
Conserved Mechanisms, Novel Anatomies: The Developmental Basis of Fin Evolution and the Origin of Limbs

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
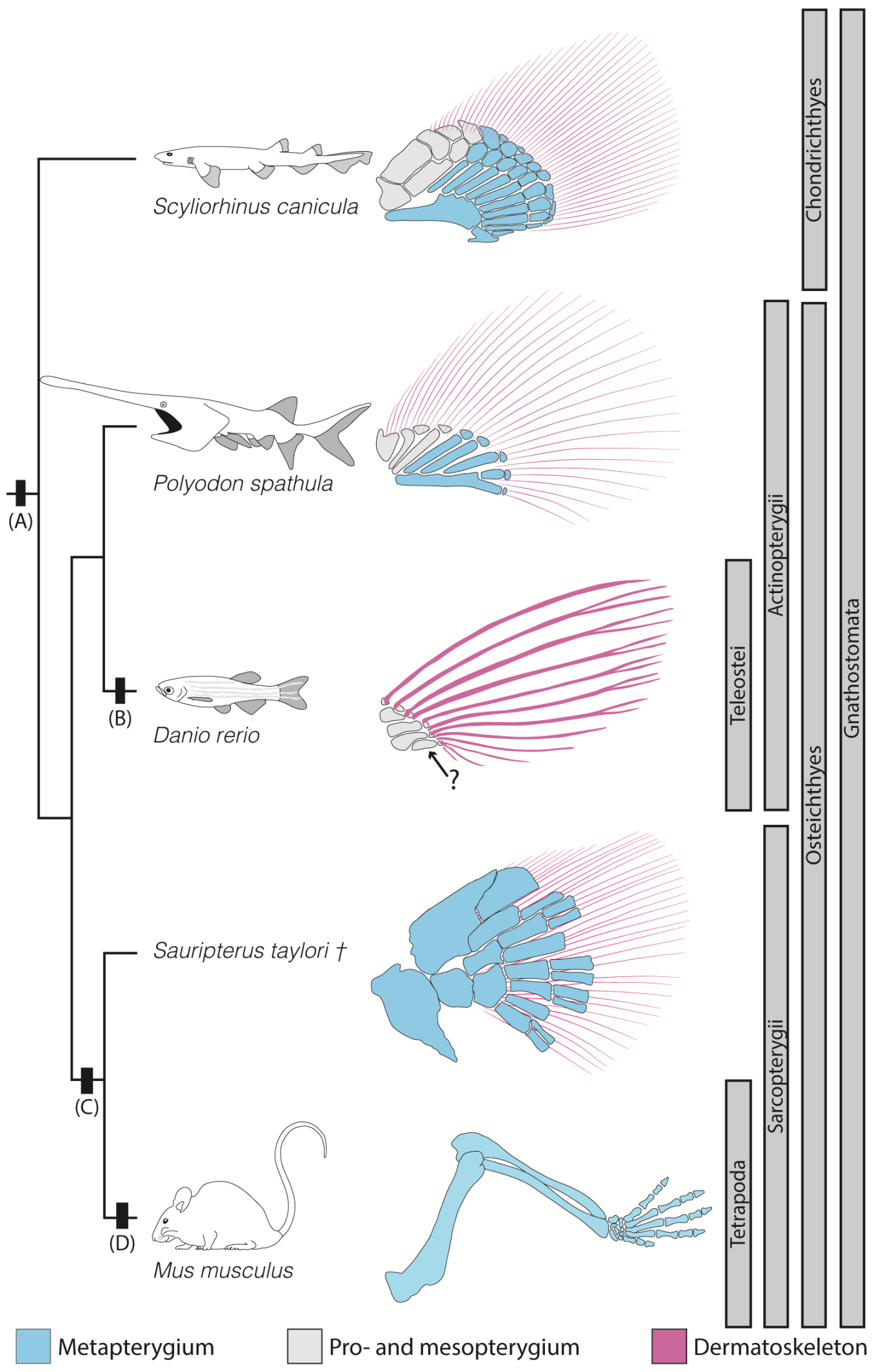
2. Anatomical Context
3. Tetrapod Limb Development
3.1. Integration of Limb Development
3.2. Limb Development: The AER and Fgf–Shh Signaling
3.3. Limb Development: Hox Regulation of Pattern
4. Comparative Developmental Studies Reveal Deep Homologies
4.1. Homologies and Novelty
4.2. Developmental Comparisons: Fgf and Shh
4.3. Developmental Comparisons: Hox Genes
4.4. Recent Functional Studies
5. In Search of Deeper Homologies Still (Deepest Homologies)
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Owen, R. On the Nature of Limbs; John Van Voorst: London, UK, 1849; 119p. [Google Scholar] [CrossRef] [Green Version]
- Balfour, F.M. On the development of the skeleton of the paired fins of Elasmobranchii, considered in relation to its bearings on the nature of the limbs of the Vertebrata. Proc. Zool. Soc. Lond. 1881, 1881, 656–671. [Google Scholar] [CrossRef]
- Davis, M.C. The deep homology of the autopod: Insights from Hox gene regulation. Integr. Comp. Biol. 2013, 53, 224–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Schneider, I.; Shubin, N.H. Evolution: The deep genetic roots of tetrapod-specific traits. Curr. Biol. 2021, 31, R467–R469. [Google Scholar] [CrossRef]
- Gegenbaur, C. Untersuchungen zur Vergleichenden Anatomie der Wirbeltiere; Wilhelm Engelmann: Lepzig, Germany, 1865; Volume II, 176p. [Google Scholar] [CrossRef] [Green Version]
- Coates, M.I. The evolution of paired fins. Theory Biosci. 2003, 122, 266–287. [Google Scholar] [CrossRef]
- Davis, M.C.; Shubin, N.H.; Force, A. Pectoral fin and girdle development in the basal actinopterygians Polyodon spathula and Acipenser transmontanus. J. Morphol. 2004, 628, 608–628. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Yu, X. Stem sarcopterygians have primitive polybasal fin articulation. Biol. Lett. 2009, 5, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Gegenbaur, C. Elements of Comparative Anatomy, 2nd ed.; Macmillan and Co.: London, UK, 1878; 645p. [Google Scholar]
- Hawkins, M.B.; Henke, K.; Harris, M.P. Latent developmental potential to form limb-like skeletal structures in zebrafish. Cell 2021, 184, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H. Approaches to a comparison of fin and limb structure and development. Theory Biosci. 2003, 122, 288–301. [Google Scholar] [CrossRef]
- Goodrich, E.S. Studies on the Structure & Development of Vertebrates; Macmillan and Co. Ltd.: London, UK, 1930; 837p. [Google Scholar]
- Janvier, P. Early Vertebrates; Oxford University Press: Oxford, UK, 1996; 408p. [Google Scholar]
- Davis, M.C.; Shubin, N.H.; Daeschler, E.B. A new specimen of Sauripterus taylori (Sarcopterygii; Osteichthyes) from the Famennian Catskill Formation of North America. J. Vertebr. Paleontol. 2004, 24, 26–40. [Google Scholar] [CrossRef]
- Shubin, N.H.; Daeschler, E.B.; Jenkins, F.A. The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 2006, 440, 764–771. [Google Scholar] [CrossRef]
- Zhang, J.; Wagh, P.; Guay, D.; Sanchez-Pulido, L.; Padhi, B.K.; Korzh, V.; Andrade-Navarro, M.A.; Akimenko, M.A. Loss of fish actinotrichia proteins and the fin-to-limb transition. Nature 2010, 466, 234–237. [Google Scholar] [CrossRef]
- Daeschler, E.B.; Shubin, N. Fish with fingers? Nature 1998, 391, 133. [Google Scholar] [CrossRef]
- Gehrke, A.R.; Schneider, I.; de la Calle-Mustienes, E.; Tena, J.J.; Gomez-Marin, C.; Chandran, M.; Nakamura, T.; Braasch, I.; Postlethwait, J.H.; Gómez-Skarmeta, J.L.; et al. Deep conservation of wrist and digit enhancers in fish. Proc. Natl. Acad. Sci. USA 2015, 112, 803–808. [Google Scholar] [CrossRef] [Green Version]
- Woltering, J.M.; Noordermeer, D.; Leleu, M.; Duboule, D. Conservation and divergence of regulatory strategies at Hox loci and the origin of tetrapod digits. PLoS Biol. 2014, 12, e1001773. [Google Scholar] [CrossRef] [Green Version]
- Shubin, N.H.; Davis, M.C. Modularity in the evolution of vertebrate appendages. In Modularity in Development and Evolution; Schlosser, G., Wagner, G., Eds.; University of Chicago Press: Chicago, IL, USA, 2004; pp. 429–440. [Google Scholar]
- Tulenko, F.J.; Massey, J.L.; Holmquist, E.; Kigundu, G.; Thomas, S.; Smith, S.M.E.; Mazan, S.; Davis, M.C. Fin-fold development in paddlefish and catshark and implications for the evolution of the autopod. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [Green Version]
- Tulenko, F.J.; Augustus, G.J.; Massey, J.L.; Sims, S.E.; Mazan, S.; Davis, M.C. HoxD expression in the fin-fold compartment of basal gnathostomes and implications for paired appendage evolution. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Gehrke, A.R.; Lemberg, J.; Szymaszek, J.; Shubin, N.H. Digits and fin rays share common developmental histories. Nature 2016, 537, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Ahn, D.; Ho, R.K. Tri-phasic expression of posterior Hox genes during development of pectoral fins in zebrafish: Implications for the evolution of vertebrate paired appendages. Dev. Biol. 2008, 322, 220–233. [Google Scholar] [CrossRef] [Green Version]
- Schneider, I.; Shubin, N.H. The origin of the tetrapod limb: From expeditions to enhancers. Trends Genet. 2013, 29, 419–426. [Google Scholar] [CrossRef]
- Cloutier, R.; Clement, A.M.; Lee, M.S.Y.; Noël, R.; Béchard, I.; Roy, V.; Long, J.A. Elpistostege and the origin of the vertebrate hand. Nature 2020, 579, 549–554. [Google Scholar] [CrossRef]
- Wagner, G.P.; Chiu, C.H. The Tetrapod Limb: A Hypothesis on Its Origin. J. Exp. Zool. 2001, 291, 226–240. [Google Scholar] [CrossRef]
- Woltering, J.M.; Duboule, D. The origin of digits: Expression patterns versus regulatory mechanisms. Dev. Cell 2010, 18, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Coates, M.I. The Devonian tetrapod Acanthostega gunnari Jarvik: Postcranialanatomy, basal tetrapod interrelationships and patterns of skeletal evolution. Trans. R. Soc. Edinb. 1996, 87, 363–421. [Google Scholar] [CrossRef]
- Tabin, C.; Wolpert, L. Rethinking the proximodistal axis of the vertebrate limb in the molecular era. Genes Dev. 2007, 21, 1433–1442. [Google Scholar] [CrossRef] [Green Version]
- Delgado, I.; Torres, M. Coordination of limb development by crosstalk among axial patterning pathways. Dev. Biol. 2017, 429, 382–386. [Google Scholar] [CrossRef]
- Cohn, M.J.; Patel, K.; Krumlauf, R.; Wilkinson, D.G.; Clarke, J.D.W.; Tickle, C. Hox9 genes and vertebrate limb specification. Nature 1997, 387, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Tickle, C. Tbx18 and boundary formation in chick somite and wing development. Dev. Biol. 2004, 268, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Teran, M.; Ros, M.A. The Apical Ectodermal Ridge: Morphological aspects and signaling pathways. Int. J. Dev. Biol. 2008, 52, 857–871. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.R. The roles of FGFs in the early development of vertebrate limbs. Genes Dev. 1998, 12, 1571–1586. [Google Scholar] [CrossRef] [Green Version]
- Lewandoski, M.; Sun, X.; Martin, G.R. Fgf8 signalling from the AER is essential for normal limb development. Nat. Genet. 2000, 26, 460–463. [Google Scholar] [CrossRef]
- Sun, X.; Mariani, F.V.; Martin, G.R. Functions of FGF signalling from the apical ectodermal ridge in limb development. Nature 2002, 418, 501–508. [Google Scholar] [CrossRef]
- Beermann, F.; Kaloulis, K.; Hofmann, D.; Murisier, F.; Bucher, P.; Trumpp, A. Identification of evolutionary conserved regulatory elements in the mouse Fgf8 locus. Genesis 2006, 44, 1–6. [Google Scholar] [CrossRef]
- Marinić, M.; Aktas, T.; Ruf, S.; Spitz, F. An integrated holo-enhancer unit defines tissue and gene specificity of the fgf8 regulatory landscape. Dev. Cell 2013, 24, 530–542. [Google Scholar] [CrossRef] [Green Version]
- Gehrke, A.R.; Shubin, N.H. Cis-regulatory programs in the development and evolution of vertebrate paired appendages. Semin. Cell Dev. Biol. 2016, 57, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddle, R.D.; Johnson, R.L.; Laufer, E.; Tabin, C. Sonic hedgehog mediates the polarizing activity of the ZPA. Cell 1993, 75, 1401–1416. [Google Scholar] [CrossRef]
- Krauss, S.; Concordet, J.P.; Ingham, P.W. A functionally conserved homolog of the Drosophila segment polarity gene hh is expressed in tissues with polarizing activity in zebrafish embryos. Cell 1993, 75, 1431–1444. [Google Scholar] [CrossRef]
- Tickle, C.; Towers, M. Sonic hedgehog signaling in limb development. Front. Cell Dev. Biol. 2017, 5, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagai, T.; Hosoya, M.; Mizushina, Y.; Tamura, M.; Shiroishi, T. Elimination of a long-range cis-regulatory module causes complete loss of limb-specific Shh expression and truncation of the mouse limb. Development 2005, 132, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Zeller, R.; López-Ríos, J.; Zuniga, A. Vertebrate limb bud development: Moving towards integrative analysis of organogenesis. Nat. Rev. Genet. 2009, 10, 845–858. [Google Scholar] [CrossRef]
- Niswander, L.; Jeffrey, S.; Martin, G.R.; Tickle, C. A positive feedback loop coordinates growth and patterning in the vertebrate limb. Nature 1994, 371, 609–612. [Google Scholar] [CrossRef]
- Tanaka, M.; Tamura, K.; Noji, S.; Nohno, T.; Ide, H. Induction of additional limb at the dorsal-ventral boundary of a chick embryo. Dev. Biol. 1997, 182, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tickle, C. The contribution of chicken embryology to the understanding of vertebrate limb development. Mech. Dev. 2004, 121, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Zakany, J.; Duboule, D. The role of Hox genes during vertebrate limb development. Curr. Opin. Genet. Dev. 2007, 17, 359–366. [Google Scholar] [CrossRef]
- Davis, A.P.; Witte, D.P.; Hsieh-Li, H.M.; Potter, S.S.; Capecchi, M.R. Absence of radius and ulna in mice lacking hoxa-11 and hoxd-11. Nature 1995, 375, 791–795. [Google Scholar] [CrossRef]
- Fromental-Ramain, C.; Warot, X.; Messadecq, N.; LeMeur, M.; Dollé, P.; Chambon, P. Hoxa-13 and Hoxd-13 play a crucial role in the patterning of the limb autopod. Development 1996, 122, 2997–3011. [Google Scholar] [CrossRef]
- Mercader, N.; Leonardo, E.; Azplazu, N.; Serrano, A.; Morata, G.; Martínez, C.A.; Torres, M. Conserved regulation of proximodistal limb axis development by Meis1/Hth. Nature 1999, 402, 425–429. [Google Scholar] [CrossRef]
- Mercader, N.; Leonardo, E.; Piedra, M.E.; Martinez-A, C.; Ros, M.A.; Torres, M. Opposing RA and FGF signals control proximodistal vertebrate limb development through regulation of Meis genes. Development 2000, 127, 3961–3970. [Google Scholar] [CrossRef]
- Delgado, I.; López-Delgado, A.C.; Alberto, R.D.; Giovinazzo, G.; Cadenas, V.; Fernández-De-Manuel, L.; Sánchez-Cabo, F.; Anderson, M.J.; Lewandoski, M.; Torres, M. Proximo-distal positional information encoded by an Fgf-regulated gradient of homeodomain transcription factors in the vertebrate limb. Sci. Adv. 2020, 6, 1–10. [Google Scholar] [CrossRef]
- Tarchini, B.; Duboule, D. Control of Hoxd genes’ collinearity during early limb development. Dev. Cell 2006, 10, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Lonfat, N.; Duboule, D. Structure, function and evolution of topologically associating domains (TADs) at HOX loci. FEBS Lett. 2015, 589, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Andrey, G.; Montavon, T.; Mascrez, B.; Gonzalez, F.; Noordermeer, D.; Leleu, M.; Trono, D.; Spitz, F.; Duboule, D. A switch between topological domains underlies HoxD genes collinearity in mouse limbs. Science 2013, 340, 1234167. [Google Scholar] [CrossRef]
- Montavon, T.; Soshnikova, N.; Mascrez, B.; Joye, E.; Thevenet, L.; Splinter, E.; De Laat, W.; Spitz, F.; Duboule, D. A regulatory archipelago controls hox genes transcription in digits. Cell 2011, 147, 1132–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitz, F.; Gonzalez, F.; Duboule, D. A global control region defines a chromosomal regulatory landscape containing the HoxD cluster. Cell 2003, 113, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Lehoczky, J.A.; Innis, J.W. BAC transgenic analysis reveals enhancers sufficient for Hoxa13 and neighborhood gene expression in mouse embryonic distal limbs and genital bud. Evol. Dev. 2008, 10, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Berlivet, S.; Paquette, D.; Dumouchel, A.; Langlais, D.; Dostie, J.; Kmita, M. Clustering of tissue-specific sub-TADs accompanies the regulation of HoxA genes in developing limbs. PLoS Genet. 2013, 9, e1004018. [Google Scholar] [CrossRef] [Green Version]
- Müller, G.B.; Wagner, G.P. Novelty in evolution: Restructuring the concept. Annu. Rev. Ecol. Syst. 1991, 22, 229–256. [Google Scholar] [CrossRef]
- Hall, B.K. Consideration of the neural crest and its skeletal derivatives in the context of novelty/innovations. J. Exp. Zool. Part B Mol. Dev. Evol. 2005, 304, 548–557. [Google Scholar] [CrossRef]
- True, J.R.; Carroll, S.B. Gene co-option in physiological and morphological evolution. Annu. Rev. Cell Dev. Biol. 2002, 18, 53–80. [Google Scholar] [CrossRef]
- Shubin, N.; Tabin, C.; Carroll, S. Deep homology and the origins of evolutionary novelty. Nature 2009, 457, 818–823. [Google Scholar] [CrossRef]
- Wray, G.A.; Abouheif, E. When is homology not homology? Curr. Opin. Genet. Dev. 1998, 8, 675–680. [Google Scholar] [CrossRef]
- Shubin, N.; Tabin, C.; Carroll, S. Fossils, genes and the evolution of animal limbs. Nature 1997, 388, 639–648. [Google Scholar] [CrossRef]
- Shubin, N.H. Origin of evolutionary novelty: Examples from limbs. J. Morphol. 2002, 252, 15–28. [Google Scholar] [CrossRef]
- Lettice, L.A.; Heaney, S.J.H.; Purdie, L.A.; Li, L.; de Beer, P.; Oostra, B.A.; Goode, D.; Elgar, G.; Hill, R.E.; de Graaff, E. A long-range Shh enhancer regulates expression in the developing limb and fin and is associated with preaxial polydactyly. Hum. Mol. Genet. 2003, 12, 1725–1735. [Google Scholar] [CrossRef]
- Dahn, R.D.; Davis, M.C.; Pappano, W.N.; Shubin, N.H. Sonic hedgehog function in chondrichthyan fins and the evolution of appendage patterning. Nature 2007, 445, 311–314. [Google Scholar] [CrossRef]
- Neumann, C.J.; Grandel, H.; Gaffield, W.; Schulte-Merker, S.; Nüsslein-Volhard, C. Transient establishment of anteroposterior polarity in the zebrafish pectoral fin bud in the absence of sonic hedgehog activity. Development 1999, 126, 4817–4826. [Google Scholar] [CrossRef]
- Grandel, H.; Schulte-Merker, S. The development of the paired fins in the zebrafish (Danio rerio). Mech. Dev. 1998, 79, 99–120. [Google Scholar] [CrossRef]
- Mercader, N. Early steps of paired fin development in zebrafish compared with tetrapod limb development. Dev. Growth Differ. 2007, 49, 421–437. [Google Scholar] [CrossRef]
- Reifers, F.; Böhli, H.; Walsh, E.C.; Crossley, P.H.; Stainier, D.Y.R.; Brand, M. Fgf8 is mutated in zebrafish acerebellar (ace) mutants and is required for maintenance of midbrain-hindbrain boundary development and somitogenesis. Development 1998, 125, 2381–2395. [Google Scholar] [CrossRef]
- Grandel, H.; Draper, B.W.; Schulte-Merker, S. Dackel acts in the ectoderm of the zebrafish pectoral fin bud to maintain AER signaling. Development 2000, 127, 4169–4178. [Google Scholar] [CrossRef]
- Jovelin, R.; He, X.; Amores, A.; Yan, Y.L.; Shi, R.; Qin, B.; Roe, B.; Cresko, W.A.; Postlethwait, J.H. Duplication and divergence of fgf8 functions in teleost development and evolution. J. Exp. Zool. Part B Mol. Dev. Evol. 2007, 308, 730–743. [Google Scholar] [CrossRef]
- Hodgkinson, V.S.; Ericsson, R.; Johanson, Z.; Joss, J.M.P. The apical ectodermal ridge in the pectoral fin of the Australian lungfish (Neoceratodus forsteri): Keeping the fin to limb transition in the fold. Acta Zool. 2009, 90, 253–263. [Google Scholar] [CrossRef]
- Wood, A. Early pectoral fin development and morphogenesis of the apical ectodermal ridge in the killifish, Aphysosemion scheeli. Anat. Rec. 1982, 204, 349–356. [Google Scholar] [CrossRef]
- Géraudie, J. No Initiation of the actinotrichial development in the early fin bud of the fish, Salmo. J. Morphol. 1977, 151, 353–361. [Google Scholar] [CrossRef]
- Nikaido, M.; Noguchi, H.; Nishihara, H.; Toyoda, A.; Suzuki, Y.; Kajitani, R.; Suzuki, H.; Okuno, M.; Aibara, M.; Ngatunga, B.P.; et al. Coelacanth genomes reveal signatures for evolutionary transition from water to land. Genome Res. 2013, 23, 1740–1748. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Wang, J.; Zhu, C.; Yang, L.; Ren, Y.; Ruan, J.; Fan, G.; Hu, J.; Xu, W.; Bi, X.; et al. African lungfish genome sheds light on the vertebrate water-to-land transition. Cell 2021, 184, 1362–1376.e18. [Google Scholar] [CrossRef]
- Thorogood, P. The development of the teleost fin and implications for our understanding of tetrapod limb evolution. Dev. Patterning Vert. Limb 1991, 205, 347–354. [Google Scholar] [CrossRef]
- Yano, T.; Abe, G.; Yokoyama, H.; Kawakami, K.; Tamura, K.; Yano, T.; Abe, G.; Yokoyama, H.; Kawakami, K.; Tamura, K. Mechanism of pectoral fin outgrowth in zebrafish development. Development 2012, 139, 4291. [Google Scholar] [CrossRef] [Green Version]
- Sordino, P.; van der Hoeven, F.; Duboule, D. Hox gene expression in teleost fins and the origin of vertebrate digits. Nature 1995, 375, 678–681. [Google Scholar] [CrossRef]
- Sordino, P.; Duboule, D. A molecular approach to the evolution of vertebrate paired appendages. Trends Ecol. Evol. 1996, 11, 114–119. [Google Scholar] [CrossRef]
- Davis, M.C.; Dahn, R.D.; Shubin, N.H. An autopodial-like pattern of Hox expression in the fins of a basal actinopterygian fish. Nature 2007, 447, 473–477. [Google Scholar] [CrossRef]
- Woltering, J.M.; Irisarri, I.; Ericsson, R.; Joss, J.M.P.; Sordino, P.; Meyer, A. Sarcopterygian fin ontogeny elucidates the origin of hands with digits. Sci. Adv. 2020, 6, 1–8. [Google Scholar] [CrossRef]
- Woltering, J.M.; Holzem, M.; Meyer, A. Lissamphibian limbs and the origins of tetrapod hox domains. Dev. Biol. 2019, 456, 138–144. [Google Scholar] [CrossRef]
- Langellotto, F.; Fiorentino, M.; Felice, E.; Caputi, L.; Nittoli, V.; Joss, J.M.P.; Sordino, P. Expression of meis and hoxa11 in dipnoan and teleost fins provides new insights into the evolution of vertebrate appendages. Evodevo 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.P.; Khan, P.A.; Blanco, M.J.; Misof, B.; Liversage, R.A. Evolution of hoxa-11 expression in amphibians: Is the urodele autopodium an innovation? Am. Zool. 1999, 39, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Bickelmann, C.; Frota-Lima, G.N.; Triepel, S.K.; Kawaguchi, A.; Schneider, I.; Fröbisch, N.B. Noncanonical Hox, Etv4, and Gli3 gene activities give insight into unique limb patterning in salamanders. J. Exp. Zool. Part B Mol. Dev. Evol. 2018, 330, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Kherdjemil, Y.; Lalonde, R.L.; Sheth, R.; Dumouchel, A.; De Martino, G.; Pineault, K.M.; Wellik, D.M.; Stadler, H.S.; Akimenko, M.A.; Kmita, M. Evolution of Hoxa11 regulation in vertebrates is linked to the pentadactyl state. Nature 2016, 539, 89–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, I.; Aneas, I.; Gehrke, A.R.; Dahn, R.D.; Nobrega, M.A.; Shubin, N.H. Appendage expression driven by the Hoxd Global Control Region is an ancient gnathostome feature. Proc. Natl. Acad. Sci. USA 2011, 108, 12782–12786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, R.; Zhang, G.; Cohn, M.J. Biphasic Hoxd gene expression in shark paired fins reveals an ancient origin of the distal limb domain. PLoS ONE 2007, 2, e754. [Google Scholar] [CrossRef] [Green Version]
- Scotti, M.; Kherdjemil, Y.; Roux, M.; Kmita, M. A Hoxa13:Cre mouse strain for conditional gene manipulation in developing limb, hindgut, and urogenital system. Genesis 2015, 53, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Onimaru, K.; Kuraku, S.; Takagi, W.; Hyodo, S.; Sharpe, J.; Tanaka, M. A shift in anterior–posterior positional information underlies the fin-to-limb evolution. Elife 2015, 4, 1–15. [Google Scholar] [CrossRef]
- Letelier, J.; De La Calle-Mustienes, E.; Pieretti, J.; Naranjo, S.; Maeso, I.; Nakamura, T.; Pascual-Anaya, J.; Shubin, N.H.; Schneider, I.; Martinez-Morales, J.R.; et al. A conserved Shh cis-regulatory module highlights a common developmental origin of unpaired and paired fins. Nat. Genet. 2018, 50, 504–509. [Google Scholar] [CrossRef]
- Kondo, T.; Zákány, J.; Innis, J.W.; Duboule, D. Of fingers, toes and penises. Nature 1997, 390, 29. [Google Scholar] [CrossRef] [Green Version]
- Cohn, M.J. Development of the external genitalia: Conserved and divergent mechanisms of appendage patterning. Dev. Dyn. 2011, 240, 1108–1115. [Google Scholar] [CrossRef] [Green Version]
- Gillis, J.A.; Dahn, R.D.; Shubin, N.H. Shared developmental mechanisms pattern the vertebrate gill arch and paired fin skeletons. Proc. Natl. Acad. Sci. USA 2009, 106, 5720–5724. [Google Scholar] [CrossRef] [Green Version]
- Gillis, J.A.; Hall, B.K. A shared role for sonic hedgehog signalling in patterning chondrichthyan gill arch appendages and tetrapod limbs. Development 2016, 143, 1313–1317. [Google Scholar] [CrossRef] [Green Version]
- Pueyo, J.I.; Couso, J.P. Parallels between the proximal–distal development of vertebrate and arthropod appendages: Homology without an ancestor? Curr. Opin. Genet. Dev. 2005, 15, 439–446. [Google Scholar] [CrossRef]
- Tarazona, O.A.; Lopez, D.H.; Slota, L.A.; Cohn, M.J. Evolution of limb development in cephalopod mollusks. Elife 2019, 8, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Stewart, T.A.; Bhat, R.; Newman, S.A. The evolutionary origin of digit patterning. Evodevo 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Turing, A.M. The chemical basis of morphogenesis. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1952, 237, 37–72. [Google Scholar] [CrossRef]
- Kondo, S.; Miura, T. Reaction-diffusion model as a framework for understanding biological pattern formation. Science 2010, 329, 1616–1620. [Google Scholar] [CrossRef] [Green Version]
- Newman, S.A. Lineage and pattern in the developing vertebrate limb. Trends Genet. 1988, 4, 329–332. [Google Scholar] [CrossRef]
- Raspopovic, J.; Marcon, L.; Russo, L.; Sharpe, J. Digit patterning is controlled by a Bmp-Sox9-Wnt Turing network modulated by morphogen gradients. Science 2014, 345, 566–569. [Google Scholar] [CrossRef]
- Onimaru, K.; Marcon, L.; Musy, M.; Tanaka, M.; Sharpe, J. The fin-to-limb transition as the re-organization of a Turing pattern. Nat. Commun. 2016, 7, 11582. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.; Lerea, K.M.; Peng, H.; Kaltner, H.; Gabius, H.-J.; Newman, S.A. A regulatory network of two galectins mediates the earliest steps of avian limb skeletal morphogenesis. BMC Dev. Biol. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Sheth, R.; Marcon, L.; Bastida, M.F.; Junco, M.; Quintana, L.; Dahn, R.; Kmita, M.; Sharpe, J.; Ros, M.A. Hox genes regulate digit patterning by controlling the wavelength of a turing-type mechanism. Science 2012, 338, 1476–1480. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.P.; Williamson, S.; Fallon, J.F.; Meinhardt, H.; Prum, R.O. Molecular evidence for an activator-inhibitor mechanism in development of embryonic feather branching. Proc. Natl. Acad. Sci. USA 2005, 102, 11734–11739. [Google Scholar] [CrossRef] [Green Version]
- Sick, S.; Reinker, S.; Timmer, J.; Schlake, T. WNT and DKK determine hair follicle spacing through a reaction-diffusion mechanism. Science 2006, 314, 1447–1450. [Google Scholar] [CrossRef] [Green Version]
- Cooper, R.L.; Thiery, A.P.; Fletcher, A.G.; Delbarre, D.J.; Rasch, L.J.; Fraser, G.J. An ancient Turing-like patterning mechanism regulates skin denticle development in sharks. Sci. Adv. 2018, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Goodrich, E.S. On the dermal fin-rays of fishes-living and extinct. Q. J. Microsc. Soc. 1904, 47, 465–522. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cass, A.N.; Elias, A.; Fudala, M.L.; Knick, B.D.; Davis, M.C. Conserved Mechanisms, Novel Anatomies: The Developmental Basis of Fin Evolution and the Origin of Limbs. Diversity 2021, 13, 384. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080384
Cass AN, Elias A, Fudala ML, Knick BD, Davis MC. Conserved Mechanisms, Novel Anatomies: The Developmental Basis of Fin Evolution and the Origin of Limbs. Diversity. 2021; 13(8):384. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080384
Chicago/Turabian StyleCass, Amanda N., Ashley Elias, Madeline L. Fudala, Benjamin D. Knick, and Marcus C. Davis. 2021. "Conserved Mechanisms, Novel Anatomies: The Developmental Basis of Fin Evolution and the Origin of Limbs" Diversity 13, no. 8: 384. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080384