
The Towakkalak System, A Hotspot of Subterranean Biodiversity in Sulawesi, Indonesia




1
Institut de Systématique, Évolution, Biodiversité (ISYEB), UMR7205 CNRS, Muséum National d’Histoire Naturelle, Sorbonne Université, EPHE, 45 Rue Buffon, 75005 Paris, France
2
Museum Zoologicum Bogoriense, Research Center for Biology, Indonesian Institute of Sciences, Cibinong Science Center, Cibinong 16911, Indonesia
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(8), 392; https://0-doi-org.brum.beds.ac.uk/10.3390/d13080392
Submission received: 27 July 2021
/
Revised: 9 August 2021
/
Accepted: 10 August 2021
/
Published: 20 August 2021
(This article belongs to the Special Issue Hotspots of Subterranean Biodiversity)
Abstract
:The Towakkalak System located in the Maros karst of South Sulawesi is currently the richest of Southeast Asia in obligate subterranean species. It comprises several caves and shafts that give access to the subterranean Towakkalak river as well as many unconnected fossil caves, stream sinks, and springs located within its footprint. The total length of the caves linked to the active system is 24,319 m and comprises two of the longest caves of Indonesia, Gua Salukkan Kallang and Gua Tanette. Studies of its fauna began in 1985. There are 10 stygobionts and 26 troglobionts that are known from the system. The smaller adjacent system of Saripa has 6 stygobionts and 18 troglobionts, of which 1 and 3, respectively, are absent from Towakkalak. Like all tropical cave inventories, our dataset has limits due to identification uncertainties, gaps in habitat (waters, guano) and taxonomic coverage (micro-crustaceans, mites), sampling methods (pitfall trapping, Karaman–Chappuis), and problems of ecological assignment. A number of additional species are therefore expected to be found in the future. The Towakkalak and Saripa cave systems are included in the Bantimurung-Bulusaraung National Park and are under efficient protection, but parts of the Maros karst outside the park are under serious threat, mainly from quarrying.

1. Introduction and Context
Cave-restricted species were long considered to be exceptional or rare in tropical caves. They are actually widespread in all tropical regions of the world [1,2]. In Indonesia, the presence of such cave-restricted species has been documented since 1985 in the karst of Maros in South Sulawesi, which has been pointed to as one of the richest tropical caves of the world in this respect [2,3,4]. In the present paper, we give a commented and updated list of the cave fauna of the Towakkalak and Saripa cave systems, which have the highest species richness within the Maros karst [5]. We confirm Towakkalak as the richest hotspot of the tropical world for subterranean biodiversity. We briefly circumvent the shortfalls that affect our cave species inventory as well as the few ones available in other regions of tropical Asia. We characterize the Towakkalak system fauna within the Maros karst and in a broader context. We skim through the threats that exist to the Towakkalak and Maros cave biodiversity, focusing on growing concerns about the limestone quarrying that affects large parts of the karst that are not currently included in the Bantimurung-Bulusaraung National Park.
1.1. Geographical Setting
The Maros karst is located in the province of Sulawesi Selatan (South Sulawesi), close to the city of Makassar, located between the latitudes 4.650 °S and 5.088 °S, and is within an altitudinal range from sea level to about 700 m. It is an increasingly popular region in Indonesia, as it combines a series of exceptional features in several domains: fine tower karst landscape, unusual geological formations, huge and beautiful caves, an abundance of prehistoric artefacts, the oldest rock painting on earth, and a rich fauna. Accessible by road in one hour from the big city of Makassar, the Maros karst is under very strong human pressure, mostly due to limestone exploitation, which has destroyed large parts of its unique landscape since the 1980s. The Bantimurung-Bulusaraung National Park (Babul) was created in 2004, and today, it protects the core of this invaluable heritage, though the northern part of the karst is not included.
The Maros region has a tropical climate, but it departs from the surrounding areas of Sulawesi by a contrasted seasonal climate. Over a 30 year period (1931–1960), annual rainfall exceeded 3000 mm per year in Maros. Monthly rainfall was 50 mm per month or less during the dry season (July to September), and was more than 400 mm per month from December to March, with a peak in January (761 mm) [6]. In January 1986, rainfall reached 1067 mm, and the whole plain from Samanggi to Maros was under water. According to [7], during the 1985–2014 period, the rainfall peak in January was 827.40 mm, and the lowest was in August (46.4 mm), with average monthly rainfall of 337.02 mm.
1.2. Archeology
The Maros karst is a major site for the prehistory of Sunda islands. Since the pioneer works of [8], a large amount of buried remains and artefacts have been discovered in several caves on the western border of the karst, especially in the Leang Leang valley and in the nearby karst of Mallawa, that characterize the Toalean culture. Since the 1950s, a number of prehistoric paintings have also been found in several caves of the region, generating a strong interest in knowledge of Southeast Asia prehistory [9,10]. More recently, Maros made headlines again in the archeological world when the dating of several cave paintings revealed ages of up to 45,000 years, leading them to become the oldest rock art in the world [11,12]. Many caves of the Maros karst harbor Pleistocene parietal art, rivaling the ‘ice age’ cave art of western Europe [13].
1.3. Geology
The geology of the Maros karst has been synthetized by [14]. The karst is composed of Tonasa limestone from the upper Eocene to mid-Miocene period. The Tonasa formation, which is 3000 m thick, is mostly composed of massive coralline limestone, bioclastic limestone, and calcarenite. These limestones are discontinuously overlaid by the volcano-sedimentary rocks of the Camba formation, dated from the mid- to upper Miocene period. The Camba formation, with its overlying limestone, subsists in the lowlands south of the Maros karst. It also subsists east of the karst at a higher altitude, suggesting that the two streamlets that sink there, far from the main karst block, probably circulate kilometers under the Camba formation to join the Towakkalak system and its resurgence.
The mid- to late-Miocene volcanic rocks (basalt, trachytes, and diorites) are visible in the karst in several areas as laccoliths, sills, and dykes. Sills are volcanic rocks interbedded horizontally between limestone layers and have a thickness of 10–30 m (Figure 1D). The small K11 spring near Gua Salukkan Kallang emerges at the foot of a sill. But the most unusual feature of the Maros karst is the presence of deep (up to 150 m), narrow (4 to 50 m), long (up to 4 km), straight, or weakly curved corridors in several areas (Figure 1A), which occur in the footprint of the Towakkalak system in particular. These corridors are predominantly generated by volcanic dykes that are more rapidly eroded than the surrounding Tonasa limestone. Interestingly, the subterranean passages that have been explored so far rarely follow corridor dykes. Conversely, in several caves, especially in the case of Gua Salukkan Kallang, narrow to large dykes (1 dm to several meters wide) have been intersected by subterranean galleries, giving large volcanic pebbles in the subterranean streams. These cave passages are therefore posterior to the late Miocene period, though no volcanic filling has been found in the explored caves, which could allow more precise dating [14].
1.4. Geomorphology
‘Such gorges, chasms, and precipices as here abound, I have nowhere seen in the Archipelago. A sloping surface is scarcely anywhere to be found, huge walls and rugged masses of rock terminating all the mountains and inclosing the valleys. In many parts there are vertical or even overhanging precipices five or six hundred feet high, yet completely clothed with a tapestry of vegetation’. This is how Alfred Russel Wallace described the Maros karst in 1890, where he spent weeks searching for insects [15].
The Maros karst is pinpointed, together with the Sierra de los Organos in Cuba, as a model of a tropical tower karst and is repeatedly documented and interpreted by karstologists [6,16]. It is actually mostly constituted by massive cliff-bound karst units, with tower morphology that is only developed in their upper part. Delimited on its western side by subvertical cliffs arising abruptly up to 200 m above the alluvial plain almost at sea level (Figure 1C), it forms large limestone plateaus dissected by a few deep valleys, up to an elevation of 700 m on the slopes of the Bulusaraung extinct volcano. Together with the nearby karsts of Mallawa to the east and Barru to the north, it covers about 700 km2 and is mostly concentrated around Bulusaraung.
The Maros karst offers an amazing variety of surface landforms. The corridors discussed above are its most remarkable feature, which have only been reported elsewhere from the Bau karst in Sarawak, though at a smaller extent [14,17]. They often give access to the core of the karst massif, while the terrain between them is practically impassable. More classical karstic features are noticeable, including vestigial karstic outcrops sparse in the alluvial plain, isolated outcrops of various morphology, flat subsurface limestone with water circulation up to 4 meters under the level of the plain, hemispherical hills, deep poljes and depressions, rocky gorges, impassable lapiez, and tsingy-like formations. The deep karst involves big karstic springs, river sinks, numerous foothill caves, long, clean and beautiful underground rivers, mega shafts (Figure 1B) comparable in size to those of Papua New Guinea, and a variety of speleothems.
1.5. Caves
A well-documented synthesis on the Maros karst and caves was published by [18]. A first expedition was organized by Denis Wellens in 1984, but its results were not published [19]. The discovery of big caves began in 1985, with the first expedition of the Association Pyrénéenne de Spéléologie to Maros, where more than 8 km in Gua Salukkan Kallang were explored and mapped [20], while an Italian team explored and mapped 3500 m of Leang Assuloang [21]. Several other expeditions followed, sometimes in collaboration with local Makassar cavers. Expedition reports provide localization and description of explored caves as well as hydrogeological and biological original data [22,23,24,25,26,27,28,29,30]. All of these caving expeditions were associated with biological surveys and were done with the participation of Indonesian biologists and researchers from Indonesian Institute of Sciences (LIPI) and the Museum Zoologicum Bogoriense. A few independent expeditions were also conducted by Italian and Indonesian cavers, in particular, the Acintyacunyata Speleological Club from Yogyakarta [31] and Korpala UNHAS from Makassar, while several biological sampling campaigns in caves and springs have also been conducted by LIPI researchers.
A total of 219 caves (68 km surveyed) are documented in the APS reports from 1985 to 2001 for the Maros karst [30], to which about 20 caves mapped by the Acintyacunyata Speleological Club from Yogyakarta [31], several tens of caves surveyed by various Indonesian biologists [32] as well as a large number of archaeological caves should be added.
There are five of these caves that have a depth of 190 m or more. Lubang Leaputte is a huge shaft with a 263 m depth and a 100 × 70 m entrance (Figure 2A); Lubang Kapa Kapasa has a 210 m depth and a vertical pit of 205 m; Lubang Beru has a 207 m depth; Gua Salukkan Kallang has a 205 m depth; and Lubang Tomanangna has a 190 m depth with a pit of 170 m and a section of 30 × 20 m [33,34,35].
There are eight caves that are more than 1 km long. A total of four of them belong to the two systems of our study: the Towakkalak system, which contains Gua Salukkan Kallang (12263 m surveyed), Gua Tanette (9692 surveyed m), and Lubang Kabut (1095 m) [35], and the Saripa system, which contains Leang Saripa (2336 m) [34,36]. There have been four other caves that have been explored and mapped outside of these systems: Leang Assuloang (10048 m surveyed) [37]; Gua Londron (5893 m) [38]; Gua Mimpi (1395 m surveyed) [33]; Gua Kacici (1058 m) [39].
2. Material and Methods
In this paper, we focus on the stygobionts and troglobionts, defined independently of their morphology as species only known from caves [1], and also consider the most characteristic stygophiles and troglophiles. Species included in these last ecological categories were all eutroglophiles. The authors of the species names are given in Tables 1 and 2 or in the text for any cited species that are not listed in these tables. The following local terms for landscape features are used: Gua (Indonesian), Leang (Makassar): cave; Gunung (Indonesian), Bulu (Makassar): mountain; Lubang (Indonesian): shaft; Sungai (Indonesian): river.
In this paper, we deal with the biodiversity of the large underground system of Towakkalak and the small adjacent system of Saripa as point of comparison, which are both located in the southern part of the Maros karst.
2.1. The Towakkalak System (Figures 2B–E and 3)
A detailed description of the system can be found in [40,41], and information on the system’s hydrology and hydrogeology can be found in [42,43]. The Towakkalak system (SKT or ‘Gua Salukkan Kallang–Towakkalak’ system in [4]) is a large underground river that can be accessed by several caves between the Sungai Gallang sinks to the big emergence of Towakkalak at Bantimurung, which is located 7 km away in a straight line. There are two caves that represent the main access to the system: Gua Salukkan Kallang and Gua Tanette.
The Towakkalak system includes the second and fourth longest caves in Indonesia, Gua Salukkan Kallang (12,263 m long, 205 m deep, 4 entrances: K1, K2, K3, K4) and Gua Tanette (9692 m, 25 m deep, a single entrance). Cave passages are typically large galleries of a regular section, often more than 15 m in diameter, where the underground Towakkalak river flows and is navigable in two sections by several kilometers. The river can be accessed through two other smaller caves located between two larger and previously cited caves: Lubang Batu Neraka (749 m long, 85 m deep) and Lubang Kabut (1095 m long, 74 m deep) [35,41]. The upstream sump of the system in Gua Salukkan Kallang was dived in 2001 on a length of more than 100 m [36]. Several other caves, unconnected to the Towakkalak underground river, are likely to belong to the same system.
The Towakkalak underground river emerges at about 40 m of altitude as a large spring which provides water to the Bantimurung waterfall, which is 700 m downstream and is a highly praised tourism spot in South Sulawesi (Figure 2C). The Towakkalak resurgence drains a hydrogeological basin of at least 57 km2, including 23.5 km2 of outcropping limestone in its lower part (highest elevation probably 500 m), and 33.5 km2 as volcanic rocks and sediments in its upper part (highest elevation 1353 m at Bulusaraung). Its discharge was measured in the dry season (July) to be about 0.6 m3/s [42], and it probably reaches several tens times this value in the rainiest months. Towakkalak is also fed by several stream sinks, those of the Salukkan Kallang and Pangni streamlets (260 m and 420 m of altitude) and the sinks of the Sungai Gallang at 420 m of altitude in particular. The Salukkan Kallang stream sinks at the contact point between volcanic-alluvia and limestone. Lubang Pangni is located more than 3 km northeast of the Salukkan Kallang sink in a very small limestone isolated outcrop; its 100-meter depth suggests a long water circulation under the volcanic and alluvia surfaces. The Sungai Gallang, which sinks in a very tiny and isolated outcrop of limestone, is supposed to circulate for 2.7–2.9 km under a volcanic ridge towards the upstream sump of Gua Salukkan Kallang. In fact, half of the supposed catchment area of the Towakkalak spring is a black box, as the extent and continuity of the limestone underneath the volcanic terrains and the recent alluvial terrain are unknown.
The limits of the Towakkalak catchment in its limestone part are also uncertain but for another reason. About 700 m downstream of the Towakkalak spring and 30–40 m below its elevation are two other springs: Jamala spring and Baharuddin spring. Dye injected into Gua Salukkan Kallang subterranean stream 4.8 km to the east strongly colored the Towakkalak spring, but none of these other springs [42]. The Baharuddin spring, the smallest (135 l/s in dry season), has no evident link with any of the surrounding caves. The Jamala spring is about two times bigger than Towakkalak (1.3 m3/s) [42]. The origin of its water is not known, and not a single cave can be attached to this spring, while at least 50 caves are located in the footprint of the Towakkalak catchment. Moreover, a long fossil gallery of Gua Tanette, the second largest cave of the Towakkalak system, heads to the west and would pass above the most likely predictable passages of Jamala according to the sump map drawn by divers [36]. Therefore, the northern limits of the Towakkalak system are currently impossible to trace.
2.2. The Saripa System
Biologically, the best documented cave of the Maros karst is Leang Saripa and its spring (Figure 3 and Figure 4). This small hydrogeological system, which is of easier access than the Towakkalak system, is adjacent to it in a same large limestone unit. Its cave fauna is slightly less rich, differing from that of Towakkalak by several taxa. The Saripa system includes the spring of Saripa, which flows out at the plain level from a small cave that is about 50 m long, and Leang Saripa, which is located about 30 m above it.
Many other caves open along the alluvial plain of the Patunuang river down to Bantimurung and are probably connected to the Saripa system during the rainy season when the plain is flooded. Leang Saripa is unusual for its complex system of galleries, contrasting with most caves of the Maros karst. In 2000, about 1700 m of passages, arranged in two or three levels, were explored and mapped [31,33,34]. In 2001, the main sump of the system was dived, giving access to 700 additional meters of big galleries, where a large number of bats were present [36]. Saripa has a large array of aquatic and terrestrial habitats that are favorable to stygobiotic and troglobiotic fauna.
2.3. Sampling
Aside taxonomic descriptions, a number of papers deal with the cave fauna of the Maros karst, in particular that of Towakkalak and Saripa systems, which have been more studied than others [4,5,23,44,45,46,47]. The fauna was mostly collected by the authors of this paper by hand picking, netting, and substrate extraction in the Berlese funnels, over the course of several field trips from 1985 to 2008 and in collaboration with cavers of the Association Pyrénéenne de Spéléologie (Toulouse, France) and the Indonesian caving clubs of Yogyakarta and Makassar. Sampled aquatic habitats include freshwater pools, percolating water, endogenous stream, exogenous streams (often eutrophic), and more rarely, phreatic water. Sampled terrestrial habitats were the oligotrophic and ‘SCAT’ habitats (where the main food supply is scattered swiftlet and bat feces) [48], periodically, inundated habitats with fine silt deposits, flood detritus, bat guano accumulation, parietal and twilight habitats, and soil within cave. Milieu Souterrain Superficiel-type habitats have not been detected, while deep soil habitats outside of the cave host a diverse and rich interstitial fauna not considered here, where troglophilic species are sometimes present.
3. Results
The Towakkalak and Saripa systems, hereafter referred to as Towakkalak and Saripa, are the best studied and richest cave systems of the Maros karst. Their stygobiotic, troglobiotic, main stygophilic, and main troglophilic species are listed in Tables 1 and 2. In 2000, Deharveng and Bedos listed 7 stygobionts and 21 troglobionts in Towakkalak (under the name Gua Salukkan Kallang system) [4]. The updated list includes 10 stygobionts and 26 troglobionts due to changes in the ecological status of several species and the integration of some overlooked species. In addition, one stygobiont and three troglobionts that were absent in Towakkalak but were present in the adjacent Saripa system were added.
3.1. Aquatic Fauna
Tricladida: Only four species of flatworms are cited from Southeast Asian caves, three of which are members of the genus Dugesia Girard, 1850 [49]. Dugesia leclerci is the only eyeless stygobiotic species from Indonesia. It is described and only known from Gua Tanette [50]. A blind flatworm, tentatively reported here as D. leclerci, is present at a very high density in the lake (sump 1) of Leang Saripa. The ecological status of D. uenorum, another cave flatworm described and only known from Lubang Pangni, a shaft probably linked to Towakkalak, is pendent, as it is oculated and weakly pigmented [50].
Aquatic Crustacea, Amphipoda, Bogidiellidae: Amphipoda are represented by a single specimen of an unidentified Bogidiellidae collected in Gua Tanette. The rarity of the Maros species is likely due to the under-sampling of interstitial habitats, as the family is represented in several caves of the Sunda Islands by described (Bogidiella deharvengi Stock and Botosaneanu, 1989 from Halmahera) and undescribed species (on Muna Island, Kalimantan, Sumatra).
Aquatic Crustacea, Isopoda, Cirolanidae (Figure 5A): There is one species of stygobiotic isopod, Cirolana marosina, that is present in the Maros karst. Though derived from a marine stock, it is only known from standing freshwater pools, contrary to several of its subterranean congeners, which live in anchialine habitats [61]. The species is known from four caves of the Maros karst: Gua Assuloang, located more than 15 km from seacoast, Gua Lumpur of the Towakkalak system and the two Saripa caves, more than 25 km from seacoast. A flat alluvial plain almost at sea level lies between these caves and the sea, with mangrove locally present along meandering channels. Interestingly, [61] noticed that the holotype of Assuloang is completely blind, while the Saripa specimens, located more than 10 km SE of Gua Assuloang, have reduced unpigmented eyes, suggesting that a process of incipient speciation might have occurred.
Aquatic Crustacea, Decapoda: The dominant species of stygobiotic and stygophilic mesofauna in the groundwaters of the Towakkalak and Saripa systems are shrimps (Caridea) and crabs (Brachyura) [62].
- -
- Caridea (Figure 5B): The shrimps found in the systems of interest belong to four genera: Caridina H. Milne-Edwards, 1837 (with 1 stygobiotic species restricted to Towakkalak system and 1 stygophilic species); Marosina, Cai and Ng, 2005 (endemic genus with two stygobiotic species only known from the Maros karst, one of which is limited to Gua Salukkan Kallang), Parisia Holthuis, 1956 (endemic genus with one rare troglobiotic species only found in Gua Tanette), and Macrobrachium lar, a large size stygophile widespread in Pacific and Indian Ocean islands [62,63,64]. These shrimps often live as large populations in springs, streams, and puddles. Their diversity in the Maros karst is reminiscent of the radiation of the genus Caridina in the lakes of Sulawesi [65], which is not matched elsewhere in tropical caves in Asia and obviously calls for further sampling. The proneness to colonize cave habitats repeats in several clades of the worldwide distributed family Atyidae [66] but is particularly marked in the Maros karst, which has the highest number of troglobiotic species in Australasia and the highest level of troglomorphy with its two Marosina species.
- -
- Brachyura (Figure 5C,D): Crabs are frequent in the Maros caves, with four species. A total of three of them belong to the speciose sundaic genus Parathelphusa H. Milne-Edwards, 1853, of which P. sorella, a cave-obligate species with reduced eyes and that is restricted to Saripa cave, where it is rather common [67]. The most remarkable crab species of Maros is the small Cancrocaeca xenomorpha, which is blind and with very long and thin legs [68]. The monospecific genus Cancrocaeca Ng, 1991, belongs to a family of mostly marine species, but the Maros species only lives in freshwater, both in the Towakkalak and Saripa systems, where it occurs sporadically in standing water puddles. A second species of Hymenosomatidae discovered more recently in a cave of the Sangkulirang karst of Kalimantan, Guaplax denticulata Naruse, Ng and Guinot, 2008, also lives in freshwater. Both differ from the third cave Hymenosomatidae of the region, Sulaplax ensifer Naruse, Ng and Guinot, 2008, from Muna Island in Southeast Sulawesi, which lives in brackish water.
Aquatic Vertebrata, Pisces (Figure 5E): Only two modified cave fish are known from Indonesia, and both are from Sulawesi. Bostrychus microphthalmus from caves of the Maros karst is one of them, which is so far restricted to the Towakkalak and Saripa systems [69,70]. It retains minute eyes covered with skin and is the only Bostrychus modified for cave life according to Proudlove [71]. The species is found sporadically in still water pools. In Leang Saripa, the species was collected in 2004 in the sump where several specimens were also observed in August 2020. We also collected a discolored but normal-eyed Glossogobius in Gua Tanette, which could be a cave form of an undescribed species found in the surface water of the Maros area [69].
3.2. Terrestrial Fauna
Gastropoda: Terrestrial snails of the family Subulinidae are frequent in Maros caves, especially in guano, as well as in most caves of Southeast Asia. They are represented by two large-range species in the systems of interest [72].
Araneae (Figure 6A,B): Spiders are present and are diversified in all of the terrestrial habitats of the Maros caves. A total of nine species, which may be either troglobiotic or troglophilic are listed in Table 1 and Table 2.
- -
- Speocera caeca: This troglobiont is widely represented in the caves of the two systems. It is the only blind species of the speciose tropical genus Speocera Berland, 1914, which includes several other cave species [73].
- -
- The species cf. Amauropelma sp.: It is a troglobiotic spider that has been found in several caves of the two systems. By its reduced and unpigmented eyes, pale body color, and rather large size, it is reminiscent of Amauropelma matakecil Miller and Rahmadi, 2012, that was recently described from caves of Central Java [74].
- -
- Psiloderces leclerci: The speciose Southeast Asia genus Psiloderces Simon, 1892, is known by one species in our study, P. leclerci, from one cave and one surface site. The latter form is ‘much deeper in color’ and has shorter legs than that of the cave [75], suggesting a possible separate species status. Psiloderces leclerci belongs to the speciose genus Psiloderces of the South Asian family Psilodercidae, widespread on Sunda island with a few other cave species.
- -
- Spermophora maros: The genus Spermophora Hentz, 1841, comprises 45 species that are widely distributed in tropical regions around the world [53] but that are rare in caves. Spermophora maros is the only species of the Sunda Islands to have reduced unpigmented eyes, long legs, and whitish coloration [76].
- -
- Uthina mimpi: The 17 species of the genus Uthina Simon, 1893 [53], mostly live in forest litter. Uthina mimpi is, however, a weakly modified species of pale coloration, slightly reduced eyes, and slender legs that seems to be fully troglobiont. In addition, two other Uthina species of less clear ecological status, the widespread troglophilic Uthina luzonica Simon, 1893, and the Maros endemic U. sulawesiensis Yao and Li, 2016, probably exist in our study area [77].
Aside from several unidentified families of troglophilic species and the Uthina spp. discussed above, two other identified species are listed in Table 2. Speocera karkari is reported from a few surface sites in Southeast Asia. In Sulawesi, it is only known from two caves, Gua Mampu in the Bone karst and Gua B2 of the Towakkalak system. The big Heteropoda beroni is a regular inhabitant of the caves of the two systems, quietly resting on cave walls, especially in non-oligotrophic habitats, and often near the cave entrance. A species of this genus that may be H. beroni has been found hunting during the night near caves at Bantimurung. It is a major predator of the giant arthropod communities, just like its congeners in other Southeast Asia karsts [4].
Opiliones: There is one unidentified species of Opiliones, possibly an Assamiidae, that is found rather frequently as isolated specimens in the caves of the Towakkalak and Saripa systems. It has small eyes, is pale yellow, and has never been found outside of caves. Similar species are sometimes present in Southeast Asian caves, but most remain unstudied.
Palpigradi: Karsts of South Sulawesi around Maros are unusually rich in Palpigradi, with two families, three genera, and four species, three of which have been described and are only known from the region. There are two species (Koeneniodes frondiger Remy, 1950 and Eukoenenia paulinae Condé, 1994) that are soil dwellers. The two other species are so far cave restricted. Eukoenenia maros is limited to two caves of the Towakkalak system, and Prokoenenia celebica is known from Gua Salukkan Kallang and Gua Mampu in the Bone karst. Always rare in caves, these minute Arachnida were found as isolated specimens in oligotrophic habitats. As the soil outside caves has been well sampled in Maros, the troglobiotic status of the cave species is reliable. In addition, E. maros is considered as troglomorphic in its original description.
Pseudoscorpiones: They are rare, represented by one or two blind unidentified species, which are present in two caves.
Schizomida (Figure 6C): Schizomida of Southeast Asia are severely under-studied compared to those of tropical America or Australia. In Maros, they are frequent in caves and soils, with species differing in particular by the morphology of the male flagellum and the relative length of the appendages. Some morphospecies seem to be cave restricted. Given the proneness to speciation in several lineages of schizomids [78], several new species are expected to occur in Maros as they do in other low elevation karsts of tropical Asia.
Amblypygi (Figure 6D): Whip spiders are, after huntsman spiders of the genus Heteropoda, the more common predators of the giant arthropod communities in most caves of Southeast Asia. There are two species of the genera Sarax Simon, 1892, and Charon Karsch, 1879, that are reported from Maros. The Sarax is probably new to science [79]. Both are common in many caves of the region, including the Towakkalak and Saripa system, where they are sometimes encountered rather deep inside caves. They may feed on the Rhaphidophora sp. abundant in all caves, probably on juveniles or eggs rather than on adults given the size of this cricket. None of these species has been collected outside caves in Maros so far. The Sarax is possibly troglobiont, as suggested by its pale-yellow color and relatively long legs [79]. Several Charon described from Southeast Asia were synonymized with C. grayi (Gervais, 1842) by Kraepelin [80], but recent publications do not accept these synonymies and consider that several species are likely to occur in the region [81,82].
Diplopoda (Figure 6E,F): Hypocambala helleri, a widely distributed saprophagous species, is the most abundant milliped and perhaps arthropod in eutrophic or mesotrophic habitats of the Maros caves. Large populations may be observed on guano, but it is also common in SCAT habitats. The species is considered as a troglophile in the literature but was only found in caves in the Maros karst. A second, less frequent species is the troglobiotic snow white Eutrichodesmus reductus, present in several caves of the Maros karst including both systems of interest [83]. It is never found in dense populations nor on guano piles. Metopidiothrix kalang has a morphology of a surface species (eyes and pigment present) but has been regularly and only found in Gua Salukkan Kallang so far.
Oniscida (Figure 7A,B): There are two cave restricted terrestrial isopods that are reported from Towakkalak [84] and that are present in Saripa system. The most frequent, undescribed species assigned to the genus Venezillo Verhoeff, 1928, which is whitish with reduced eyes, has been found in many caves of the Maros karst, where it lives in oligotrophic to mesotrophic habitats, feeding on organic debris. It has never been found outside of caves. The second species, assigned to the genus Papuaphiloscia Vandel, 1970, is an eyeless and transparent species that is also found in several caves of the Towakkalak and Saripa systems. Both genera have cave species in other regions of Southeast Asia.
Collembola [85] (Figure 7C–E): Springtails are at the basis of trophic chains for the terrestrial invertebrate compartment in most caves of the world, especially in oligotrophic habitats. In guano, they are usually second to mites but reach huge density locally. They are preyed upon by beetles, mites, spiders, pseudoscorpions, and probably schizomids. Guano is not as abundant in the Maros karst as it is in other tropical regions of Asia or the Pacific, but three caves of the Towakkalak and Saripa system nevertheless host large bat colonies. The dominant springtails in these habitats are the parthenogenetic pantropical species listed in Table 2. In the oligotrophic or mesotrophic habitats of Maros caves, Collembola are moderately diversified. In the Towakkalak and Saripa systems, a single troglobitic species of the family Entomobryidae, Pseudosinella maros, is known which is also present in other caves of the Maros karst. This species is apparently related to blind small surface Pseudosinella Schäffer, 1897 that are abundant in the forest soils of the region. P. maros mostly differs from them by several troglomorphic characters, i.e., larger body size, proportionally longer appendages, and thinner claws [86]. Similar to its surface relatives in Maros, P. maros is eyeless. Though weakly modified, it is more troglomorphic than other tropical cave Pseudosinella that have been described so far [87]. Similar undescribed forms of this genus are present in caves of Halmahera [88] and Papua (Fak-Fak) [89]. Pseudosinella maros exhibits noticeable variability in claw elongation and antennal S-chaetotaxy. Specimens collected in underground systems of the Maros karst north and south of the type locality (Gua Restauran, close to but independent from Saripa system) might be taxonomically different [86]. This has been corroborated by subsequent barcode analyses, which indicate species-level divergences of 15–20% between populations within and around the systems of interest (unpublished data).
Diplura: Lepidocampa (Lepidocampa) hypogaea is a clearly troglomorphic species and is widespread in the caves of Towakkalak and Saripa (Table 1). Interestingly, all 11 specimens collected in Gua Baharuddin, a system adjacent to Towakkalak, differ morphologically from those of Towakkalak [90].
Orthoptera, Rhaphidophoridae (Figure 8A): Diestrammena heinrichi (Ramme, 1943) is a large Orthoptera described from the Maros region, but we ignore from which habitat it is described. Allegrucci et al. [91] mention a Rhaphidophora Serville, 1838, from Leang Saripa, and we assigned the very abundant cricket present in all of the caves of the Maros karst to this last genus, pending a redescription of D. heinrichi. This Rhaphidophora is the biggest and most common species of the giant arthropod community in the caves of the Towakkalak and Saripa systems. It seems to be saprophagous, but it has not been ascertained that it feeds outside of the cave during night as observed for some other species of the family. The species has normal eyes but a rather pale and uniform coloration.
Blattodea, Nocticolidae (Figure 8C): Nocticolidae are regular troglobionts in caves in Southeast Asia and the Pacific and are among the most troglomorphic cave arthropods in this area, often combining anophthalmy, marked appendage elongation, and depigmentation. Specimens collected in the Towakkalak and Saripa systems have not been analyzed by specialists, but two eyeless troglomorphic species are present (Fred Stone in litt.), differing by slenderness, degree of troglomorphy, and male tergal structure. They are commonly found in loose groups deep inside caves and mostly in oligotrophic habitats, where they run very quickly when disturbed. Nocticolidae are considered as saprophagous in the literature, probably feeding on degraded organic matter, which is consistent with what we observed in our caves. Interestingly, blind as well as microphthalmic species have been collected in several other caves of the Maros karst.
Coleoptera, Carabidae (Figure 8B,D): Cave obligate beetles are exceptional in tropical cave systems. There are two species of Carabidae (ground beetles) from two different subfamilies that are known in the Towakkalak and Saripa systems, a case that is unique to the Indonesian archipelago: Mateuellus troglobioticus (Pterostichinae) and Eustra saripaensis (Paussinae). The only species of the genus Mateuellus Deuve, 1990, M. troglobioticus, is also the only clearly troglobiotic species of the large tribe Abacetini. It exhibits moderately troglomorphic characteristics, i.e., slightly reduced eyes and slightly elongate appendages. The species is a regular inhabitant of the caves of the Maros karst, with a subspecies described from the nearby Tompobulu karst, M. troglobioticus faillei Deuve, 2010. Interestingly, another abacetin beetle was recently described from Java caves (Metabacetus willi Guéorguiev 2013). Though only found in caves, it is considered to be a troglophile or a trogloxene by its author on the basis of its dark color and inconspicuously modified eyes and leg length. Eustra saripaensis is a rare micro-endemic species that is blind and unpigmented, exclusively known from Leang Saripa. It seems to be restricted to a single location, a clay slope in the unflooded part of the cave, where it usually occurs in rotten bamboo or wood. The genus is known by another cave species (E. pseudomatanga cavernicola Deuve, 2001) and an edaphic species in Sulawesi. It is also present in a few caves of other Southeast Asian regions [57], where it is always rare.
Lepidoptera: Tineidae are very abundant on guano piles and are represented by several unidentified species.
Swiftlets: Swiftlets are common in the Towakkalak and Saripa systems as well in several caves of the Maros karst. They are an important provider of organic matter in the caves where they nest, though large piles of their guano have not been observed. There have been three species recorded in South Sulawesi—two Aerodramus Oberholser, 1906, which echolocate, and Collocalia esculenta (Linnaeus, 1758), which does not [92], but their exact identification cannot be ascertained. Aerodramus swiftlets were qualified of ‘good speciator’ [92] for having both extremely good flying, hence dispersal ability, and a strong link to their home caves. Of the many species recognized in Southeast Asia and the Pacific, these two are present in the caves of Sulawesi, but it is not known if they are both present in the Maros karst.
Bats: Bats are present in many Maros caves, where 15 species are recorded [93]. Insectivorous species are much more frequent than frugivorous or nectarivorous ones. The former are encountered, sometimes deep inside caves, as isolated individuals and small groups in many caves of the karst. Rather large colonies with large piles of guano are surprisingly rare in our study area: unidentified species in Gua Alolu and Emballonura alecto (Eydoux and Gervais, 1836) in Leang Saripa and in Gua Mimpi. The only very large colony of insectivorous bats, of the species Chaerephon plicatus (Buchanan, 1800), is located 13 km NW from Towakkalak in a cave near Salenrang. Fruit bats are recorded from Gua Kelelawar near Kappang as a rather large colony of two species, Dobsonia exoleta K. Andersen, 1909, and Rousettus amplexicaudatus (E. Geoffroy, 1810). Most records of bats in the Maros karst are actually from other caves outside of our study area, such as Gua Londron, Gua Mattampa Belakang, Gua Togendra, or Gua Peceng [93; Rahmadi pers. comm.].
4. Discussion and Conclusions
4.1. Limitations of The Checklist
The number of species of an underground site makes sense by comparison to others. Comparisons make sense if sampling is comparable in terms of sampled habitats, sampled groups, and sampling efforts. This condition is rarely fulfilled, resulting in false or uncertain species absence (or presence) in lists, which should lead to cautious interpretation and comparison of the results. We provide hereafter an overview of gaps and bias of our dataset, which was summarized in Table 1 and Table 2.
4.1.1. Taxonomy
A large proportion of the listed species are undescribed, which is usual in tropical cave species inventories, even the best documented ones [94]. Several groups have been collected but not studied, such as Opiliones, Pseudoscorpiones, or Schizomida, which may uncover several species.
4.1.2. Habitats
Habitats have been unevenly sampled. Aquatic, especially interstitial habitats, guano, and hanging roots, all known to often be species-rich, were not sampled, under-, or minimally sampled, which may account for the low diversity of microcrustacea, guanophiles, and euedaphomorphic species in our caves. Stygobiotic microinvertebrates represent a major part of subterranean biodiversity. They have been sporadically collected but not identified. Guano has only been marginally sampled. Hanging roots have not been sampled in the systems of interest, but in Gua Assuloang, where they were sampled, they hosted the remarkable genus Celebenna Hoch and Wessel, 2011 (Cixiidae), which is so far endemic of this cave. The global richness of the fauna has probably been severely truncated for the concerned taxa, a major bias that has to be taken into account when comparing species richness.
4.1.3. Sampling Methods
The collection methods were mostly direct hand-picking and Berlese extraction of substrate cores in terrestrial habitats and netting using large mesh nets in aquatic habitats. Pitfall trapping, baiting, the use of fine mesh nets, or Karaman–Chappuis in sediments, to maintain basic collecting methods, were not or marginally used, and would have probably provided additional species.
4.1.4. Species Ecological Status
Assignment to troglobionts of terrestrial species found in caves on the basis of their anophthalmy and depigmentation is widespread in the literature, while these traits are more common in euedaphic species from deep soil than in troglobionts [95]. Here, we only assigned species that were collected in caves to troglobionts or stygobionts [1], a status that has to be considered provisional, at least for non-troglomorphic species, as it depends on our knowledge of surface fauna, especially soil. To limit this uncertainty hanging over most species lists, we sampled as much as possible inside of and outside of caves.
4.2. Cave Fauna Features of The Towakkalak and Saripa Systems
4.2.1. Species Richness
A total of 26 troglobionts and 10 stygobionts, i.e., 36 obligate cave species, are known for Towakkalak; the numbers are 19, 6, and 25 for Saripa. These values are the highest recorded for any Southeast Asian caves, and probably for any tropical cave. Most species are shared by both systems, but 15 species are only present in Towakkalak, and 4 species are only in Saripa. The much larger size of Towakkalak is probably linked to more diverse habitats that could explain the difference, which might increase in the future, as Towakkalak has been much less studied than Saripa.
4.2.2. Endemism
There are thirty-four cave restricted species that are endemic to the area, having not been detected in the nearest karsts of Bone (east of South Sulawesi), in Muna (Southeast Sulawesi), nor elsewhere in Indonesia. The remaining species are morphospecies of unknown distribution but that are likely to be endemic as well. The figure is completely different for stygophiles and troglophiles, with only 4 probably endemic species of a total of 28. Endemism at the generic level is a marker of stronger geographic isolation. There are four genera that are endemic to the Maros karst. A total of three of them are present in the systems of interest with four species: the minute crab Cancrocaeca xenomorpha, two shrimps species of the genus Marosina, and the terrestrial beetle Mateuellus troglobioticus. The three former species are highly troglomorphic, the latter, only moderately. There are two more genera that are endemic of the Maros karst but were not found in the systems of interest: the stygobiotic beetle Speonoterus bedosae Spangler, 1996, from Gua Mangana [42] and Celebenna thomarosa, a non-troglomorphic Cixiidae (Homoptera) from Gua Assuloang. The presence of five endemic genera confers an exceptional biological value to the Maros karst. All of the described cave restricted species of Towakkalak and Saripa are endemic to the Maros karst. Other karsts of South Sulawesi are much less known, but none of their cave restricted species have been found in the Maros karst [96].
4.2.3. Shared Diversity Features
The species composition of the Towakkalak and Saripa caves have two common characteristic features: a high diversity of Decapoda among stygobionts and a high diversity of arachnids among troglobionts. Decapoda dominate the aquatic fauna diversity, with 6 stygobionts (5 in Towakkalak, 3 in Saripa) out of a total of 11 (10 in Towakkalak, 6 in Saripa). This dominance seems to be characteristic of Sulawesi caves, which is in line with the radiation undergone by Caridea in the lakes of Southeast Sulawesi (see Caridea above). With 12 troglobiotic or likely troglobiotic species (11 in Towakkalak, 5 in Saripa) of a total of 29 troglobionts (26 in Towakkalak, 18 in Saripa), Arachnida dominate the cave terrestrial fauna in terms of diversity, as is the case in other tropical caves of the region [4], although this figure seems less clear for tropical Brazilian caves [97,98].
4.2.4. Troglomorphy
Patterns of troglomorphy are contrasted among cave species of the systems of interest. In Southeast Asia, for several taxa, the occurrence and degree of troglomorphy seem to roughly decrease with decreasing elevation and with decreasing distance to the equator [2]. Due to the geographical location near equator and low altitude, Maros cave species were therefore not expected to exhibit significant troglomorphic traits. However, they do. The ultimate driver of cave colonization, seasonality, is strong in the Maros karst region and may have led to these modest cave-related morphological modifications that affect several species of Palpigradi, springtails, Diplura, beetles, and fish. On the other hand, Nocticolidae and aquatic taxa generally do not respond to the elevation and latitudinal gradients nor to seasonality and exhibit highly troglomorphic morphologies in many low altitude caves of tropical Asia, as they do in the Towakkalak and Saripa systems.
4.2.5. Guano
As the major food resource for cave fauna in tropical caves, guano habitats deserve some comments. SCAT habitats in Maros caves host, as in most caves of the world, the largest number of cave restricted species [48,99]. On the other hand, guano accumulations have a much richer overall fauna, especially diversified in tropical caves, which remain poorly documented [4]. Remote fossil passages where bats or swiftlets do not venture, such as the large fossil galleries of Gua Tanette, are almost azoic. Surprisingly for a tropical karst of Southeast Asia, guano accumulations are not common in the Maros karst (see above ‘bats’). Leang Saripa hosted isolated specimens of insectivorous bats and swiftlets in the first part of the cave but hosted large bat colonies beyond sump 1. Guano accumulation in Maros caves is mostly produced by insectivorous bats, exceptionally by frugivorous ones, and never by swiftlets. As is the case in Vanuatu caves [100], significant faunistic differences between guano of different types were not detected, but the dominant group in this habitat, mites, have not been analyzed. The identified fauna was mostly represented by large populations of troglophiles-guanophiles of wide distribution: Hypocambala helleri and several pantropical parthenogenetic Collembola (Table 2). The former species seems to be restricted to caves in the Maros karst, where it is very common, but is reported from surface habitats in other tropical regions. Among Collembola, Xenylla yucatana, a pantropical species, is frequent in cave guano and is scarce in surface habitats. The other cited Collembola, mostly pantropical and parthenogenetic, may form dense populations in guano piles but all are more frequent in outside soils. Species of the giant arthropod community as well as, though less frequently, some troglobionts, such as Mateuellus troglobioticus, Venezillo sp. or Schizomida, are often wandering near guano piles.
4.2.6. Invasives
Big cockroaches are often present at a huge density in low altitude and warm tropical caves of Southeast Asia, especially when they have been disturbed by humans. Pycnoscelus surinamensis L., 1767, is reported, for instance, from Gua Mampu in the Bone karst, which is 35 km NE of the Maros karst [101]. However, these giant cockroaches are absent in all of the caves that were surveyed in the Maros karst, including those of Towakkalak and Saripa. In Leang Saripa, the occurrence of rats was noticed in several parts of the cave, indicated by bottles of pitfall traps that were missing or removed from their original places. Some kinds of rodent nest were also found in small holes on the cave walls, mostly composed of plastic garbage.
4.3. Conservation Issues
The cave fauna of the Maros karst, the richest spot of subterranean biodiversity in the tropics, is well protected in the core of the Bantimurung-Bulusaraung National Park, but several peripherical zones are at risk and experience the impact of human pressure that have dramatically increased during the last two decades. Forest logging and land use changes, which may induce important modifications in water flow and circulation, disturbing food supply for underground fauna, are active in these non-protected parts of the karst. Given the terrain roughness, this is not, however, the main threat on cave fauna. Contrary to Gunungsewu in Java, Maros is lucky enough to have agriculture related pollution sources that are mostly downstream karst resurgences, not upstream sinking rivers. Cave over-frequentation linked to the impressive increase in local tourism in Bantimurung and its associated degradations, particularly garbage, is a more recent concern that seriously affects cave habitats in various ways (Gua Mimpi and Leang Saripa). Its direct effects on invertebrate fauna are likely very local. Walls are sometimes extensively tagged, even in the deepest parts of Leang Saripa, but this landscape degradation does not directly impact the fauna. The main biological concern related to over-frequentation is actually that large bat colonies have already abandoned the most visited cave (Gua Mimpi).
However, the major concern for landscape and cave fauna is the multiplication of limestone exploitations during the last two decades, affecting some of the best karst landscape and also several archaeological caves. The extent to which they impact cave invertebrate communities is unknown because they are mostly located in unsurveyed areas north of the National Park boundaries. The only well biologically surveyed outcrops in this area, Mattampa and Lancina northeast of Pangkadjene, host an original and rich cave fauna, including a blind scorpion (Chaerilus sabinae Lourenço, 1995) and a large Japygidae that are unknown elsewhere in Maros. This raises serious concerns about the potential impact of these numerous limestone quarries on Maros subterranean biodiversity. Even when the habitat is not destroyed itself, noise and dust linked to quarrying are known to disturb swiftlets, bats, and bat colonies, which are the main food providers for obligate cave fauna and the only one for guanobionts, but we also lack information on the distribution of swiftlets and bats in this part of the karst. They are also, together with climate change, the greatest threat on the preservation of the invaluable prehistoric rock art of the Maros karst [13] as well the main cause of the current disfiguration of the unique landscape of the karst outside of the National Park boundaries.
The upgrade of the Karaenta Nature Reserve to National Park status in 2004 placed the core of the karst, including the Towakkalak system, under strong protection, but the problems have worsened for several limestone blocks outside the Park boundaries, which are currently without guardrail as a response to the impact of quarrying on cave biodiversity. It is hoped that the candidature of the Maros-Pangkep karst as a World Heritage site, a status which it deserves in so many respects, will lead to the reconsideration of the multiplication of limestone exploitations in its northern part from a landscape and biodiversity perspective.
Author Contributions
Prepared the framework of the article, L.D.; all contributed to data compilation, data interpretation, writing, and editing, L.D., C.R., Y.R.S., and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
Funding for field work in 2001–2002 was provided by the ARCBC joint cooperation project between the European Union and ASEAN; by the Museum National d’Histoire Naturelle (Paris) in 2007; and during these few last years by LIPI (DIPA Pusat Penelitian Biologi: 2003–2015) for Maros & Gunungsewu Karst Project for C.R. and Y.R.S.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This paper is dedicated to the memory of the late Baharuddin, who introduced us to the beautiful karst of Maros, for his unforgettable sense of humor. Our campaigns in the Maros karst were backed by teams of cavers and divers, both Indonesian and French, without whom we would have been unable to sample a number of caves. Thanks to François Brouquisse, Didier Rigal, Franck Brehier, and all those who participated in the Association Pyrénéenne de Spéléologie expeditions to Maros. Philippe Leclerc actively contributed to biological sampling together with several LIPI researchers (late Renny K. Hadiaty, Agustinus Suyanto, Ristiyanti M. Marwoto and Daisy Wowor). Jean-Yves Rasplus and Didier Rigal kindly provided several of their photographs. We are indebted to Roland Barkey from Maros for his kind support from the beginning of the work on the karst, and to many people of Bantimurung and around for their help in the field. Special thanks to the National Park staff, particularly Iqbal Abadi Rasjid, Chaeril, and Dedy Asriady, for their support and kindness during field works. Last but not least, we are grateful to the numerous taxonomists who have efficiently studied our collections.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Deharveng, L.; Bedos, A. Diversity of Terrestrial Invertebrates in Subterranean Habitats. In Cave Ecology, Ecological Studies 235; Moldovan, O.T., Kovác, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 107–172. [Google Scholar]
- Deharveng, L.; Bedos, A. Chapter 18—Diversity Patterns in the Tropics. In Encyclopedia of Caves; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 146–162. [Google Scholar]
- Culver, D.C.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Caves Karst Studies 2000, 62, 11–17. [Google Scholar]
- Deharveng, L.; Bedos, A. The cave fauna of Southeast Asia. Origin, evolution and ecology. In Ecosystems of the World 30. Subterranean Ecosystems; Wilkens, H., Culver, D.C., Humphreys, W.F., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 603–632. [Google Scholar]
- Suhardjono, Y.R.; Ubaidillah, R. (Eds.) Fauna Karst dan gua Maros, Sulawesi Selatan; LIPI Press: Jakarta, Indonesia, 2012; 258p. [Google Scholar]
- Balazs, D. Karst Regions in Indonesia. Karszt Es Barlang 1968, 5, 3–61. [Google Scholar]
- Arsyad, M.; Sulistiawaty, U.; Tiwow, V.A. Analysis of Characteristics and Classification of Rainfall in the Maros Karst Region, South Sulawesi. In Proceedings of the International Seminar on Mathematic, Science and Computer Education, Bandung, Indonesia, 15 October 2016. [Google Scholar]
- Sarasin, P.; Sarasin, F. Die Toala-Hoehlen von Lamontjong. Versuch Einer Anthropologie der Insel Celebes; C.W. Kriedel’s Verlag: Wiesbaden, Germany, 1905. [Google Scholar]
- Van Heekeren, H.R. Rock-paintings and other prehistoric discoveries near Maros (South West Celebes). Laporan Tahunan 1952, 22–35. [Google Scholar]
- Glover, I.C.; Sinha, P. Changes in stone tool use 10,000 years ago: A microwear analysis of flakes with use gloss from Leang Burung 2 and Ulu Leang 1 caves, Sulawesi, Indonesia. Mod. Quat. Res. S. E. Asia 1984, 8, 137–164. [Google Scholar]
- Aubert, M.; Brumm, A.; Ramli, M.; Sutikna, T.; Saptomo, E.W.; Hakim, B.; Morwood, M.J.; van den Bergh, G.D.; Kinsley, L.; Dosseto, A. Pleistocene cave art from Sulawesi, Indonesia. Nature 2014, 514, 223–227. [Google Scholar] [CrossRef]
- Brumm, A.; Oktaviana, A.A.; Burhan, B.; Hakim, B.; Lebe, R.; Zhao, J.X.; Sulistyarto, P.H.; Ririmasse, M.; Adhityatama, S.; Sumantri, I.; et al. Oldest cave art found in Sulawesi. Sci. Adv. 2021, 7, eabd4648. [Google Scholar] [CrossRef]
- Huntley, J.; Aubert, M.; Oktaviana, A.A.; Lebe, R.; Hakim, B.; Burhan, B.; Muhammad Aksa, L.; Made Geria, I.; Ramli, M.; Siagian, L.; et al. The effects of climate change on the Pleistocene rock art of Sulawesi. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brouquisse, F. 13. Cadre géologique. In Expédition Thaï-Maros 85, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1986; pp. 101–118. [Google Scholar]
- Wallace, A.R. The Malay Archipelago: The Land of the Orang-Utan and the Bird of Paradise: A Narrative of Travel with Studies of Man and Nature; Macmillan and Co.: London, UK, 1890. [Google Scholar]
- Sunartadirdja, M.A.; Lehmann, H. Der tropische Karst von Maros und Nord-Bone in SW-Celebes (Sulawesi). Z. Geomorph. 1960, 4 (Suppl. 2), 49–65. [Google Scholar]
- Crabtree, S.; Friederich, H. The Caves of the Bau District, Sarawak. Cave Sci. 1982, 9, 83–93. [Google Scholar]
- Kusch, H. Speläologische Forschungen auf der Insel Sulawesi (Celebes, Indonesien) zwischen 1857 und 1977. Die Höhle 1981, 32, 91–102. [Google Scholar]
- Deharveng, L.; Bedos, A. 10. Les cavités des environs de Bantimurung. In Expédition Thaï-Maros 85, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1986; pp. 81–95. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expédition Thaï-Maros 85, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1986; pp. 1–215. [Google Scholar]
- De Vivo, A.; Campion, N.; Menin, A.; Viviani, F. Vecchie storie Indonesiane. Speleologia 1992, 27, 32–41. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expédition Thaï-Maros 86, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1987; pp. 1–174. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expéditions de l’A.P.S. en Asie du Sud-Est. Travaux Scientifiques—1; APS: Toulouse, France, 1988; pp. 1–52. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expéditions Maros 88-Maros 89, Rapport Spéléologique; APS: Toulouse, France, 1990; pp. 1–51. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expédition Indonésie 90, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1992; pp. 1–104. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expédition Maros 94, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1997; pp. 1–40. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Indonésie 92, Rapport Spéléologique; APS: Toulouse, France, 2001; pp. 1–67. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Expédition Maros 99, Rapport Spéléologique; APS: Toulouse, France, 2002; pp. 1–39. [Google Scholar]
- Association Pyrénéenne de Spéléologie. Maros 2001—Indonésie—Sulawesi Selatan, Rapport Spéléologique, unpublished, no date. 25p.
- Brouquisse, F.; Deharveng, L.; Laumanns, M. Indonesia 1985–2001 Expeditions of the Association Pyrénéenne de Spéléologie. Berl. Höhlenkundliche Ber. 2015, 59, 1–197. [Google Scholar]
- Acintyacunyata Speleological Club. Laporan Ekspedisi Maros 1989—Sulawesi Selatan, Unpublished, no date. 1–146.
- Suhardjono, Y.R.; Rahmadi, C.; Nugroho, H.; Wiantoro, S. Bab 2 Karst dan Gua. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 13–52. [Google Scholar]
- Bedos, A.; Deharveng, L.; Deharveng, L.; Leclerc, P.; Rigal, D.; Solier, P. 3. Résultats spéléologiques. In Expéditions Maros 88-Maros 89, Rapport Spéléologique; APS: Toulouse, France, 1990; pp. 15–48. [Google Scholar]
- Brouquisse, F.; Lacas, M.; Rigal, D. 5. Sulawesi: Résultats spéléologiques. In Expédition Indonésie 90, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1992; pp. 37–82. [Google Scholar]
- Bedos, A.; Brouquisse, F.; Deharveng, L.; Leclerc, P.; Rigal, D. 4. Grandes Cavités du Karst de Maros. In Indonésie 92, Rapport Spéléologique; APS: Toulouse, France, 2001; pp. 39–45. [Google Scholar]
- Brehier, F. Reconnaissance de quelques siphons du karst de Maros. Maros 2001—Indonésie—Sulawesi Selatan, Rapport Spéléologique, Unpublished. 16–20.
- Rigal, D. Leang Assuloang. Maros 2001—Indonésie—Sulawesi Selatan, Rapport Spéléologique, Unpublished. 7–10.
- Rigal, D.; Lacas, M. 3. Nouvelles découvertes sur le karst de Maros. In Indonésie 92, Rapport Spéléologique; APS: Toulouse, France, 2001; pp. 15–37. [Google Scholar]
- Brouquisse, F.; Brouquisse, R. 4. Résultats spéléologiques. In Expédition Maros 94, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1997; pp. 14–28. [Google Scholar]
- Brouquisse, F. 9. Le secteur de Kappang et le réseau de Gua Salukkan Kallang. In Expédition Thaï-Maros 85, Rapport Spéléologique et scientifique; APS: Toulouse, France, 1986; pp. 68–80. [Google Scholar]
- Rigal, D. Gua Salukkan Kallang, karst de Maros, Célèbes Sud, Indonésie. Spelunca 1987, 28, 32–39. [Google Scholar]
- Brouquisse, F.; Rigal, D. 6. Résultats spéléologiques Sulawesi. In Expédition Thaï–Maros 86, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1987; pp. 47–74. [Google Scholar]
- Brouquisse, F.; Dalger, D. 8. Hydrogéochimie. In Expédition Thaï-Maros 86, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1987; pp. 85–96. [Google Scholar]
- Deharveng, L. 10. Programme zoologique: Bilan général et principaux résultats. In Expédition Thaï-Maros 86, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1987; pp. 111–116. [Google Scholar]
- Galletti, I. Nota biospeleologica della spedizione Sulawesi ‘94. Speleologia Iblea 1996, 4, 98–101. [Google Scholar]
- Deharveng, L.; Bedos, A. Salukkan Kallang, Indonesia: Biospeleology. In Encyclopedia of Cave and Karst Science; Gunn, J., Ed.; Fitzroy Dearborn: London, UK, 2004; pp. 631–633. [Google Scholar]
- Suhardjono, Y.R. Review of Biospeleology in Sulawesi Island. In Proceedings of the International Symposium on The Ecology and Limnology of the Malili Lakes, Bogor, Indonesia, 20–23 March 2006; pp. 29–38. [Google Scholar]
- Chapman, P. The ecology of caves in the Gunung Mulu National Park, Sarawak. Trans. British Cave Research Assoc. 1982, 9, 142–162. [Google Scholar]
- Brancelj, A.; Boonyanusith, C.; Watiroyram, S.; Sanoamuang, L.O. The groundwater-dwelling fauna of Southeast Asia. J. Limnol. 2013, 72, e16. [Google Scholar] [CrossRef] [Green Version]
- Kawakatsu, M.; Mitchell, R.W. Two new freshwater cavernicole planarians (Turbellaria, Tricladida, Paludicola) from Sulawesi (Celebes), Indonesia. Spec. Bull. Jpn. Soc. Coleopterol. 1995, 4, 81–104. [Google Scholar]
- WoRMS Editorial Board. World Register of Marine Species. Available online: https://www.marinespecies.orgatVLIZ (accessed on 15 June 2021).
- Harvey, M.S. Whip Spiders of the World, Version 1.0. Western Australian Museum, Perth. 2013. Available online: http://www.museum.wa.gov.au/catalogues/whip-spiders (accessed on 13 June 2021).
- World Spider Catalog. Version 22.5. Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 5 July 2021).
- Bu, Y.; Souza, M.F.V.R.; Mayoral, J. New and interesting palpigrades (Arachnida, Palpigradi) of the genera Koeneniodes Silvestri, 1913 and Prokoenenia Börner, 1901 from Asia. Zootaxa 2021, 4990, 45–64. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World 1996–2021. Available online: http://www.collembola.org (accessed on 10 June 2021).
- Sendra, A.; Jiménez-Valverde, A.; Rochat, J.; Legros, V.; Gasnier, S.; Cazanove, G. A new and remarkable troglobitic Lepidocampa Oudemans, 1890 species from La Réunion Island, with a discussion on troglobiomorphic adaptations in campodeids (Diplura). Zoologischer Anzeiger 2017, 266, 95–104. [Google Scholar] [CrossRef]
- Faille, A. Les Coléoptères troglobies de l'île de Sulawesi (Indonésie); description du mâle du Paussidae cavernicole Eustra saripaensis Deuve, 2002 (Coleoptera). Bull. Soc. Entomol. Fr. 2010, 115, 375–380. [Google Scholar]
- Deuve, T. Sur une population différenciée de Mateuellus troglobioticus (Deuve, 1990) dans le sud de Sulawesi (Col., Caraboidea, Harpalidae, Pterostichinae, Abacetini). Bull. Soc. Entom. Fr. 2010, 115, 310. [Google Scholar]
- Cigliano, M.M.; Braun, H.; Eades, D.C.; Otte, D. Orthoptera Species File. Version 5.0/5.0. Available online: http://Orthoptera.SpeciesFile.org (accessed on 5 July 2021).
- Integrated Taxonomic Information System (ITIS). Available online: www.itis.gov (accessed on 4 July 2021).
- Botosaneanu, L. New stygobiontic isopods (Isopoda: Cirolanidae, Anthuridae) from caves in Sulawesi, Indonesia. Bull. Inst. R. Sci. Nat. Belg. 2003, 73, 91–105. [Google Scholar]
- Wowor, D.; Rahmadi, C. Bab 8 Krustasea. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 165–190. [Google Scholar]
- Cai, Y.; Ng, P.K.L. Marosina, a New Genus of Troglobitic Shrimps (Decapoda, Atyidae) from Sulawesi, Indonesia, with Descriptions of Two New Species. Crustaceana 2005, 78, 129–139. [Google Scholar]
- Cai, Y.; Ng, P.K.L. The freshwater shrimps of the genera Caridina and Parisia from karst caves of Sulawesi Selatan, Indonesia, with descriptions of three new species (Crustacea: Decapoda: Caridea: Atyidae. J. Nat. Hist. 2009, 43, 1093–1114. [Google Scholar] [CrossRef]
- von Rintelen, K.; Glaubrecht, M.; Schubart, C.; Wessel, A.; von Rintelen, T. Adaptive radiation and ecological diversification of Sulawesi’s ancient lake shrimps. Evolution 2010, 64, 3287–3299. [Google Scholar] [CrossRef]
- von Rintelen, K.; Page, T.J.; Cai, Y.; Roe, K.; Stelbrink, B.; Kuhajda, B.R.; Iliffe, T.M.; Hughes, J.; von Rintelen, T. Drawn to the dark side: A molecular phylogeny of freshwater shrimps (Crustacea: Decapoda: Caridea: Atyidae) reveals frequent cave invasions and challenges current taxonomic hypotheses. Mol. Phylogenet. Evol. 2012, 63, 82–96. [Google Scholar] [CrossRef]
- Chia, O.K.S.; Ng, P.K.L. The freshwater crabs of Sulawesi, with descriptions of two new genera and four new species (Crustacea: Decapoda: Brachyura: Parathelphusidae). Raffles Bull. Zool. 2006, 54, 381–428. [Google Scholar]
- Ng, P.K.L. Cancrocaeca xenomorpha, new genus and species, a blind troglomorphic freshwater hymenosomatid (Crustacea: Decapoda: Brachyura) from Sulawesi, Indonesia. Raffles Bull. Zool. 1991, 39, 59–73. [Google Scholar]
- Hoese, D.F.; Kottelat, M. Bostrychus microphthalmus, a new microphthalmic cavefish from Sulawesi (Teleostei: Gobiidae). Ichthyol. Explor. Freshwaters 2005, 16, 183–191. [Google Scholar]
- Hadiaty, R.K. Bab 5 Ikan. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 89–113. [Google Scholar]
- Proudlove, G. Subterranean Fishes of the World. Available online: https://cavefishes.org.uk (accessed on 14 June 2021).
- Marwoto, R.M.; Isnaningsih, N.R. Bab 6 Molluska. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 115–148. [Google Scholar]
- Brescovit, A.D.; Zampaulo, R.D.A.; Cizauskas, I. The first two blind troglobitic spiders of the genus Ochyrocera from caves in Floresta Nacional de Carajás, state of Pará, Brazil (Araneae, Ochyroceratidae). Zookeys 2021, 1031, 143. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Rahmadi, C. A troglomorphic spider from Java (Araneae, Ctenidae, Amauropelma). Zookeys 2012, 163, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeleman-Reinhold, C.L. The Ochyroceratidae of the Indo-Pacific region (Araneae). Raffles Bull. Zool. 1995, 43 (Suppl. 2), 1–103. [Google Scholar]
- Huber, B.A. Revision of the genus Spermophora Hentz in Southeast Asia and on the Pacific Islands, with descriptions of three new genera (Araneae: Pholcidae). Zool. Meded. 2005, 79, 61–114. [Google Scholar]
- Huber, B.A.; Caspar, K.R.; Eberle, J. New species reveal unexpected interspecific microhabitat diversity in the genus Uthina Simon, 1893 (Araneae: Pholcidae). Invertebr. Syst. 2019, 33, 181–207. [Google Scholar] [CrossRef]
- Abrams, K.M.; Huey, J.A.; Hillyer, M.J.; Humphreys, W.F.; Didham, R.K.; Harvey, M.S. Too hot to handle: Cenozoic aridification drives multiple independent incursions of Schizomida (Hubbardiidae) into hypogean environments. Mol. Phylogenet. Evol. 2019, 139, 106532. [Google Scholar] [CrossRef]
- Rahmadi, C. Bab 9 Arthropoda Gua. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 191–214. [Google Scholar]
- Kraepelin, K. Das Tierreich 8—Scorpiones und Pedipalpi; R. Friedländer und Sohn: Berlin, Germany, 1899; pp. 1–265. [Google Scholar]
- Harvey, M.S.; West, P.L.J. New species of Charon (Amblypygi, Charontidae) from northern Australia and Christmas Island. J. Arachnol. 1998, 26, 273–284. [Google Scholar]
- Rahmadi, C.; Harvey, M.S.; Kojima, J.I. The status of the whip spider subgenus Neocharon (Amblypygi: Charontidae) and the distribution of the genera Charon and Stygophrynus. J. Arachnol. 2011, 39, 223–229. [Google Scholar] [CrossRef]
- Golovatch, S.I.; Geoffroy, J.J.; Mauriès, J.P.; van den Spiegel, D. Review of the millipede genus Eutrichodesmus Silvestri, 1910 (Diplopoda, Polydesmida, Haplodesmidae), with descriptions of new species. Zookeys 2009, 12, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Dalens, H. 12. Données préliminaires sur les Isopodes terrestres récoltés dans les grottes de Sulawesi et des Moluques. In Expédition Thaï-Maros 86, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1987; pp. 129–132. [Google Scholar]
- Suhardjono, Y.R. Bab 11 Ekorpegas. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 227–246. [Google Scholar]
- Deharveng, L.; Suhardjono, Y.R. Pseudosinella maros sp. n., a troglobitic Entomobryidae (Collembola) from Sulawesi Selatan, Indonesia. Rev. Suisse Zool. 2004, 111, 979–984. [Google Scholar] [CrossRef]
- Cipola, N.G.; Oliveira, J.V.L.C.; Bellini, B.C.; Ferreira, A.S.; Lima, E.C.A.; Brito, R.A.; Stievano, L.C.; Souza, P.G.C.; Zeppelini, D. Review of Eyeless Pseudosinella Schäffer (Collembola, Entomobryidae, Lepidocyrtinae) from Brazilian Caves. Insects 2020, 11, 194. [Google Scholar] [CrossRef] [Green Version]
- Deharveng, L. 4. La faune souterraine de Batu Lubang. In Expédition Batukarst 88, Rapport Spéléologique et Scientifique; APS: Toulouse, France, 1989; pp. 37–46. [Google Scholar]
- Deharveng, L.; Whitten, T.; Leclerc, P. 5.13. Caves of Papua. In The Ecology of Papua; Marshall, A., Beehler, B., Eds.; Periplus: Budapest, Hungary, 2007; pp. 1064–1082. [Google Scholar]
- Condé, B. Campodéides des grottes des Célèbes (Insectes, Diploures). Mém. Biospéol. 1992, 19, 155–158. [Google Scholar]
- Allegrucci, G.; Trewick, S.A.; Fortunato, A.; Carchini, G.; Sbordoni, V. Cave crickets and cave weta (Orthoptera, Rhaphidophoridae) from the southern end of the World: A molecular phylogeny test of biogeographical hypotheses. J. Orthoptera Res. 2010, 19, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Rheindt, F.E.; Norman, J.A.; Christidis, L. Extensive diversification across islands in the echolocating Aerodramus swiftlets. Raffles Bull. Zool. 2014, 62, 89–99. [Google Scholar]
- Suyanto, A.; Wiantoro, S. Bab 3 Kelelawar. In Fauna Karst dan gua Maros, Sulawesi Selatan; Suhardjono, Y.R., Ubaidillah, R., Eds.; LIPI Press: Jakarta, Indonesia, 2012; pp. 53–76. [Google Scholar]
- Chapman, P. Species diversity in a tropical cave ecosystem. Proc. Univ. Bristol Spelaeol. Soc. 1983, 16, 201–213. [Google Scholar]
- Coiffait, H. Les Coléoptères du sol. Vie et Milieu 1958, 9 (Suppl. 7), 1–204. [Google Scholar]
- Beron, P. Comparative study of the invertebrate cave faunas of Southeast Asia and New Guinea. Hist. Nat. Bulg. 2015, 21, 169–210. [Google Scholar]
- Souza Silva, M.; Ferreira, R.L. The first two hotspots of subterranean biodiversity in South America. Subterr. Biol. 2016, 19, 1–21. [Google Scholar] [CrossRef]
- Trajano, E.; Gallão, J.E.; Bichuette, M.E. Spots of high diversity of troglobites in Brazil: The challenge of measuring subterranean diversity. Biodivers. Conserv. 2016, 25, 1805–1828. [Google Scholar] [CrossRef]
- Jeannel, R.; Racovitza, E.G. Biospeologica XXXIX. Enumération des grottes visitées 1913–1917 (sixième série). Arch. Zool. Exp. Gen. 1918, 57, 203–470. [Google Scholar]
- Deharveng, L.; Lips, J.; Rahmadi, C. Focus on guano. In The Natural History of Santo: Caves and Soils; Bouchet, P., Le Guyader, H., Pascal, O., Eds.; MNHN; IRD; PNI: Paris, France, 2011; pp. 300–305. [Google Scholar]
- Hanitsch, R. On some cave-dwelling Blattids from Celebes. Tijdschr. Entomol. 1932, 75, 264–265. [Google Scholar]
Figure 1.
Karst landscape of the Maros karst; (A) corridors generated by volcanic dykes in the Towakkalak area (Google Earth); (B) Batuputte giant shafts (Google Earth); (C) limestone cliffs NW of Bantimurung (Leang Leang valley); (D) volcanic sill topped with limestone near Pangea. Reproduced with permission from Louis Deharveng (C,D).
Figure 1.
Karst landscape of the Maros karst; (A) corridors generated by volcanic dykes in the Towakkalak area (Google Earth); (B) Batuputte giant shafts (Google Earth); (C) limestone cliffs NW of Bantimurung (Leang Leang valley); (D) volcanic sill topped with limestone near Pangea. Reproduced with permission from Louis Deharveng (C,D).

Figure 2.
(A) Lubang Leaputte; (B) Gua Salukkan Kallang underground river; (C) Bantimurung waterfall, downstream Towakkalak spring; (D) Fossil gallery in Gua Tanette; (E) underground river in Gua Tanette. Reproduced with permission from Didier Rigal.
Figure 2.
(A) Lubang Leaputte; (B) Gua Salukkan Kallang underground river; (C) Bantimurung waterfall, downstream Towakkalak spring; (D) Fossil gallery in Gua Tanette; (E) underground river in Gua Tanette. Reproduced with permission from Didier Rigal.

Figure 3.
Hydrogeological synthesis of the Maros karst east of Bantimurung, including Towakkalak and Saripa systems [35] updated. Upper left: map of Indonesia with the localization of the studied region in Sulawesi. Legend. Limestone, area filled with rectangles; recent alluvial deposit, area filled with dots, yellow background; basalt (ß), orange; diorite (d), grey; volcanic sediments of the Camba formation, pale green. Square, village; cross, elevation point; empty blue circle, stream sink; filled blue circle, karstic spring; blue line, surface river; blue arrow, underground river; filled red circle, big shaft; red line, explored cave; dotted line, underground water passage, proven by coloration or hypothesized. Encircled numbers, caves and springs (white on black, sampled caves; black on white, large unsampled caves): 1, Gua Burung Salangan Geram; 2, Gua Salukkan Kallang; 3, K11 spring; 4, Gua K9; 5, Gua Alolu; 6, Gua Broukiss; 7, Lubang Kabut; 8, Lubang Batu Neraka; 9, Gua Tanette (entrance); 10, Gua Wattanang (a) and Gua Uri (b); 11, Towakkalak spring; 12, Gua Lumpur (a) and Gua Bantimurung (b); 13, Gua B2 and Gua B3; 14, Jamala spring; 15, Gua Mimpi and Gua Istani Toakala; 16, Gua Baharuddin; 17, Leang Saripa and Saripa spring; 18, Gua Restauran; 19, sinks of the Sungai Gallang.
Figure 3.
Hydrogeological synthesis of the Maros karst east of Bantimurung, including Towakkalak and Saripa systems [35] updated. Upper left: map of Indonesia with the localization of the studied region in Sulawesi. Legend. Limestone, area filled with rectangles; recent alluvial deposit, area filled with dots, yellow background; basalt (ß), orange; diorite (d), grey; volcanic sediments of the Camba formation, pale green. Square, village; cross, elevation point; empty blue circle, stream sink; filled blue circle, karstic spring; blue line, surface river; blue arrow, underground river; filled red circle, big shaft; red line, explored cave; dotted line, underground water passage, proven by coloration or hypothesized. Encircled numbers, caves and springs (white on black, sampled caves; black on white, large unsampled caves): 1, Gua Burung Salangan Geram; 2, Gua Salukkan Kallang; 3, K11 spring; 4, Gua K9; 5, Gua Alolu; 6, Gua Broukiss; 7, Lubang Kabut; 8, Lubang Batu Neraka; 9, Gua Tanette (entrance); 10, Gua Wattanang (a) and Gua Uri (b); 11, Towakkalak spring; 12, Gua Lumpur (a) and Gua Bantimurung (b); 13, Gua B2 and Gua B3; 14, Jamala spring; 15, Gua Mimpi and Gua Istani Toakala; 16, Gua Baharuddin; 17, Leang Saripa and Saripa spring; 18, Gua Restauran; 19, sinks of the Sungai Gallang.


Figure 5.
(A) Cirolana marosina from Leang Saripa; (B) Marosina longirostris from Leang Saripa; (C) Cancrocaeca xenomorpha from Lubang Batu Neraka; (D) Parathelphusa sorella from Leang Saripa; (E) Bostrychus microphthalmus from Gua Tanette. Reproduced with permission from Louis Deharveng (A,D); Jean-Yves Rasplus (B); Didier Rigal (C,E).
Figure 5.
(A) Cirolana marosina from Leang Saripa; (B) Marosina longirostris from Leang Saripa; (C) Cancrocaeca xenomorpha from Lubang Batu Neraka; (D) Parathelphusa sorella from Leang Saripa; (E) Bostrychus microphthalmus from Gua Tanette. Reproduced with permission from Louis Deharveng (A,D); Jean-Yves Rasplus (B); Didier Rigal (C,E).

Figure 6.
(A) Heteropoda beroni from Leang Saripa; (B) cf. Amauropelma sp. from Saripa spring cave; (C) schizomid from Gua Tanette; (D) Charon sp. from Saripa spring cave; (E) Eutrichodesmus reductus from Leang Saripa; (F) Hypocambala helleri from Leang Saripa. Reproduced with permission from Jean-Yves Rasplus (A,E) and Louis Deharveng (B–D,F).
Figure 6.
(A) Heteropoda beroni from Leang Saripa; (B) cf. Amauropelma sp. from Saripa spring cave; (C) schizomid from Gua Tanette; (D) Charon sp. from Saripa spring cave; (E) Eutrichodesmus reductus from Leang Saripa; (F) Hypocambala helleri from Leang Saripa. Reproduced with permission from Jean-Yves Rasplus (A,E) and Louis Deharveng (B–D,F).

Figure 7.
(A) Venezillo sp. from Leang Saripa; (B) Papuaphiloscia sp. from Leang Saripa; (C) Pseudosinella maros from Gua Lumpur; (D) Oncopodura sp. from Gua Tanette; (E) Pararrhopalites sp. from Gua Tanette. Reproduced with permission from Louis Deharveng.
Figure 7.
(A) Venezillo sp. from Leang Saripa; (B) Papuaphiloscia sp. from Leang Saripa; (C) Pseudosinella maros from Gua Lumpur; (D) Oncopodura sp. from Gua Tanette; (E) Pararrhopalites sp. from Gua Tanette. Reproduced with permission from Louis Deharveng.

Figure 8.
(A) Rhaphidophora sp. from Saripa spring cave; (B) Eustra saripaensis from Leang Saripa; (C) Nocticolidae sp. from Leang Saripa; (D) Mateuellus troglobioticus from Leang Saripa. Reproduced with permission from Anne Bedos (A), Jean-Yves Rasplus (B,C) and Louis Deharveng (D).
Figure 8.
(A) Rhaphidophora sp. from Saripa spring cave; (B) Eustra saripaensis from Leang Saripa; (C) Nocticolidae sp. from Leang Saripa; (D) Mateuellus troglobioticus from Leang Saripa. Reproduced with permission from Anne Bedos (A), Jean-Yves Rasplus (B,C) and Louis Deharveng (D).


{kind=link}
{kind=link}
{kind=link}
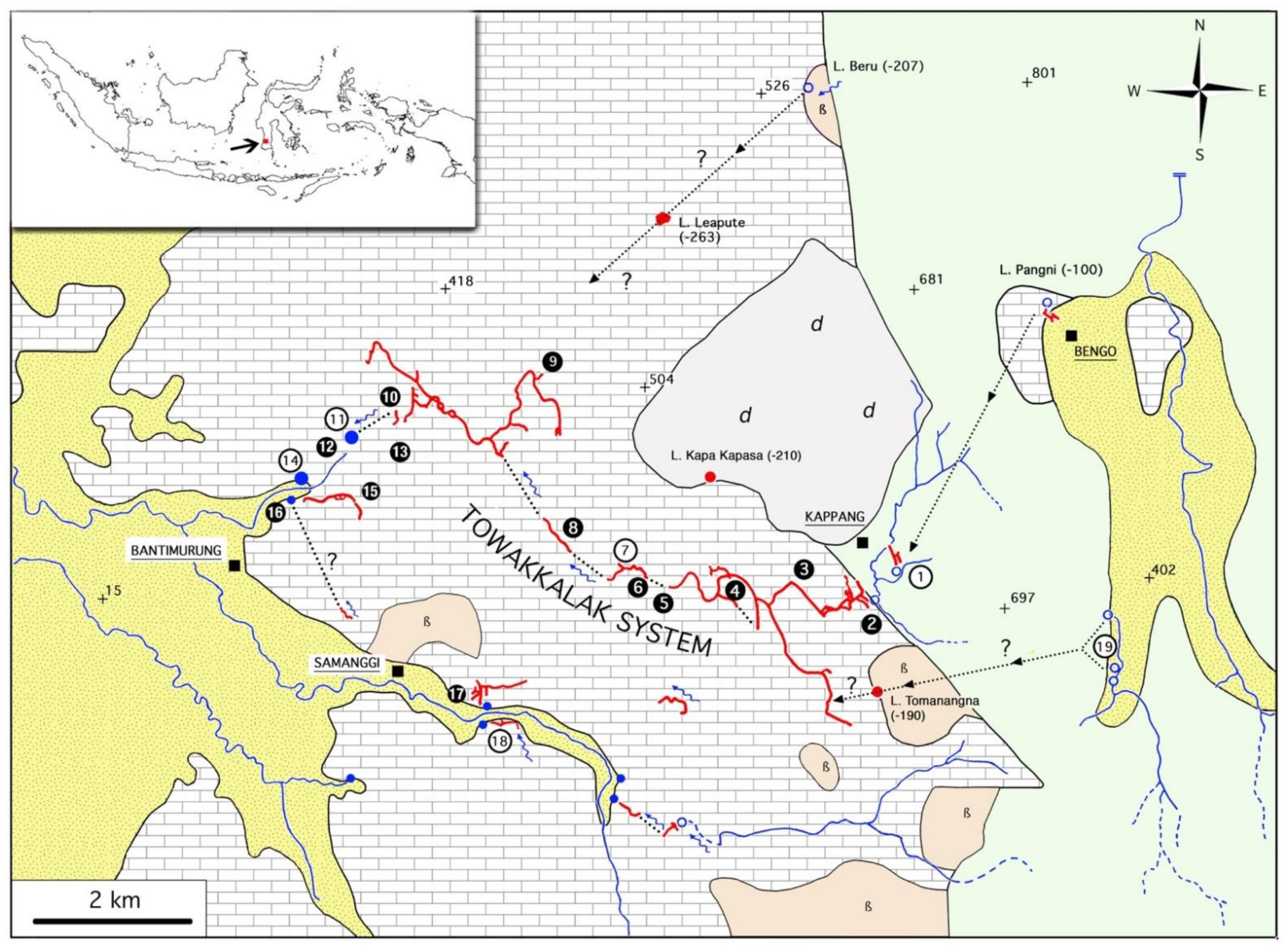{kind=link}
{kind=link}
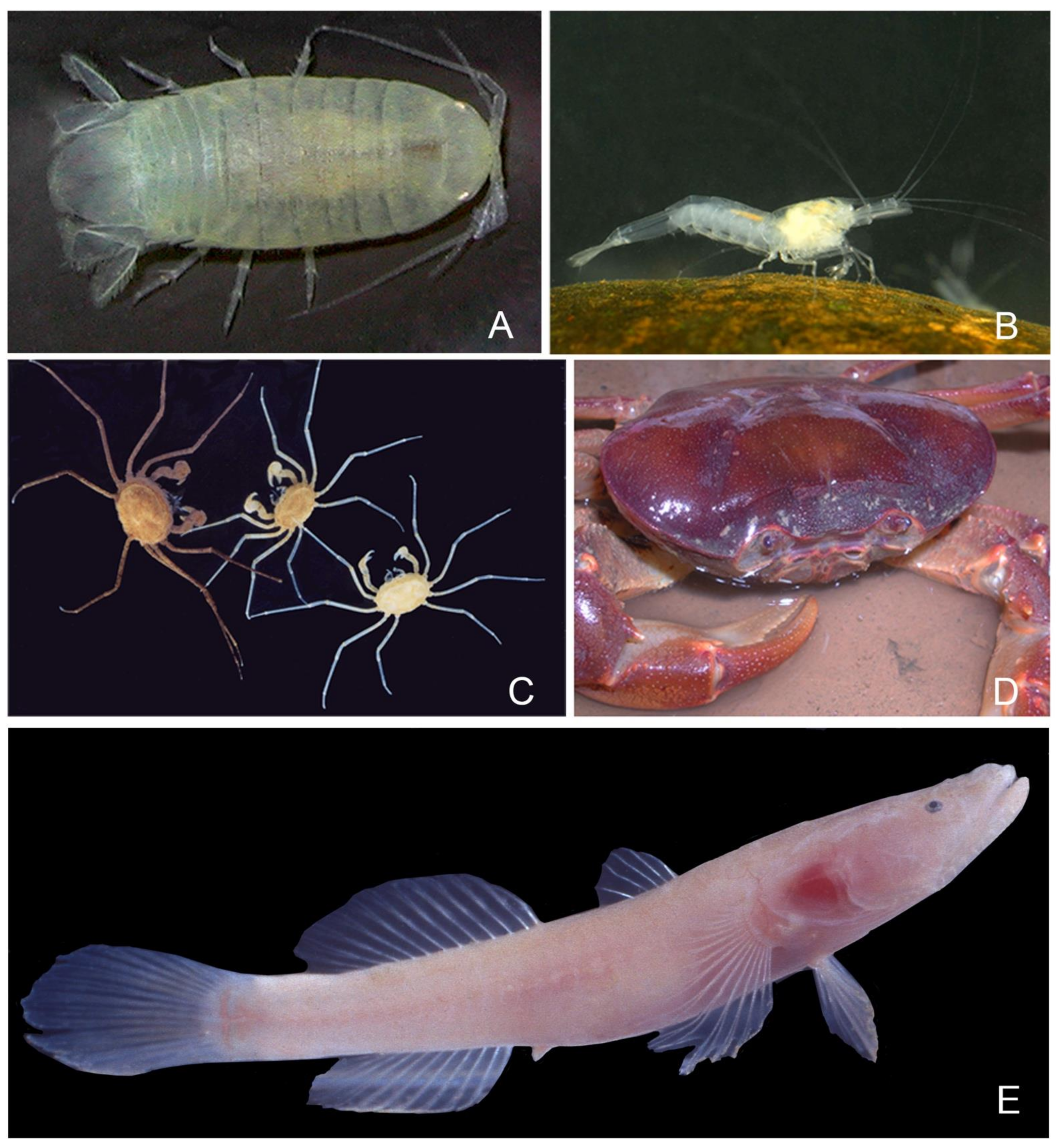{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Stygobiotic and troglobiotic species of the Towakkalak and Saripa systems. Abbreviations—Ecology: SB, stygobiont (aquatic); TB, troglobiont (terrestrial). Distribution (dist): end, restricted to the Maros karst; end *, restricted to a single system. Towakkalak: see Figure 3 for cave numbering; 10a, Gua Wattanang; 12a, Gua Lumpur; 12b, Gua Bantimurung; Pa, Lubang Pangni. Saripa: L, Leang Saripa; S, Saripa Spring; ?, uncertain identification. Number of species in the genus and taxonomic validity of the species (gen val): +, corrected numbers from various sources. n.a., not applicable.
Table 1.
Stygobiotic and troglobiotic species of the Towakkalak and Saripa systems. Abbreviations—Ecology: SB, stygobiont (aquatic); TB, troglobiont (terrestrial). Distribution (dist): end, restricted to the Maros karst; end *, restricted to a single system. Towakkalak: see Figure 3 for cave numbering; 10a, Gua Wattanang; 12a, Gua Lumpur; 12b, Gua Bantimurung; Pa, Lubang Pangni. Saripa: L, Leang Saripa; S, Saripa Spring; ?, uncertain identification. Number of species in the genus and taxonomic validity of the species (gen val): +, corrected numbers from various sources. n.a., not applicable.
| Taxonomic Group | Taxon | Ecology | Dist | Towakkalak | Saripa | Gen Val |
|---|---|---|---|---|---|---|
| Tricladida: Dugesiidae | Dugesia leclerci Kawakatsu and Mitchell, 1995 | SB | end | 9 | L?,S? | 121 [51] |
| Tricladida: Dugesiidae | Dugesia uenorum Kawakatsu and Mitchell, 1995 | SB? | end * | Pa | 121 [51] | |
| Amphipoda: Bogidiellidae | Bogidiellidae sp. | SB | end | 9 | n.a. | |
| Isopoda: Cirolanidae | Cirolana marosina Botosaneanu, 2003 | SB | end | 12a | S | 146 [51] |
| Caridea: Atyidae | Caridina leclerci Cai and Ng, 2009 | SB | end * | 2,3,9 | 344 [51] | |
| Caridea: Atyidae | Marosina brevirostris Cai and Ng, 2005 | SB | end * | 2 | 2 [51] | |
| Caridea: Atyidae | Marosina longirostris Cai and Ng, 2005 | SB | end | 2,8,9 | L | 2 [51] |
| Caridea: Atyidae | Parisia deharvengi Cai and Ng, 2009 | SB | end * | 9 | 9 [51] | |
| Brachyura: Hymenosomatidae | Cancrocaeca xenomorpha Ng, 1991 | SB | end | 8,9 | L,S | 1 [51] |
| Brachyura: Gecarcinucidae | Parathelphusa sorella Chia and Ng, 2006 | SB | end * | L | 49 [51] | |
| Pisces: Eleotridae | Bostrychus microphthalmus Hoese and Kottelat, 2005 | SB | end | 9 | L | 9 [51] |
| Acari: Leeuwenhoekiidae | Leeuwenhoekiidae sp. (cf.) | TB? | n.a. | L,S | n.a. | |
| Amblypygi: Charinidae | Sarax sp. | TB | end? | 2 | 17 [52]+ | |
| Araneae: Ctenidae | Amauropelma cf. sp. | TB | end | 4,9,12a | L | 24 [53] |
| Araneae: Psilodercidae | Psiloderces leclerci Deeleman-Rheinhold, 1995 | TB? | end | 2,4,13 | 38 [53] | |
| Araneae: Ochyroceratidae | Speocera caeca Deeleman-Rheinhold, 1995 | TB | end | 2,4,13 | L | 84 [53] |
| Araneae: Pholcidae | Spermophora maros Huber, 2005 | TB | end * | 2,9 | 45 [53] | |
| Araneae: Pholcidae | Uthina mimpi Huber, Caspar and Eberle, 2019 | TB | end | 2,15 | 17 [53] | |
| Opiliones | Opiliones sp. | TB | end | 2,9,12b,15 | n.a. | |
| Palpigradi: Eukoeneniidae | Eukoenenia maros Condé, 1992 | TB | end * | 9,13 | 93 [54] | |
| Palpigradi: Prokoeneniidae | Prokoenenia celebica Condé, 1994 | TB | end * | 2 | 6 [54] | |
| Pseudoscorpiones | Pseudoscorpiones sp. | TB | n.a. | 15 | L | n.a. |
| Schizomida | Schizomida spp. | TB | n.a. | 2,4,9,10a,15 | L | n.a. |
| Diplopoda: Haplodesmidae | Eutrichodesmus reductus Golovatch et al., 2009 | TB | end | 9 | L,S | 53 [51] |
| Diplopoda: Metopidiothrichidae | Metopidiothrix kalang Shear, 2002 | TB | end * | 2 | 39 [51] | |
| Oniscida: Philosciidae | Papuaphiloscia sp. | TB | end | 2,9,13 | L,S | 15 [51] |
| Oniscida: Armadillidae | Venezillo sp. | TB | end | 4,10a,12b,13,15 | S | 137 [51] |
| Collembola: Neanuridae | Deuterobella sp. | TB-TP? | end | 9 | L,S | 4 [55]+ |
| Collembola: Neelidae | Megalothorax sp. | TB-TP? | ? | 2 | L | 33 [55]+ |
| Collembola: Oncopoduridae | Oncopodura sp. | TB | end | 9 | L | 49 [55]+ |
| Collembola: Sminthuridae | Pararrhopalites sp. | TB | end? | 2,9,12b,15 | L | 17 [55]+ |
| Collembola: Entomobryidae | Pseudosinella maros Deharveng and Suhardjono, 2004 | TB | end | 2,4,5,9,12a,12b,15 | L,S | 352 [55]+ |
| Collembola: Entomobryidae | Sinella sp. | TB? | ? | 15 | 86 [55]+ | |
| Diplura: Campodeidae | Lepidocampa (Lepidocampa) hypogaea Condé, 1992 | TB | end | 2,4,6,9,12b,13 | 17 [56] | |
| Zygentoma: Nicoletiidae | Nicoletiidae | TB? | end | S | n.a. | |
| Blattodea: Nocticolidae | Nocticolidae sp. 1 | TB | end | 2?,9?,12a,12b?,15? | L?,S? | n.a. |
| Blattodea: Nocticolidae | Nocticolidae sp. 2 | TB | end | 2?,9,12a,12b?,15? | L,S? | n.a. |
| Coleoptera: Carabidae | Eustra saripaensis Deuve, 2002 | TB | end * | L | 28 [57] | |
| Coleoptera: Carabidae | Mateuellus troglobioticus troglobioticus Deuve, 1990 | TB | end | 6,15 | L,S | 2 [58] |
| Hemiptera: Cixiidae | Cixiidae sp. | TB? | ? | 15 | n.a. |
Table 2.
Main stygophilic and troglophilic species of the Towakkalak and Saripa systems. Abbreviations: Ecology: SP, stygophile (aquatic); TP, troglophile (terrestrial); Gu: guanophile (terrestrial); eu, euedaphic (terrestrial). Distribution (dist): cos, cosmopolite; end, restricted to the Maros karst; ipa, Indo-Pacific; pan, pantropical; sul, Sulawesi; wid, widespread. Towakkalak: see Figure 3 for cave numbering; 10b, Gua Uri; 12b, Gua Bantimurung; Pa, Lubang Pangni. Saripa: same as Table 1. Number of species in the genus and taxonomic validity of the species (gen val): same as Table 1.
Table 2.
Main stygophilic and troglophilic species of the Towakkalak and Saripa systems. Abbreviations: Ecology: SP, stygophile (aquatic); TP, troglophile (terrestrial); Gu: guanophile (terrestrial); eu, euedaphic (terrestrial). Distribution (dist): cos, cosmopolite; end, restricted to the Maros karst; ipa, Indo-Pacific; pan, pantropical; sul, Sulawesi; wid, widespread. Towakkalak: see Figure 3 for cave numbering; 10b, Gua Uri; 12b, Gua Bantimurung; Pa, Lubang Pangni. Saripa: same as Table 1. Number of species in the genus and taxonomic validity of the species (gen val): same as Table 1.
| Taxonomic Group | Taxon | Ecology | Dist | Towakkalak | Saripa | Gen Val |
|---|---|---|---|---|---|---|
| Caridea: Atyidae | Caridina parvidentata Roux, 1904 | SP | end | 9 | 344 [51] | |
| Caridea: Palaemonidae | Macrobrachium lar (Fabricius, 1798) | SP | pan | 9 | 276 [51] | |
| Brachyura: Gecarcinucidae | Parathelphusa celebensis (De Man, 1892) | SP | sul | Pa | 49 [51] | |
| Brachyura: Gecarcinucidae | Parathelphusa pareparensis (De Man, 1892) | SP | end | 2,9 | 49 [51] | |
| Pisces: Gobiidae | Glossogobius sp. | SP? | ? | 9 | 35 [51] | |
| Gastropoda: Subulinidae | Allopeas gracile (Hutton, 1834) | TP | pan | 15 | L | 21 [51] |
| Gastropoda: Subulinidae | Paropeas achatinaceum Pfeiffer, 1846 | TP | ipa | 15 | 4 [51] | |
| Amblypygi: Charontidae | Charon sp. | TP | ? | 10b,15 | L,S | 5 [52]+ |
| Araneae: Sparassidae | Heteropoda beroni Jaeger, 2005 | TP | end | 15 | L | 189 [53] |
| Araneae: Ochyroceratidae | Speocera karkari (Baert, 1980) | TP | ? | 13 | 84 [53] | |
| Diplopoda: Glomeridesmidae | Glomeridesmus sp. | TP? | ? | 15 | 28 [51] | |
| Diplopoda: Cambalopsidae | Hypocambala helleri Silvestri, 1897 | TP-TB | wid | 10b,15 | L | 15 [51] |
| Diplura: Campodeidae | Lepidocampa (L.) weberi borneensis Silvestri, 1933 | TP | wid | 2 | 17 [56] | |
| Collembola: Hypogastruridae | Acherontiella sp. | TP(eu) | ? | 15 | 20 [55]+ | |
| Collembola: Isotomidae | Isotomodes sp. | TP(eu) | ? | 15 | 36 [55]+ | |
| Collembola: Isotomidae | Folsomides centralis (Denis, 1931) | TP(Gu) | pan | 2,9 | 70 [55]+ | |
| Collembola: Isotomidae | Folsomides parvulus Stach, 1922 | TP(Gu) | cos | 2,5 | 70 [55]+ | |
| Collembola: Isotomidae | Folsomides pseudoparvulus Martynova, 1978 | TP(Gu) | pan | 2 | 70 [55]+ | |
| Collembola: Isotomidae | Folsomina onychiurina Denis, 1931 | TP(Gu) | pan | 2,5,12b | 5 [55]+ | |
| Collembola: Isotomidae | Isotomiella nummulifer Deharveng and Oliveira, 1990 | TP | pan | 2 | 55 [55]+ | |
| Collembola: Isotomidae | Isotomiella symetrimucronata Najt and Thibaud, 1988 | TP | pan | 2,15 | 55 [55]+ | |
| Collembola: Hypogastruridae | Willemia cf. buddenbrocki Hüther, 1959 | TP | ? | 2 | 47 [55]+ | |
| Collembola: Hypogastruridae | Xenylla yucatana Mills, 1938 | TP(Gu) | pan | 5 | 139 [55]+ | |
| Orthoptera: Rhaphidophoridae | Rhaphidophora sp. | TP | end | 2,9,12a,15 | L,S | 102 [59]+ |
| Coleoptera: Aderidae | Aderidae sp. | TP(Gu) | ? | 15 | L | n.a. |
| Coleoptera: Histeridae | Aeletes sp. | TP | ? | 2 | 87 [60] | |
| Coleoptera: Staphylinidae | Staphylinidae spp. | TP(Gu) | ? | 2,4,12a,12b,15 | L | n.a. |
| Lepidoptera: Tineidae | Tineidae spp. | TP(Gu) | ? | 5 | n.a. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deharveng, L.; Rahmadi, C.; Suhardjono, Y.R.; Bedos, A. The Towakkalak System, A Hotspot of Subterranean Biodiversity in Sulawesi, Indonesia. Diversity 2021, 13, 392. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080392
AMA Style
Deharveng L, Rahmadi C, Suhardjono YR, Bedos A. The Towakkalak System, A Hotspot of Subterranean Biodiversity in Sulawesi, Indonesia. Diversity. 2021; 13(8):392. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080392
Chicago/Turabian StyleDeharveng, Louis, Cahyo Rahmadi, Yayuk Rahayuningsih Suhardjono, and Anne Bedos. 2021. "The Towakkalak System, A Hotspot of Subterranean Biodiversity in Sulawesi, Indonesia" Diversity 13, no. 8: 392. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080392
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.