
The Ethanol Extract of Musa sapientum Linn. Peel Inhibits Melanogenesis through AKT Signaling Pathway

 , and
, and 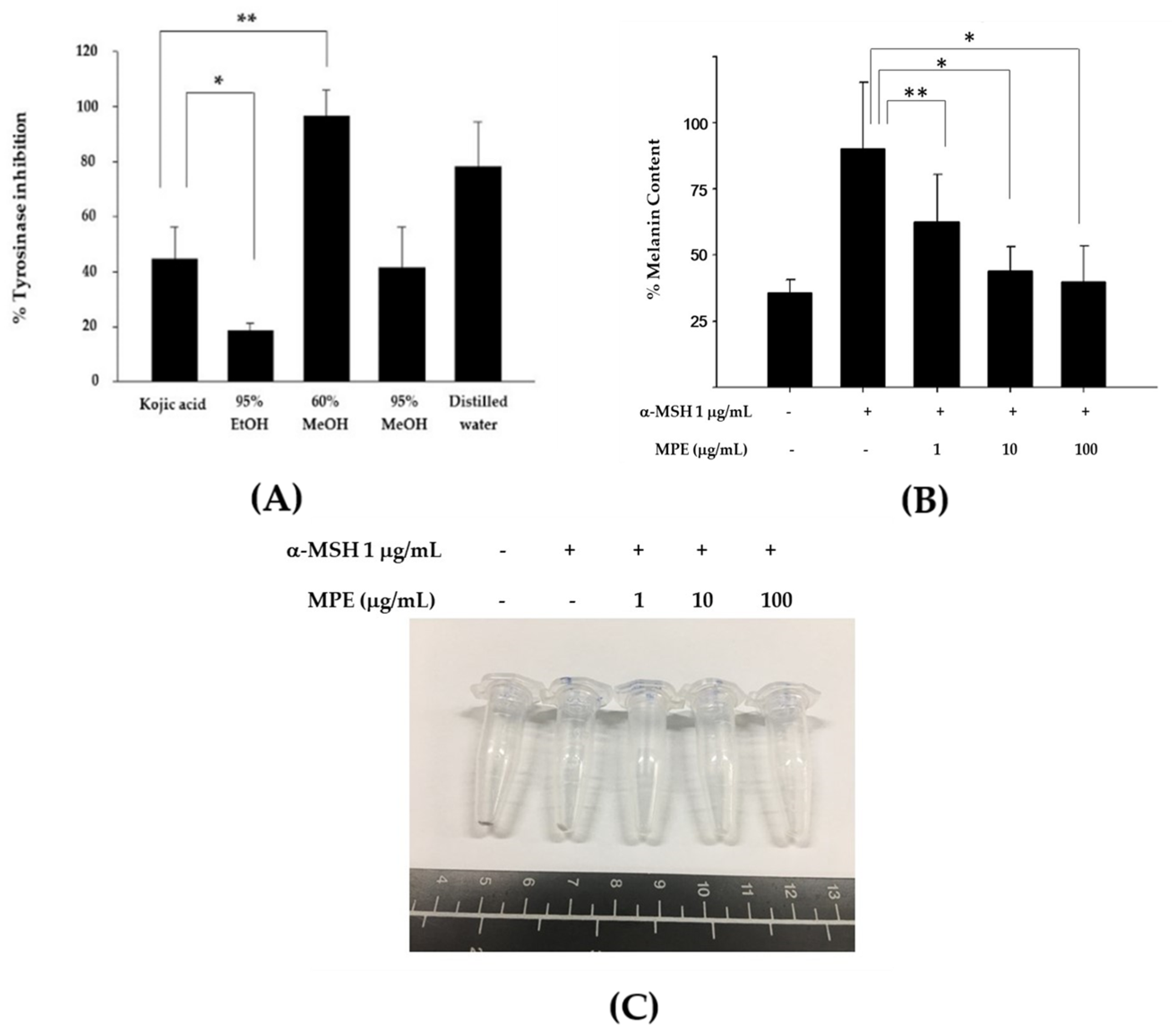{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Cell Culture
2.3. Mushroom Tyrosinase Assay
2.4. Melanin Content Assay
2.5. Protein Content Determination
2.6. Western Blot Analysis
2.7. Immunofluorescence Assay
2.8. Statistical Analysis
3. Results
3.1. MPE Diminished Mushroom Tyrosinase Activity and Melanin Production in B16F10 Mouse Melanoma Cells
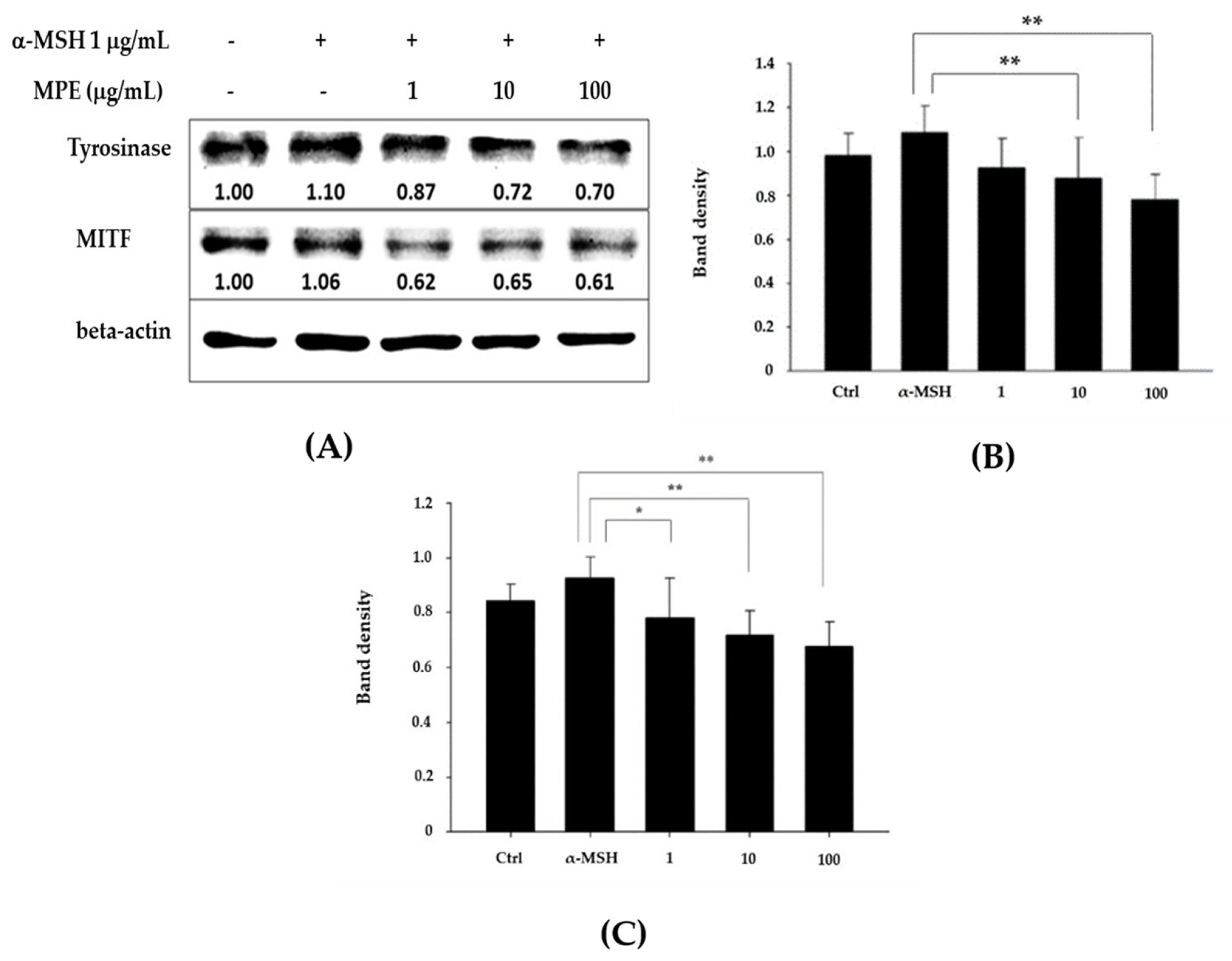
3.2. MPE Reduced Tyrosinase and MITF Protein Expression
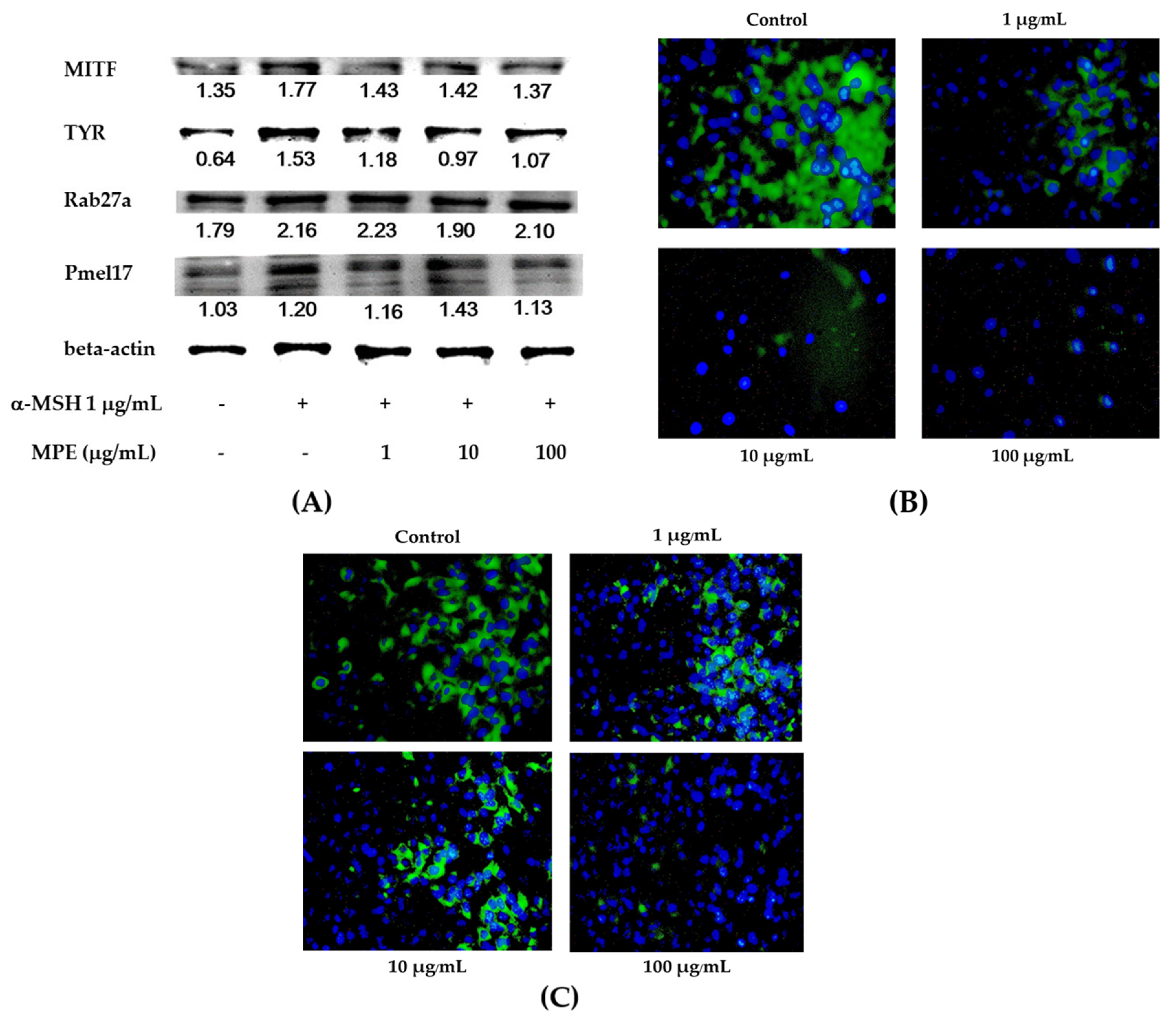
3.3. Effect of MPE on Melanogenesis Inhibition through Melanosome Transfer Process
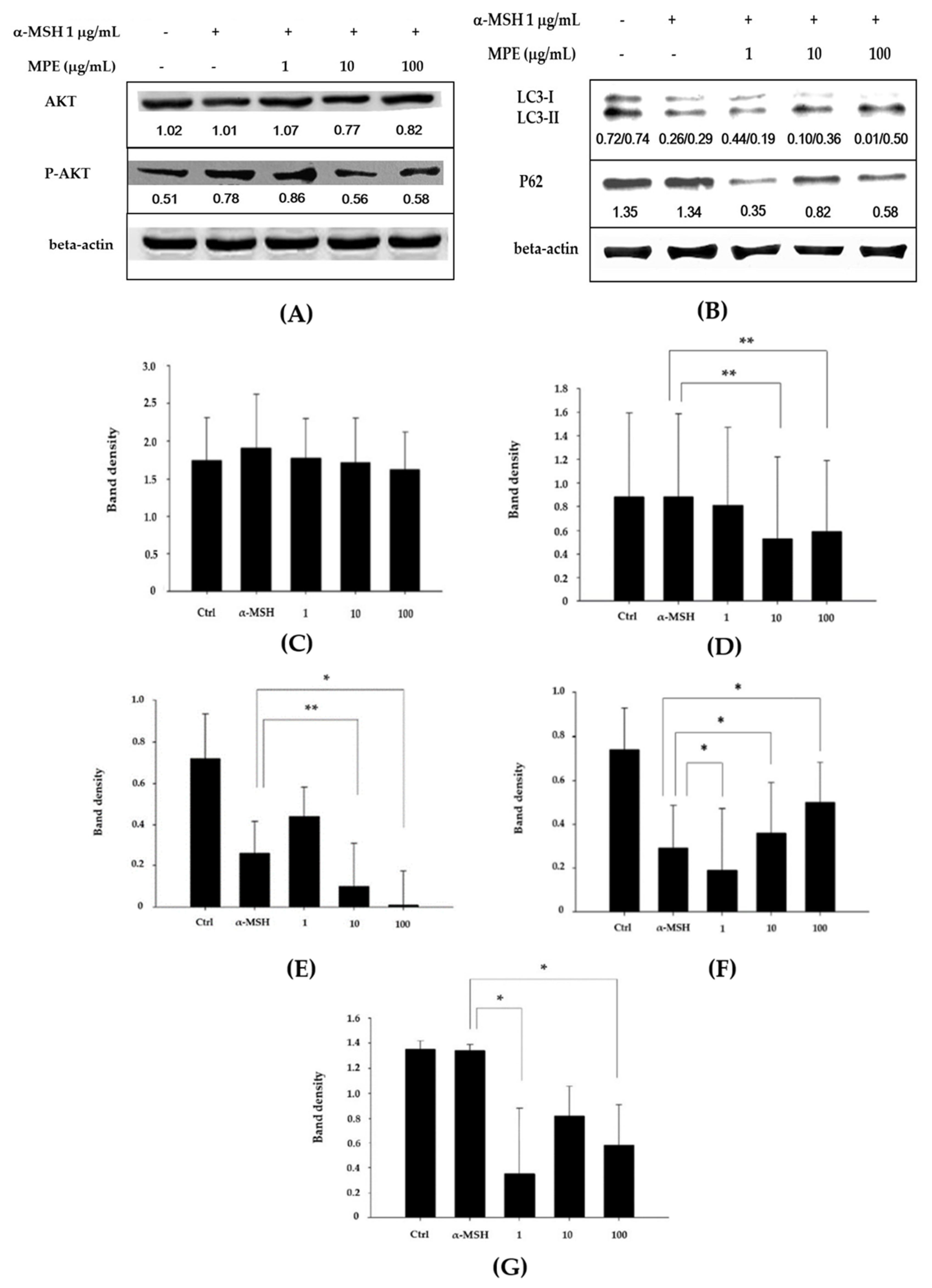
3.4. Effect of MPE on Melanogenesis Inhibition through the AKT Signaling and Autophagy Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masson Regnault, M.; Gadaud, N.; Boulinguez, S.; Tournier, E.; Lamant, L.; Gladieff, L.; Roche, H.; Guenounou, S.; Recher, C.; Sibaud, V. Chemotherapy-related reticulate hyperpigmentation: A case series and review of the literature. J. Dermatol. 2015, 231, 312–318. [Google Scholar] [CrossRef]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]
- Serre, C.; Busuttil, V.; Botto, J.M. Intrinsic and extrinsic regulation of human skin melanogenesis and pigmentation. Int. J. Cosmet. Sci. 2018, 40, 328–347. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Ren, T.; Li, Y.; Cheng, A.; Xie, W.; Xu, L.; Peng, L.; Lin, J.; Lian, L.; Diao, Y.; et al. Oleoylethanolamide inhibits α-melanocyte stimulating hormone-stimulated melanogenesis via ERK, Akt and CREB signaling pathways in B16 melanoma cells. Oncotarget 2017, 23, 56868–56879. [Google Scholar] [CrossRef] [Green Version]
- Tsao, Y.T.; Huang, Y.F.; Kuo, C.Y.; Lin, Y.C.; Chiang, W.C.; Wang, W.K.; Hsu, C.W.; Lee, C.H. Hinokitiol inhibits melanogenesis via AKT/mTOR signaling in B16F10 mouse melanoma cells. Int. J. Mol. Sci. 2016, 17, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Bioactive compounds in banana and their associated health benefits—A review. Food Chem. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Hang, T.V.; Christopher, J.S.; Quan, V.V. Phenolic compounds within banana peel and their potential uses: A review. J. Funct. Foods 2018, 40, 238–248. [Google Scholar] [CrossRef]
- Anhwange, B.A. Chemical composition of Musa sapientum (Banana) Peels. J. Food Technol. 2008, 6, 263–266. [Google Scholar]
- Pereira, A.; Maraschin, M. Banana (Musa spp.) from peel to pulp: Ethnopharmacology, source of bioactive compounds and its relevance for human health. J. Ethnopharmacol. 2015, 160, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Youryon, P.; Supapvanich, S. Physicochemical quality and antioxidant changes in ‘Leb Mue Nang’ banana fruit during ripening. Agric. Nat. Resour. 2017, 50, 47–52. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effect of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Bowman, J.P.; Deshpande, J. Overall skin tone and skin-lightening-improving effects with oral supplementation of lutein and zeaxanthin isomers: A double-blind, placebo-controlled clinical trial. Clin. Cosmet. Investig. Dermatol. 2016, 7, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passo Tsamo, C.V.; Herent, M.F.; Tomekpe, K.; Happi Emaga, T.; Quetin-Leclercq, J.; Rogez, H.; Larondelle, Y.; Andre, C. Phenolic profiling in the pulp and peel of nine plantain cultivars (Musa sp.). Food Chem. 2015, 167, 197–204. [Google Scholar] [CrossRef]
- Shen, T.; Heo, S.I.; Wang, M.H. Involvement of the p38 MAPK and ERK signaling pathway in the anti-melanogenic effect of methyl 3,5-dicaffeoyl quinate in B16F10 mouse melanoma cell. Chem. Biol. Interact. 2012, 199, 106–111. [Google Scholar] [CrossRef]
- Swangpol, S.; Somana, J.; Chattrakhom, S. Leb Mue Nang. In 108 Thai Banana Cultivars; Swangpol, S., Somana, J., Chattrakhom, S., Eds.; H.M. Queen Sirikit Park Foundation: Bangkok, Thailand, 2009; pp. 208–209. [Google Scholar]
- Phacharapiyangkul, N.; Thirapanmethee, K.; Sa-Ngiamsuntorn, K.; Panich, U.; Lee, C.H.; Chomnawang, M.T. Effect of sucrier banana peel extracts on inhibition of melanogenesis through the ERK signaling pathway. Int. J. Med. Sci. 2019, 16, 602–606. [Google Scholar] [CrossRef] [Green Version]
- Tsao, Y.T.; Kuo, C.Y.; Kuan, Y.D.; Lin, H.C.; Wu, L.H.; Lee, C.H. The extracts of astragalus membranaceus inhibit melanoenesis through the ERK signaling pathway. Int. J. Med. Sci. 2017, 14, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Yun, W.J.; Kim, E.Y.; Park, J.E.; Jo, S.Y.; Bang, S.H.; Chang, E.J.; Chang, S.E. Microtubule-associated protein light chain 3 is involved in melanogenesis via regulation of MITF expression in melanocytes. Sci. Rep. 2016, 6, 19914. [Google Scholar] [CrossRef]
- Kim, E.S.; Shin, J.H.; Seok, S.H.; Kim, J.B.; Chang, H.; Park, S.J.; Jo, Y.K.; Choi, E.S.; Park, J.S.; Yeom, M.H.; et al. Autophagy mediates anti-melanogenic activity of 3’-ODI in B16F1 melanoma cells. Biochem. Biophys. Res. Commun. 2013, 442, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Ruesgas-Ramón, M.; Figueroa-Espinoza, M.C.; Durand, E. Application of deep eutectic solvents (DES) for phenolic compounds extraction: Overview, challenges, and opportunities. J. Agric. Food Chem. 2017, 10, 3591–3601. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.E.; Levy, C. The three m’s: Melanoma, micro-phthalmia-associated transcription factor and microRNA. Pigment Cell Melanoma Res. 2011, 24, 1088–1106. [Google Scholar] [CrossRef]
- Bu, J.; Ma, P.C.; Chen, Z.Q.; Zhou, W.Q.; Fu, Y.J.; Li, L.J.; Li, C.R. Inhibition of MITF and tyrosinase by paeonol-stimulated JNK/SAPK to reduction of phosphorylated CREB. Am. J. Chin. Med. 2008, 36, 245–263. [Google Scholar] [CrossRef]
- Lee, D.H.; Ahn, S.S.; Kim, J.B.; Lim, Y.; Lee, Y.H.; Shin, S.Y. Downregulation of α-melanocyte-stimulating hormone-induced activation of the Pax3-MITF-tyrosinase axis by sorghum ethanolic extract in B16F10 melanoma cells. Int. J. Mol. Sci. 2018, 19, 1640. [Google Scholar] [CrossRef] [Green Version]
- Wasmeier, C.; Hume, A.N.; Bolasco, G.; Seabra, M.C. Melanosomes at a glance. J. Cell Sci. 2008, 121, 3995–3999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hume, A.N.; Ushakov, D.S.; Tarafder, A.K.; Ferenczi, M.A.; Seabra, M.C. Rab27a regulates the peripheral distribution of melanosomes in melanocytes. J. Cell Biol. 2001, 152, 795–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadokoro, R.; Takahashi, Y. Intercellular transfer of organelles during body pigmentation. Curr. Opin. Genet. Dev. 2017, 45, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.K.; Lin, S.T.; Chang, W.W.; Liu, L.W.; Li, T.Y.; Kuo, C.Y.; Hsieh, J.L.; Lee, C.H. Hinokitiol induces autophagy in murine breast and colorectal cancer cells. Environ. Toxicol. 2016, 31, 77–84. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Taira, N.; Yoshioka, M.; Okano, Y.; Masaki, H. Disruption of melanosome transport in melanocytes treated with theophylline causes their degradation by autophagy. Biochem. Biophy. Res. Commun. 2017, 485, 126–130. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phacharapiyangkul, N.; Thirapanmethee, K.; Sa-ngiamsuntorn, K.; Panich, U.; Lee, C.-H.; Chomnawang, M.T. The Ethanol Extract of Musa sapientum Linn. Peel Inhibits Melanogenesis through AKT Signaling Pathway. Cosmetics 2021, 8, 70. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8030070
Phacharapiyangkul N, Thirapanmethee K, Sa-ngiamsuntorn K, Panich U, Lee C-H, Chomnawang MT. The Ethanol Extract of Musa sapientum Linn. Peel Inhibits Melanogenesis through AKT Signaling Pathway. Cosmetics. 2021; 8(3):70. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8030070
Chicago/Turabian StylePhacharapiyangkul, Naphichaya, Krit Thirapanmethee, Khanit Sa-ngiamsuntorn, Uraiwan Panich, Che-Hsin Lee, and Mullika Traidej Chomnawang. 2021. "The Ethanol Extract of Musa sapientum Linn. Peel Inhibits Melanogenesis through AKT Signaling Pathway" Cosmetics 8, no. 3: 70. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8030070