Description of Echolocation Call Parameters for Urban Bats in Vietnam as a Step Towards a More Integrated Acoustic Monitoring of Urban Wildlife in Southeast Asia


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bat Captured
2.2. Species Identification
2.3. Acoustics Recording and Data Analysis
3. Results
3.1. The Species Diversity and Their Acoustic Parameters
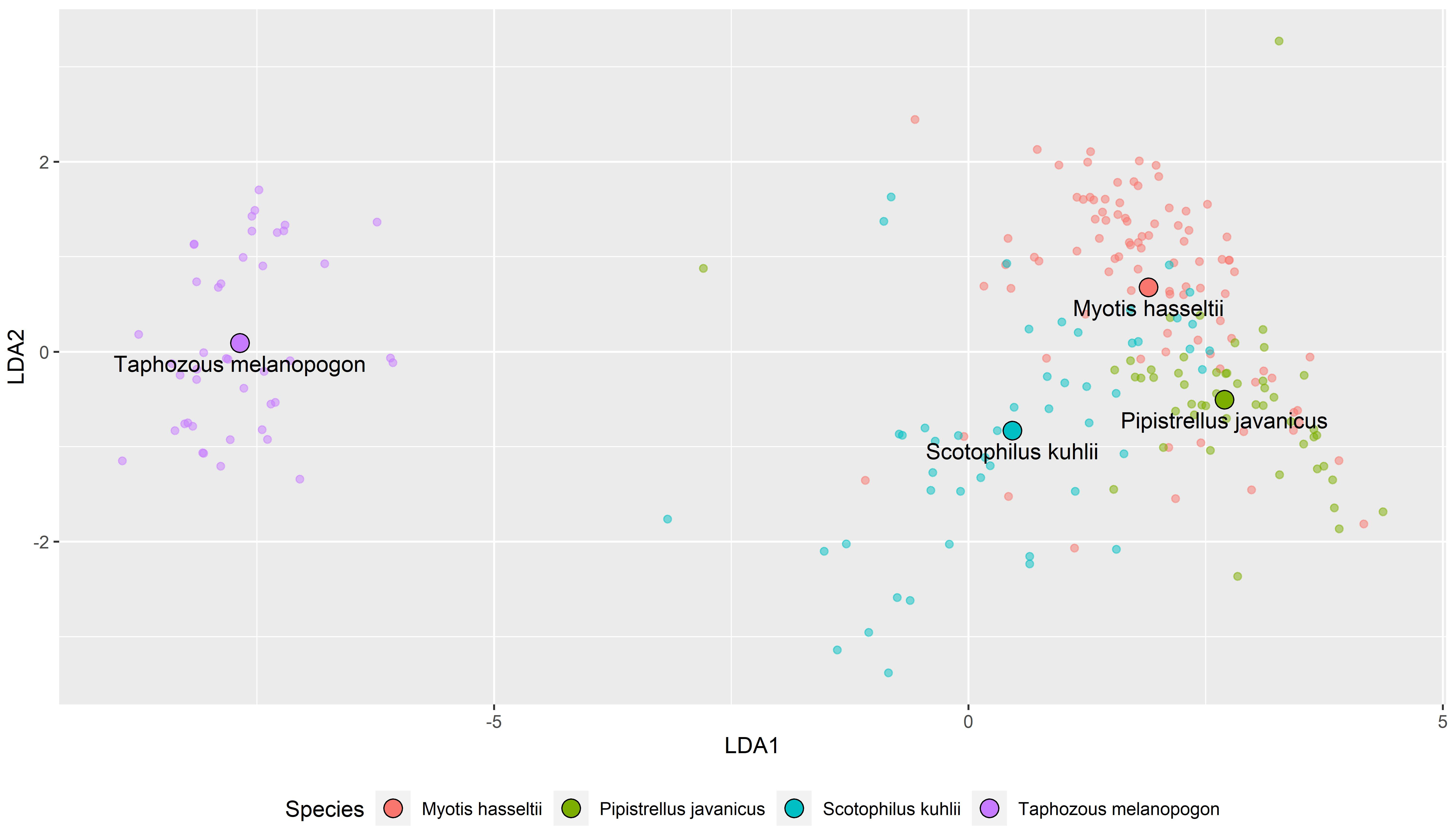
3.2. The Echolocation Call Classification by Discriminant Function Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kruskop, S.V. Bats of Vietnam: Checklist and an Identification Manual; Joint Russian-Vietnamese Science and Technological Tropical Centre: Moscow, Russia, 2013; p. 300. [Google Scholar]
- Thong, V.D.; Mao, X.; Csorba, G.; Bates, P.; Ruedi, M.; Van Viet, N.; Loi, D.N.; Van Nha, P.; Chachula, O.; Tuan, T.A.; et al. First Records of Myotis altarium (Chiroptera: Vespertilionidae) from India and Vietnam. Mammal. Study 2018, 43, 1–7. [Google Scholar] [CrossRef]
- Kruskop, S.V.; Borissenko, A.V. A New Species of South-East AsianMyotis(Chiroptera: Vespertilionidae), with Comments on Vietnamese “Whiskered Bats”. Acta Chiropterologica 2013, 15, 293–305. [Google Scholar] [CrossRef]
- Soisook, P.; Karapan, S.; Satasook, C.; Thong, V.D.; Khan, F.A.A.; Maryanto, I.; Csorba, G.; Furey, N.; Aul, B.; Bates, P.J. A Review of the Murina cyclotis Complex (Chiroptera: Vespertilionidae) with Descriptions of a New Species and Subspecies. Acta Chiropterologica 2013, 15, 271–292. [Google Scholar] [CrossRef]
- Son, N.T.; Görföl, T.; Francis, C.M.; Motokawa, M.; Estók, P.; Endo, H.; Thong, V.D.; Dang, N.X.; Oshida, T.; Csorba, G. Description of a New Species ofMyotis(Vespertilionidae) from Vietnam. Acta Chiropterologica 2013, 15, 473–483. [Google Scholar] [CrossRef]
- Görföl, T.; Kruskop, S.V.; Tu, V.T.; Estók, P.; Son, N.T.; Csorba, G. A new genus of vespertilionid bat: The end of a long journey for Joffre’s Pipistrelle (Chiroptera: Vespertilionidae). J. Mammal. 2020, 101, 331–348. [Google Scholar] [CrossRef]
- Furey, N.M.; Mackie, I.J.; Racey, P.A. Bat diversity in Vietnamese limestone karst areas and the implications of forest degradation. Biodivers. Conserv. 2010, 19, 1821–1838. [Google Scholar] [CrossRef]
- Thong, V.D.; Dietz, C.; Denzinger, A.; Bates, P.J.J.; Furey, N.M.; Csorba, G.; Glenn, M.E.; Hoye; Thuy, L.D.; Schnitzler, H.-U. Further Records of Murina tiensa from Vietnam with first information on its echolocation calls. Hystrix It. J. Mammalog. 2011, 22, 129–138. [Google Scholar]
- Thong, V.D. Bats of Cat Tien national park: Diversity, echolocation and taxonomic remarks. TAP CHI SINH HOC 2016, 37, 336–343. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef]
- Jones, G.; Jacobs, D.S.; Kunz, T.H.; Willig, M.R.; Racey, P.A. Carpe noctem: The importance of bats as bioindicators. Endanger. Species Res. 2009, 8, 93–115. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.C.; Scholl, J.M.; Bauer, J.; Teige, T.; Yovel, Y.; Kramer-Schadt, S.; Gras, P. Movement responses of common noctule bats to the illuminated urban landscape. Landsc. Ecol. 2019, 35, 189–201. [Google Scholar] [CrossRef]
- Jung, K.; Kalko, E.K.V. Adaptability and vulnerability of high flying Neotropical aerial insectivorous bats to urbanization. Divers. Distrib. 2011, 17, 262–274. [Google Scholar] [CrossRef]
- Lane, D.J.; Kingston, T.; Lee, B.P.-H. Dramatic decline in bat species richness in Singapore, with implications for Southeast Asia. Biol. Conserv. 2006, 131, 584–593. [Google Scholar] [CrossRef]
- General Statistics Office. Vietnam Population and Housing Census 2009—Migration and Urbanization in Vietnam: Patterns, Trend and Differentials; Ministry of Planning and Investment: Hanoi, Vietnam, 2011; p. 140. [Google Scholar]
- Nong, D.H.; Fox, J.M.; Miura, T.; Saksena, S. Built-up Area Change Analysis in Hanoi Using Support Vector Machine Classification of Landsat Multi-Temporal Image Stacks and Population Data. Land 2015, 4, 1213–1231. [Google Scholar] [CrossRef] [Green Version]
- MacSwiney, M.C.; Clarke, F.M.; Racey, P.A. What you see is not what you get: the role of ultrasonic detectors in increasing inventory completeness in Neotropical bat assemblages. J. Appl. Ecol. 2008, 45, 1364–1371. [Google Scholar] [CrossRef]
- Avila-Flores, R.; Fenton, M.B. Use of spatial features by foraging insectivorous bats in a large urban landscape. J. Mammal. 2005, 86, 1193–1204. [Google Scholar] [CrossRef]
- Straka, T.M.; Lentini, P.E.; Lumsden, L.F.; Wintle, B.A.; Van Der Ree, R. Urban bat communities are affected by wetland size, quality, and pollution levels. Ecol. Evol. 2016, 6, 4761–4774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straka, T.; Wolf, M.; Gras, P.; Buchholz, S.; Voigt, C.C. Tree Cover Mediates the Effect of Artificial Light on Urban Bats. Front. Ecol. Evol. 2019, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Threlfall, C.G. Trait-dependent tolerance of bats to urbanization: A global meta-analysis. In Proceedings of the Proceedings of the Royal Society of London. Series B: Biological Sciences; The Royal Society: London, UK, 2018; Volume 285, p. 20181222. [Google Scholar]
- Voigt, C.C.; Kingston, T. Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer Science and Business Media LLC: Berlin, Germany, 2016. [Google Scholar]
- Gallo, T.; Lehrer, E.W.; Fidino, M.; Kilgour, R.J.; Wolff, P.J.; Magle, S.B. Need for multiscale planning for conservation of urban bats. Conserv. Biol. 2018, 32, 638–647. [Google Scholar] [CrossRef]
- Jones, G.; Vaughan, N.; Parsons, S. Acoustic identification of bats from directly sampled and time expanded recordings of vocalizations. Acta Chiropterologica 2000, 22, 155–170. [Google Scholar]
- Wu, H.; Jiang, T.; Liu, S.; Lu, G.; Feng, J. Acoustic identification of two morphologically similar bat species, Miniopterus magnater and Miniopterus fuliginosus (Chiroptera, Miniopteridae). Mamalia 2020, 84, 201–206. [Google Scholar] [CrossRef]
- Thong, V.D. Acoustic identification and taxonomic remarks of Hipposiderids and Rhinolophids (Chiroptera: Hipposideridae, Rhinolophidae) in Tam Dao national park, Northeastern Nietnam. TAP CHI SINH HOC 2015, 36, 487–493. [Google Scholar] [CrossRef]
- Hughes, A.C.; Satasook, C.; Bates, P.J.J.; Soisook, P.; Sritongchuay, T.; Jones, G.; Bumrungsri, S. Using Echolocation Calls to Identify Thai Bat Species: Vespertilionidae, Emballonuridae, Nycteridae and Megadermatidae. Acta Chiropterologica 2011, 13, 447–455. [Google Scholar] [CrossRef]
- Wordley, C.F.R.; Foui, E.K.; Mudappa, D.; Sankaran, M.; Altringham, J.D. Acoustic Identification of Bats in the Southern Western Ghats, India. Acta Chiropterologica 2014, 16, 213–222. [Google Scholar] [CrossRef]
- Sophany, P.; Sarith, P.; Furey, N.M. Cambodian bat echolocation: A first description of assemblage call parameters and assessment of their utility for species identification. Cambod. J. Nat. History 2013, 2013, 16–26. [Google Scholar]
- Faraway, J. Practical Regression and Anova Using R. Available online: https://cran.r-project.org/doc/contrib/Faraway-PRA.pdf (accessed on 15 August 2020).
- Hair, J.F.; Black, W.C.; Babin, B.J.; Anderson, R.E. Multivariate Data Analysis, 7th ed.; Pearson: New York, NY, USA, 2009; p. 816. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 30 July 2020).
- Francis, C. A Guide to the Mammals of South-East Asia; Princeton University Press: Princeton, NJ, USA, 2008; p. 392. [Google Scholar]
- Tu, V.T. Final Project Report: Conservation of Bat Assemblages in Hanoi City, Vietnam (RGS 55.02.0); Institute of Ecology and Biological Resources: Hanoi, Vietnam, 2010. [Google Scholar]
- Tu, V.T.; Arai, S.; Kikuchi, F.; Hang, C.T.; Tuan, T.A.; Csorba, G.; Görföl, T. Rediscovery of Van Hasselt’s Mouse-eared Bat Myotis hasseltii (Temminck, 1840) and its first genetic data from Hanoi, northern Vietnam. J. Threat. Taxa 2019, 11, 13915–13919. [Google Scholar] [CrossRef]
- Heller, K.G. Echolocation calls of Malaysian bats. Säugetierkunde 1989, 54, 1–8. [Google Scholar]
- Sơn, N.T.; O’Shea, T.J.; Gore, J.A.; Csorba, G.; Tu, V.T.; Oshida, T.; Endo, H.; Motokawa, M. Bats (Mammalia: Chiroptera) of the southeastern Truong Son Mountains, Quang Ngai Province, Vietnam. J. Threat. Taxa 2016, 8, 8953–8969. [Google Scholar] [CrossRef] [Green Version]
- Habitat and Acoustic Survey and an Action Plan for Bats Conservation in the Kathmandu Valley, Nepal. Available online: https://smcrf.org/project/habitat-and-acoustic-survey-and-an-action-plan-for-bats-conservation-in-the-kathmandu-valley-nepal/ (accessed on 27 September 2020).
- Raghuram, H.; Jain, M.; Balakrishnan, R. Species and acoustic diversity of bats in a palaeotropical wet evergreen forest in southern India. Curr. Sci. 2014, 107, 631–641. [Google Scholar]
- Zhu, G.; Chmura, A.A.; Zhang, L. Morphology, Echolocation Calls and Diet of Scotophilus kuhlii (Chiroptera: Vespertilionidae) on Hainan Island, South China. Acta Chiropterologica 2012, 14, 175–181. [Google Scholar] [CrossRef]
- Thomas, D.W.; Bell, G.P.; Fenton, M.B. Variation in Echolocation Call Frequencies Recorded from North American Vespertilionid Bats: A Cautionary Note. J. Mammal. 1987, 68, 842–847. [Google Scholar] [CrossRef]
- Wund, M.A. Variation in the Echolocation Calls of Little Brown Bats (Myotis lucifugus) in Response to Different Habitats. Am. Midl. Nat. 2006, 156, 99–108. [Google Scholar] [CrossRef]
- Wei, L.; Han, N.; Zhang, L.; Helgen, K.M.; Parsons, S.; Zhou, S.; Zhang, S. Wing morphology, echolocation calls, diet and emergence time of black-bearded tomb bats (Taphozous melanopogon, Emballonuridae) from southwest China. Acta Chiropterologica 2008, 10, 51–59. [Google Scholar] [CrossRef]
- Schnitzler, H.-U.; Moss, C.F.; Denzinger, A. From spatial orientation to food acquisition in echolocating bats. Trends Ecol. Evol. 2003, 18, 386–394. [Google Scholar] [CrossRef]


{kind=link}
| # | City | Latitude | Longitude | Time | # Net | # Night | Note |
|---|---|---|---|---|---|---|---|
| 1 | Tra Vinh | 9.921870008 | 106.346854 | May 2020 | 1 | 2 | Indoor parking area |
| 2 | Tra Vinh | 10.82854802 | 106.7282675 | May 2020 | 2 | 1 | A small pond |
| 3 | Tra Vinh | 10.87740471 | 106.6799775 | August 2019 May 2020 | 2 | 4 | Grassland with some trees |
| 4 | Tra Vinh | 9.922454478 | 106.3476558 | May 2020 | 1 | 1 | Under the bridge |
| 5 | Tra Vinh | 9.919190397 | 106.349743 | May 2020 | 1 | 1 | Abondance house |
| 6 | Ho Chi Minh | 9.923449242 | 106.3434506 | May 2020 | 1 | 1 | Old building |
| 7 | Ho Chi Minh | 9.938479504 | 106.337781 | June 2020 | 2 | 1 | Near Nhieu Loc canal |
| 8 | Ho Chi Minh | 10.78259474 | 106.7016182 | July 2020 | 2 | 1 | Small canal |
| 9 | Ho Chi Minh | 10.79325811 | 106.6957258 | July 2020 | 2 | 1 | Small canal |
| 10 | Ho Chi Minh | 10.76487179 | 106.6356857 | July 2020 | 2 | 1 | Small canal |
| 11 | Ho Chi Minh | 10.76132323 | 106.618494 | June 2020 | 1 | 2 | Small canal |
| Species | Location | d (ms) | FMAXE (kHz) | hf (kHz) | lf (kHz) | sf (kHz) | tf (kHz) | fr (kHz) | N | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Scotophilus kuhlii | Vietnam | 5.48 ± 23.8 | 45.64 ± 3.32 | 76.27 ± 6.56 | 38.94 ± 2.40 | 60.41 ± 7.21 | 42.66 ± 3.18 | 37.32 ± 7.06 | 49 | This study |
| Scotophilus kuhlii | India | 2.96 ± 0.38 | 45.26 ± 0.77 | N/a | N/a | 56.67 ± 2.06 | 43.53 ± 0.76 | 13.1 ± 2.25 | 2 | [1] |
| Taphozous melanopogon | Vietnam | 6.45 ± 1.13 | 33.07 ± 5.90 | 59.10 ± 5.11 | 23.19 ± 1.25 | 32.70 ± 3.27 | 31.09 ± 5.12 | 35.91 ± 5.22 | 43 | This study |
| Taphozous melanopogon | Thailand | 6.02 ± 3.4 | 29.71 ± 2.67 | 76.15 ± 20.18 | 20.37 ± 6.2 | 36.6 ± 10.44 | 22.58 ± 5.58 | 55.78 ± 20.32 | 33 | [2] |
| Taphozous melanopogon | Malaysia | 6–14 | N/a | N/a | N/a | 26–30 | 24–26 | N/a | N/a | [3] |
| Pipistrellus javanicus | Vietnam | 4.78 ± 1.06 | 50.30 ± 5.09 | 88.27 ± 3.50 | 42.27 ± 2.66 | 68.23 ± 4.05 | 48.03 ± 3.61 | 46.00 ± 3.48 | 50 | This study |
| Pipistrellus javanicus | Nepal | 3.84 | 36.85 | 42 | 36.19 | 52.81 | [4] | |||
| Myotis hasseltii | Vietnam | 3.86 ± 0.50 | 50.06 ± 3.09 | 84.63 ± 5.42 | 38.82 ± 3.12 | 67.55 ± 3.75 | 47.80 ± 2.31 | 45.81 ± 6.31 | 91 | This study |
| Myotis hasseltii | Malaysia | 2.5–5.5 | N/a | N/a | N/a | 82-104 | N/a | N/a | N/a | [3] |
| Acoustic Parameter | Pipistrellus javanicus | Scotophilus kuhlii | Myotis hasseltii | Taphozous melanopogon |
|---|---|---|---|---|
| duration | −7.1731 (0) * | −7.4182 (0) * | −9.8947 (0) * | −3.8096 (0.0) * |
| FMAXE(kHz) | 2.691 (0.01) * | 3.6796 (0.001) * | −0.4311 (0.67) | 0.5517 (0.58) |
| hf (kHz) | −1.4085 (0.17) | 5.9849 (0) * | 1.7652 (0.08) | 0.8711 (0.39) |
| lf (kHz) | −1.3701 (0.19) | −1.9089 (0.07) | −7.4933 (0) * | −8.9108 (0.0) * |
| sf (kHz) | −0.0027 (0.99) | 3.2563 (0.002) * | 2.2077 (0) * | −2.6367 (0.01) * |
| tf (kHz) | −0.0937 (0.92) | 3.7237 (0.001) * | 0.7845 (0.43) | 0.8740 (0.38) |
| fr (kHz) | −0.4044 (0.69) | 7.4684 (0) * | 4.692 (0) * | 2.3334 (0.02) * |
| True Group | |||||
|---|---|---|---|---|---|
| As predicted group | Myotis hasseltii | Pipistrellus javanicus | Scotophilus kuhlii | Taphozous melanopogon | Sum |
| Myotis hasseltii | 73 | 14 | 15 | 102 | |
| Pipistrellus javanicus | 13 | 35 | 1 | 49 | |
| Scotophilus kuhlii | 5 | 1 | 33 | 39 | |
| Taphozous melanopogon | 0 | 0 | 43 | 43 | |
| Total | 91 | 50 | 49 | 43 | 233 |
| n correct | 73 | 35 | 33 | 43 | 184 |
| % correct | 80.80 | 70.00 | 67.34 | 100.0 | 78.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, L.K.; Tran, B.V.; Le, Q.T.; Nguyen, T.T.; Voigt, C.C. Description of Echolocation Call Parameters for Urban Bats in Vietnam as a Step Towards a More Integrated Acoustic Monitoring of Urban Wildlife in Southeast Asia. Diversity 2021, 13, 18. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010018
Pham LK, Tran BV, Le QT, Nguyen TT, Voigt CC. Description of Echolocation Call Parameters for Urban Bats in Vietnam as a Step Towards a More Integrated Acoustic Monitoring of Urban Wildlife in Southeast Asia. Diversity. 2021; 13(1):18. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010018
Chicago/Turabian StylePham, Long Kim, Bang Van Tran, Quy Tan Le, Trung Thanh Nguyen, and Christian C. Voigt. 2021. "Description of Echolocation Call Parameters for Urban Bats in Vietnam as a Step Towards a More Integrated Acoustic Monitoring of Urban Wildlife in Southeast Asia" Diversity 13, no. 1: 18. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010018