High Prevalence of Novel Beak and Feather Disease Virus in Sympatric Invasive Parakeets Introduced to Spain From Asia and South America


Abstract
:
1. Introduction
2. Materials and Methods
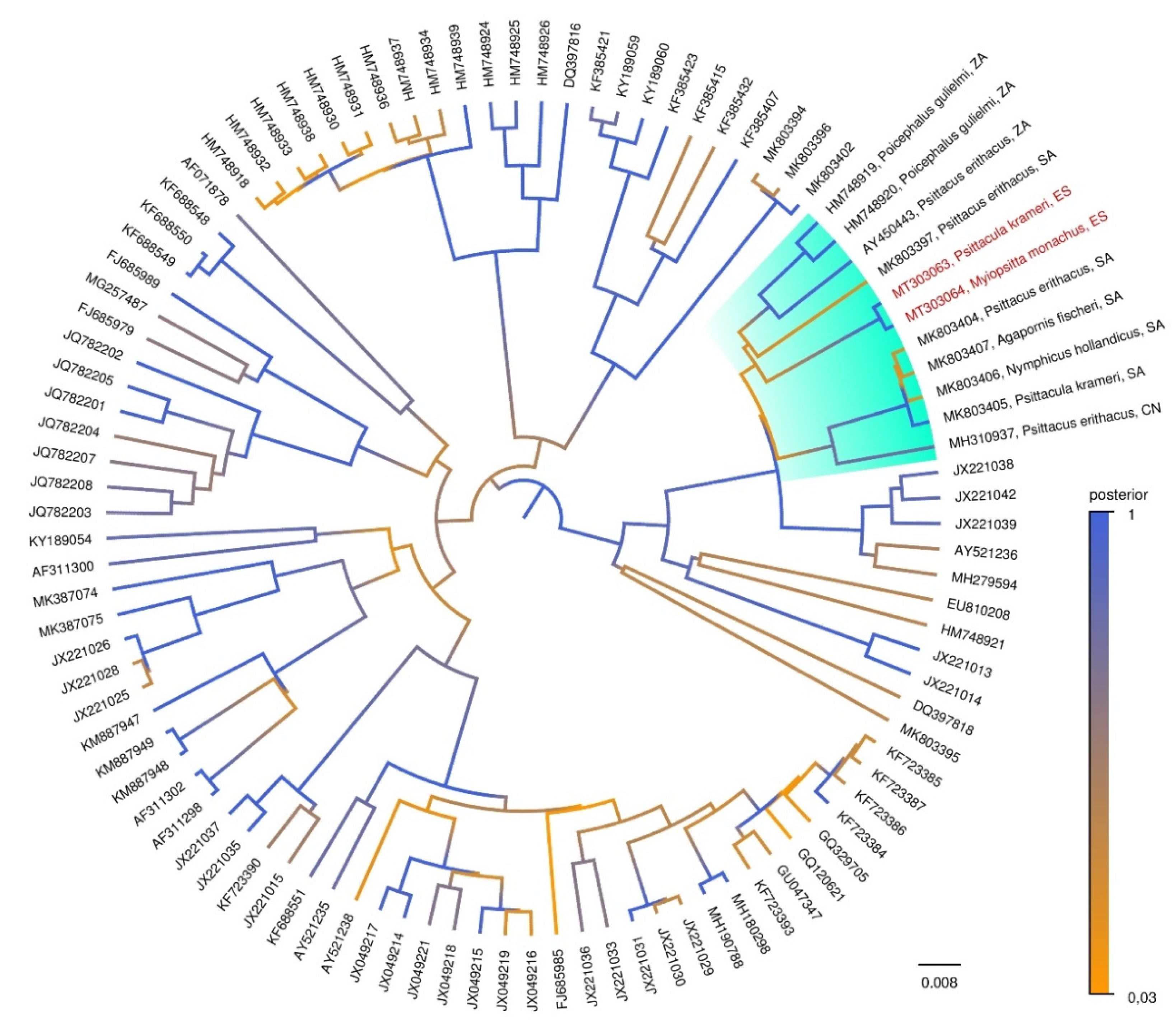
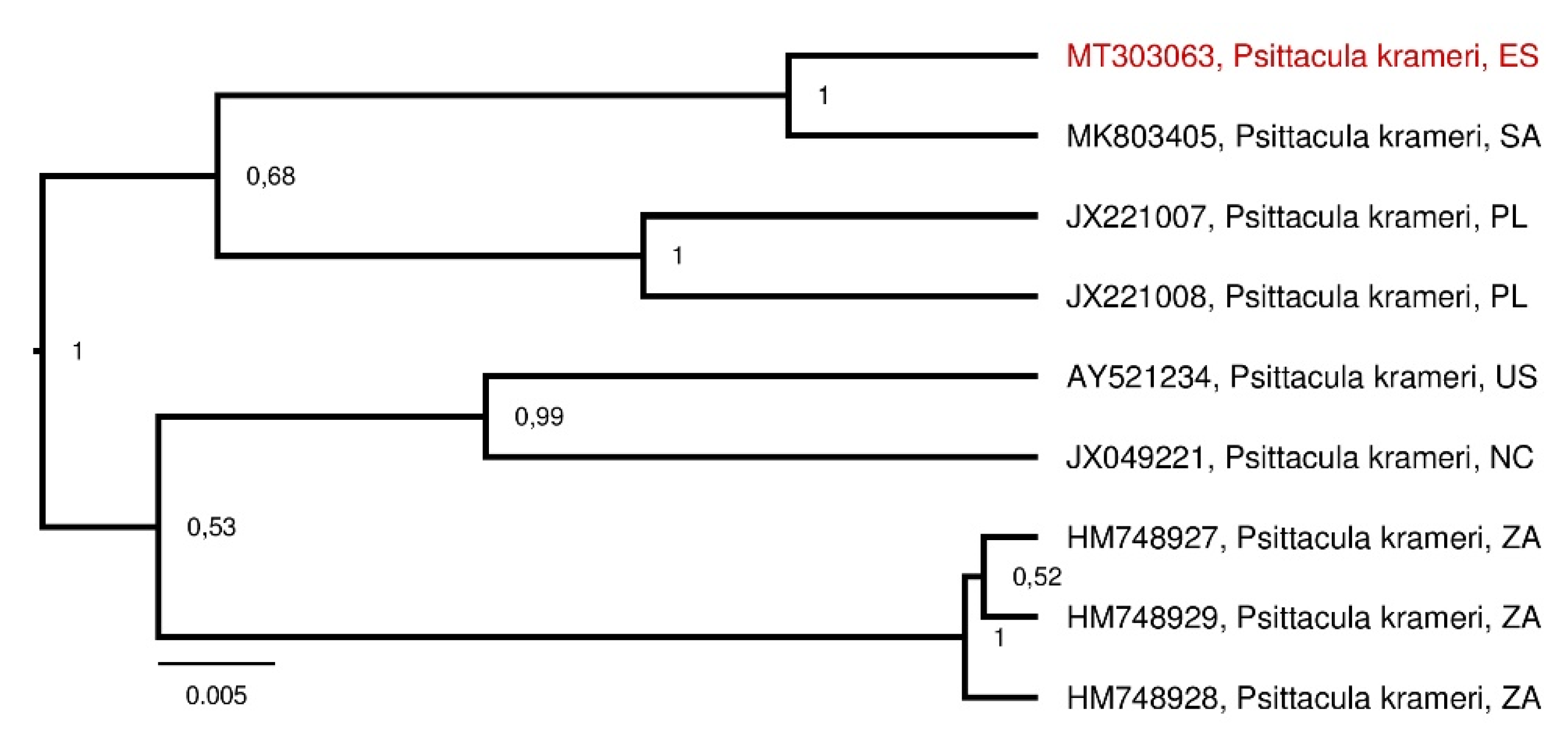
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fogell, D.J.; Martin, R.O.; Groombridge, J.J. Beak and feather disease virus in wild and captive parrots: An analysis of geographic and taxonomic distribution and methodological trends. Arch. Virol. 2016, 161, 2059–2074. [Google Scholar] [CrossRef] [Green Version]
- Raidal, S.R.; Peters, A. Psittacine beak and feather disease: Ecology and implications for conservation. Emu 2018, 118, 80–93. [Google Scholar] [CrossRef]
- Sarker, S.; Patterson, E.I.; Peters, A.; Baker, G.B.; Forwood, J.K.; Ghorashi, S.A.; Holdsworth, M.; Baker, R.; Murray, N.; Raidal, S.R. Mutability dynamics of an emergent single stranded DNA virus in a naïve host. PLoS ONE 2014, 9, e85370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pass, D.A.; Perry, R.A. The pathology of psittacine beak and feather disease. Aust. Vet. J. 1984, 61, 69–74. [Google Scholar] [CrossRef]
- Harkins, G.W.; Martin, D.P.; Christoffels, A.; Varsani, A. Towards inferring the global movement of beak and feather disease virus. Virology 2014, 450, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Warburton, L.; Perrin, M. Evidence of psittacine beak and feather disease in wild Black-cheeked Lovebirds in Zambia. Papageien 2002, 5, 166–169. [Google Scholar]
- Regnard, G.L.; Boyes, R.S.; Martin, R.O.; Hitzeroth, I.I.; Rybicki, E.P. Beak and feather disease viruses circulating in Cape parrots (Poicepahlus robustus) in South Africa. Arch. Virol. 2015, 160, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Fogell, D.J.; Groombridge, J.J.; Tollington, S.; Canessa, S.; Henshaw, S.; Zuel, N.; Jones, C.G.; Andrew Greenwood, A.; Ewen, J.G. Hygiene and biosecurity protocols reduce infection prevalence but do not improve fledging success in an endangered parrot. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Faulkes, C.G.; Greenwood, A.G.; Jones, C.G.; Kaiser, P.; Lyne, O.D.; Black, S.A.; Aurelie Chowrimootoo, A.; Groombridge, J.J. Tracking viral evolution during a disease outbreak: The rapid and complete selective sweep of a circovirus in the endangered Echo parakeet. J. Virol. 2012, 86, 5221–5229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, A.V.; Andery, D.A.; Ferreira, F.C., Jr.; Ortiz, M.C.; Marques, M.V.R.; Marin, S.Y.; Vilela, D.A.R.; Resende, J.S.; Resende, M.; Donatti, R.V.; et al. Molecular diagnosis of beak and feather disease in native Brazilian psittacines. Braz. J. Poult. Sci. 2015, 17, 451–458. [Google Scholar] [CrossRef] [Green Version]
- González-Hein, G.; Gil, I.A.; Sanchez, R.; Huaracan, B. Prevalence of Aves Polyomavirus 1 and Beak and Feather Disease Virus From Exotic Captive Psittacine Birds in Chile. J. Avian Med. Surg. 2019, 33, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Massaro, M.; Ortiz-Catedral, L.; Julian, L.; Galbraith, J.; Kurenbach, B.; Kearvell, J.; Kemp, J.; Hal, J.; Elkington, S.; Taylor, G.; et al. Molecular characterisation of beak and feather disease virus (BFDV) in New Zealand and its implications for managing an infectious disease. Arch. Virol. 2012, 157, 1651–1663. [Google Scholar] [CrossRef] [PubMed]
- Varsani, A.; Regnard, G.L.; Bragg, R.; Hitzeroth, I.I.; Rybicki, E.P. Global genetic diversity and geographical and host-species distribution of beak and feather disease virus isolates. J. Gen. Virol. 2011, 92, 752–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardador, L.; Lattuada, M.; Strubbe, D.; Tella, J.L.; Reino, L.; Figueira, R.; Carrete, M. Regional bans on wild-bird trade modify invasion risks at a global scale. Conserv. Lett. 2017, 10, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.; Reino, L.; Schindler, S.; Strubbe, D.; Vall-llosera, M.; Bastos Araújo, M.; Capinha, C.; Carrete, M.; Mazzoni, S.; Monteiro, M.; et al. Trends in legal and illegal trade of wild birds: A global assessment based on expert knowledge. Biodivers. Conserv. 2019, 28, 3343–3369. [Google Scholar] [CrossRef]
- Fogell, D.J.; Martin, R.O.; Bunbury, N.; Lawson, B.; Sells, J.; McKeand, A.M.; Tatayah, V.; Trung, C.T.; Groombridge, J.J. Trade and conservation implications of new beak and feather disease virus detection in native and introduced parrots. Conserv. Biol. 2018, 32, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Amery-Gale, J.; Marenda, M.S.; Owens, J.; Eden, P.A.; Browning, G.F.; Devlin, J.M. A high prevalence of beak and feather disease virus in non-psittacine Australian birds. J. Med. Microbiol. 2017, 66, 1005–1013. [Google Scholar] [CrossRef]
- Rahaus, M.; Wolff, M.H. Psittacine beak and feather disease: A first survey of the distribution of beak and feather disease virus inside the population of captive psittacine birds in Germany. J. Vet. Med. B 2003, 50, 368–371. [Google Scholar] [CrossRef]
- Abellán, P.; Tella, J.L.; Carrete, M.; Cardador, L.; Anadón, J.D. Climatic matching drives spread rate but not establishment success in recent unintentional bird introductions. PNAS 2017, 114, 9385–9390. [Google Scholar] [CrossRef] [Green Version]
- Jackson, H.; Strubbe, D.; Tollington, S.; Prys-Jones, R.; Matthysen, E.; Groombridge, J.J. Ancestral origins and invasion pathways in a globally invasive bird correlate with climate and influences from bird trade. Mol. Ecol. 2015, 24, 4269–4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardador, L.; Carrete, M.; Gallardo, B.; Tella, J.L. Combining trade data and niche modelling improves predictions of the origin and distribution of non-native European populations of a globally invasive species. J. Biogeogr. 2016, 43, 967–978. [Google Scholar] [CrossRef]
- Edelaar, P.; Roques, S.; Hobson, E.A.; Gonçalves da Silva, A.; Avery, M.L.; Russello, M.A.; Senar, J.C.; Wright, T.F.; Carrete, M.; Tella, J.L. Shared genetic diversity across the global invasive range of the Monk Parakeet suggests a common restricted geographic origin and the possibility of convergent selection. Mol. Ecol. 2015, 24, 2164–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumschick, S.; Blackburn, T.M.; Richardson, D.M. Managing alien bird species: Time to move beyond “100 of the worst” lists? Bird Conserv. Int. 2016, 26, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Avery, M.L.; Shiels, A.B. Monk and rose-ringed parakeets. In Ecology and Management of Terrestrial Vertebrate Invasive Species in the United States, 1st ed.; Pitt, W.C., Beasley, J.C., Witmer, G.W., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 333–358. [Google Scholar]
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pyšek, P.; Vilà, M. More than “100 worst” alien species in Europe. Biol. Invasions 2018, 20, 1611–1621. [Google Scholar] [CrossRef] [Green Version]
- Mazza, G.; Tricarico, E.; Genovesi, P.; Gherardi, F. Biological invaders are threats to human health: An overview. Ethol. Ecol. Evol. 2014, 26, 112–129. [Google Scholar] [CrossRef]
- Mori, E.; Meini, S.; Strubbe, D.; Ancillotto, L.; Sposimo, P.; Menchetti, M. Do alien free-ranging birds affect human health? A global summary of known zoonoses. In Invasive Species and Human Health, 1st ed.; Mazza, G., Tricarcio, E., Eds.; CABI Editions: Wallingford, UK, 2018; pp. 120–129. [Google Scholar]
- Sa, R.C.; Cunningham, A.A.; Dagleish, M.P.; Wheelhouse, N.; Pocknell, A.; Borel, N.; Hannah, L.; Peck, H.L.; Lawson, B. Psittacine beak and feather disease in a free-living ring-necked parakeet (Psittacula krameri) in Great Britain. Eur. J. Wildl. Res. 2014, 60, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Kessler, S.; Heenemann, K.; Krause, T.; Twietmeyer, S.; Fuchs, J.; Lierz, M.; Corman, V.M.; Vahlenkamp, T.M.; Rubbenstroth, D. Monitoring of free-ranging and captive Psittacula populations in Western Europe for avian bornaviruses, circoviruses and polyomaviruses. Avian Pathol. 2020, 49, 119–130. [Google Scholar] [CrossRef]
- Hernández-Brito, D.; Carrete, M.; Popa-Lisseanu, A.; Ibáñez, C.; Tella, J.L. Crowding in the city: Losing and winning competitors of an invasive bird. PLoS ONE 2014, 9, e100593. [Google Scholar] [CrossRef]
- Ypelaar, I.; Bassami, M.R.; Wilcox, G.E.; Raidal, S.R. A universal polymerase chain reaction for the detection of psittacine beak and feather disease virus. Vet. Microbiol. 1999, 68, 141–148. [Google Scholar] [CrossRef]
- Ritchie, P.A.; Anderson, I.L.; Lambert, D.M. Evidence for specificity of psittacine beak and feather disease viruses among avian hosts. Virology 2003, 306, 109–115. [Google Scholar] [CrossRef]
- Sergeant, E. 2018. Epitools Epidemiological Calculators. Ausvet Pty Ltd. Available online: http://epitools.ausvet.com.au/ (accessed on 15 April 2020).
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, W29–W33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. Figtree Software (v.1.4.4). 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 February 2020).
- Genovart, M.; Negre, N.; Tavecchia, G.; Bistuer, A.; Parpal, L.; Oro, D. The young, the weak and the sick: Evidence of natural selection by predation. PLoS ONE 2010, 5, e9774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.J.; Anderson, I.L.; Alley, M.R.; Springett, B.P.; Gartrell, B.D. The prevalence of beak and feather disease virus infection in wild populations of parrots and cockatoos in New Zealand. N. Z. Vet. J. 2007, 55, 235–238. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; McInnes, K.; Hauber, M.E.; Brunton, D.H. First report of beak and feather disease virus (BFDV) in wild Red-fronted Parakeets (Cyanoramphus novaezelandiae) in New Zealand. Emu 2009, 109, 244–247. [Google Scholar] [CrossRef]
- Eastwood, J.R.; Berg, M.L.; Ribot, R.F.; Buchanan, K.L.; Walder, K.; Bennett, A.T. Prevalence of BFDV in wild breeding Platycercus elegans. J. Ornithol. 2019, 160, 557–565. [Google Scholar] [CrossRef]
- Sarker, S.; Moylan, K.G.; Ghorashi, S.A.; Forwood, J.K.; Peters, A.; Raidal, S.R. Evidence of a deep viral host switch event with beak and feather disease virus infection in rainbow bee-eaters (Merops ornatus). Sci. Rep. 2015, 5, 14511. [Google Scholar] [CrossRef] [Green Version]
- Kalodimos, N. First account of a nesting population of Monk parakeets (Myiopsitta monachus) with nodule-shaped bill lesions in Katehaki, Athens, Greece. Bird Popul. 2013, 12, 1–6. [Google Scholar]
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.J.; Abbud, M.C.; Aguilar, J.M.R.E.; Alemán, U.; Aramburú, R.M.; Arce Arias, A.; Balas McNab, R.; Balsby, T.J.S.; et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef] [Green Version]
- CITES Trade Database. Available online: https://trade.cites.org/ (accessed on 10 April 2020).
- Fèvre, E.M.; Bronsvoort, B.M.D.C.; Hamilton, K.A.; Cleaveland, S. Animal movements and the spread of infectious diseases. Trends Microbiol. 2006, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, M.; Mori, E. Worldwide impact of alien parrots (Aves Psittaciformes) on native biodiversity and environment: A review. Ethol. Ecol. Evol. 2014, 26, 172–194. [Google Scholar] [CrossRef]
- Sarker, S.; Lloyd, C.; Forwood, J.; Raidal, S.R. Forensic genetic evidence of beak and feather disease virus infection in a powerful owl (Ninox strenua). Emu 2016, 116, 71–74. [Google Scholar] [CrossRef]
- Circella, E.; Legretto, M.; Pugliese, N.; Caroli, A.; Bozzo, G.; Accogli, G.; Lavazza, A.; Camarda, A. Psittacine Beak and Feather Disease–like Illness in Gouldian Finches (Chloebia gouldiae). Avian Dis. 2014, 58, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Mori, E.; Malfatti, L.; Le Louarn, M.; Brito, D.H.; Ten Cate, B.; Ricci, M.; Menchetti, M. ‘Some like it alien’: Predation on invasive ring–necked parakeets by the long–eared owl in an urban area. Anim. Biodiv. Conserv. 2020, 43, 151–158. [Google Scholar]
- Hernández-Brito, D.; Blanco, G.; Tella, J.L.; Carrete, M.A. protective nesting association with native species counteracts biotic resistance for the spread of an invasive parakeet from urban to rural habitats. Front. Zool. 2020, 17, 1–13. [Google Scholar] [CrossRef]
- Abellán, P.; Carrete, M.; Anadón, J.D.; Cardador, L.; Tella, J.L. Non-random patterns and temporal trends (1912-2012) in the transport, introduction and establishment of exotic birds in Spain and Portugal. Divers. Distrib. 2016, 22, 263–273. [Google Scholar] [CrossRef]
- Turbé, A.; Strubbe, D.; Mori, E.; Carrete, M.; Chiron, F.; Clergeau, P.; González-Moreno, P.; Le Louarn, M.; Luna, A.; Menchetti, M.; et al. Assessing the assessments: Evaluation of four impact assessment protocols for invasive alien species. Divers. Distrib. 2017, 23, 297–307. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| MT303063 (ES) | MK803405 (SA) | JX221007 (PL) | JX221008 (PL) | AY521234 (US) | JX049221 (NC) | HM748927 (ZA) | HM748929 (ZA) | HM748928 (ZA) | |
|---|---|---|---|---|---|---|---|---|---|
| MT303063 (ES) | - | 1 (0.5) | 11 (5.4) | 4 (1.9) | 11 (5.4) | 7 (3.5) | 7 (3.5) | 7 (3.5) | 8 (4.0) |
| MK803405 (SA) | 9 (1.5) | - | 10 (5.0) | 3 (1.5) | 10 (5.0) | 6 (3.0) | 8 (4.0) | 8 (4.0) | 9 (4.5) |
| JX221007 (PL) | 39 (6.5) | 33 (5.5) | - | 9 (4.5) | 16 (8.0) | 12 (6.0) | 14 (7.0) | 14 (7.0) | 15 (7.5) |
| JX221008 (PL) | 31 (5.1) | 25 (4.1) | 20 (3.3) | - | 9 (4.5) | 5 (2.5) | 7 (3.5) | 7 (3.5) | 8 (4.0) |
| AY521234 (US) | 33 (5.5) | 27 (4.4) | 46 (7.6) | 36 (6.0) | - | 7 (3.5) | 9 (4.5) | 9 (4.5) | 10 (5.0) |
| JX049221 (NC) | 28 (4.6) | 22 (3.6) | 44 (7.3) | 32 (5.3) | 22 (3.6) | - | 6 (3.0) | 6 (3.0) | 7 (3.5) |
| HM748927 (ZA) | 31 (5.1) | 31 (5.1) | 47 (7.8) | 40 (6.6) | 34 (5.6) | 33 (5.5) | - | 0 (0.0) | 1 (0.5) |
| HM748929 (ZA) | 32 (5.3) | 31 (5.1) | 47 (7.8) | 40 (6.6) | 33 (5.5) | 34 (5.6) | 1 (0.2) | - | 1 (0.5) |
| HM748928 (ZA) | 33 (5.5) | 33 (5.5) | 49 (8.1) | 42 (7.0) | 36 (6.0) | 33 (5.5) | 2 (0.3) | 3 (0.5) | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morinha, F.; Carrete, M.; Tella, J.L.; Blanco, G. High Prevalence of Novel Beak and Feather Disease Virus in Sympatric Invasive Parakeets Introduced to Spain From Asia and South America. Diversity 2020, 12, 192. https://0-doi-org.brum.beds.ac.uk/10.3390/d12050192
Morinha F, Carrete M, Tella JL, Blanco G. High Prevalence of Novel Beak and Feather Disease Virus in Sympatric Invasive Parakeets Introduced to Spain From Asia and South America. Diversity. 2020; 12(5):192. https://0-doi-org.brum.beds.ac.uk/10.3390/d12050192
Chicago/Turabian StyleMorinha, Francisco, Martina Carrete, José L. Tella, and Guillermo Blanco. 2020. "High Prevalence of Novel Beak and Feather Disease Virus in Sympatric Invasive Parakeets Introduced to Spain From Asia and South America" Diversity 12, no. 5: 192. https://0-doi-org.brum.beds.ac.uk/10.3390/d12050192