In Vitro Hair Dermal Papilla Cells Induction by Fagraea berteroana, a Tree of the Marquesan Cosmetopoeia (French Polynesia)

 , ,
, ,
Abstract
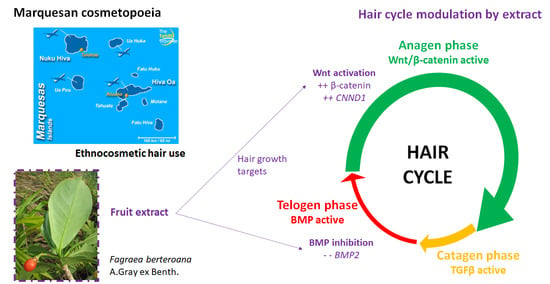
:
1. Introduction
2. Materials and Methods
2.1. Plant Material Collection, Extraction, and Fractionation
2.2. Cell Viability via the MTT Assay
2.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. Direct Immunofluorescence Staining
2.5. Statistical Analysis
2.6. UHPLC-MS/MS Analysis of F. berteroana Fraction FEAE-F0
3. Results and Discussion
3.1. Hair Follicle Dermal Papilla Cells (HDPCs) Proliferation Dynamic after Treatment with Extracts and Target Genes Regulation
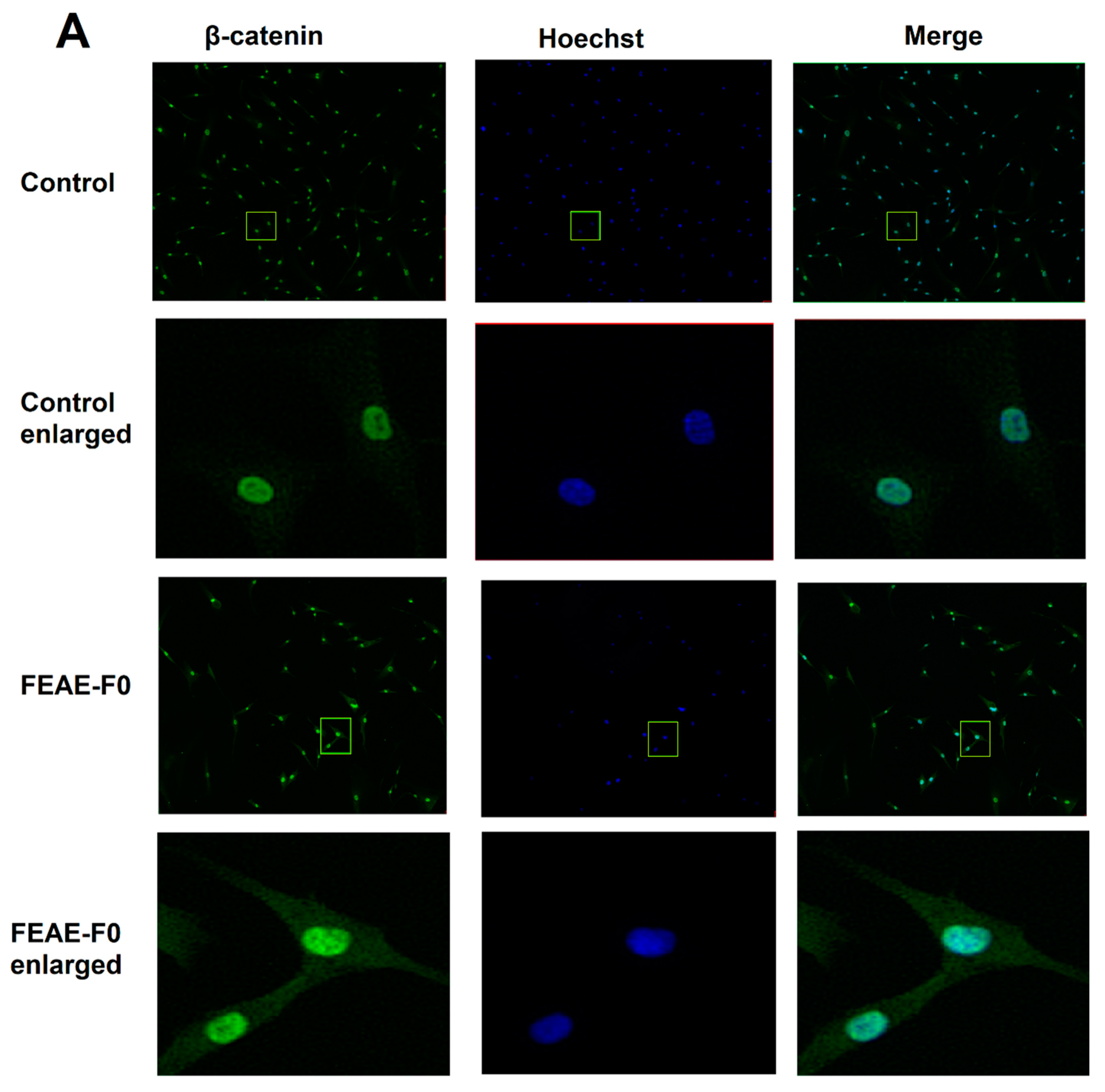
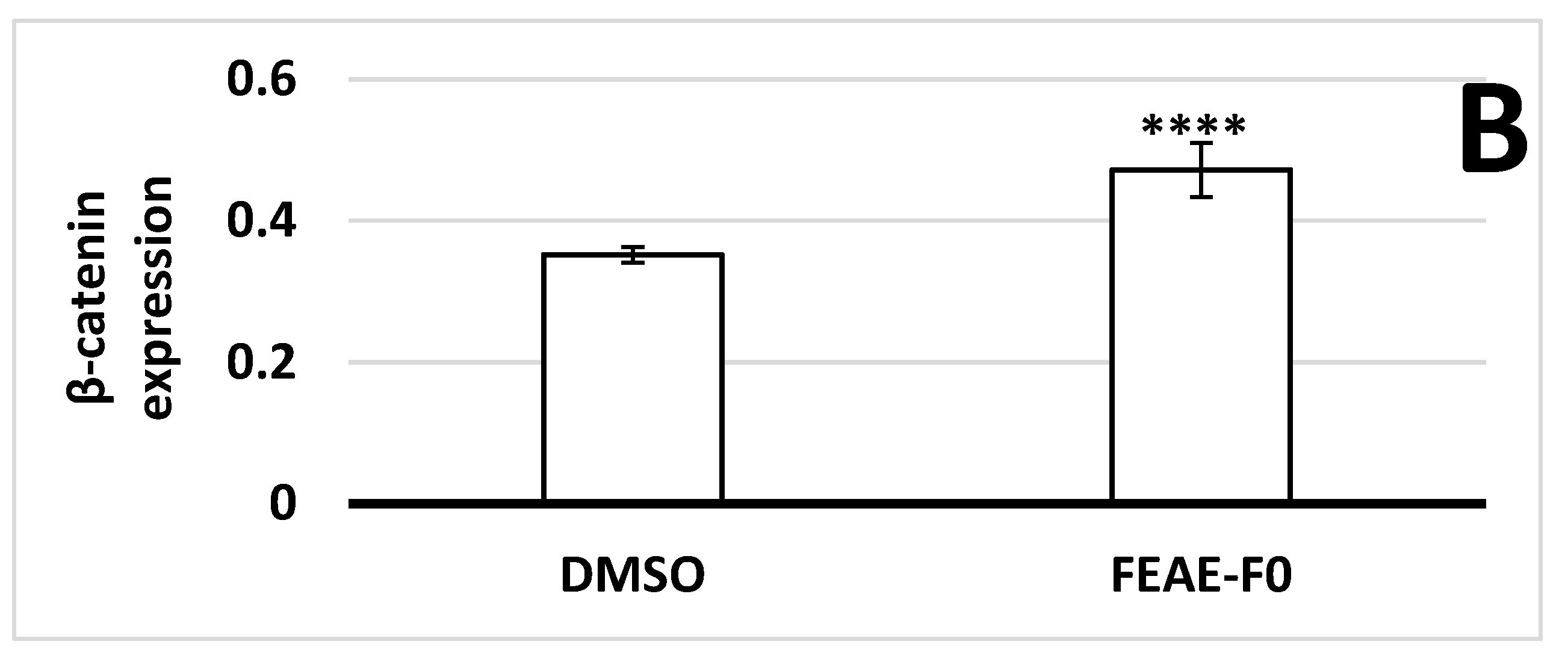
3.2. Modulation of β-Catenin Production by FEAE-F0
3.3. Chemical Composition of F. berteroana FEAE-F0
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Butaud, J.-F. Synthèse Bibliographique Portant Sur Les Plantes Utilisées Dans La Cosmétopée de Polynésie Française; Cosmetic Valley: Chartres, France, 2013. [Google Scholar]
- Ansel, J.-L.; Ly, Q.; Butaud, J.-F.; Nicolas, M.; Herbette, G.; Peno-Mazzarino, L.; Lati, E.; Raharivelomanana, P. Activité anti-âge de l’extrait de Fitchia nutans, un ingrédient cosméticeutique d’un monoï traditionnel polynésien. Comptes Rendus Chim. 2016, 19, 1049–1055. [Google Scholar] [CrossRef]
- Jost, X.; Ansel, J.-L.; Lecellier, G.; Raharivelomanana, P.; Butaud, J.-F. Ethnobotanical Survey of Cosmetic Plants Used in Marquesas Islands (French Polynesia). J. Ethnobiol. Ethnomed. 2016, 12. [Google Scholar] [CrossRef]
- Martin, A. The Enlightenment in Paradise: Bougainville, Tahiti, and the Duty of Desire. Eighteenth Century Stud. 2007, 41, 203–216. [Google Scholar] [CrossRef]
- Gauthier, L. TAHITI—1904-1921; Les éditions du Pacifique: Paris, France, 1985. [Google Scholar]
- Hughes, K.; Ho, R.; Butaud, J.-F.; Filaire, E.; Ranouille, E.; Berthon, J.-Y.; Raharivelomanana, P. A Selection of Eleven Plants Used as Traditional Polynesian Cosmetics and Their Development Potential as Anti-Aging Ingredients, Hair Growth Promoters and Whitening Products. J. Ethnopharmacol. 2019, 245, 112159. [Google Scholar] [CrossRef] [PubMed]
- Handy, E.S.C. The Native Culture in the Marquesas; Bernice P. Bishop Museum: Honolulu, HI, USA, 1923. [Google Scholar]
- Pétard, P. Plantes Utiles de Polynésie Française et Raau Tahiti; Editions Haere Po No Tahiti: Papeete, French Polynesia, 1986; ISBN 978-2-904171-06-2. [Google Scholar]
- Whistler, W.A.; Elevitch, C.R. Fagraea berteroana (Pua kenikeni), Ver. 3.2.; Volume Species Profiles for Pacific Island Agroforestry; Elevitch, C.R., Ed.; Permanent Agriculture Resources (PAR): Honolulu, HI, USA, 2006. [Google Scholar]
- Hayashi, S.; Kameoka, H.; Hashimoto, S.; Furukawa, K.; Arai, T. Volatile Compounds of Fagraea berteriana Flowers. J. Essent. Oil Res. 1995, 7, 505–510. [Google Scholar] [CrossRef]
- Whistler, W.A. Plants in Samoan Culture: The Ethnobotany of Samoa; Isle Botanica: Honolulu, HI, USA, 2000; ISBN 978-0-9645426-6-2. [Google Scholar]
- Motley, T.J. The Ethnobotany of Fagraea Thunb. (Gentianaceae): The Timber of Malesia and the Scent of Polynesia. Econ. Bot. 2004, 58, 396–409. [Google Scholar] [CrossRef]
- Cuendet, M.; Hostettmann, K.; Potterat, O.; Dyatmiko, W. Iridoid Glucosides with Free Radical Scavenging Properties from Fagraea blumei. Helv. Chim. Acta 1997, 80, 1144–1152. [Google Scholar] [CrossRef]
- Cambie, R.C.; Lal, A.R.; Rickard, C.E.F.; Tanaka, N. Chemistry of Fijian Plants. V.: Constituents of Fagraea gracilipes A. GRAY. Chem. Pharm. Bull. 1990, 38, 1857–1861. [Google Scholar] [CrossRef] [Green Version]
- Ferdinal, N.; Alfajri, R.; Arifin, B. Isolation and Characterization of Scopoletin from The Bark of Fagraea ceilanica Thumb and Antioxidants Tests. Int. J. Adv. Sci. Eng. Inf. Technol. 2015, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Bangprapai, A.; Thongphasuk, P.; Songsak, T. Determination of Swertiamarin Content by Tlc-Densitometer in Fagraea fragrans Roxb. Leaves. Bull. Health Sci. Technol. 2016, 14, 13–18. [Google Scholar]
- Jonville, M.-C.; Capel, M.; Frédérich, M.; Angenot, L.; Dive, G.; Faure, R.; Azas, N.; Ollivier, E. Fagraldehyde, a Secoiridoid Isolated from Fagraea fragrans. J. Nat. Prod. 2008, 71, 2038–2040. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, E.; Suzumura, K.; Yamazaki, M. Pharmacologically Active Components of Todopon Puok (Fagraea racemosa), a Medicinal Plant from Borneo. Chem. Pharm. Bull. 1995, 43, 2200–2204. [Google Scholar] [CrossRef] [Green Version]
- Madmanang, S.; Cheyeng, N.; Heembenmad, S.; Mahabusarakam, W.; Saising, J.; Seeger, M.; Chusri, S.; Chakthong, S. Constituents of Fagraea fragrans with Antimycobacterial Activity in Combination with Erythromycin. J. Nat. Prod. 2016, 79, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Rattanaburi, S.; Kaikaew, K.; Watanapokasin, R.; Phongpaichit, S.; Mahabusarakamb, W. A New Lignan from the Stem Bark of Fagraea fragrans Roxb. Nat. Prod. Res. 2020, 1–6. [Google Scholar] [CrossRef]
- Wan, A.S.C.; Chow, Y.L. Alkaloids of Fagraea fragrans Roxb. J. Pharm. Pharmacol. 1964, 16, 484–486. [Google Scholar] [CrossRef] [PubMed]
- Basir, D.; Hanafi, M.; Saputra, A.; Wati, T. Free Solvent Amidation of Ursolic and Oleanolic Acids of Fagraea fragrans Fruits: Their P-388 Antitumor Activity. J. Phys. Conf. Ser. 2018, 1095, 012006. [Google Scholar] [CrossRef]
- Tamura, Y.; Takata, K.; Eguchi, A.; Kataoka, Y. In Vivo Monitoring of Hair Cycle Stages via Bioluminescence Imaging of Hair Follicle NG2 Cells. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Plikus, M.V. New Activators and Inhibitors in the Hair Cycle Clock: Targeting Stem Cells’ State of Competence. J. Investig. Dermatol. 2012, 132, 1321–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inui, S.; Itami, S. Molecular Basis of Androgenetic Alopecia: From Androgen to Paracrine Mediators through Dermal Papilla. J. Dermatol. Sci. 2011, 61, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kwack, M.H.; Kang, B.M.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Minoxidil Activates β-catenin Pathway in Human Dermal Papilla Cells: A Possible Explanation for Its Anagen Prolongation Effect. J. Dermatol. Sci. 2011, 62, 154–159. [Google Scholar] [CrossRef]
- Zhang, H.; Su, Y.; Wang, J.; Gao, Y.; Yang, F.; Li, G.; Shi, Q. Ginsenoside Rb1 Promotes the Growth of Mink Hair Follicle via PI3K/AKT/GSK-3β Signaling Pathway. Life Sci. 2019, 229, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Dastan, M.; Najafzadeh, N.; Abedelahi, A.; Sarvi, M.; Niapour, A. Human Platelet Lysate versus Minoxidil Stimulates Hair Growth by Activating Anagen Promoting Signaling Pathways. Biomed. Pharmacother. 2016, 84, 979–986. [Google Scholar] [CrossRef]
- Oshimori, N.; Fuchs, E. Paracrine TGF-β Signaling Counterbalances BMP-Mediated Repression in Hair Follicle Stem Cell Activation. Cell Stem Cell 2012, 10, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Veltri, A.; Lang, C.; Lien, W.-H. Concise Review: Wnt Signaling Pathways in Skin Development and Epidermal Stem Cells: Wnt Signaling in Skin Development and Stem Cells. Stem Cells 2018, 36, 22–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, L. QBI, The University of Queensland Australia Measuring Cell Fluorescence Using ImageJ—The Open Lab Book v1.0. Available online: https://theolb.readthedocs.io/en/latest/imaging/measuring-cell-fluorescence-using-imagej.html (accessed on 15 October 2020).
- MassBank of North America (MoNA)—PubChem Data Source. Available online: https://pubchem.ncbi.nlm.nih.gov/source/22043 (accessed on 20 August 2020).
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A Public Repository for Sharing Mass Spectral Data for Life Sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Inui, S.; Itami, S. Androgen Actions on the Human Hair Follicle: Perspectives. Exp. Dermatol. 2013, 22, 168–171. [Google Scholar] [CrossRef]
- Warner, D.R.; Greene, R.M.; Pisano, M.M. Cross-Talk between the TGFβ and Wnt Signaling Pathways in Murine Embryonic Maxillary Mesenchymal Cells. FEBS Lett. 2005, 579, 3539–3546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.-Q.; Wu, Z.-B.; Chu, X.-Y.; Bi, Z.-G.; Fan, W.-X. An Investigation of Crosstalk between Wnt/β-catenin and Transforming Growth Factor-β Signaling in Androgenetic Alopecia. Medicine 2016, 95, e4297. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.Y. Targeting Wnt/β-catenin Pathway for Developing Therapies for Hair Loss. Int. J. Mol. Sci. 2020, 21, 4915. [Google Scholar] [CrossRef] [PubMed]
- Kimelman, D.; Xu, W. β -catenin Destruction Complex: Insights and Questions from a Structural Perspective. Oncogene 2006, 25, 7482–7491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamos, J.L.; Weis, W.I. The β-catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef]
- Shtutman, M.; Zhurinsky, J.; Simcha, I.; Albanese, C.; D’Amico, M.; Pestell, R.; Ben-Ze’ev, A. The Cyclin D1 Gene Is a Target of the β-catenin/LEF-1 Pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5522–5527. [Google Scholar] [CrossRef] [Green Version]
- Schneider, J.A.; Logan, S.K. Revisiting the Role of Wnt/β-catenin Signaling in Prostate Cancer. Mol. Cell. Endocrinol. 2018, 462, 3–8. [Google Scholar] [CrossRef]
- Hughes, K.; Ho, R.; Greff, S.; Filaire, E.; Ranouille, E.; Chazaud, C.; Herbette, G.; Butaud, J.-F.; Berthon, J.-Y.; Raharivelomanana, P. Hair Growth Activity of Three Plants of the Polynesian Cosmetopoeia and Their Regulatory Effect on Dermal Papilla Cells. Molecules 2020, 25, 4360. [Google Scholar] [CrossRef] [PubMed]
- Suciati, S.; Lambert, L.K.; Ross, B.P.; Deseo, M.A.; Garson, M.J. Phytochemical Study of Fagraea Spp. Uncovers a New Terpene Alkaloid with Anti-Inflammatory Properties. Aust. J. Chem. 2011, 64, 489. [Google Scholar] [CrossRef]
- Villasenor, I. Bioactivities of Iridoids. AIAAMC 2007, 6, 307–314. [Google Scholar] [CrossRef]
- Viljoen, A.; Mncwangi, N.; Vermaak, I. Anti-Inflammatory Iridoids of Botanical Origin. CMC 2012, 19, 2104–2127. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Man, X.-Y.; Li, C.-M.; Chen, J.-Q.; Zhou, J.; Cai, S.-Q.; Lu, Z.-F.; Zheng, M. VEGF Induces Proliferation of Human Hair Follicle Dermal Papilla Cells through VEGFR-2-Mediated Activation of ERK. Exp. Cell Res. 2012, 318, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Lachgar; Charveron; Gall; Bonafe Minoxidil Upregulates the Expression of Vascular Endothelial Growth Factor in Human Hair Dermal Papilla Cells. Br. J. Dermatol. 1998, 138, 407–411. [CrossRef]
- Messenger, A.G.; Rundegren, J. Minoxidil: Mechanisms of Action on Hair Growth. Br. J. Dermatol. 2004, 150, 186–194. [Google Scholar] [CrossRef]
- Shumbe, L.; Bott, R.; Havaux, M. Dihydroactinidiolide, a High Light-Induced β-Carotene Derivative That Can Regulate Gene Expression and Photoacclimation in Arabidopsis. Mol. Plant 2014, 7, 1248–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, C.L.R. Terpene Extract for the Treatment of Hair Loss 2013. Available online: https://patentscope2.wipo.int/search/en/detail.jsf?docId=WO2013174854 (accessed on 31 January 2021).
- Takahashi, H.; Hara, S.c/oK.K.Y.C.K.; Matsui, R.c/oK.K.Y.C.K. Diterpenes and Flavonoids as 5-Alpha-Reductase Inhibitors 1996. Available online: https://patents.google.com/patent/EP0747048A2/en (accessed on 31 January 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Forward Primer | Reverse Primer |
|---|---|---|
| BMP2 | 5′-CCC-ACT-TGG-AGG-AGA-AAC-AA-3′ | 5′-GCT-GTT-TGT-GTT-TGG-CTT-GA-3′ |
| CALM2 | 5′-GGG-AAC-ATC-TGG-GTT-ATG-CC-3′ | 5′-GAC-TGT-CCA-TAG-TCC-ACG-CA-3′ |
| CCND1 | 5′-AAC-TAC-CTG-GAC-CGC-TTC-CT-3′ | 5′-CCA-CTT-GAG-CTT-GTT-CAC-CA-3′ |
| GAPDH | 5′-CCA-GCA-AGA-GCA-CAA-GAG-GA-3′ | 5′-TGG-TTG-AGC-ACA-GGG-TAC-TT-3′ |
| PUM1 | 5′-GGT-GCC-CTT-GTA-GTG-AAT-GC-3′ | 5′-TGT-TGT-TCC-AGC-AAG-ACC-AC-3′ |
| TGFB1 | 5′-CTG-GCG-ATA-CCT-CAG-CAA-CC-3′ | 5′-CGG-TAG-TGA-ACC-CGT-TGA-TGT-C-3′ |
| Positive Mode | Negative Mode | UV (nm) | Database Search (21/01/2021) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Peak No. | Rt (min) | [M + H]+ | [M + Na]+ | Product Ions (MS/MS, 20 and 40 eV) | [M − H]− | Product Ions (MS/MS) | Metlin | MetFrag (vs. Pubchem) | Molecular Formula | Compound Annotation | |
| 1 | 7.67 | 171.1019 | 193.084 | 20 eV: 57 (100, C4H9+), 83 (10, C5H7O+), 111 (19, C7H11O+), 125 (4, C8H13O+) 40 eV: 55 (21, C4H7+), 57 (100, C4H9+), 59 (10, C3H7O+), 97 (17, -), 145 (10, -) | - | - | 224 | 3 entries, no experimental MS2 spectra | 5240 candidates | C9H14O3 | boonein |
| 2 | 7.82 | 181.0861 | 203.0681 | 20 eV: 57 (100, C3H5O+), 117 (6, C9H9+), 131 (4, -), 151 (3, C8H7O3+) 40 eV: 57 (100, C3H5O+), 91 (57, C7H7+), 109 (36, -), 115 (50, C9H7+), 127 (21, -), 152 (20, -) | - | - | 225, 274 | 21 entries, 7 experimental MS2 spectra, no correspondence | 3909 candidates | C10H12O3 | - |
| 3 | 8.22 | 153.0915 | 175.0734 | 20 eV: 67 (46, C5H7+), 79 (85, C6H7+), 91 (74, C7H7+), 105 (25, C8H9+), 107 (100, C8H11+) 40 eV: 53 (28, C4H5+), 55 (22, C4H7+), 65 (71, C5H5+), 67 (27, C5H7+), 79 (44, C6H7+), 91 (100, C7H7+) | - | - | 226, 279 | 13 entries, 2 experimental MS2 spectra, no correspondence | 3668 candidates | C9H12O2 | - |
| 4 | 8.57 | 213.1454 | - | 20 eV: - 40 eV: 55 (39, C4H7+), 57 (28, -), 91 (67, -), 91 (64, C7H7+), 95 (41, C6H7O+), 145 (100, -) | - | - | 226, 279 | 13 entries, 2 experimental MS2 spectra, no correspondence | 4703 candidates | C12H20O3 | - |
| 5 | 8.83 | - | - | - | - | - | 226, 279 | - | - | ||
| 6 | 9.00 | 181.1223 | 203.1044 | 20 eV: 79 (21, C6H7+), 81 (29, C6H9+), 93 (64, C7H9+), 95 (56, C7H11+), 107 (93, C8H11+), 121 (42, C9H13+), 135 (84, C10H15+), 145 (22, C11H13+), 163 (58, C11H15O+), 181 (100, C11H17O2+) 40 eV: 53 (33, C4H5+), 55 (40, C4H7+), 67 (31, C5H7+), 69 (17, C5H9+), 79 (69, C6H7+), 91 (100, C7H7+), 93 (47, C7H9+), 105 (47, C8H9+), 107 (24, C8H11+) | - | - | 226, 284 | 12 entries, 1 experimental MS2 spectrum hypothetical match Metlin ID 90552 | 6090 candidates | C11H16O2 | - |
| 7 | 9.36 | - | - | - | 217.0874 | 20 eV: 117.0346 (33, C8H5O−), 119.0503 (100, C8H7O−), 145.0296 (51, C9H5O2−), 163.0402 (28, C9H7O3−), 217.0873 (9, C13H13O3−) 40 eV: 89 (3, C7H5−), 93 (5, C6H5O−), 117 (100, C8H5O−), 119 (38, C8H7O−), 145 (5, C9H5O2−) | 227 | 7 entries, 1 experimental MS2 spectrum, no correspondence | 5081 candidates | C13H14O3 | - |
| 8 | 9.45 | 213.1484 | 235.1306 | 20 eV: 57 (100, C4H9+), 81 (4, C6H9+), 83 (5, C6H11+), 109 (5, C7H9O+), 127 (48, C7H11O2+), 139 (6, C8H11O2+), 195 (10, C12H19O2+) 40 eV: 57 (100, C4H9+), 79 (8, C6H7+), 81 (14, C6H9+), 109 (3, C7H9O+), 127 (8, C7H11O2+) | 211.1344 (low) | - | 227 | 13 entries, 2 experimental MS2 spectra, no correspondence | 4703 candidates | C12H20O3 | - |
| 9 | 9.52 | 223.0965 | 247.0745 | 20 eV: 121 (2, C7H5O2+), 149 (100, C8H5O3+), 177 (7, C10H9O3+) 40 eV: 65 (80, C5H5+), 93 (19, C6H5O+), 111 (10, C6H7O2+), 121 (72, C7H5O2+), 149 (100, C8H5O3+), 167 (4, C8H7O4+) | - | - | 228, 277 | 13 entries, 1 experimental MS2 spectrum, no correspondence | 6105 candidates | C12H14O4 | - |
| 10 | 9.80 | - | - | - | 195.1393 | 20 eV: 109 (8, C7H9O−), 111 (6, C7H11O−), 125 (3, C8H13O−), 138 (12, C8H10O2−), 167 (16, C11H19O−), 179 (24, C11H15O2−), 195 (100, C12H19O2−) 40 eV: 53 (100, C3HO−), 109 (11, C7H9O−), 122 (14, C7H6O2−), 138 (14, C8H10O2−) | 227 | 59 entries, 1 experimental MS2 spectrum, no correspondence | 6334 candidates | C12H20O2 | - |
| 11 | 10.50 | 181.1224 | 203.1045 | 20 eV: 93 (53, C7H9+), 95 (28, C6H7O+), 97 (25, C6H9O+), 107 (32, C8H11+), 111 (17, C7H11O+), 125 (100, C7H9O2+), 135 (37, C10H15+), 138 (22, C8H10O2+), 163 (28, C11H15O+), 181 (94, C11H17O2+) 40 eV: 53 (50, C4H5+), 55 (40, C4H7+), 65 (20, C5H5+), 67 (44, C5H7+), 77 (19, C6H5+), 79 (38, C6H7+), 81 (12, C6H9+), 91 (100, C7H7+), 93 (42, C7H9+), 95 (70, C6H7O+), 97 (22, C6H9O+), 105 (21, C8H9+), 109 (20, C7H9O+), 123 (11, C7H7O2+), 125 (12, C7H9O2+) | - | - | 228, 276 | 12 entries, 1 experimental MS2 spectrum no close match | 6090 candidates | C11H16O2 | - |
| 12 | 10.98 | 209.1538 | 231.1357 | 20 eV: - 40 eV: 53 (10, C4H5+), 55 (36, C4H7+), 57 (100, C4H9+), 59 (30, C3H7O+), 65 (7, C5H5+), 67 (56, C5H7+), 69 (15, C5H9+), 77 (5, C6H5+), 79 (42, C6H7+), 81 (31, C6H9+), 83 (4, C5H7O+, 5, C6H11+), 91 (49, C7H7+), 93 (31, C7H9+), 95 (9, C6H7O+, 15, C7H11+), 97 (12, C6H9O+), 103 (6, C8H7+), 105 (46, C8H9+), 107 (37, C8H11+), 109 (8, C7H9O+), 111 (4, C7H11O+), 119 (18, C9H11+), 121 (17, C9H13+), 123 (3, C7H7O2+, C8H11O+, C9H15+), 133 (10, C10H13+), 135 (8, C10H15+), 151 (6, C10H15O+), 179 (11, C11H15O2+), 194 (4, C12H18O2+) | - | - | 228, 292 | 13 entries, no experimental MS2 spectra | 6905 candidates | C13H20O2 | - |
| 13 | 11.45 | 279.1594 | 301.1414 | 20 eV: 57 (10, C4H9+), 109 (2, C8H13+), 149 (100, C8H5O3+), 167 (5, C8H7O4+), 209 (2, C10H9O5+) 40 eV: 57 (19, C4H9+), 121 (23, C7H5O2+), 149 (100, C8H5O3+), 167 (5, C8H7O4+) | - | - | 227, 279 | 4 entries, 1 experimental MS2 spectrum, no correspondence | 5392 candidates | C16H22O4 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hughes, K.; Ho, R.; Chazaud, C.; Hermitte, S.; Greff, S.; Butaud, J.-F.; Filaire, E.; Ranouille, E.; Berthon, J.-Y.; Raharivelomanana, P. In Vitro Hair Dermal Papilla Cells Induction by Fagraea berteroana, a Tree of the Marquesan Cosmetopoeia (French Polynesia). Cosmetics 2021, 8, 13. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010013
Hughes K, Ho R, Chazaud C, Hermitte S, Greff S, Butaud J-F, Filaire E, Ranouille E, Berthon J-Y, Raharivelomanana P. In Vitro Hair Dermal Papilla Cells Induction by Fagraea berteroana, a Tree of the Marquesan Cosmetopoeia (French Polynesia). Cosmetics. 2021; 8(1):13. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010013
Chicago/Turabian StyleHughes, Kristelle, Raimana Ho, Claire Chazaud, Stéphanie Hermitte, Stéphane Greff, Jean-François Butaud, Edith Filaire, Edwige Ranouille, Jean-Yves Berthon, and Phila Raharivelomanana. 2021. "In Vitro Hair Dermal Papilla Cells Induction by Fagraea berteroana, a Tree of the Marquesan Cosmetopoeia (French Polynesia)" Cosmetics 8, no. 1: 13. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010013