Reciprocal Role of Salamanders in Aquatic Energy Flow Pathways


Área de Biodiversidad y Conservación, Departamento de Biología y Geología, Física y Química Inorgánica, Universidad Rey Juan Carlos, Móstoles, 28933 Madrid, Spain
Diversity 2020, 12(1), 32; https://0-doi-org.brum.beds.ac.uk/10.3390/d12010032
Submission received: 19 November 2019
/
Revised: 13 January 2020
/
Accepted: 15 January 2020
/
Published: 17 January 2020
(This article belongs to the Special Issue The Ecological Role of Salamanders as Predators and Prey)
Abstract
:Many species of salamanders (newts and salamanders per se) have a pivotal role in energy flow pathways as they include individuals functioning as prey, competitors, and predators. Here, I synthesize historic and contemporary research on the reciprocal ecological role of salamanders as predators and prey in aquatic systems. Salamanders are a keystone in ecosystem functioning through a combination of top–down control, energy transfer, nutrient cycling processes, and carbon retention. The aquatic developmental stages of salamanders are able to feed on a wide variety of invertebrate prey captured close to the bottom as well as on small conspecifics (cannibalism) or other sympatric species, but can also consume terrestrial invertebrates on the water surface. This capacity to consume allochthonous resources (terrestrial invertebrates) highlights the key role of salamanders as couplers of terrestrial and aquatic ecosystems (i.e., aquatic–terrestrial linkages). Salamanders are also an important food resource for other vertebrates such as fish, snakes, and mammals, covering the energy demands of these species at higher trophic levels. This study emphasizes the ecological significance of salamanders in aquatic systems as central players in energy flow pathways, enabling energy mobility among trophic levels (i.e., vertical energy flow) and between freshwater and terrestrial habitats (i.e., lateral energy flow).
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Ever since the concept of energy transfer was introduced in the mid-19th century [1,2], much attention has been paid to exploring predator–prey interactions and energy flow pathways to understand ecosystem complexity and functioning e.g., [3,4,5]. From a theoretical standpoint, every organism is a potential source of energy for successive categories of consumers [2]. In aquatic food webs, energy and nutrients flow from basal to higher trophic levels (i.e., bottom–up flow to consumers), but also energy and nutrients can be transferred from terrestrial to aquatic systems and vice versa [6,7,8]. The structure and functioning of aquatic food webs and energy flows are mediated through top–down (i.e., consumers/predators can regulate the abundance of their prey at lower trophic levels) and bottom–up (i.e., the availability of resources can limit the abundance of consumers/predators in the trophic level above them) regulations [8,9]. Many species of newts and salamanders (henceforth termed “salamanders”) can impact invertebrate populations at successively lower trophic levels (top–down control), driving trophic cascades [10,11,12], but they also can be an important energy budget for higher trophic levels (i.e., apex predators such as fish, snakes, and mammals) through bottom–up energy flow pathways [3,13]. Thus, it is reasonable to posit that salamanders can display a pivotal role in aquatic energy flow pathways as these species include individuals functioning as prey, competitors, and predators in the aquatic food webs.
The feeding habits of aquatic developmental stages of salamanders are well documented, being able to feed on a wide variety of invertebrate prey captured close to the bottom as well as terrestrial invertebrates on the water surface [14,15,16,17,18]. In general, salamanders have been considered as generalist–opportunist foragers, with remarkable ontogenetic dietary shifts, and their diets reflect the relative abundance of prey in the environment [14,18,19,20]. In addition, salamanders have been widely used as model organisms to test key ecological questions related to, for example, top–down trophic cascades, nutrient cycling processes, and carbon retention in ecosystems e.g., [3,12,21,22]. However, little attention has been given to exploring the reciprocal role of salamanders (both prey and predators) in aquatic energy flow pathways. Here, I provide a summary review on the ecological role of salamanders as predators and prey in aquatic systems, addressing the knowledge gaps and highlighting promising areas for future research.
2. Salamanders as Predators
2.1. Diet Description
Primary dietary components for salamanders depend upon the literature consulted. For example, Regester et al. [23] concluded that most of the interspecific dietary variations among salamander larvae assemblages are attributed to copepods (Cyclopidae) and three families of aquatic insects (Chironomidae, Chaoboridae, and Culicidae). By contrast, Sánchez-Hernández et al. [18] observed that the diets of different developmental stages of salamanders (larva, juvenile, and adult) primarily consisted of aquatic invertebrates (mainly Plecoptera, Diptera, and Ephemeroptera). Thus, salamanders in aquatic systems feed mainly on invertebrates captured close to the bottom, such as Ostracoda, Cladocera, Copepoda, Chironomidae, Asellus, or trichopteran larvae, and can also consume prey items on the water surface [14,15,17,19,24,25]. In addition, some salamanders such as, for example, the Pacific giant salamander (Dicaptodon tenebrosus), can consume juvenile fish [26]. The two-toed amphiuma (Amphiuma means) was also observed to prey on juvenile green sunfish (Lepomis cyanellus) [27]. The consumption of conspecifics (cannibalism) or other sympatric species is often observed in larval salamanders [27,28,29]. Overall, salamander species can adapt their diets in response to spatial and temporal variation in prey availability [30,31,32].
Since salamanders are gape-limited predators, their feeding is closely related to prey characteristics, such as prey size [33,34,35]. Therefore, it is thought that prey items with small size, high body flexibility, without hard external structures (cases/shells), or with weak agility may be easy to capture and handle [36] according to the optimal foraging theory (OFT) [37,38,39]. Salamanders are able to consume highly mobile (e.g., Baetidae) and less mobile (e.g., Limnephilidae and gastropods) prey [14], although they may also accidentally consume immobile items such as vegetal fragments [40]. Nevertheless, some immobile items, such as eggs, are taken deliberately, and most researchers consider eggs a high-energy source for amphibians [41,42]. Thus, the view of diet description in terms of prey traits can be an important feature to generalize concepts across multiple systems or species. In this regard, prey trait analysis has been proposed as a functional approach to understand mechanisms involved in predator–prey relationships [36,43,44], and consequently, it may be useful for understanding prey-handling efficiency, interspecific interactions, and the mechanisms that determine food partitioning among species [36,45]. It is important to keep in mind that the feeding behavior and prey-handling efficiency of salamanders have been mainly studied in laboratory conditions [46,47,48], and thus future empirical studies attempting to integrate prey characteristics (e.g., prey traits or prey guild members) with diet descriptions will help to understand feeding and the key role of salamanders as predators.
Salamanders normally undergo a change in diet composition during ontogeny as well as across seasons e.g., [14,16,18]. Metamorphs (i.e., individuals already metamorphosed) rely highly on terrestrial invertebrates, whereas paedomorphs (i.e., individuals with larval traits) prefer planktonic resources [16]. In fact, larval salamanders feed mainly on aquatic invertebrates, whereas adults usually eat more terrestrial invertebrates on the water surface or even forage in terrestrial environments [14,15,30,49,50]. Some species, such as the Pyrenean newt (Euproctus asper), can undergo discrete ontogenetic dietary shifts with no major ontogenetic shifts in prey-type consumption but shifts in maximum prey width and terrestrial invertebrate consumption [18]. Most salamander species, on the other hand, exhibit remarkable ontogenetic dietary shifts associated with metamorphosis and concomitant shifts in habitat use (between freshwater and terrestrial environments) [14,15,16,30,49,50]. Thus, ontogenetic dietary shifts can be explained through shifts in habitat utilization for feeding and body size (i.e., processes scaling with mouth size). First, terrestrial invertebrates are more frequently consumed by metamorphosed individuals, which is linked to their capacity to forage in terrestrial habitats and, second, individuals within a population can share the same prey categories but partitioning of the food resources can occur at prey-size level, in line with limitations imposed by mouth gape.
The diets of salamanders can change across seasons regardless of geographic area (e.g., Europe or America) e.g., [16,51]. Lunghi et al. [52] demonstrated that prey size and richness differed significantly between seasons (in both cases, higher in autumn than in spring). An illustrative example of changes in diet composition across seasons can be found in Vignoli [16], who observed that feeding intensity (number of prey), niche breadth, and diet composition changed noticeably across seasons. More specifically, the dominant prey of paedomorphs changed between summer (planktonic crustaceans) and autumn (terrestrial arthropods); but they remained similar for metamorphs (terrestrial arthropods; Homoptera in summer and aphids in autumn) [16]. Thus, seasonal changes in diet composition can be discrete in some cases, as highlighted for ontogenetic dietary shifts. By contrast, high similarity has been found in the Pyrenean newts, with the same dietary components (Diptera and Plecoptera) dominant in all seasons [18]. However, a distinct use of prey taxa and size was observed over the seasons, as juvenile individuals tended to consume fewer Diptera and Plecoptera in spring and autumn, respectively [18]. One of the main criticisms of the current knowledge of the trophic ecology of salamanders is that many studies have focused on only a single season e.g., [17,19,25], despite ecological processes, such as predator–prey interactions, usually having marked seasonality.
Knowledge of the niche use of salamanders has become pivotal to understanding growth and survival through predation and competition. Although many studies rely on a single model organism, the theory behind niche use needs to be set in a broad ecological framework that includes a delineation of general patterns in the feeding of salamanders. In this regard, this review demonstrates that dietary patterns can be common among salamander species. These patterns include larval individuals preying on aquatic taxa that are easy to capture and handle, such as zooplankton in lentic systems and aquatic insects (Diptera, Ephemeroptera, and Plecoptera) in lotic systems, but there is a greater dependence on terrestrial invertebrates after metamorphosis. As already pointed out, the diet composition of salamanders can be explained by site-specific prey community structures and intrinsic factors related to body size (i.e., processes scaling with mouth size). However, it is still unexplored if the diet composition of salamanders follows broadscale patterns and whether environmental factors related to biogeography can be important for determining global patterns in the feeding of salamander species, as previously demonstrated in other vertebrate taxa [53,54]. This represents a promising avenue for future research that needs to be explored in order to accept or refute the view that the diet composition of salamanders can converge across multiple systems or species.
2.2. Top–Down Control
Salamanders provide direct and indirect biotic control of species diversity and ecosystem processes along grazer and detritus pathways [7]. Salamanders may act as top predators in systems without fish and semi-aquatic vertebrates (e.g., snakes and mammals), such as lentic systems (lakes and ponds) and headwater streams where apex predators are absent e.g., [55,56,57,58]. For example, Wissinger et al. [57] highlighted the effects of salamander predation on the invertebrate communities of subalpine wetlands, showing a key influence of salamanders on the composition of benthic and planktonic assemblages. Urban [59] provided a good example about the understanding of the functioning and top–down regulations in salamander assemblages. That is, intermediate consumers (spotted salamanders, Ambystoma maculatum) can exacerbate prey biomass declines associated with apex predation (marbled salamander, Ambystoma opacum), but buffer the top–down effects of marbled salamander predation on prey diversity [59]. Thus, the strength of top–down effects via salamanders can vary with the structure of the salamander assemblage. In some scenarios, salamander-mediated top–down effects can be weak, because interactions between apex predators and intermediate consumers are very strong (i.e., they select for compensatory evolutionary responses in other species) instead of classical top–down considerations (i.e., apex predators have remarkable effects on lower trophic levels) [59]. Atlas and Palen [60] showed theoretically (using a multitrophic modeling framework) that prey vulnerability (i.e., vulnerable versus armored aquatic invertebrates) can limit the top–down effect of the Pacific giant salamander. That is, the absence of armored aquatic invertebrates (e.g., Order Trichoptera and Coleoptera) exacerbates top–down trophic cascades (reducing insect emergence, but increasing algal and detrital biomass) [60]. This conclusion is supported by the above-mentioned gape limitations and prey-handling efficiency of salamanders [36]; thus, armored prey items with hard external structures (cases/shells) can limit salamander-mediated top–down effects, because those prey items are difficult to capture and handle. However, empirical research under controlled (e.g., mesocosms) or natural conditions is required for this to be accepted as a general theory. The top–down model with salamanders as top predators exemplifies a case where salamander predation is the strongest driver of changes in invertebrate and primary producers (i.e., salamanders suppress aquatic invertebrates, which releases freshwater macrophytes, algae, and detritus from herbivory and detritivory, respectively). These top–down effects of salamanders on ecosystem functions (animal community, nutrient dynamics, and primary production) have also been found in terrestrial systems e.g., [61], which underlines the global importance of salamander species on top–down control. However, it should be kept in mind that some studies have reported no evidence of trophic cascades with larval salamanders [62,63] and weak or inconclusive top–down effects of salamanders on invertebrate communities [64,65,66]. This highlights that salamanders do not always drive top–down effects, but they are definitively important to understand food-web dynamics through predator–prey interactions.
3. Salamanders as Prey
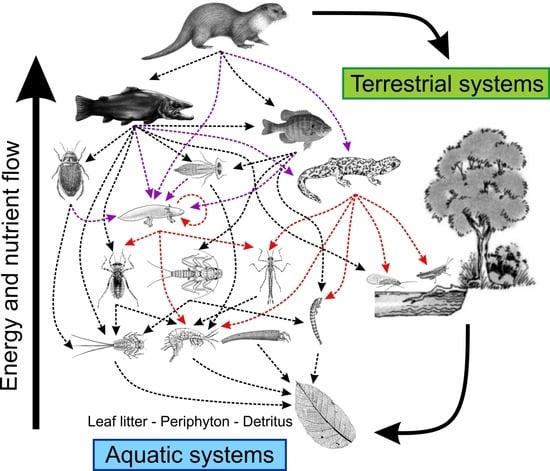
Salamanders are profitable food resources for aquatic apex predators (e.g., snake, otter, and fish species), representing a key taxon to understand energy and nutrient transfer from invertebrates up to higher trophic levels (Figure 1). Preston and Johnson [67] highlighted the key importance of amphibians as food resources for apex predators, which in turn is key for the understanding of apex predator distributions. Therefore, declines of amphibian species may potentially drive negative consequences for apex predators that prey on amphibians [67]. Jobe et al. [68] recently listed 69 salamander species consumed by 89 predators, with snakes being the most frequently reported predator (35% of predations reported), followed by salamanders (24%) and birds (16%). This list would increase as more research becomes available.
3.1. Consumption of Salamanders in Higher Trophic Levels
Although many salamander species develop defensive strategies (i.e., toxic secretions and cryptic and aposematic colorations) to deter predators, the consumption of salamanders by other vertebrates at higher trophic levels has been reported widely e.g., [69,70,71,72,73]. Salamanders provide food for birds and snakes e.g., [68,69,74]. For example, Preston and Johnson [67] found amphibians in most (93% of frequency of occurrence) of the studied aquatic gartersnakes (Thamnophis atratus) in California. Another example of predation of salamanders can be found in Escoriza and Hassine [75], who documented the first record of the viperine snake (Natrix maura) preying on the endemic Edough newt (Pleurodeles poireti) in northeastern Algeria. Willson and Winne [76] observed that the occurrence of the mole salamander (Ambystoma talpoideum) accounted for the majority (90%) of the diet composition of two snake species inhabiting the wetlands of South Carolina. Indeed, several researchers accept the view that aquatic salamanders are optimal prey for aquatic snakes because of their morphology traits (i.e., salamanders show elongated morphology), which enables snakes to eat a large number of salamanders with little effects on crawling speed [77].
The consumption of salamanders by mustelid species has also been reported widely e.g., [72,73,78,79]. Smiroldo et al. [80] recently reviewed the amphibians in the diet of the Eurasian otter (Lutra lutra) and, in the case of salamanders, the list included eight species (see Table 2 in the work by Smiroldo et al.). However, Jobe et al. [68] only listed one salamander species (great crested newt Triturus cristatus) as prey for Eurasian otter (see Appendix A in the work by Jobe et al.). This calls for a better compilation of information to create lists of predators that are capable (i.e., with published records) of feeding upon salamanders. Clavero et al. [81] noticed important seasonal effects in the consumption of amphibians by Eurasian otters, with the highest prevalence during the spawning periods of most amphibian species (late winter–spring). Novais et al. [82] also noticed that amphibians are the most frequent food resource in spring, but fish are the most frequent food resource in autumn, winter, and summer. Parry et al. [79] found that the palmate newt (Lissotriton helveticus) remains in Eurasian otter spraints in all months except for February and March. Cogălniceanu et al. [83] observed that Eurasian otters use temporary ponds with high concentrations of amphibians to consume vulnerable prey such as ribbed newts (Pleurodeles waltl). These authors also found that Eurasian otters consumed only the soft organs through an incision in the upper part of the thorax [83]. This is most likely because many salamanders produce tetrodotoxin, which is a powerful neurotoxin and anti-predatory skin secretion [84]. Bringsøe and Nørgaard [73] identified the predation of the great crested newt by the Eurasian otter in Denmark. Therefore, predators (here Eurasian otters) can overcome the defensive strategies (i.e., toxic secretions and cryptic and aposematic colorations) of salamanders. Additionally, several researchers support the notion that amphibian consumption by Eurasian otters is inversely related to fish availability (i.e., fish biomass) [85]. Thus, it is possible that spatial and temporal variations in fish availability determine predation on salamanders by mustelid species.
The consumption of salamanders by fish is also well known [54,86,87,88], with much research focusing on the effect of fish introductions on amphibian assemblages e.g., [89,90,91,92]. Overall, amphibian species richness is significantly lower in aquatic systems where fish have been introduced [93], demonstrating the importance of predatory fish species as consumers of salamanders. However, not all amphibian species show the same response to fish introductions e.g., [91,93]. For example, Orizaola and Braña [91] found no effect of salmonid presence on two widespread anuran species (common toad Bufo bufo and common midwife toad Alytes obstetricans), but they did find a negative impact on newt species (palmate newt, Alpine newt Triturus alpestris, and marbled newt Triturus Marmoratus) as well as on an anuran species (European common frog Rana temporaria). It should be kept in mind that salamanders usually represent a small portion of fish stomach contents [54,94,95]. Taking the brown trout (Salmo trutta) as an example, the primary dietary components are commonly Ephemeroptera, Diptera, Trichoptera, Plecoptera, and surface prey (terrestrial arthropods and emerged aquatic insects), with salamander remains recorded only occasionally [54].
The above examples illustrate the importance of salamanders as prey for many vertebrate species. Thus, salamanders are central players in energy and nutrient flow pathways from invertebrates up to higher trophic levels along multiple systems. Knowing the potential consumers of salamanders is integral to understanding the structure and function of communities and ecosystems through predator–prey interactions. This is key to improving our ability to predict the effects of animal introductions or changes in species distributions due to climate change on ecological processes that function at the individual, population, and community levels. From another point of view, Sánchez-Hernández et al. [54] observed an inverse association between mean annual temperature and the global dietary contribution of salamanders in brown trout. Adrián and Delibes [96] concluded that the frequency of occurrence of amphibians in the feces of Eurasian otters tends to decrease with increasing latitude. By contrast, Clavero et al. [53] observed only small differences between Mediterranean and temperate locations in the consumption of amphibians by Eurasian otters. This is seemingly a contradictory standpoint regarding studies in favor of geographical variation in the consumption of salamanders. It is possible that predation upon salamanders follows geographical patterns, representing a particularly promising area for future research. In this regard, the direction of future research could be toward the link between geographical variations and classical observations of latitudinal diversity gradients in amphibians [97,98].
Cannibalism and intraguild predation can also have important implications for the population dynamics, community structure, and distribution patterns of salamanders via growth/mortality trade-offs [99,100,101,102]. For example, Resetarits [103] found that the spring salamander (Gyrinophilus porphyriticus), a relatively big species, can impact the growth of a smaller salamander (the northern two-lined salamander, Eurycea bislineata) and a crayfish (Cambarus barton), but have no effect on the fitness (relative condition or fecundity) of brook trout (Salvelinus fontinalis). From a broader perspective, the study also recognized the notion that not all salamander species respond in the same way to the presence of predators; some are more vulnerable than those that can behaviorally avoid predation with high growth costs [103]. The consumption of conspecifics (cannibalism) is often observed in larval salamanders [28,29] and needs to be taken into account for a complete understanding of the dynamics and structural properties of populations and food webs. Cannibalism is commonly promoted by low food availability, high conspecific density, and long breeding seasons [104,105]. Long breeding seasons enable interactions among conspecifics of different sizes [105], with bigger individuals (cannibals show proportionally broader heads) being responsible for the cannibalism [104]. Thus, body size emerges as a key precursor of cannibalism, but it is not entirely clear whether body size is a cause (e.g., early hatching and thus bigger individuals having an advantage over smaller or late hatching individuals) or a consequence (e.g., increases in body size are facilitated by cannibalism) of cannibalism [104]. In this regard, the main advantages of cannibalism are the increasing survival and growth rate of individuals that become cannibalistic [106]. Cannibalism and intraguild predation have a key importance to population and community structure via larval production and metamorph emergence, which, in turn, have consequences in energy flow within aquatic systems and between systems (aquatic–terrestrial coupling) [106,107]. These studies demonstrated that cannibalism (intraspecific predation) and intraguild predation represent an energy loop that maintains nutrients and energy within salamander populations or communities, but with high costs because of larval mortality and thus loss of recruitment. In addition, competitive interactions among sympatric salamander species with similar size and structure may reduce the ability to predict changes in both population and community structure [102]. Thus, cannibalism and intraguild predation should not be neglected in studies attempting to explore the influence of population structure, behavior, life history, resource competition, and energy flow. In this regard, cannibalism and intraguild predation have direct consequences on the trophic levels of individuals by feeding on conspecifics and sympatric salamander species, and thus on the energy and nutrient flows in the aquatic ecosystem.
3.2. Salamanders as Energy Subsides for Higher Trophic Levels
Salamanders represent a large standing stock of nutrients in aquatic systems [108], but the nutrient composition of salamander species may change across ontogenetic stages [109]. Although salamanders usually have a lower capacity of nutrient recycling compared to other aquatic vertebrates such as fish species because excretion rates are higher in fish species [110,111], it is expected that salamanders can be important recyclers of biologically essential nutrients in fishless systems [21,111].
Despite there being good examples showing energy pathways in aquatic systems (e.g., leaf litter decomposition and detritus through bacteria into shredder and grazer invertebrates and then directly, or indirectly via mid-level consumers, to the main predators) [112,113], the role of salamanders in energy transfer to higher trophic levels is not completely known. It is thought that salamanders show a highly efficient conversion of invertebrate to vertebrate biomass, but low rates of energy flow compared to birds and mammals [114]. Regester et al. [23] yielded results consistent with the view that assimilation efficiencies for larval marbled salamanders are relatively high, demonstrating that this species efficiently converts ingested prey into biomass. Aquatic salamander communities provide a considerable energy flow (average net flux: 349.5 ± 140.8 g ash-free dry mass per year) into pond habitats [107]. Considering a density of 171 aquatic snakes ha−1, Willson and Winne [76] estimated that aquatic snakes consume over 200 kg (>55,000 individuals) of amphibian prey per year, which represents >150,000 kJ ha−1 of energy flow from salamanders (secondary consumers) to snakes (tertiary consumers). Thus, most of our current understanding of energy transfer in aquatic systems provides compelling evidence that salamanders are important energy subsides for higher trophic levels [76,79,108]. However, it is possible that salamanders are largely consumed by predators when salamanders are accessible and vulnerable to predators (i.e., breeding season) or when prey taxa other than salamanders (e.g., fish) are scarce in the environment e.g., [7,85]. Therefore, salamanders have a key role as energy subsides for higher trophic levels, but also need to be considered as a resource pulse (i.e., “episodes of increased resource availability in space and time that combine low frequency, large magnitude, and short duration”, which is a definition provided by Yang et al. [115]). Consistent with this view, there is ubiquitous evidence that salamanders can be an important energy source for consumers at higher trophic levels, but often they are spatially and temporally variable.
A better understanding of the role of salamanders in energy transfer demands a complete knowledge of energy inputs and outputs. Research in resource inputs for salamanders has identified the most important prey, in terms of energy inputs, for salamander species. For example, larval ambystomatid salamanders (marbled salamander and spotted salamanders) acquire most of their energy from copepods and larval dipterans [23]. It is possible that freshwater crustaceans other than copepods, such as isopods, cladocerans, and amphipods, may also be a major energy food source for salamanders in aquatic systems [116]. Denoël [117] quantified the origin (aquatic prey versus terrestrial invertebrates) of the energy intake by juvenile Alpine newts, concluding that 62% of the energy intake came from aquatic prey and the other 38% came from terrestrial invertebrates caught in water. Considering that around 10% of the energy ingested during larval development is associated with the production of metamorphosed salamanders [23], it can be expected that salamander species provide a key energy subsidy for higher trophic levels in aquatic systems. For example, newts represent a profitable prey for Eurasian otters [79]. Pagacz and Witczuk [118] estimated that the consumption of amphibians by Eurasian otters can be equally important as fish, constituting 54% of the biomass of consumed prey. Smiroldo et al. [80] revealed that the frequency of occurrence of amphibians in dietary studies in Eurasian otters averaged 12%. Considering that the frequency of occurrence only covers the total number of stomachs with prey and thus without taking the relative abundance of each prey type into account [119], it is doubtful that the use of literature sources only including frequency of occurrence can draw conclusions regarding the importance of salamanders as energy resources for otter species compared to papers reporting the abundance (numerical, biomass, or volume) of prey categories. Salamanders commonly represent a small portion of the stomach contents in brown trout and northern pike (Exos lucius) [54,77,78], which underscores the small importance of salamanders as energy subsides for piscivorous fish species such as, for example, many salmonid and pike species. On the other hand, salamanders are thought to be important energy sources for centrarchid species [86,87,120].
Except for works focused on aquatic systems [23,107], remarkable advances in the understanding of energy transfer through salamanders came from studies carried out in terrestrial systems. This is the case of the classical work by Burton and Likens [3], who showed that salamander populations in a forest ecosystem only utilized around 0.02% of the net primary productivity. The authors also provided solid evidence that salamander populations are a remarkable energy source for predators (20% of the energy flow through bird and mammal populations) [3]. Semlitsch et al. [13] suggested that salamanders in forest ecosystems may play a greater role in the trophic transfer of energy and nutrients, as salamander densities were 2–4 times greater than the values previously reported by Burton and Likens [3]. However, to the best of my knowledge, the trophic efficiency mediated by salamanders (i.e., the percentage of energy assimilated by salamanders through the consumption of aquatic invertebrates and the percentage of energy available for higher trophic levels) in aquatic systems has yet to be quantified. The relatively sparse literature on this topic suggests that this would be a fruitful area for future research. Such studies will likely reveal whether trophic efficiency mediated by salamanders can be generalized across multiple systems or species.
4. Salamanders as Promoters of Aquatic–Terrestrial Coupling (Lateral Energy Transfers)
Aquatic and terrestrial systems are not isolated, but interconnected [6,121,122]. Salamanders can couple the link between freshwater and terrestrial habitats through seasonal migrations (breeding adults) and emergences (metamorphosing larvae) e.g., [7,23,107]. Regester et al. [23] demonstrated that metamorphosed salamanders export 3–8% of total prey production to adjacent forest. Thus, the production of metamorphosed salamanders is key for predicting the magnitude of energy subsidies transferred from aquatic systems to terrestrial systems. As already pointed out, cannibalism and intraguild predation represent important regulation mechanisms that are responsible for larval production, and thus of emerging metamorphs [106,107]. In this regard, Regester et al. [107] quantified the grams of ash-free dry mass per year exported from ponds to surrounding forest by emerging metamorphs (between 21 and 135.2 g). This highlights the relevance of salamanders on the energy flow between aquatic and terrestrial habitats, which evidences the aquatic–terrestrial coupling promoted by emerging metamorphs. The coupling of aquatic and terrestrial systems can also be promoted by feeding on allochthonous food resources (terrestrial invertebrates). In fact, inputs of terrestrial invertebrates from riparian canopy cover may represent an important food resource for salamanders inhabiting aquatic environments [18,117]. The importance has been identified in multiple ways: in terms of energy subsidy [117], food resource partitioning between sympatric salamander species [18], or ecosystem functioning [6]. For example, Denoël [117] demonstrated that aquatic juveniles of Alpine newts in Drakolimni lake increase their energy intake using allochthonous food resources up to 60%. Thus, terrestrial invertebrates can provide high-energy gains [117], and they can represent more than 25% of salamanders’ diet in terms of biomass during the summer [123]. This represents a first level of aquatic–terrestrial coupling, by which salamanders rely on allochthonous food resources. However, it is reasonable to posit that, at least in the presence of drift feeders such as salmonids, this aquatic–terrestrial coupling is not promoted by salamanders (diets dominated by benthic aquatic invertebrates), as terrestrial subsides are monopolized by fish species [65].
The aquatic–terrestrial coupling can also be promoted when salamanders act as prey for terrestrial predators ([68] and references therein). For example, it is well known that predatory birds frequently feed on salamander species [68,69], which represents an important link between terrestrial and aquatic habitats. Another level of interface between aquatic and terrestrial habitats can be related to the direct movement of salamanders between these habitats. Due to the life cycle and water dependency of salamanders, they commonly migrate and disperse from their overwintering terrestrial spots to aquatic breeding sites and vice versa [124]. These movements link the energy and nutrient budgets between environments and drive new interactions in the colonized habitat [7,125]. Thus, salamander species intervene in ecosystem processes, enhancing the flux of energy and nutrients between terrestrial and aquatic habitats.
Overall, it is reasonable to posit that aquatic and terrestrial ecosystems can be strongly linked through salamanders, which mobilize nutrients and energy between both interconnected ecosystems (Figure 1). More specifically, this review provides fundamental evidence that the aquatic–terrestrial coupling can be promoted by salamanders at three levels: (i) the consumption of allochthonous food resources by salamanders (i.e., transfer of terrestrial resources to aquatic consumers), (ii) consumption of salamanders by terrestrial predators (i.e., transfer of aquatic resources to terrestrial consumers), and (iii) direct movement of salamanders between aquatic and terrestrial habitats (e.g., seasonal migrations of breeding adults and emergences of metamorphosed larval). These three points where coupling can be mediated by salamander species exemplify the importance of these taxa for understanding unidirectional and bidirectional lateral energy transfers across aquatic and terrestrial ecosystems.
5. Conclusions
This review synthetizes the role of salamanders in aquatic energy flow pathways, pointing out their crucial role in ecosystem functioning as promoters of vertical and lateral energy flows. The role of salamanders in the vertical dimension accepts the view that salamanders promote energy flow among trophic levels, mobilizing energy and nutrients from aquatic invertebrates up to salamanders, but also efficiently converting ingested prey into tissue biomass to become a profitable resource for higher trophic levels (Figure 1). The lateral dimension represents the processes by which energy and nutrients are transferred between aquatic and terrestrial habitats, as well as within trophic levels promoted by cannibalism and intraguild predation in aquatic ecosystems.
Despite research progress on the trophic ecology of salamanders, our knowledge and understanding mostly come from local geographic zones and single model organisms; thus, future research needs to be set in a broad ecological framework to answer global research questions. Studies focusing on the role of salamanders in energy flow pathways will need to be extended to the whole life cycle and ecosystem (aquatic and terrestrial) dimensions to be accepted as a general theory, representing a particularly promising area for future research. An additional promising avenue could be the exploration of global patterns in the trophic ecology of salamanders. In this regard, because high-level salamander phylogenies reflecting the link between diet and morphology (tooth shapes and dentition patterns) are available [126], further studies that integrate geographical and evolutionary frameworks into the feeding and dietary specialization of salamanders are required (see Rabosky et al. [127] for an example using fish as model organisms) to identify the basis of global patterns. Another fruitful area for future research could be exploration of the trophic efficiency of salamanders between trophic levels (i.e., between aquatic invertebrates and apex predators via salamanders as mid-level vertebrate predators). Future studies will likely reveal whether the role of salamanders in energy flow pathways varies geographically along ecosystem (aquatic and terrestrial) or latitudinal dimensions, generating novel insights into the implications of salamanders for communities and ecosystem processes and functioning at global scales.
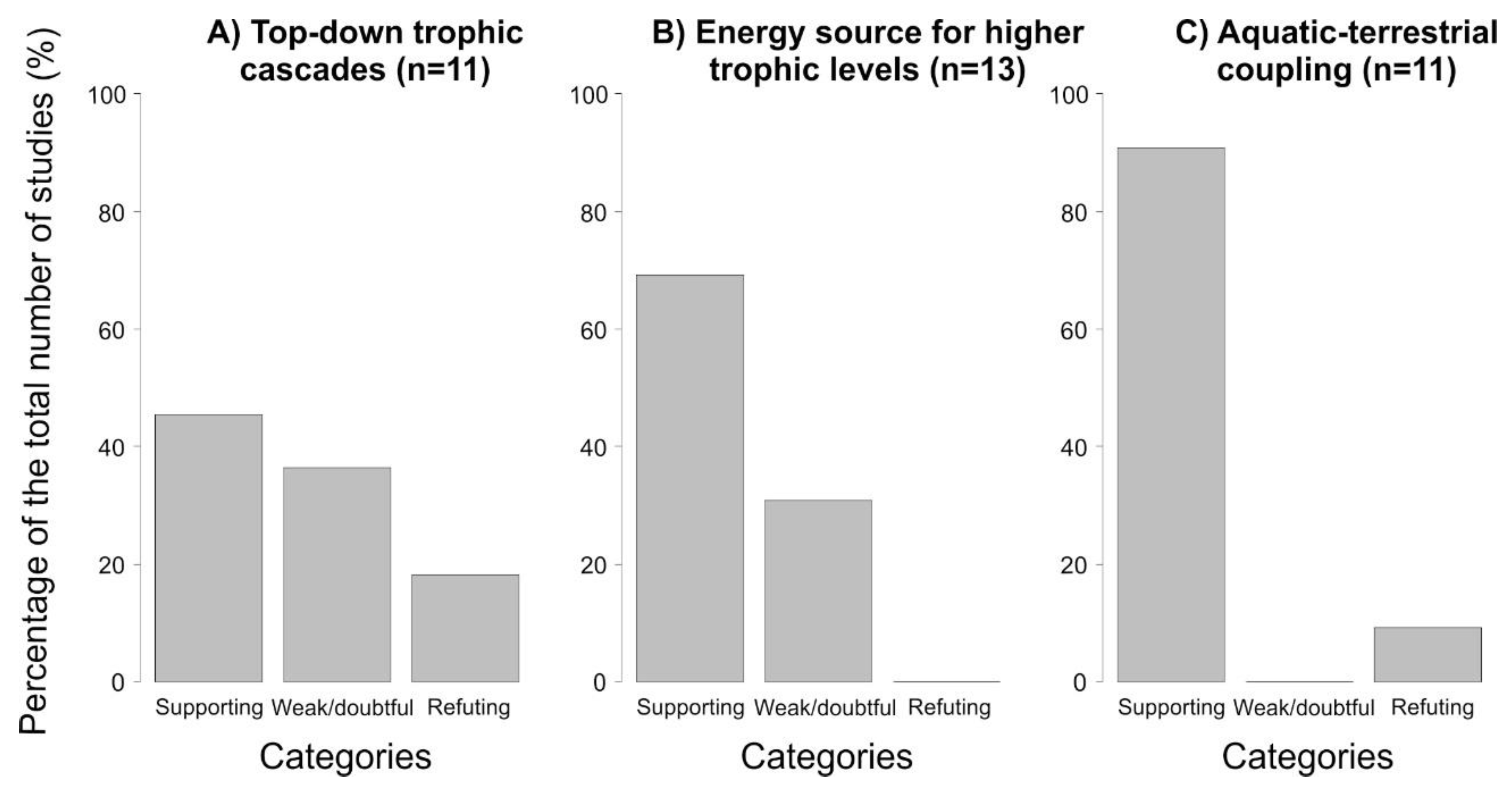
To date, there is both empirical and theoretical evidence showing that salamanders can be key in aquatic ecosystem functioning through several key processes related to top–down control, becoming an energy source for higher trophic levels, and aquatic–terrestrial coupling (Figure 2). As a caveat, caution should be exercised regarding this conclusion because this figure (i.e., summarizing the number of papers supporting or refuting the potential importance of salamanders in the different roles: aquatic top–down trophic cascades and vertical and lateral energy flows) is based on the literature used in each section (Section 2.2, Section 3.2 and Section 4, respectively) and not on an extensive literature review on the topics. Yet, this review emphasizes the ecological significance of salamanders in aquatic systems as central players in energy flow pathways, enabling energy mobility among trophic levels (i.e., vertical energy flow) and between freshwater and terrestrial habitats (i.e., lateral energy flow).
Funding
This research received no external funding.
Acknowledgments
Andy Nunn is acknowledged for valuable comments and corrections on the manuscript. I also appreciate constructive comments from three anonymous reviewers, which considerably improved the quality of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ivlev, V.S. Transformation of energy by aquatic animals. Int. Rev. Ges. Hydrobiol. Hydrogr. 1939, 38, 449–458. [Google Scholar] [CrossRef]
- Lindeman, R. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–418. [Google Scholar] [CrossRef]
- Burton, T.M.; Likens, G.E. Energy flow and nutrient cycling in salamander populations in the Hubbard Brook experimental forest, New Hampshire. Ecology 1975, 56, 1068–1080. [Google Scholar] [CrossRef]
- Thompson, R.M.; Brose, U.; Dunne, J.A.; Hall, R.O.; Hladyz, S.; Kitching, R.L.; Martinez, N.D.; Rantala, H.; Romanuk, T.N.; Stouffer, D.B.; et al. Food webs: Reconciling the structure and function of biodiversity. Trends Ecol. Evol. 2012, 27, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.J.; Cavanagh, R.D.; Drinkwater, K.F.; Grant, S.M.; Heymans, J.J.; Hofmann, E.E.; Hunt, G.L.; Johnston, N.M. Understanding the structure and functioning of polar pelagic ecosystems to predict the impacts of change. Proc. R. Soc. B 2016, 283, 20161646. [Google Scholar] [CrossRef] [Green Version]
- Nakano, S.; Miyasaka, H.; Kuhara, N. Terrestrial aquatic linkages: Riparian arthropod inputs alter trophic cascades in a stream food web. Ecology 1999, 80, 2435–2441. [Google Scholar]
- Davic, R.D.; Welsh, H.H.J. On the ecological roles of salamanders. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 405–434. [Google Scholar] [CrossRef] [Green Version]
- Power, M.E.; Holomuzki, J.R.; Lowe, R.L. Food webs in Mediterranean rivers. Hydrobiologia 2013, 719, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Allan, J.D.; Castillo, M.M. Stream Ecology Structure and Function of Running Waters, 2nd ed.; Chapman and Hall: New York, NY, USA, 2007. [Google Scholar]
- Holomuzki, J.R.; Collins, J.P.; Brunkow, P.E. Trophic control of fishless ponds by tiger salamander larvae. Oikos 1994, 71, 55–64. [Google Scholar] [CrossRef]
- Hocking, D.J.; Babbitt, K.J. Effects of Red-Backed Salamanders on Ecosystem Functions. PLoS ONE 2014, 9, e86854. [Google Scholar] [CrossRef] [Green Version]
- Nery, T.; Schmera, D. The effects of top-down and bottom-up controls on macroinvertebrate assemblages in headwater streams. Hydrobiologia 2016, 763, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Semlitsch, R.D.; O’Donnell, K.M.; Thompson, F.R. Abundance, biomass production, nutrient content, and the possible role of terrestrial salamanders in Missouri Ozark forest ecosystems. Can. J. Zool. 2014, 92, 997–1004. [Google Scholar] [CrossRef]
- Fasola, M.; Canova, L. Feeding habits of Triturus vulgaris, T. cristatus and T. alpestris (Amphibia, Urodela) in the northern Apennines (Italy). Boll Zool. 1992, 59, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Montori, A. Alimentación de las larvas de tritón pirenaico, Euproctus asper, en el prepirineo de la Cerdaña, España. Amphibia–Reptilia 1992, 13, 157–167. [Google Scholar] [CrossRef]
- Vignoli, L.; Bombi, P.; D’Amen, M.; Bologna, M.A. Seasonal variation in the trophic niche of a heterochronic population of Triturus alpestris apuanus from the south-western Alps. Herpetol. J. 2007, 17, 183–191. [Google Scholar]
- Romano, A.; Salvidio, S.; Palozzi, R.; Sbordoni, V. Diet of the newt, Triturus carnifex (Laurenti, 1768), in the flooded karst sinkhole Pozzo del Merro, central Italy. J. Caves Karst. Stud. 2012, 74, 271–277. [Google Scholar] [CrossRef]
- Sánchez-Hernández, J.; Montori, A.; Llorente, G.A. Ontogenetic dietary shifts and food resource partitioning in a stream-dwelling Urodela community: Mechanisms to allow coexistence across seasons. Russ. J. Herpetol. 2019, 26, 135–149. [Google Scholar] [CrossRef]
- Joly, P.; Giacoma, C. Limitation of similarity and feeding habits in three syntopic species of newts (Triturus, Amphibia). Ecography 1992, 15, 401–411. [Google Scholar] [CrossRef]
- Salvidio, S.; Oneto, F.; Ottonello, D.; Costa, A.; Romano, A. Trophic specialization at the individual level in a terrestrial generalist salamander. Can. J. Zool. 2015, 93, 79–83. [Google Scholar] [CrossRef]
- Keitzer, S.C.; Goforthm, R.R. Spatial and seasonal variation in the ecological significance of nutrient recycling by larval salamanders in Appalachian headwater streams. Freshw. Sci. 2013, 32, 1136–1147. [Google Scholar] [CrossRef]
- Best, M.L.; Welsh, H.H. The trophic role of a forest salamander: Impacts on invertebrates, leaf litter retention, and the humification process. Ecosphere 2014, 5, 16. [Google Scholar] [CrossRef]
- Regester, K.J.; Whiles, M.R.; Lips, K.R. Variation in the trophic basis of production and energy flow associated with emergence of larval salamander assemblages from forest ponds. Freshwat. Biol. 2008, 53, 1754–1767. [Google Scholar] [CrossRef]
- Braña, F.; de la Hoz, M.; Lastra, C. Alimentación y relaciones tróficas entre las larvas de Triturus marmoratus, T. alpestris y T. helveticus (Amphibia, Caudata). Doñana Acta Vert. 1986, 13, 21–33. [Google Scholar]
- Rosa, G.; Costa, A.; Salvidio, S. Trophic strategies of two newt populations living in contrasting habitats on a Mediterranean island. Ethol. Ecol. Evol. 2019. [Google Scholar] [CrossRef]
- Parker, M.S. Predation by Pacific giant salamander larvae on juvenile steelhead trout. Northwest Nat. 1993, 74, 77–81. [Google Scholar] [CrossRef]
- Montaña, C.G.; Ceneviva-Bastos, M.; Schalk, C.M. New vertebrate prey for the aquatic salamander Amphiuma means (Caudata: Amphiumidae). Herpetol. Notes 2014, 7, 755–756. [Google Scholar]
- Park, S.-R.; Jeong, J.-Y.; Park, D. Cannibalism in the Korean salamander (Hynobius leechii: Hynobiidae, caudata, amphibia) larvae. Integr. Biosci. 2005, 9, 13–18. [Google Scholar] [CrossRef]
- Jefferson, D.M.; Ferrari, M.C.O.; Mathis, A.; Hobson, K.A.; Britzke, E.R.; Crane, A.L.; Blaustein, A.R.; Chivers, D.P. Shifty salamanders: Transient trophic polymorphism and cannibalism within natural populations of larval ambystomatid salamanders. Front. Zool. 2014, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, F.; Perez-Mellado, V.; Gil, M.J.; Lizana, M. Food habits and trophic availability in the high mountain population of the spotted salamander from Spain (Salamandra salamandra almanzoris) (Caudata: Salamandridae). Folia Zool. 1990, 89, 841–853. [Google Scholar]
- Jaeger, R.G. Diet diversity and clutch size of aquatic and terrestrial salamanders. Oecologia 1981, 48, 190–193. [Google Scholar] [CrossRef]
- Salvidio, S.; Crovetto, F.; Costa, A. Individual trophic specialisation in the Alpine newt increases with increasing resource diversity. Ann. Zool. Fenn. 2019, 56, 17–24. [Google Scholar] [CrossRef]
- Ranta, E.; Tjossem, S.F.; Leikola, N. Female-male activity and zooplankton foraging by the smooth newt (Triturus vulgaris). Ann. Zool. Fennici. 1987, 24, 79–88. [Google Scholar]
- Denoël, M.; Joly, P. Size-related predation reduces intramorph competition in paedomorphic alpine newts. Can. J. Zool. 2001, 79, 943–948. [Google Scholar] [CrossRef]
- Maerz, J.C.; Myers, E.M.; Adams, D.C. Trophic polymorphism in a terrestrial salamander. Evol. Ecol. Res. 2006, 8, 23–35. [Google Scholar]
- Sánchez-Hernández, J. Disentangling prey-handling efficiency of larval newts through multivariate prey trait analysis. J. Nat. Hist. 2014, 48, 1957–1969. [Google Scholar] [CrossRef]
- Emlen, J.M. The role of time and energy in food preference. Am. Nat. 1966, 100, 611–617. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Pianka, E.R. On optimal use of a patchy environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Pyke, G.H.; Pulliam, H.R.; Charnov, E.L. Optimal foraging: A selective review of theory and tests. Q. Rev. Biol. 1977, 52, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Cicort-Lucaciu, A.-Ş.; Ardeleanu, A.; Cupşa, D.; Naghi, N. The trophic spectrum of a Triturus cristatus (Laurentus 1768) population from Plopiş Mountains area (Bihor County, Romania). North-West J. Zool. 2005, 1, 31–39. [Google Scholar]
- Crump, M.L. Opportunistic cannibalism by amphibian larvae in temporary aquatic environments. Am. Nat. 1983, 121, 281–287. [Google Scholar] [CrossRef]
- Crossland, M.R. Predation by tadpoles on toxic toad eggs: The effect of tadpole size on predation success and tadpole survival. J. Herpetol. 1998, 32, 443–446. [Google Scholar] [CrossRef]
- de Crespin de Billy, V.; Usseglio-Polatera, P. Traits of brown trout prey in relation to habitat characteristics and benthic invertebrate communities. J. Fish Biol. 2002, 60, 687–714. [Google Scholar] [CrossRef]
- de Crespin de Billy, V.; Dumont, B.; Lagarrigue, T.; Baran, P.; Statzner, B. Invertebrate accessibility and vulnerability in the analysis of brown trout (Salmo trutta L.) summer habitat suitability. River Res. Appl. 2002, 18, 533–553. [Google Scholar] [CrossRef]
- Sánhez-Hernández, J.; Vieira-Lanero, R.; Servia, M.J.; Cobo, F. Feeding habits of four sympatric fish species in the Iberian Peninsula: Keys to understanding coexistence using prey traits. Hydrobiologia 2011, 667, 119–132. [Google Scholar] [CrossRef]
- Ranta, E.; Nuutinen, V. Foraging by the smooth newt (Triturus vulgaris) on zooplankton: Functional response and diet choice. J. Anim. Ecol. 1985, 54, 275–293. [Google Scholar] [CrossRef]
- Ranta, E.; Saloheimo, M.; Nuutinen, V. Orientation of the smooth newt (Triturus vulgaris) foraging on zooplankton. Ann Zool Fennici. 1986, 23, 281–287. [Google Scholar]
- Heiss, E.; Natchev, N.; Gumpenberger, M.; Weissenbacher, A.; Van Wassenbergh, S. Biomechanics and hydrodynamics of prey capture in the Chinese giant salamander reveal a high-performance jaw-powered suction feeding mechanism. J. R. Soc. Interface 2013, 10, 20121028. [Google Scholar] [CrossRef] [Green Version]
- Kopecký, O.; Vojar, J.; Šusta, F.; Rehák, I. Composing and scaling of male and female alpine newt (Mesotriton alpestris) prey, with related site and seasonal effect. Ann. Zool. Fenn. 2012, 9, 231–239. [Google Scholar] [CrossRef]
- Farasat, H.; Sharifi, M. Food habit of the endangered yellow-spotted newt Neurergus microspilotus (Caudata, Salamandridae) in Kavat Stream, western Iran. Zool. Stud. 2014, 53, 61. [Google Scholar] [CrossRef] [Green Version]
- Falke, L.P.; Henderson, J.S.; Novak, M.; Preston, D.L. Temporal shifts in intraspecific and interspecific diet variation: Effects of predator body size and identity across seasons in a stream community. bioRxiv 2018. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What shapes the trophic niche of European plethodontid salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef] [PubMed]
- Clavero, M.; Prenda, J.; Delibes, M. Trophic diversity of the otter (Lutra lutra L.) in temperate and Mediterranean freshwater habitats. J. Biogeogr. 2003, 30, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Hernández, J.; Finstad, A.G.; Arnekleiv, J.V.; Kjærstad, G.; Amundsen, P.-A. Drivers of diet patterns in a globally distributed freshwater fish species. Can. J. Fish. Aquat. Sci. 2019, 76, 1263–1274. [Google Scholar] [CrossRef]
- Holomuzki, J.R.; Collins, J.P. Trophic dynamics of a top predator, Ambystoma tigrinum nebulosum (Caudata: Ambystomatidae), in a lentic community. Copeia 1987, 1987, 949–957. [Google Scholar] [CrossRef]
- Parker, M.S. Feeding Ecology of Stream-Dwelling Pacific Giant Salamander Larvae (Dicamptodon tenebrosus). Copeia 1994, 1994, 705–718. [Google Scholar] [CrossRef]
- Wissinger, S.A.; Whiteman, H.H.; Sparks, G.B.; Rouse, G.L.; Brown, W.S. Foraging trade-offs along a predator-permanence gradient in subalpine wetlands. Ecology 1999, 80, 2102–2116. [Google Scholar]
- Sánchez-Hernández, J.; Cobo, F.; Amundsen, P.-A. Food Web Topology in High Mountain Lakes. PLoS ONE 2015, 10, e0143016. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C. Evolution mediates the effects of apex predation on aquatic food webs. Proc. R. Soc. B 2013, 280, 20130859. [Google Scholar] [CrossRef]
- Atlas, W.I.; Palen, W.J. Prey Vulnerability Limits Top-Down Control and Alters Reciprocal Feedbacks in a Subsidized Model Food Web. PLoS ONE 2014, 9, e85830. [Google Scholar] [CrossRef] [Green Version]
- Hickerson, C.A.M.; Anthony, C.D.; Walton, B.M. Eastern Red-backed Salamanders regulate top-down effects in a temperate forest-floor community. Herpetologica 2017, 73, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.L.; Rowland, F.E.; Semlitsch, R.D. Variation in phenology and density differentially affects predator–prey interactions between salamanders. Oecologia 2017, 185, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Rowland, F.E.; Rawlings, M.B.; Semlitsch, R.D. Joint effects of resources and amphibians on pond ecosystems. Oecologia 2017, 183, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Reice, S.R.; Edwards, R.L. The effect of vertebrate predation on lotic macroinvertebrate communities in Quebec, Canada. Can. J. Zool. 1986, 64, 1930–1936. [Google Scholar] [CrossRef]
- Atlas, W.I.; Palen, W.J.; Courcelles, D.M.; Munshaw, R.G.; Monteith, Z.L. Dependence of stream predators on terrestrial prey fluxes: Food web responses to subsidized predation. Ecosphere 2013, 4, 69. [Google Scholar] [CrossRef]
- Reinhardt, T.; Brauns, M.; Steinfartz, S.; Weitere, M. Effects of salamander larvae on food webs in highly subsidised ephemeral ponds. Hydrobiologia 2017, 799, 37–48. [Google Scholar] [CrossRef]
- Preston, D.L.; Johnson, P.T.J. Importance of Native Amphibians in the Diet and Distribution of the Aquatic Gartersnake (Thamnophis atratus) in the San Francisco Bay Area of California. J. Herpetol. 2012, 46, 221–227. [Google Scholar] [CrossRef]
- Jobe, K.L.; Montaña, C.G.; Schalk, C.M. Emergent patterns between salamander prey and their predators. Food Webs 2019. [Google Scholar] [CrossRef]
- Brandon, R.A.; Huheey, J.E. Diurnal Activity, Avian Predation, and the Question of Warning Coloration and Cryptic Coloration in Salamanders. Herpetologica 1975, 31, 252–255. [Google Scholar]
- Brodie, E.D.; Ridenhour, B.J.; Brodie, E.D. The evolutionary response of predators to dangerous prey: Hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts. Evolution 2002, 56, 2067–2082. [Google Scholar] [CrossRef]
- Brodie, E.D.; Feldman, C.R.; Hanifin, C.T.; Motychak, J.E.; Mulcahy, D.G.; Williams, B.L.; Brodie, E.D. Parallel Arms Races between Garter Snakes and Newts Involving Tetrodotoxin as the Phenotypic Interface of Coevolution. J. Chem. Ecol. 2005, 31, 343–356. [Google Scholar] [CrossRef]
- Stokes, A.N.; Ray, A.M.; Buktenica, M.W.; Gall, B.G.; Paulson, E.; Paulson, D.; French, S.S.; Brodie, E.D.; Brodie, E.D. Otter Predation on Taricha granulosa and Variation in Tetrodotoxin Levels with Elevation. Northwest Nat. 2015, 96, 13–21. [Google Scholar] [CrossRef]
- Bringsøe, H.; Nørgaard, J. Predation of Triturus cristatus (Caudata: Salamandridae) by the Eurasian otter, Lutra lutra (Carnivora: Mustelidae). Herpetol. Notes 2018, 11, 279–280. [Google Scholar]
- Gregory, P.T.; Isaac, L.A. Food Habits of the Grass Snake in Southeastern England: Is Natrix natrix a Generalist Predator? J. Herpetol. 2004, 38, 88–95. [Google Scholar] [CrossRef]
- Escoriza, D.; Hassine, J.B. First case of predation in Pleurodeles poireti (Gervais, 1835). Bol. Asoc. Herpetol. Esp. 2017, 28, 19–20. [Google Scholar]
- Willson, J.D.; Winne, C.T. Evaluating the functional importance of secretive species: A case study of aquatic snake predators in isolated wetlands. J. Zool. 2015, 298, 266–273. [Google Scholar] [CrossRef]
- Willson, J.D.; Hopkins, W.A. Prey morphology constrains the feeding ecology of an aquatic generalist predator. Ecology 2011, 92, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Velo-Antón, G.; Cordero-Rivera, A. Predation by invasive mammals on an insular viviparous population of Salamandra salamandra. Herpetol. Notes 2011, 4, 299–301. [Google Scholar]
- Parry, G.S.; Yonow, N.; Forman, D. Predation of newts (Salamandridae, Pleurodelinae) by Eurasian otters Lutra lutra (Linnaeus). Herpetol. Bull. 2015, 132, 9–14. [Google Scholar]
- Smiroldo, G.; Villa, A.; Tremolada, P.; Gariano, P.; Balestrieri, A.; Delfino, M. Amphibians in Eurasian otter Lutra lutra diet: Osteological identification unveils hidden prey richness and male-biased predation on anurans. Mammal Rev. 2019. [Google Scholar] [CrossRef]
- Clavero, M.; Prenda, J.; Delibes, M. Amphibian and reptile consumption by otters (Lutra lutra) in a coastal area in Southern Iberian Peninsula. Herpetol. J. 2005, 15, 125–131. [Google Scholar]
- Novais, A.; Sedlmayr, A.; Moreira-Santos, M.; Goncalves, F.; Ribeiro, R. Diet of the otter Lutra lutra in an almost pristine Portuguese river: Seasonality and analysis of fish prey through scale and vertebrae keys and length relationships. Mammalia 2010, 74, 71–81. [Google Scholar] [CrossRef]
- Cogălniceanu, D.; Márquez, R.; Beltrán, J.F. Impact of otter (Lutra lutra) predation on amphibians in temporary ponds in Southern Spain. Acta Herpetol. 2010, 5, 217–222. [Google Scholar]
- Hanifin, C.T. The chemical and evolutionary ecology of tetrodotoxin (TTX) toxicity in terrestrial vertebrates. Mar. Drugs 2010, 8, 577–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiroldo, G.; Gariano, P.; Balestrieri, A.; Manenti, R.; Pini, E.; Tremolada, P. Predation on Amphibians May Enhance Eurasian Otter Recovery in Southern Italy. Zool. Sci. 2019, 36, 273–283. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Interactions between fish and salamander larvae. Oecologia 1987, 72, 481–486. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Allotopic Distribution of Two Salamanders: Effects of Fish Predation and Competitive Interactions. Copeia 1988, 1988, 290–298. [Google Scholar] [CrossRef]
- Domínguez, J.; Pena, J.C. Alimentación del lucio Esox lucius en un área de reciente colonización (cuenca del Esla, noroeste de España). Variaciones en función de la talla. Ecologia 2001, 15, 293–308. [Google Scholar]
- Knapp, R.A. Effects of nonnative fish and habitat characteristics on lentic herpetofauna in Yosemite National Park, USA. Biol. Conserv. 2005, 121, 265–279. [Google Scholar] [CrossRef]
- Kats, L.B.; Ferrer, R.P. Alien predators and amphibian declines: Review of two decades of science and the transition to conservation. Divers. Distrib. 2003, 9, 99–110. [Google Scholar] [CrossRef]
- Orizaola, G.; Braña, F. Effect of salmonid introduction and other environmental characteristics on amphibian distribution and abundance in mountain lakes of northern Spain. Anim. Conservat. 2006, 9, 171–178. [Google Scholar] [CrossRef]
- Tiberti, R.; Bogliani, G.; Brighenti, S.; Iacobuzio, R.; Liautaud, K.; Rolla, M.; von Hardenberg, A.; Bassano, B. Recovery of high mountain Alpine lakes after the eradication of introduced brook trout Salvelinus fontinalis using non-chemical methods. Biol. Invasions 2019, 21, 875–894. [Google Scholar] [CrossRef]
- Martínez-Solano, I.; Barbadillo, J.; Lapeña, M. Effect of introduced fish on amphibian species richness and densities at a montane assemblage in the Sierra de Neila, Spain. Herpetol. J. 2003, 13, 167–173. [Google Scholar]
- Domínguez, J.; Pena, J.C. Spatio-temporal variation in the diet of northern pike (Esox lucius) in a colonised area (Esla basin, NW Spain). Limnetica 2000, 19, 1–20. [Google Scholar]
- Cobo, F.; Sánchez-Hernández, J.; Vieira-Lanero, R.; Servia, M.J. Organic pollution induces domestication-like characteristics in feral populations of brown trout (Salmo trutta). Hydrobiologia 2013, 705, 119–134. [Google Scholar] [CrossRef]
- Adrián, M.I.; Delibes, M. Food habits of the otter (Lutra lutra) in two habitats of the Doñana National Park, SW Spain. J. Zool. 1987, 212, 399–406. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Gustafson, M.P. Intraguild predation among larval plethodontid salamanders: A field experiment in artificial stream pools. Oecologia 1993, 96, 271–275. [Google Scholar] [CrossRef]
- Yurewicz, K.L. A growth/mortality trade-off in larval salamanders and the coexistence of intraguild predators and prey. Oecologia 2004, 138, 102–111. [Google Scholar] [CrossRef]
- Anderson, T.L.; Semlitsch, R.D. Top predators and habitat complexity alter an intraguild predation module in pond communities. J. Anim. Ecol. 2016, 85, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.L.; Linares, C.; Dodson, K.N.; Semlitsch, R.D. Variability in functional response curves among larval salamanders: Comparisons across species and size classes. Can. J. Zool. 2016, 94, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Resetarits, W.J., Jr. Ecological interactions among predators in experimental stream communities. Ecology 1991, 72, 1782–1793. [Google Scholar] [CrossRef]
- Nyman, S.; Wilkinson, R.F.; Hutcherson, J.E. Cannibalism and size relations in a cohort of larval ringed salamanders (Ambystoma annulatum). J. Herpet. 1993, 27, 78–84. [Google Scholar] [CrossRef]
- Wildy, E.L.; Chivers, D.P.; Kiesecker, J.M.; Blaustein, A.R. The effects of food level and conspecific density on biting and cannibalism in larval long-toed salamanders, Ambystoma macrodactylum. Oecologia 2001, 128, 202–209. [Google Scholar] [CrossRef]
- Polis, G.A. The Evolution and Dynamics of Intraspecific Predation. Annu. Rev. Ecol. Evol. Syst. 1981, 12, 225–251. [Google Scholar] [CrossRef]
- Regester, K.J.; Lips, K.R.; Whiles, M.R. Energy flow and subsidies associated with the complex life cycle of ambystomatid salamanders in ponds and adjacent forest in southern Illinois. Oecologia 2006, 147, 303–314. [Google Scholar] [CrossRef]
- Milanovich, J.R.; Maerz, J.C.; Rosemond, A.D. Stoichiometry and estimates of nutrient standing stocks of larval salamanders in Appalachian headwater streams. Freshw. Biol. 2015, 60, 1340–1353. [Google Scholar] [CrossRef]
- Prater, C.; Scott, D.E.; Lance, S.L.; Nunziata, S.O.; Sherman, R.; Tomczyk, N.; Capps, K.A.; Jeyasingh, P.D. Understanding variation in salamander ionomes: A nutrient balance approach. Freshw. Biol. 2019, 64, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Munshaw, R.G.; Palen, W.J.; Courcelles, D.M.; Finlay, J.C. Predator-Driven Nutrient Recycling in California Stream Ecosystems. PLoS ONE 2013, 8, e58542. [Google Scholar] [CrossRef] [Green Version]
- Milanovich, J.R.; Hopton, M.E. Stoichiometry of Excreta and Excretion Rates of a Stream-dwelling Plethodontid Salamander. Copeia 2016, 104, 26–34. [Google Scholar] [CrossRef]
- Efford, I.E. Energy transfer in Marion Lake, British Columbia; with particular reference to fish feeding. Verh. Int. Ver. Limnol. 1969, 17, 104–108. [Google Scholar] [CrossRef]
- Wallace, J.B.; Eggert, S.L.; Meyer, J.L.; Webster, J.R. Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 1997, 277, 102–104. [Google Scholar] [CrossRef] [Green Version]
- Pough, F.H. Amphibians and reptiles as low energy systems. In Behavioral Energetics; The Cost of Survival in Vertebrates; Aspey, W.P., Lustick, S.I., Eds.; Ohio State University Press: Columbus, OH, USA, 1983; pp. 141–188. [Google Scholar]
- Yang, L.H.; Bastow, J.L.; Spence, K.O.; Wright, A.N. What can we learn from resource pulses? Ecology 2008, 89, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Whiles, M.R.; Jensen, J.B.; Palis, J.G. Diets of larval flatwoods salamanders, Ambystoma cingulatum, from Florida and South Carolina. J. Herpetol. 2004, 38, 208–214. [Google Scholar] [CrossRef]
- Denoël, M. Terrestrial versus aquatic foraging in juvenile Alpine newts (Triturus alpestris). Ecoscience 2004, 11, 404–409. [Google Scholar] [CrossRef]
- Pagacz, S.; Witczuk, J. Intensive Exploitation of Amphibians by Eurasian Otter (Lutra lutra) in the Wołosaty Stream, Southeastern Poland. Ann. Zool. Fenn. 2010, 47, 403–410. [Google Scholar] [CrossRef]
- Amundsen, P.-A.; Sánchez-Hernández, J. Feeding studies take guts—Critical review and recommendations of methods for stomach contents analysis in fish. J. Fish Biol. 2019, 95, 1364–1373. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.R.; Rettig, J.E.; Mittelbach, G.G.; Valiulis, J.L.; Schaack, S.R. The effects of fish on assemblages of amphibians in ponds: A field experiment. Freshw. Biol. 1999, 41, 829–837. [Google Scholar] [CrossRef]
- Nakano, S.; Murakami, M. Reciprocal subsidies: Dynamic interdependence between terrestrial and aquatic food webs. Proc. Natl. Acad. Sci. USA 2001, 98, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Greene, B.T.; Lowe, W.H.; Likens, G.E. Forest succession and prey availability influence the strength and scale of terrestrial-aquatic linkages in a headwater salamander system. Freshw. Biol. 2008, 53, 2234–2243. [Google Scholar] [CrossRef]
- Montori, A. Alimentación de los adultos de Euproctus asper (Dugès, 1852) en la montaña media del Prepirineo catalán (España). Rev. Esp. Herpetol. 1991, 5, 23–36. [Google Scholar]
- Semlitsch, R.D. Differentiating migration and dispersal processes for pond-breeding amphibians. J. Wildl. Manag. 2008, 72, 260–267. [Google Scholar] [CrossRef]
- Reinhardt, T.; Steinfartz, S.; Paetzold, A.; Weitere, M. Linking the evolution of habitat choice to ecosystem functioning: Direct and indirect effects of pond-reproducing fire salamanders on aquatic-terrestrial subsidies. Oecologia 2013, 173, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.L.; Sears, B.R.; Wooten, J.A.; Camp, C.D.; Falk, A.; O’Quin, K.; Pauley, T.K. Evolution of dentition in salamanders: Relative roles of phylogeny and diet. Biol. J. Linn. Soc. 2016, 119, 960–973. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Chang, J.; Title, P.O.; Cowman, P.F.; Sallan, L.; Friedman, M.; Kaschner, K.; Garilao, C.; Near, T.J.; Coll, M.; et al. An inverse latitudinal gradient in speciation rate for marine fishes. Nature 2018, 559, 392–395. [Google Scholar] [CrossRef]
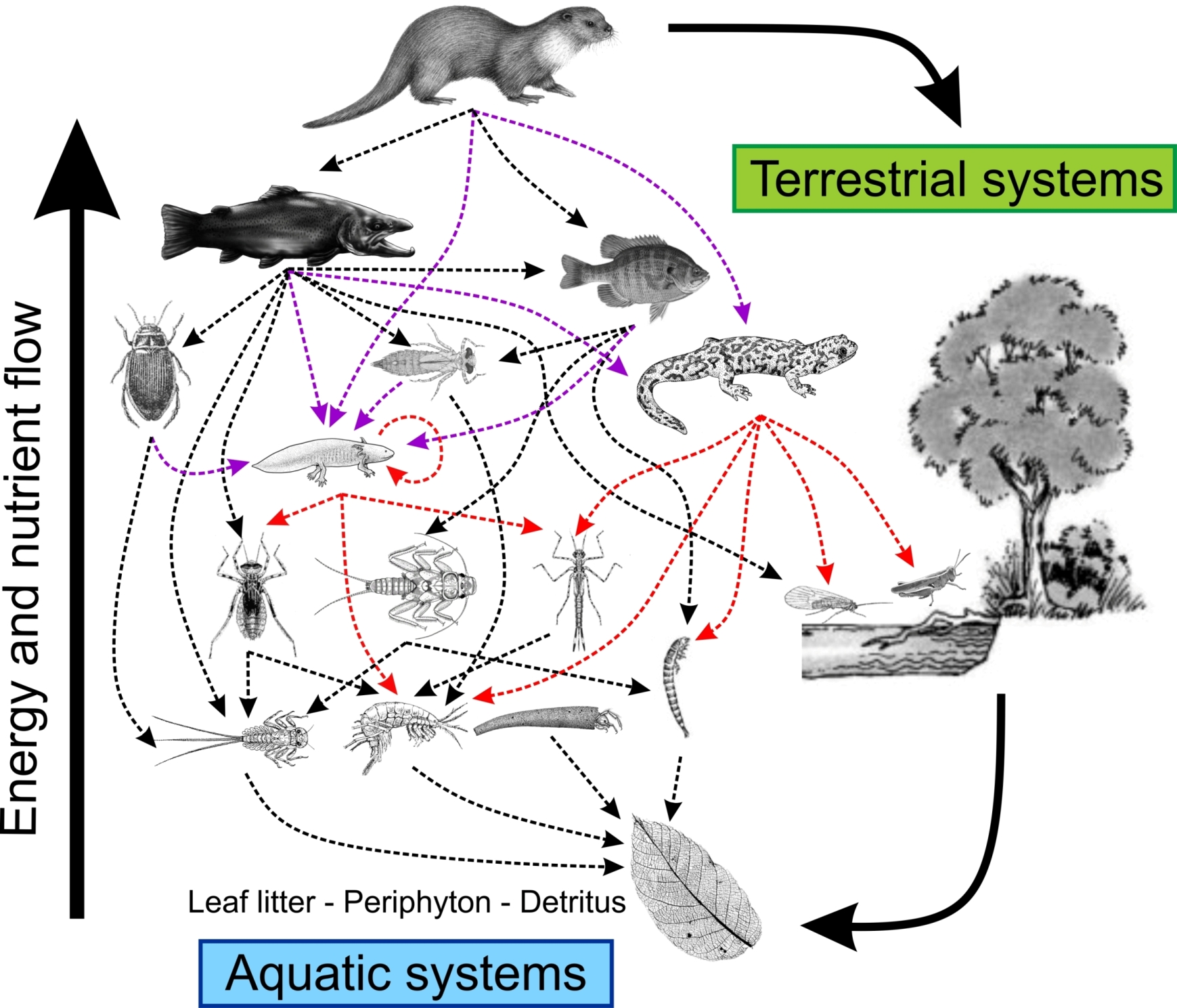
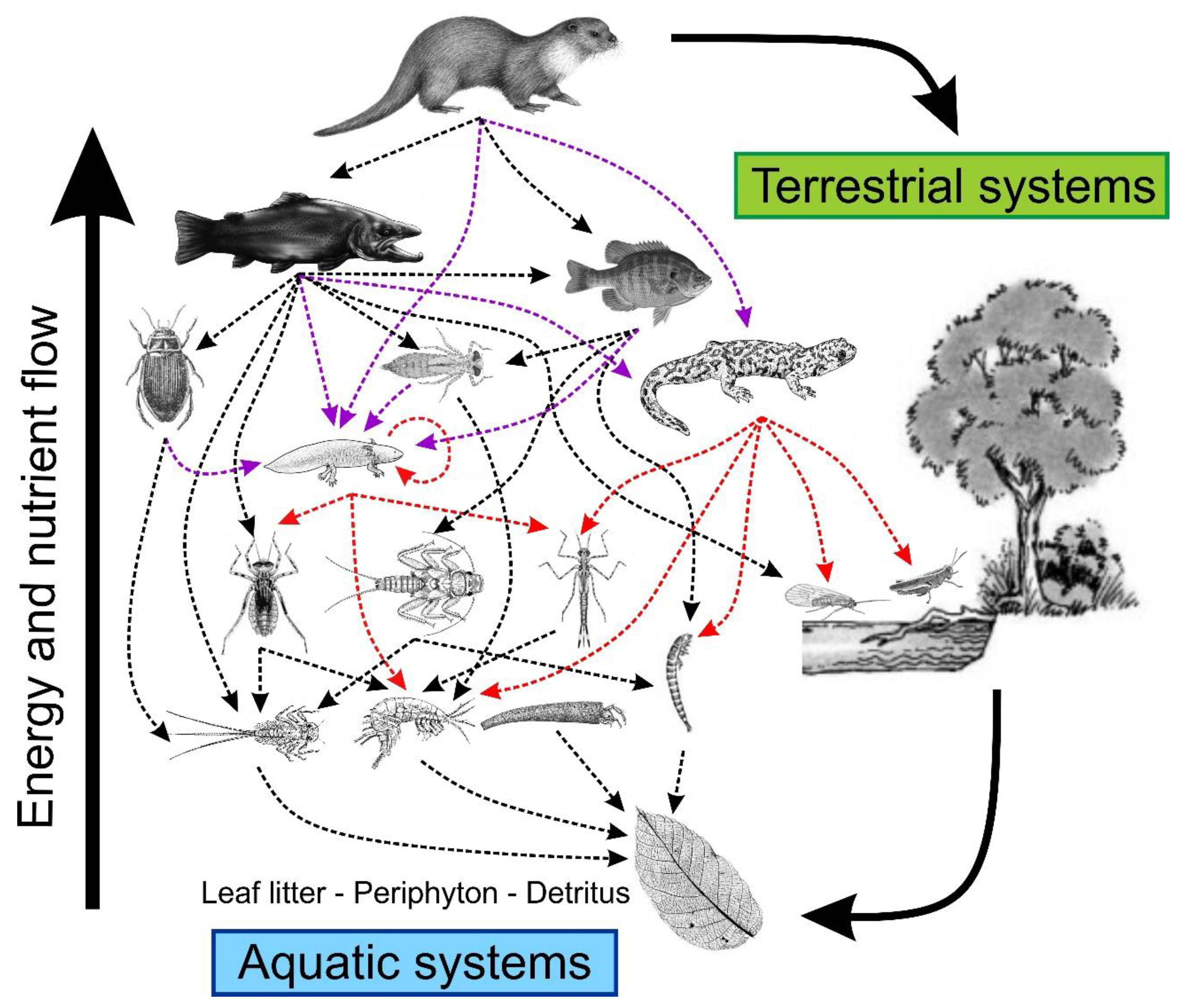
Figure 1.
Conceptual view of a hypothetical aquatic food web dominated by semi-aquatic vertebrates (here Eurasian otter, Lutra lutra), with salamanders as mid-level vertebrate predators and preys. Red dashed lines represent the interaction of salamanders as predators. Purple dashed lines represent the interaction of salamanders as prey. Arrows indicate the direction of predator–prey interaction and energy flow.
Figure 1.
Conceptual view of a hypothetical aquatic food web dominated by semi-aquatic vertebrates (here Eurasian otter, Lutra lutra), with salamanders as mid-level vertebrate predators and preys. Red dashed lines represent the interaction of salamanders as predators. Purple dashed lines represent the interaction of salamanders as prey. Arrows indicate the direction of predator–prey interaction and energy flow.

Figure 2.
Number of papers supporting or refuting the potential importance of salamanders in aquatic top–down trophic cascades (A), energy source for higher trophic levels (i.e., vertical energy flow) (B), and energy mobility between freshwater and terrestrial habitats (i.e., lateral energy flow) (C). This plot is made according to literature used in each section (Section 2.2, Section 3.2 and Section 4, respectively) of this review. Papers were classified according to three categories: (i) supporting, (ii) weak/doubtful, and (iii) refuting.
Figure 2.
Number of papers supporting or refuting the potential importance of salamanders in aquatic top–down trophic cascades (A), energy source for higher trophic levels (i.e., vertical energy flow) (B), and energy mobility between freshwater and terrestrial habitats (i.e., lateral energy flow) (C). This plot is made according to literature used in each section (Section 2.2, Section 3.2 and Section 4, respectively) of this review. Papers were classified according to three categories: (i) supporting, (ii) weak/doubtful, and (iii) refuting.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sánchez-Hernández, J. Reciprocal Role of Salamanders in Aquatic Energy Flow Pathways. Diversity 2020, 12, 32. https://0-doi-org.brum.beds.ac.uk/10.3390/d12010032
AMA Style
Sánchez-Hernández J. Reciprocal Role of Salamanders in Aquatic Energy Flow Pathways. Diversity. 2020; 12(1):32. https://0-doi-org.brum.beds.ac.uk/10.3390/d12010032
Chicago/Turabian StyleSánchez-Hernández, Javier. 2020. "Reciprocal Role of Salamanders in Aquatic Energy Flow Pathways" Diversity 12, no. 1: 32. https://0-doi-org.brum.beds.ac.uk/10.3390/d12010032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.