
Increased Serum Thromboxane A2 and Prostacyclin but Lower Complement C3 and C4 Levels in COVID-19: Associations with Chest CT Scan Anomalies and Lowered Peripheral Oxygen Saturation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Measurements
2.3. Statistical Analysis
3. Results
3.1. Socio-Demographic Data
3.2. Differences in Biomarkers among Groups
3.3. Multivariate Differences between COVID-19 Patients and Controls
3.4. Associations of Biomarkers with Anti-SARS-CoV-2 IgG Antibodies, CCTAs, and SpO2
4. Discussion
4.1. Changes in Complement in COVID-19
4.2. Increased TxA2 and PGI2 in COVID-19
4.3. Lowered Albumin, Calcium, and Magnesium in COVID-19
4.4. Biomarkers and IgG Positivity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Tan, C.W.; Tan, J.Y.; Wong, W.H.; Cheong, M.A.; Ng, I.M.; Conceicao, E.P.; Low, J.G.H.; Ng, H.J.; Lee, L.H. Clinical and laboratory features of hypercoagulability in COVID-19 and other respiratory viral infections amongst predominantly younger adults with few comorbidities. Sci. Rep. 2021, 11, 1793. [Google Scholar] [CrossRef] [PubMed]
- Song, J.W.; Zhang, C.; Fan, X.; Meng, F.P.; Xu, Z.; Xia, P.; Cao, W.J.; Yang, T.; Dai, X.P.; Wang, S.Y.; et al. Immunological and inflammatory profiles in mild and severe cases of COVID-19. Nat. Commun. 2020, 11, 3410. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.S.; Capstick, T.; Ahmed, R.; Kow, C.S.; Mazhar, F.; Merchant, H.A.; Zaidi, S.T.R. Mortality in COVID-19 patients with acute respiratory distress syndrome and corticosteroids use: A systematic review and meta-analysis. Expert Rev. Respir. Med. 2020, 14, 1149–1163. [Google Scholar] [CrossRef]
- Amini Pouya, M.; Afshani, S.M.; Maghsoudi, A.S.; Hassani, S.; Mirnia, K. Classification of the present pharmaceutical agents based on the possible effective mechanism on the COVID-19 infection. DARU J. Fac. Pharm. Tehran. Univ. Med. Sci. 2020, 28, 745–764. [Google Scholar] [CrossRef]
- Merdji, H.; Mayeur, S.; Schenck, M.; Oulehri, W.; Clere-Jehl, R.; Cunat, S.; Herbrecht, J.E.; Janssen-Langenstein, R.; Nicolae, A.; Helms, J.; et al. Histopathological features in fatal COVID-19 acute respiratory distress syndrome. Med. Intensiva 2021, 45, 261–270. [Google Scholar] [CrossRef]
- Java, A.; Apicelli, A.J.; Liszewski, M.K.; Coler-Reilly, A.; Atkinson, J.P.; Kim, A.H.; Kulkarni, H.S. The complement system in COVID-19: Friend and foe? JCI Insight 2020, 5, e140711. [Google Scholar] [CrossRef]
- Gralinski, L.E.; Sheahan, T.P.; Morrison, T.E.; Menachery, V.D.; Jensen, K.; Leist, S.R.; Whitmore, A.; Heise, M.T.; Baric, R.S. Complement Activation Contributes to Severe Acute Respiratory Syndrome Coronavirus Pathogenesis. MBio 2018, 9, e01753-18. [Google Scholar] [CrossRef] [Green Version]
- Sarma, V.J.; Huber-Lang, M.; Ward, P.A. Complement in lung disease. Autoimmunity 2006, 39, 387–394. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [Green Version]
- Bosmann, M.; Ward, P.A. Role of C3, C5 and anaphylatoxin receptors in acute lung injury and in sepsis. Adv. Exp. Med. Biol. 2012, 946, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Ghazavi, A.; Mosayebi, G.; Keshavarzian, N.; Rabiemajd, S.; Ganji, A. Reduction of Inflammatory C3 and C4 Complement Proteins in Severe COVID-19 Patients. Preprints 2020. [Google Scholar] [CrossRef]
- Fang, S.; Wang, H.; Lu, L.; Jia, Y.; Xia, Z. Decreased complement C3 levels are associated with poor prognosis in patients with COVID-19: A retrospective cohort study. Int. Immunopharmacol. 2020, 89, 107070. [Google Scholar] [CrossRef]
- Gavriilaki, E.; Brodsky, R.A. Severe COVID-19 infection and thrombotic microangiopathy: Success does not come easily. Br. J. Haematol. 2020, 189, e227–e230. [Google Scholar] [CrossRef]
- Satyam, A.; Tsokos, M.G.; Brook, O.R.; Hecht, J.L.; Moulton, V.R.; Tsokos, G.C. Activation of classical and alternative complement pathways in the pathogenesis of lung injury in COVID-19. Clin. Immunol. 2021, 226, 108716. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.M.; Kahwash, R. Will complement inhibition be the new target in treating COVID-19–related systemic thrombosis? Circulation 2020, 141, 1739–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klok, F.; Kruip, M.; Van der Meer, N.; Arbous, M.; Gommers, D.; Kant, K.; Kaptein, F.; van Paassen, J.; Stals, M.; Huisman, M. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thromb. Res. 2020, 191, 145–147. [Google Scholar] [CrossRef]
- Amara, U.; Flierl, M.A.; Rittirsch, D.; Klos, A.; Chen, H.; Acker, B.; Bruckner, U.B.; Nilsson, B.; Gebhard, F.; Lambris, J.D.; et al. Molecular intercommunication between the complement and coagulation systems. J. Immunol. 2010, 185, 5628–5636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offermanns, S. Activation of platelet function through G protein-coupled receptors. Circ. Res. 2006, 99, 1293–1304. [Google Scholar] [CrossRef] [Green Version]
- Ruan, K.H. Advance in understanding the biosynthesis of prostacyclin and thromboxane A2 in the endoplasmic reticulum membrane via the cyclooxygenase pathway. Mini Rev. Med. Chem. 2004, 4, 639–647. [Google Scholar] [CrossRef]
- Ruan, K.-H. Thrombogenesis and Anti-Thrombogenesis in Atherosclerosis Mediated by Prostanoids through the Cyclooxygenase Pathway; Nova Science Publishers Inc.: New York, NY, USA, 2005. [Google Scholar]
- FitzGerald, G.A.; Oates, J.A.; Hawiger, J.; Maas, R.L.; Roberts, L.J., 2nd; Lawson, J.A.; Brash, A.R. Endogenous biosynthesis of prostacyclin and thromboxane and platelet function during chronic administration of aspirin in man. J. Clin. Investig. 1983, 71, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Kritas, S.K.; Frydas, I.; Younes, A.; Di Emidio, P.; Ronconi, G.; Toniato, E. IL-1 induces throboxane-A2 (TxA2) in COVID-19 causing inflammation and micro-thrombi: Inhibitory effect of the IL-1 receptor antagonist (IL-1Ra). J. Biol. Regul. Homeost. Agents 2020, 34, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.Y.; Liou, J.Y.; Wu, K.K. Prostacyclin protects vascular integrity via PPAR/14-3-3 pathway. Prostaglandins Other Lipid Mediat. 2015, 118-119, 19–27. [Google Scholar] [CrossRef]
- McAdam, B.F.; Catella-Lawson, F.; Mardini, I.A.; Kapoor, S.; Lawson, J.A.; FitzGerald, G.A. Systemic biosynthesis of prostacyclin by cyclooxygenase (COX)-2: The human pharmacology of a selective inhibitor of COX-2. Proc. Natl. Acad. Sci. USA 1999, 96, 272–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Austin, S.C.; Rocca, B.; Koller, B.H.; Coffman, T.M.; Grosser, T.; Lawson, J.A.; FitzGerald, G.A. Role of prostacyclin in the cardiovascular response to thromboxane A2. Science 2002, 296, 539–541. [Google Scholar] [CrossRef]
- Fleming, I. The factor in EDHF: Cytochrome P450 derived lipid mediators and vascular signaling. Vasc. Pharm. 2016, 86, 31–40. [Google Scholar] [CrossRef]
- Vila, L. Cyclooxygenase and 5-lipoxygenase pathways in the vessel wall: Role in atherosclerosis. Med. Res. Rev. 2004, 24, 399–424. [Google Scholar] [CrossRef]
- Manieri, N.A.; Mack, M.R.; Himmelrich, M.D.; Worthley, D.L.; Hanson, E.M.; Eckmann, L.; Wang, T.C.; Stappenbeck, T.S. Mucosally transplanted mesenchymal stem cells stimulate intestinal healing by promoting angiogenesis. J. Clin. Investig. 2015, 125, 3606–3618. [Google Scholar] [CrossRef]
- Martini, R. The compelling arguments for the need of microvascular investigation in COVID-19 critical patients. Clin. Hemorheol. Microcirc. 2020, 75, 27–34. [Google Scholar] [CrossRef]
- Al-Hakeim, H.K.; Al-Jassas, H.K.; Morris, G.; Maes, M. Increased angiotensin-converting enzyme 2, sRAGE and immune activation, but lowered calcium and magnesium in COVID-19: Association with chest CT abnormalities and lowered peripheral oxygen saturation. medRxiv 2021. [Google Scholar] [CrossRef]
- Huang, J.; Cheng, A.; Kumar, R.; Fang, Y.; Chen, G.; Zhu, Y.; Lin, S. Hypoalbuminemia predicts the outcome of COVID-19 independent of age and co-morbidity. J. Med. Virol. 2020, 92, 2152–2158. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Liu, L.; Zhao, M.; Xiao, J.; Zhao, Q. Liver impairment in COVID-19 patients: A retrospective analysis of 115 cases from a single centre in Wuhan city, China. Liver Int. Off. J. Int. Assoc. Study Liver 2020, 40, 2095–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, M.T.; Zhang, J.; Lyon, A.W.; Hemmelgarn, B.R. Derivation and internal validation of an equation for albumin-adjusted calcium. BMC Clin. Pathol. 2008, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhang, Z.; Wu, Y.; Shi, Q.; Yan, H.; Mei, N.; Tolleson, W.H.; Guo, L. Endoplasmic Reticulum Stress and Store-Operated Calcium Entry Contribute to Usnic Acid-Induced Toxicity in Hepatic Cells. Toxicol. Sci. 2015, 146, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Zhang, I.X.; Ren, J.; Vadrevu, S.; Raghavan, M.; Satin, L.S. ER stress increases store-operated Ca(2+) entry (SOCE) and augments basal insulin secretion in pancreatic beta cells. J. Biol. Chem. 2020, 295, 5685–5700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, T.S.; Liu, D.X. Coronavirus infection, ER stress, apoptosis and innate immunity. Front. Microbiol. 2014, 5, 296. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.; Athan, E.; Walder, K.; Bortolasci, C.C.; O’Neil, A.; Marx, W.; Berk, M.; Carvalho, A.F.; Maes, M.; Puri, B.K. Can endolysosomal deacidification and inhibition of autophagy prevent severe COVID-19? Life Sci. 2020, 262, 118541. [Google Scholar] [CrossRef]
- Kwee, T.C.; Kwee, R.M. Chest CT in COVID-19: What the radiologist needs to know. RadioGraphics 2020, 40, 1848–1865. [Google Scholar] [CrossRef]
- Moustafa, H.E.-D.S.; El-Seddek, M. Accurate diagnosis of covid-19 based on deep neural networks and chest x-ray images.(dept. e (electronics)). Bull. Fac. Eng. Mansoura Univ. 2020, 45, 11–15. [Google Scholar] [CrossRef]
- Marcos-Jiménez, A.; Sánchez-Alonso, S.; Alcaraz-Serna, A.; Esparcia, L.; López-Sanz, C.; Sampedro-Núñez, M.; Mateu-Albero, T.; Sánchez-Cerrillo, I.; Martínez-Fleta, P.; Gabrie, L.; et al. Deregulated cellular circuits driving immunoglobulins and complement consumption associate with the severity of COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Zinellu, A.; Mangoni, A.A. Serum complement C3 and C4 and COVID-19 severity and mortality: A systematic review and meta-analysis with meta-regression. Front. Immunol. 2021, 12, 696085. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Peffault de Latour, R.; Bergeron, A.; Lengline, E.; Dupont, T.; Marchal, A.; Galicier, L.; de Castro, N.; Bondeelle, L.; Darmon, M.; Dupin, C.; et al. Complement C5 inhibition in patients with COVID-19—A promising target? Haematologica 2020, 105, 2847–2850. [Google Scholar] [CrossRef] [PubMed]
- Holter, J.C.; Pischke, S.E.; de Boer, E.; Lind, A.; Jenum, S.; Holten, A.R.; Tonby, K.; Barratt-Due, A.; Sokolova, M.; Schjalm, C.; et al. Systemic complement activation is associated with respiratory failure in COVID-19 hospitalized patients. Proc. Natl. Acad. Sci. USA 2020, 117, 25018–25025. [Google Scholar] [CrossRef]
- Ciceri, F.; Beretta, L.; Scandroglio, A.M.; Colombo, S.; Landoni, G.; Ruggeri, A.; Peccatori, J.; D’Angelo, A.; De Cobelli, F.; Rovere-Querini, P.; et al. Microvascular COVID-19 lung vessels obstructive thromboinflammatory syndrome (MicroCLOTS): An atypical acute respiratory distress syndrome working hypothesis. Crit. Care Resusc. 2020, 22, 95–97. [Google Scholar]
- Reilly, M.; Fitzgerald, G.A. Cellular activation by thromboxane A2 and other eicosanoids. Eur. Heart J. 1993, 14 (Suppl. K), 88–93. [Google Scholar]
- Gupta, A.; Kalantar-Zadeh, K.; Reddy, S.T. Ramatroban as a Novel Immunotherapy for COVID-19. J. Mol. Genet. Med. 2020, 14, 1–9. [Google Scholar] [CrossRef]
- Wise, J. Covid-19 and thrombosis: What do we know about the risks and treatment? Br. Med. J. Publ. Group 2020, 369, 1–2. [Google Scholar] [CrossRef]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet gene expression and function in patients with COVID-19. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Bilaloglu, S.; Aphinyanaphongs, Y.; Jones, S.; Iturrate, E.; Hochman, J.; Berger, J.S. Thrombosis in Hospitalized Patients With COVID-19 in a New York City Health System. JAMA 2020, 324, 799–801. [Google Scholar] [CrossRef] [PubMed]
- Robb, C.T.; Goepp, M.; Rossi, A.G.; Yao, C. Non-steroidal anti-inflammatory drugs, prostaglandins, and COVID-19. Br. J. Pharm. 2020, 177, 4899–4920. [Google Scholar] [CrossRef] [PubMed]
- Midgett, C.; Stitham, J.; Martin, K.A.; Hwa, J. Prostacyclin receptor regulation-from transcription to trafficking. Curr. Mol. Med. 2011, 11, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kunapuli, S.P. P2Y12 receptor in platelet activation. Platelets 2011, 22, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Toki, S.; Reiss, S.; Goleniewska, K.; Moore, M.L.; FitzGerald, G.; Peebles, R.S. Prostaglandin I2 Receptor (IP) Signaling Inhibits Lung Type I Interferon Expression by RSV Infection. J. Allergy Clin. Immunol. 2013, 131, AB191. [Google Scholar] [CrossRef]
- Dorris, S.L.; Peebles, R.S. PGI2 as a regulator of inflammatory diseases. Mediat. Inflamm. 2012, 2012, 926968. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Segi-Nishida, E.; Miyachi, Y.; Narumiya, S. Prostacyclin-IP signaling and prostaglandin E2-EP2/EP4 signaling both mediate joint inflammation in mouse collagen-induced arthritis. J. Exp. Med. 2006, 203, 325–335. [Google Scholar] [CrossRef]
- Nakajima, S.; Honda, T.; Sakata, D.; Egawa, G.; Tanizaki, H.; Otsuka, A.; Moniaga, C.S.; Watanabe, T.; Miyachi, Y.; Narumiya, S. Prostaglandin I2–IP signaling promotes Th1 differentiation in a mouse model of contact hypersensitivity. J. Immunol. 2010, 184, 5595–5603. [Google Scholar] [CrossRef] [Green Version]
- Canzano, P.; Brambilla, M.; Porro, B.; Cosentino, N.; Tortorici, E.; Vicini, S.; Poggio, P.; Cascella, A.; Pengo, M.F.; Veglia, F.; et al. Platelet and Endothelial Activation as Potential Mechanisms Behind the Thrombotic Complications of COVID-19 Patients. JACC. Basic Transl. Sci. 2021, 6, 202–218. [Google Scholar] [CrossRef]
- Perico, L.; Benigni, A.; Casiraghi, F.; Ng, L.F.P.; Renia, L.; Remuzzi, G. Immunity, endothelial injury and complement-induced coagulopathy in COVID-19. Nat. Rev. Nephrol. 2021, 17, 46–64. [Google Scholar] [CrossRef]
- Magro, C.; Mulvey, J.J.; Berlin, D.; Nuovo, G.; Salvatore, S.; Harp, J.; Baxter-Stoltzfus, A.; Laurence, J. Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: A report of five cases. Transl. Res. 2020, 220, 1–13. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Teuwen, L.-A.; Geldhof, V.; Pasut, A.; Carmeliet, P. COVID-19: The vasculature unleashed. Nat. Rev. Immunol. 2020, 20, 389–391. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Golshiri, K.; Ataei Ataabadi, E.; Portilla Fernandez, E.C.; Jan Danser, A.H.; Roks, A.J.M. The importance of the nitric oxide-cGMP pathway in age-related cardiovascular disease: Focus on phosphodiesterase-1 and soluble guanylate cyclase. Basic Clin. Pharmacol. Toxicol. 2020, 127, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Maes, M. A review on the acute phase response in major depression. Rev. Neurosci. 1993, 4, 407–416. [Google Scholar] [CrossRef]
- Soeters, P.B.; Wolfe, R.R.; Shenkin, A. Hypoalbuminemia: Pathogenesis and Clinical Significance. JPEN J. Parenter. Enter. Nutr. 2019, 43, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronit, A.; Kirkegaard-Klitbo, D.M.; Dohlmann, T.L.; Lundgren, J.; Sabin, C.A.; Phillips, A.N.; Nordestgaard, B.G.; Afzal, S. Plasma Albumin and Incident Cardiovascular Disease: Results From the CGPS and an Updated Meta-Analysis. Arter. Thromb. Vasc. Biol. 2020, 40, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thachil, J.; Tang, N.; Gando, S.; Falanga, A.; Cattaneo, M.; Levi, M.; Clark, C.; Iba, T. ISTH interim guidance on recognition and management of coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Paar, M.; Rossmann, C.; Nusshold, C.; Wagner, T.; Schlagenhauf, A.; Leschnik, B.; Oettl, K.; Koestenberger, M.; Cvirn, G.; Hallstrom, S. Anticoagulant action of low, physiologic, and high albumin levels in whole blood. PLoS ONE 2017, 12, e0182997. [Google Scholar] [CrossRef]
- Yadav, H.; Kor, D.J. Platelets in the pathogenesis of acute respiratory distress syndrome. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L915–L923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, E.A.; Weyrich, A.S.; Zimmerman, G.A. Platelets in Pulmonary Immune Responses and Inflammatory Lung Diseases. Physiol. Rev. 2016, 96, 1211–1259. [Google Scholar] [CrossRef] [PubMed]
- Koupenova, M.; Clancy, L.; Corkrey, H.A.; Freedman, J.E. Circulating Platelets as Mediators of Immunity, Inflammation, and Thrombosis. Circ. Res. 2018, 122, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.-L.; Liu, Z.-H.; Cao, C.-F.; Liu, M.-K.; Liao, J.; Zou, J.-B.; Kong, L.-X.; Wan, K.-Q.; Zhang, J.; Wang, Q.-B.; et al. COVID-19 Myocarditis and Severity Factors: An Adult Cohort Study. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Ma, X.; Wu, J.; Han, J.; Zheng, Z.; Duan, H.; Liu, Q.; Wu, C.; Dong, Y.; Dong, L. Low serum calcium and phosphorus and their clinical performance in detecting COVID-19 patients. J. Med. Virol. 2021, 93, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Torres, B.; Alcubilla, P.; Gonzalez-Cordon, A.; Inciarte, A.; Chumbita, M.; Cardozo, C.; Meira, F.; Gimenez, M.; de Hollanda, A.; Soriano, A.; et al. Impact of low serum calcium at hospital admission on SARS-CoV-2 infection outcome. Int. J. Infect. Dis. 2021, 104, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Di Filippo, L.; Formenti, A.M.; Rovere-Querini, P.; Carlucci, M.; Conte, C.; Ciceri, F.; Zangrillo, A.; Giustina, A. Hypocalcemia is highly prevalent and predicts hospitalization in patients with COVID-19. Endocrine 2020, 68, 475–478. [Google Scholar] [CrossRef]
- Guo, L.; Ren, L.; Yang, S.; Xiao, M.; Chang, D.; Yang, F.; Dela Cruz, C.S.; Wang, Y.; Wu, C.; Xiao, Y.; et al. Profiling Early Humoral Response to Diagnose Novel Coronavirus Disease (COVID-19). Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, 778–785. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Mehta, H.; Drevets, D.A.; Coggeshall, K.M. IL-6 increases B-cell IgG production in a feed-forward proinflammatory mechanism to skew hematopoiesis and elevate myeloid production. Blood 2010, 115, 4699–4706. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.H.; Fischer, E.M.; Leslie, R.G. The role of complement in the acquired immune response. Immunology 2000, 100, 4–12. [Google Scholar] [CrossRef]
- Zeng, F.; Dai, C.; Cai, P.; Wang, J.; Xu, L.; Li, J.; Hu, G.; Wang, Z.; Zheng, F.; Wang, L. A comparison study of SARS-CoV-2 IgG antibody between male and female COVID-19 patients: A possible reason underlying different outcome between sex. J. Med. Virol. 2020, 92, 2050–2054. [Google Scholar] [CrossRef]


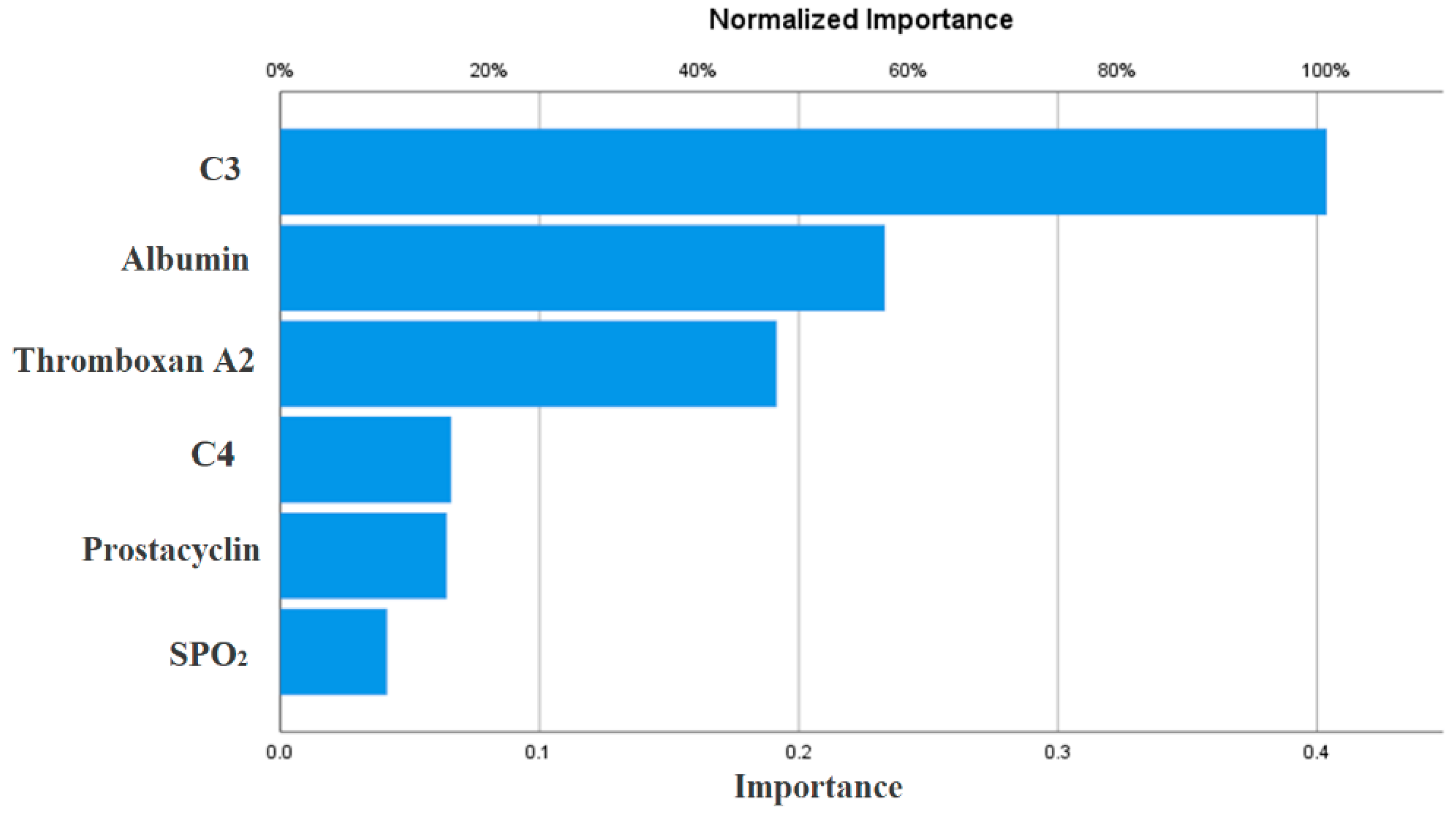{kind=link}
| Variables | HC (n = 30) | COVID-19 (n = 60) | F/FEPT/χ2 | df | p |
|---|---|---|---|---|---|
| Age (years) | 40.1 ± 8.8 | 41.0 ± 10.2 | 0.17 | 1/88 | 0.681 |
| BMI (kg/m2) | 26.05 ± 4.02 | 27.07 ± 3.62 | 1.50 | 1/88 | 0.225 |
| Sex (Female/Male) | 6/24 | 17/43 | 0.73 | 1 | 0.393 |
| Urban/Rural | 28/2 | 52/8 | 0.486 | ||
| Single/married | 10/20 | 17/43 | 0.24 | 1 | 0.626 |
| TUD (No/Yes) | 20/10 | 39/21 | 0.03 | 1 | 0.875 |
| Employment (No/Yes) | 9/21 | 21/39 | 0.23 | 1 | 0.635 |
| Education (years) | 10.7 ± 3.2 | 9.8 ± 4.0 | 1.07 | 1/88 | 0.303 |
| Admission room / ICU / RCU | 35/16/9 | ||||
| Zinc (No/Yes) | 23/7 | 1/59 | 57.53 | 1 | <0.001 |
| Vitamin D (No/Yes) | 27/3 | 0/60 | 77.14 | 1 | <0.001 |
| Vitamin C (No/Yes) | 23/7 | 0/60 | 61.79 | 1 | <0.001 |
| Dexamethasone (No/Yes) | 9/51 | ||||
| Azithromycin (No/Yes) | 25/35 | ||||
| Enoxaparin (No/Yes) | 28/32 | ||||
| O2 therapy (No/Yes) | 0/60 | ||||
| Bromhexine (No/Yes) | 21/39 | ||||
| Paracetamol (No/Yes) | 27/3 | 0/60 | 77.14 | 1 | <0.001 |
| Omeprazole (No/Yes) | 23/37 | ||||
| Ceftriaxone (No/Yes) | 35/25 | ||||
| IgG (Positive / Negative) | 36/24 |
| Biomarkers | HC A (n = 30) | COVID-CCTA B (n = 15) | COVID + CCTA C (n = 45) | F | df | p |
|---|---|---|---|---|---|---|
| SPO2 | 98.53 ± 0.68 C | 97.13 ± 0.74 C | 92.62 ± 3.40 A,B | 56.37 | 2/87 | <0.001 |
| Albumin g/L | 43.10 ± 3.11 B,C | 33.47 ± 5.08 A,C | 29.71 ± 3.79 A,B | 111.12 | 2/87 | <0.001 |
| Magnesium mM | 0.933 ± 0.102 C | 1.018 ± 0.223 | 1.010 ± 0.172 A | 2.42 | 2/87 | 0.005 |
| Calcium mM | 2.264 ± 0.096 B,C | 2.089 ± 0.148 A | 2.021 ± 0.128 A | 36.02 | 2/87 | <0.001 |
| Thromboxane A2 pg/mL | 236.4 ± 64.6 B,C | 337.9 ± 64.1 A | 366.0 ± 98.7 A | 22.17 | 2/87 | <0.001 |
| Prostacyclin pg/mL | 104.7 ± 36.9 B,C | 153.6 ± 45.4 A,C | 188.7 ± 45.0 A,B | 35.00 | 2/87 | <0.001 |
| Complement C3 mg/L | 872.1 ± 285.5 B, C | 346.7 ± 134.0 A | 302.9 ± 88.2 A | 92.00 | 2/87 | <0.001 |
| Complement C4 mg/L | 381.7 ± 138.7 B, C | 266.3 ± 110.0 A | 273.1 ± 87.3 A | 9.98 | 2/87 | <0.001 |
| Model | COVID-19 vs. HC | |
|---|---|---|
| Input Layer | No. of units | 6 |
| Rescaling method | Normalized | |
| Hidden layers | No. of hidden layers | 1 |
| No. of units in hidden layer 1 | 4 | |
| Activation Function | Hyperbolic tangent | |
| Output layer | Dependent variables | COVID-19 vs. HC |
| Number of units | 2 | |
| Activation function | Identity | |
| Error function | Sum of squares | |
| Training | Sum of squares error term | 0.420 |
| Incorrect or relative error % | 0.0% | |
| Prediction (sens, spec) | 100%, 100% | |
| Testing | Sum of Squares error | 0.252 |
| Incorrect or relative error % | 0.0% | |
| Prediction (sens spec) | 100%, 100% | |
| AUC ROC | 1.00 | |
| Holdout | Incorrect or relative error % | 0.0% |
| Prediction (sens, spec) | 100%, 100% |
| Biomarkers | Negative IgG n = 24 | Positive IgG n = 36 | F | df | p |
|---|---|---|---|---|---|
| SpO2 | 92.25 ± 4.31 | 94.75 ± 2.56 | 7.97 | 1/58 | 0.007 |
| Albumin g/L | 29.83 ± 4.37 | 31.19 ± 4.43 | 1.37 | 1/58 | 0.246 |
| Magnesium mM | 1.017 ± 0.201 | 1.009 ± 0.174 | 0.02 | 1/58 | 0.880 |
| Calcium mM | 2.012 ± 0.132 | 2.056 ± 0.137 | 1.54 | 1/58 | 0.219 |
| Thromboxane A2 pg/mL | 306.3 ± 57.6 | 394.1 ± 93.8 | 16.78 | 1/58 | <0.001 |
| Prostacyclin pg/mL | 160.2 ± 40.9 | 193.1 ± 47.2 | 7.78 | 1/58 | 0.007 |
| Complement C3 mg/L | 329.0 ± 120.2 | 303.7 ± 88.4 | 0.88 | 1/58 | 0.351 |
| Complement C4 mg/L | 227.8 ± 81.1 | 300.4 ± 89.2 | 10.23 | 1/58 | 0.002 |
| Biomarkers | No CCTAs n = 15 | CCTAs n = 45 | F | df | p |
| SpO2 | 97.13 ± 0.74 | 92.62 ± 3.40 | 25.94 | 1/58 | <0.001 |
| Albumin g/L | 33.47 ± 5.08 | 29.71 ± 3.79 | 9.25 | 1/58 | 0.004 |
| Magnesium mM | 1.0178 ± 0.233 | 1.010 ± 0.172 | 0.02 | 1/58 | 0.895 |
| Calcium mM | 2.089 ± 0.147 | 2.038 ± 0.135 | 2.94 | 1/58 | 0.092 |
| Thromboxane A2 pg/mL * | 337.9 ± 64.1 | 366.0 ± 98.7 | 1.09 | 1/58 | 0.301 |
| Prostacyclin pg/mL | 153.6 ± 45.4 | 188.7 ± 45.0 | 6.81 | 1/58 | 0.011 |
| Complement C3 mg/L ** | 346.7 ± 134.0 | 302.9 ± 88.2 | 1.92 | 1/58 | 0.171 |
| Complement C4 mg/L | 266.3 ± 110.0 | 273.4 ± 87.3 | 0.59 | 1/58 | 0.809 |
| Biomarkers | SpO2 | Thromboxane A2 | Prostacyclin |
|---|---|---|---|
| Thromboxane A2 | −0.362 (0.001) | ||
| Prostacyclin | −0.380 (0.001) | 0.539 (0.002) | |
| Complement C3 | 0.598 (0.001) | −0.544 (0.002) | −0.593 (0.001) |
| Complement C4 | 0.355 (0.001) | 0.028 (0.800) | −0.014 (0.9) |
| Albumin | 0.617 (0.001) | −0.549 (0.002) | −0.572 (0.001) |
| Magnesium | −0.208 (0.055) | 0.178 (0.115) | 0.308 (0.005) |
| Calcium | 0.490 (0.001) | −0.379 (0.002) | −0.496 (0.001) |
| Dependent Variables | Explanatory Variables | β | t | p | F Model | df | p | R2 |
|---|---|---|---|---|---|---|---|---|
| #1. LnTxA2 | Model | 28.128 | 2/87 | <0.001 | 0.393 | |||
| Albumin | −0.387 | −3.923 | <0.001 | |||||
| Prostacyclin | 0.328 | 3.319 | 0.001 | |||||
| #2. LnTxA2 | Model | 20.779 | 3/86 | <0.001 | 0.420 | |||
| sqrC3 | −0.525 | −4.471 | <0.001 | |||||
| C4 | 0.241 | 2.498 | 0.014 | |||||
| Prostacyclin | 0.227 | 2.118 | 0.037 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Hakeim, H.K.; Al-Hamami, S.A.; Almulla, A.F.; Maes, M. Increased Serum Thromboxane A2 and Prostacyclin but Lower Complement C3 and C4 Levels in COVID-19: Associations with Chest CT Scan Anomalies and Lowered Peripheral Oxygen Saturation. COVID 2021, 1, 489-502. https://0-doi-org.brum.beds.ac.uk/10.3390/covid1020042
Al-Hakeim HK, Al-Hamami SA, Almulla AF, Maes M. Increased Serum Thromboxane A2 and Prostacyclin but Lower Complement C3 and C4 Levels in COVID-19: Associations with Chest CT Scan Anomalies and Lowered Peripheral Oxygen Saturation. COVID. 2021; 1(2):489-502. https://0-doi-org.brum.beds.ac.uk/10.3390/covid1020042
Chicago/Turabian StyleAl-Hakeim, Hussein Kadhem, Shaymaa Ali Al-Hamami, Abbas F. Almulla, and Michael Maes. 2021. "Increased Serum Thromboxane A2 and Prostacyclin but Lower Complement C3 and C4 Levels in COVID-19: Associations with Chest CT Scan Anomalies and Lowered Peripheral Oxygen Saturation" COVID 1, no. 2: 489-502. https://0-doi-org.brum.beds.ac.uk/10.3390/covid1020042