biol. Chem. 2006, 281, 8981–8990. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef]
- Kameyama, K.; Takemura, T.; Hamada, Y.; Sakai, C.; Kondoh, S.; Nishiyama, S.; Urabe, K.; Hearing, V.J. Pigment production in murine melanoma cells is regulated by tyrosinase, tyrosinase-related protein 1 (TRP-1), DOPAchrome tautomerase (TRP2), and a melanogenic inhibitor. J. Investig. Dermatol. 1993, 100, 126–131. [Google Scholar] [CrossRef]
- Streingrimsson, E.; Copeland, N.G.; Jenkins, N.A. Melanocytes and the microphthalmia transcription factor network. Annu. Rev. Genet. 2004, 38, 365–411. [Google Scholar] [CrossRef]
- Sato, K.; Yamaguchi, Y.; Sakaki, S.; Takenaka, H. Pleurochrysis carterae hot-water extract inhibits melanogenesis in murine melanoma cells. Cosmetics 2019, 6, 60. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ito, Y.; Sato, K. Salicylamide enhances melanin synthesis in B16F1 melanoma cells. Biomol. Ther. 2021, 29, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takahashi, H.; Toriyama, M. Depigmenting mechanism of NSAIDs on B16F1 melanoma cells. Arch. Dermatol. Res. 2011, 303, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takei, M.; Iyota, R.; Muraoka, Y.; Nagashijma, M.; Yoshimura, Y. Indomethacin inhibits melanogenesis via down-regulation of Mitf mRNA transcription. Biosci. Biotechnol. Biochem. 2017, 81, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Khuanekkaphan, M.; Khobjai, W.; Noysang, C.; Wisidsri, N.; Thungmungmee, S. Bioactivities of Karanda (Carissa carandas Linn.) fruit extracts for novel cosmeceutical applicationsof Completion. J. Adv. Pharm. Technol. Res. 2021, 12, 162–168. [Google Scholar] [CrossRef]
- Worrapan Poomanee, W.; Khunkitti, W.; Chaiyana, W.; Intasai, N.; Lin, W.-C.; Lue, S.-L.; Leelapornpisid, P. Multifunctional biological properties and phytochemical constituents of Mangifera indica L. seed kernel extract for preventing skin aging. Toxicol. Res. 2021, 37, 459–472. [Google Scholar] [CrossRef]
- Shin, K.K.; Park, S.H.; Lim, H.Y.; Lorza, L.R.; Qomaladewia, N.P.; You, L.; Aziz, N.; Kim, S.A.; Lee, J.S.; Choung, E.S.; et al. In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract. Plants 2022, 11, 1383. [Google Scholar] [CrossRef]
- Swope, V.B.; Jameson, J.A.; McFarland, K.L.; Supp, D.M.; Miller, W.E.; McGraw, D.W.; Patel, M.A.; Nix, M.A.; Millhauser, G.L.; Babcock, G.F.; et al. Defining MC1R regulation in human melanocytes by its agonist α-melanocortin and antagonists agouti signaling protein and β-defensin 3. J. Investig. Dermatol. 2012, 132, 2255–2262. [Google Scholar] [CrossRef]
- Hasegawa, K.; Furuya, R.; Mizuno, H.; Umishio, K.; Suetsugu, M.; Sato, K. Inhibitory effect of Elephantopus mollis H.B. and K. extract on melanogenesis in B16 murine melanoma cells by downregulating microphthalmia-associated transcription factor expression. Biosci. Biotechnol. Biochem. 2010, 74, 1908–1912. [Google Scholar] [CrossRef]
- Seong, I.; Min, H.J.; Lee, J.-H.; Yeo, C.-Y.; Kang, D.M.; Oh, E.-S.; Hwang, E.S.; Kim, J. Sox10 controls migration of B16F10 melanoma cells through multiple regulatory target genes. PLoS ONE 2012, 7, e31477. [Google Scholar] [CrossRef]
- Park, J.; Jeong, D.; Jang, B.; Oh, E.-S. The melanocortin-1 receptor reversely regulates the melanin synthesis and migration of melanoma cells via dimerization-induced conformational changes. Biochem. Biophys. Res. Commun. 2019, 518, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Chuliá-Peris, L.; Carreres-Rey, C.; Gabasa, M.; Alcaraz, J.; Carretero, J.; Pereda, J. Matrix metalloproteinases and their inhibitors in pulmonary fibrosis: EMMPRIN/CD147 comes into play. Int. J. Mol. Sci. 2022, 23, 6894. [Google Scholar] [CrossRef] [PubMed]
- Parrado, C.; Mascaraque, M.; Gilaberte, Y.; Juarranz, A.; Gonzalez, S. Fernblock (Polypodium leucotomos Extract): Molecular mechanisms and pleiotropic effects in light-related skin conditions, photoaging and skin cancers, a review. Int. J. Mol. Sci. 2016, 17, 1026. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, L.; Xu, P.; Li, Y.; Ji, Z.; Kang, X. Filamin A is a potential driver of breast cancer metastasis via regulation of MMP-1. Front. Oncol. 2022, 12, 836126. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, A.; Cheng, M.; Bhujwalla, Z.M.; Krishnamachary, B.; Jiang, L.; Glunde, K. Breast cancer cell adhesome and degradome interact to drive metastasis. NPJ Breast Cancer 2015, 1, 15017. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kato, Y.; Erzinger, S.A.; Kiriakova, G.M.; Qian, Y.; Palmieri, D.; Steeg, P.S.; Price, J.E. The role of MMP-1 in breast cancer growth and metastasis to the brain in a xenograft model. BMC Cancer 2012, 12, 583. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Kim, J.Y.; Heo, J.; Cho, D.-H.; Kim, H.-S.; An, I.-S.; An, S.; Bae, S. The inhibition of melanogenesis via the PKA and ERK signaling pathways by Chlamydomonas reinhardtii extract in B16F1 melanoma cells and artificial human skin equivalents. J. Microbiol. Biotechnol. 2018, 28, 2121–2132. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kwon, Y.M.; Kim, K.W.; Kim, J.Y.H. Exploring the potential of Nannochloropsis sp. extract for cosmeceutical applications. Mar. Drugs 2021, 19, 690. [Google Scholar] [CrossRef]
- Oliva, A.K.; Bejaoui, M.; Hirano, A.; Arimura, T.; Linh, T.N.; Uchiage, E.; Nukaga, S.; Tominaga, K.; Nozaki, H.; Isoda, H. Elucidation of the potential hair growth-promoting effect of Botryococcus terribilis, its novel compound methylated-meijicoccenem and C32 botryococcene on cultured hair follicle dermal papilla cells using DNA microarray gene expression analysis. Biomedicines 2022, 10, 1186. [Google Scholar] [CrossRef]
Figure 1.
Nostoc verrucosum (ashitsuki) colony in a shallow stream.

Figure 2.
Nostoc verrucosum cultured in a cylindrical tank under a fluorescent light.

Figure 3.
Anti-oxidant activity of N. verrucosum (ashitsuki) extracts. Anti-oxidant activity of N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts were measured using the assay of ABTS radical scavenging activity. Results are represented as ABTS radical scavenging activity (%); the data are shown as mean ± SEM of three separate experiments. * p < 0.05, ** p < 0.01 versus the control group.
Figure 3.
Anti-oxidant activity of N. verrucosum (ashitsuki) extracts. Anti-oxidant activity of N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts were measured using the assay of ABTS radical scavenging activity. Results are represented as ABTS radical scavenging activity (%); the data are shown as mean ± SEM of three separate experiments. * p < 0.05, ** p < 0.01 versus the control group.

Figure 4.
Effect of N. verrucosum (ashitsuki) extracts on tyrosinase activity. Effect of N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts on monophenolase activity was measured. Results are represented as percentages of the control; the data are shown as mean ± SEM of three separate experiments. * p < 0.05, ** p < 0.01 versus the control group.
Figure 4.
Effect of N. verrucosum (ashitsuki) extracts on tyrosinase activity. Effect of N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts on monophenolase activity was measured. Results are represented as percentages of the control; the data are shown as mean ± SEM of three separate experiments. * p < 0.05, ** p < 0.01 versus the control group.

Figure 5.
Effect of N. verrucosum (ashitsuki) extracts on the proliferation of B16F1 melanoma cells. B16F1 melanoma cells were treated with N. verrucosum aqueous (A), methanol (B), and hexane (C) extract for 3 d, in the presence of 10 nM α-melanocyte-stimulating hormone (α-MSH). After treatment, the trypan blue exclusion test was performed, and the cell proliferation rate was assessed. Results are represented as a percentage of the α-MSH-alone-treated control group; the data are shown as mean ± SEM of three separate experiments. # p < 0.05 versus the control group. * p < 0.05, ** p < 0.01 versus the α-MSH-alone-treated control group.
Figure 5.
Effect of N. verrucosum (ashitsuki) extracts on the proliferation of B16F1 melanoma cells. B16F1 melanoma cells were treated with N. verrucosum aqueous (A), methanol (B), and hexane (C) extract for 3 d, in the presence of 10 nM α-melanocyte-stimulating hormone (α-MSH). After treatment, the trypan blue exclusion test was performed, and the cell proliferation rate was assessed. Results are represented as a percentage of the α-MSH-alone-treated control group; the data are shown as mean ± SEM of three separate experiments. # p < 0.05 versus the control group. * p < 0.05, ** p < 0.01 versus the α-MSH-alone-treated control group.

Figure 6.
Effect of N. verrucosum (ashitsuki) extracts on melanin synthesis in B16F1 melanoma cells. B16F1 melanoma cells were treated with N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts for 3 d in the presence of 10 nM α-MSH. After treatment, intracellular melanin content was measured as described in the materials and methods section. Results are represented as percentages of the α-MSH-alone-treated control group, the data are shown as mean ± SEM of three separate experiments. # p < 0.05 versus the control group. * p < 0.05, and ** p < 0.01 versus the α-MSH-alone-treated control group.
Figure 6.
Effect of N. verrucosum (ashitsuki) extracts on melanin synthesis in B16F1 melanoma cells. B16F1 melanoma cells were treated with N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts for 3 d in the presence of 10 nM α-MSH. After treatment, intracellular melanin content was measured as described in the materials and methods section. Results are represented as percentages of the α-MSH-alone-treated control group, the data are shown as mean ± SEM of three separate experiments. # p < 0.05 versus the control group. * p < 0.05, and ** p < 0.01 versus the α-MSH-alone-treated control group.

Figure 7.
Effect of N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts on intracellular active tyrosinase. B16F1 melanoma cells were treated with N. verrucosum extracts for 3 d in the presence of 10 nM α-MSH. After treatment, cell lysates were subjected to SDS-PAGE and DOPA staining was performed.
Figure 7.
Effect of N. verrucosum aqueous (A), methanol (B), and hexane (C) extracts on intracellular active tyrosinase. B16F1 melanoma cells were treated with N. verrucosum extracts for 3 d in the presence of 10 nM α-MSH. After treatment, cell lysates were subjected to SDS-PAGE and DOPA staining was performed.

Figure 8.
Effect of N. verrucosum (ashitsuki) hexane extract on tyrosinase and Mitf protein levels. B16F1 melanoma cells were treated with N. verrucosum hexane extract for 72 h in the presence of 10 nM α-MSH. After treatment, total cell lysates were prepared, and western blot analysis was performed.
Figure 8.
Effect of N. verrucosum (ashitsuki) hexane extract on tyrosinase and Mitf protein levels. B16F1 melanoma cells were treated with N. verrucosum hexane extract for 72 h in the presence of 10 nM α-MSH. After treatment, total cell lysates were prepared, and western blot analysis was performed.

Figure 9.
Effect of N. verrucosum (ashitsuki) hexane extract on melanogenic gene transcription. B16F1 melanoma cells were treated with N. verrucosum hexane extract and stimulated with 10 nM α-MSH for 72 h (A) and 6 h (B). (C) The cells were pre-treated with N. verrucosum hexane extract for 1 h; thereafter, α-MSH (10 nM) was added to the cells, which were then incubated for 30 or 60 min in the presence of α-MSH. Total RNA was extracted and cDNA was prepared. An equivalent amount of cDNA was amplified using primers specific for tyrosinase, TRP-1, TRP-2, Mitf, and MC1R. Results are represented as arbitrary unit of the α-MSH-alone-treated control group; the data are mean ± SEM of three separate experiments. # p < 0.05 versus the control group. ** p < 0.01 versus the α-MSH-alone-treated control group.
Figure 9.
Effect of N. verrucosum (ashitsuki) hexane extract on melanogenic gene transcription. B16F1 melanoma cells were treated with N. verrucosum hexane extract and stimulated with 10 nM α-MSH for 72 h (A) and 6 h (B). (C) The cells were pre-treated with N. verrucosum hexane extract for 1 h; thereafter, α-MSH (10 nM) was added to the cells, which were then incubated for 30 or 60 min in the presence of α-MSH. Total RNA was extracted and cDNA was prepared. An equivalent amount of cDNA was amplified using primers specific for tyrosinase, TRP-1, TRP-2, Mitf, and MC1R. Results are represented as arbitrary unit of the α-MSH-alone-treated control group; the data are mean ± SEM of three separate experiments. # p < 0.05 versus the control group. ** p < 0.01 versus the α-MSH-alone-treated control group.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sato, K.; Hiraga, Y.; Yamaguchi, Y.; Sakaki, S.; Takenaka, H. Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts. Cosmetics 2023, 10, 30. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10010030
AMA Style
Sato K, Hiraga Y, Yamaguchi Y, Sakaki S, Takenaka H. Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts. Cosmetics. 2023; 10(1):30. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10010030
Chicago/Turabian StyleSato, Kazuomi, Yosuke Hiraga, Yuji Yamaguchi, Setsuko Sakaki, and Hiroyuki Takenaka. 2023. "Anti-Melanogenic and Anti-Oxidative Effects of Nostoc verrucosum (ashitsuki) Extracts" Cosmetics 10, no. 1: 30. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics10010030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.