
Relationship between the Microbiome and Indoor Temperature/Humidity in a Traditional Japanese House with a Thatched Roof in Kyoto, Japan

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
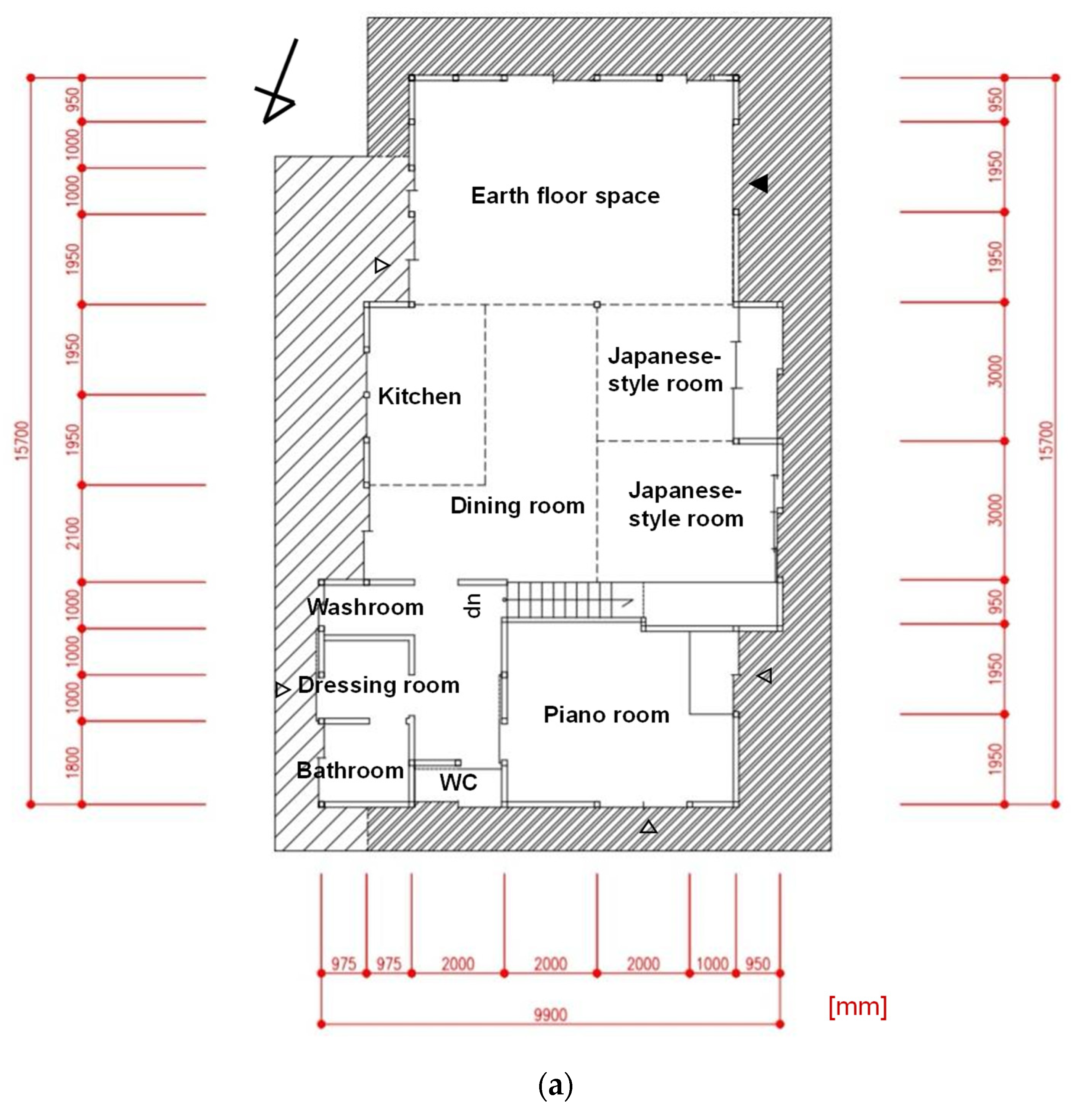
2.1. Research Site and Ways of Living
2.2. Temperature/Humidity Measurements
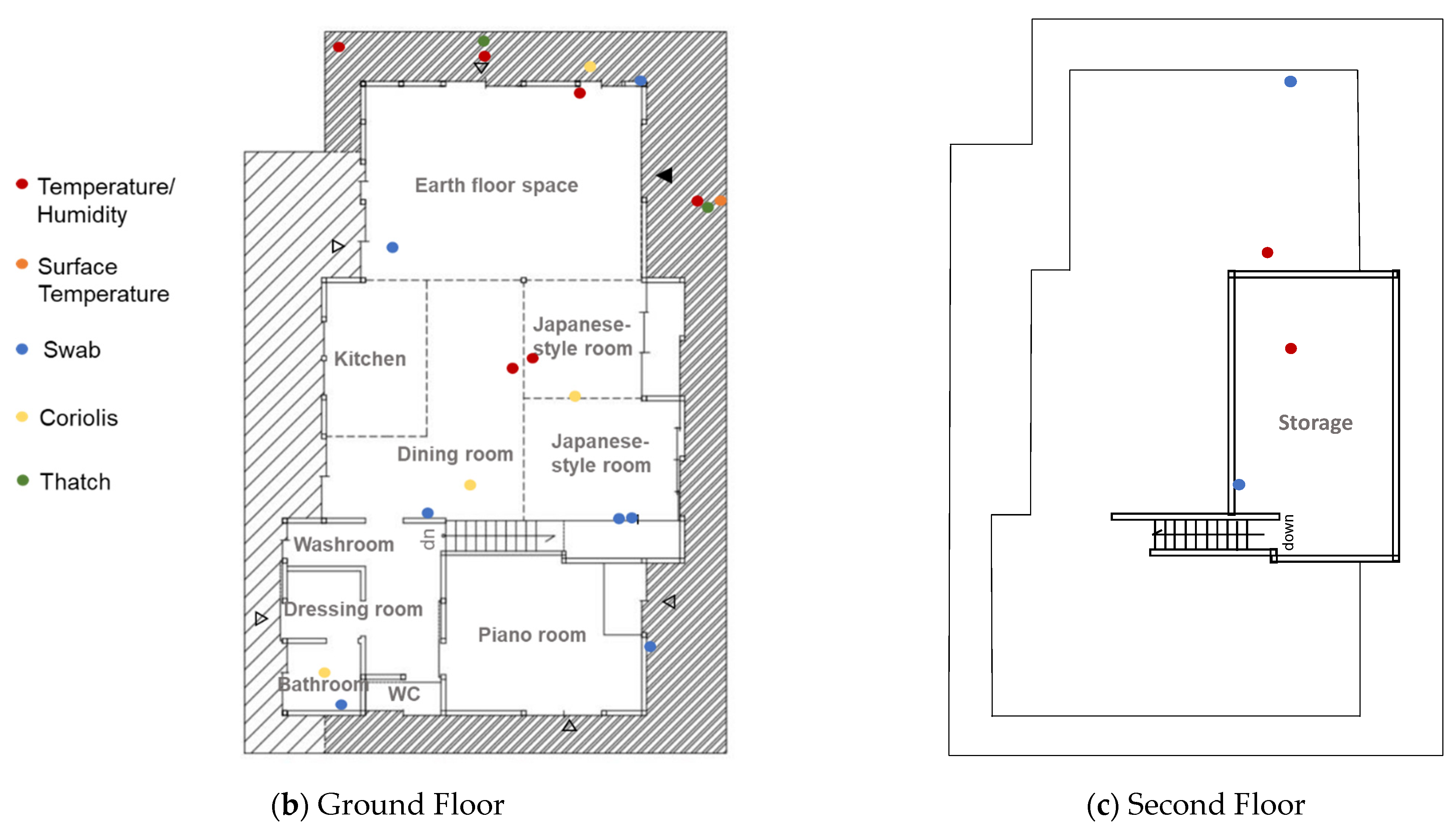
2.3. Microbial Sampling
2.4. Numerical Analysis of Surface Temperature/Humidity
2.5. DNA Extraction and Sequencing
2.6. Data Analysis and Availability
3. Results and Discussion
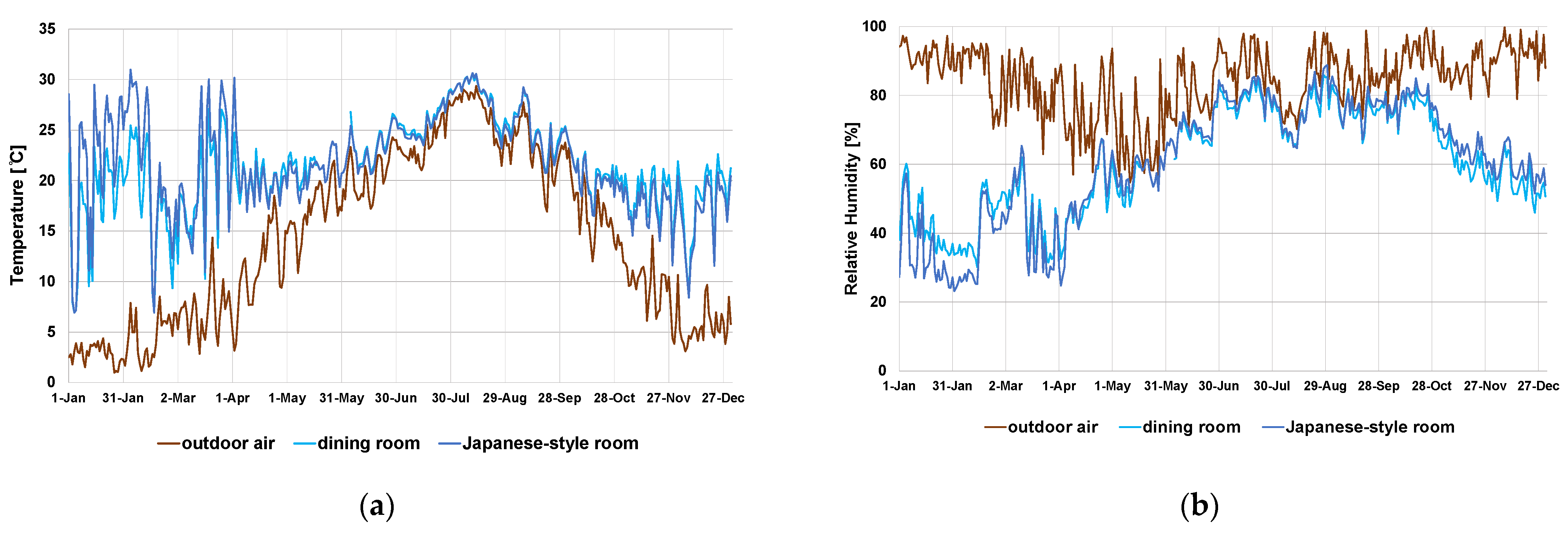
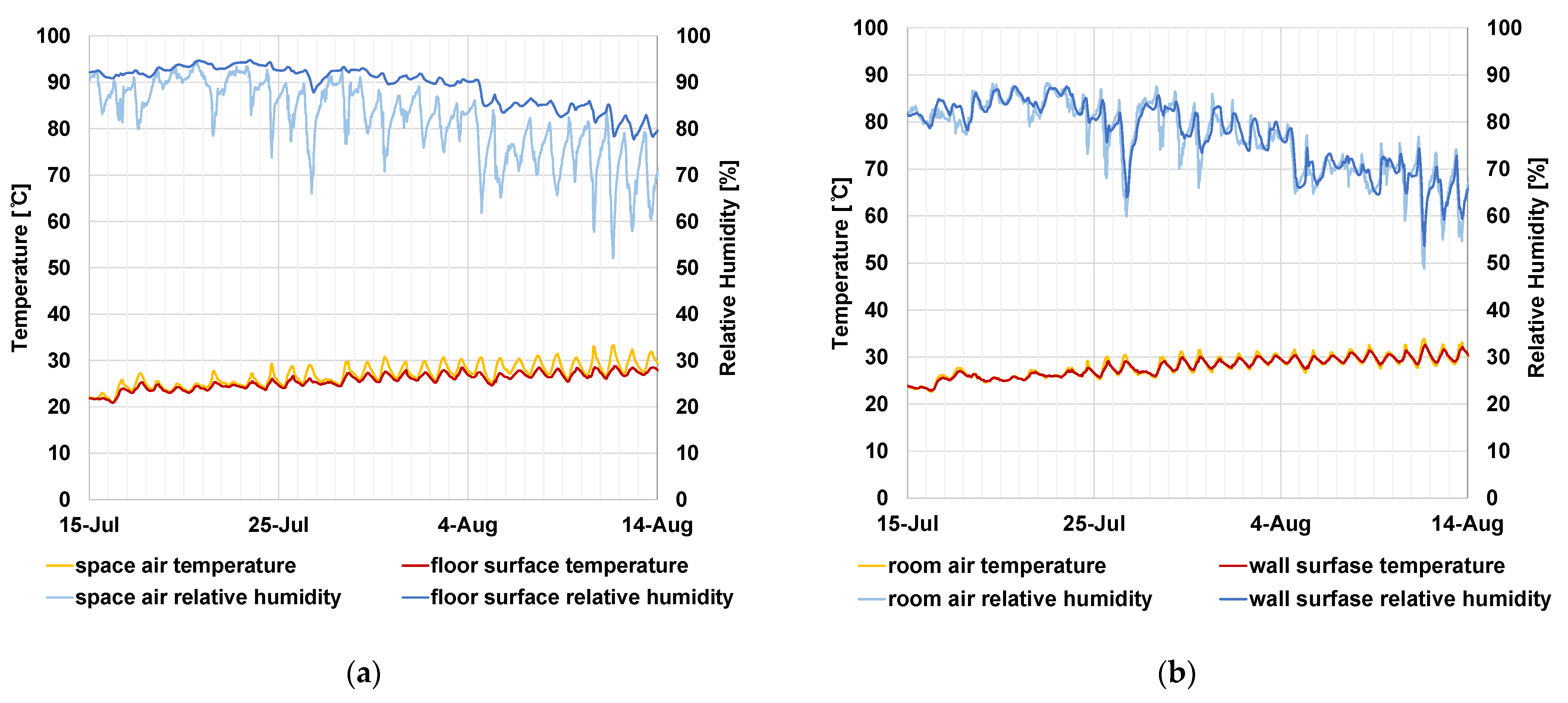
3.1. Outdoor Air and Indoor Air Temperature/Humidity
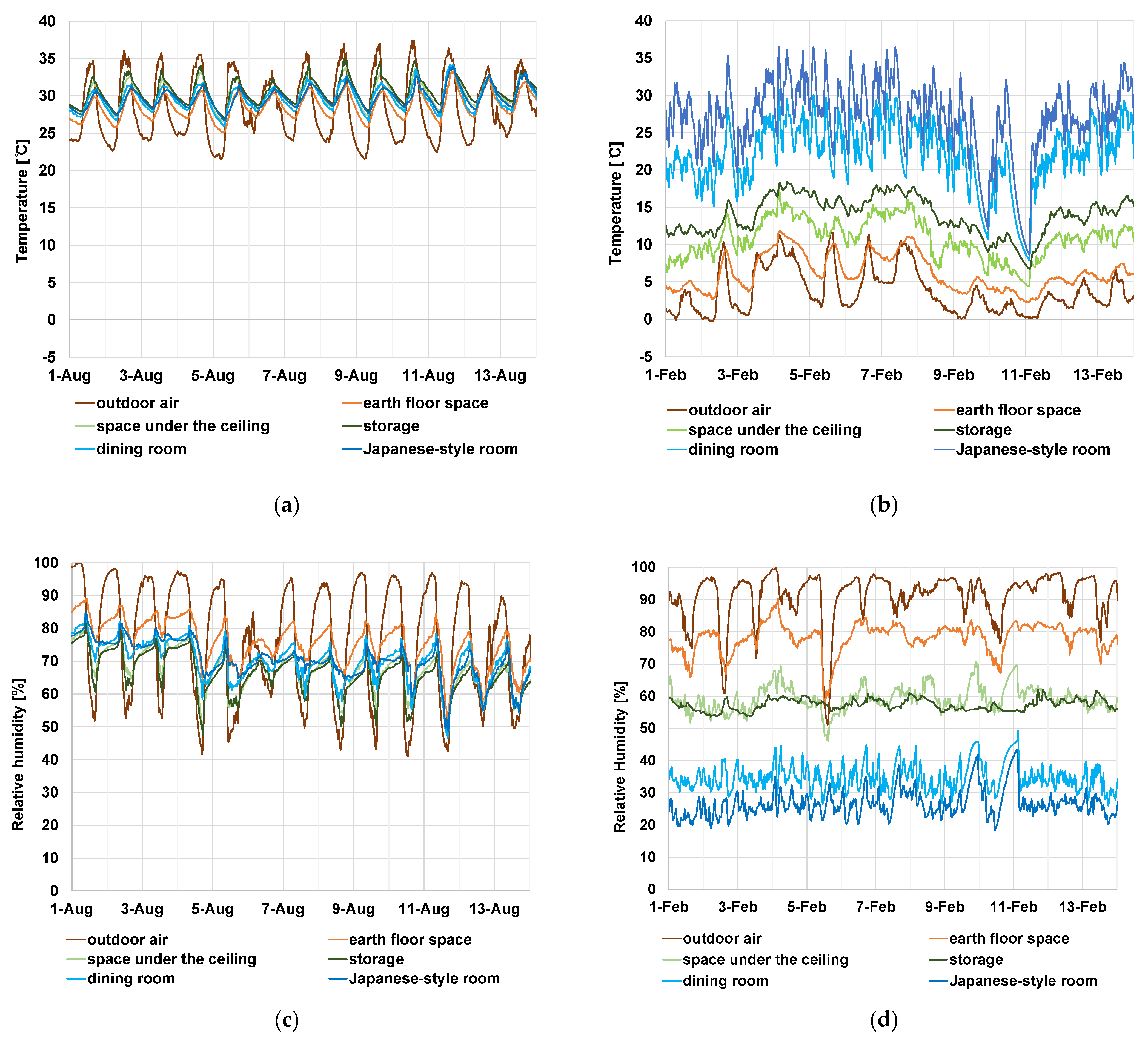
3.2. Result of Numerical Analysis of Surface Temperature/Humidity
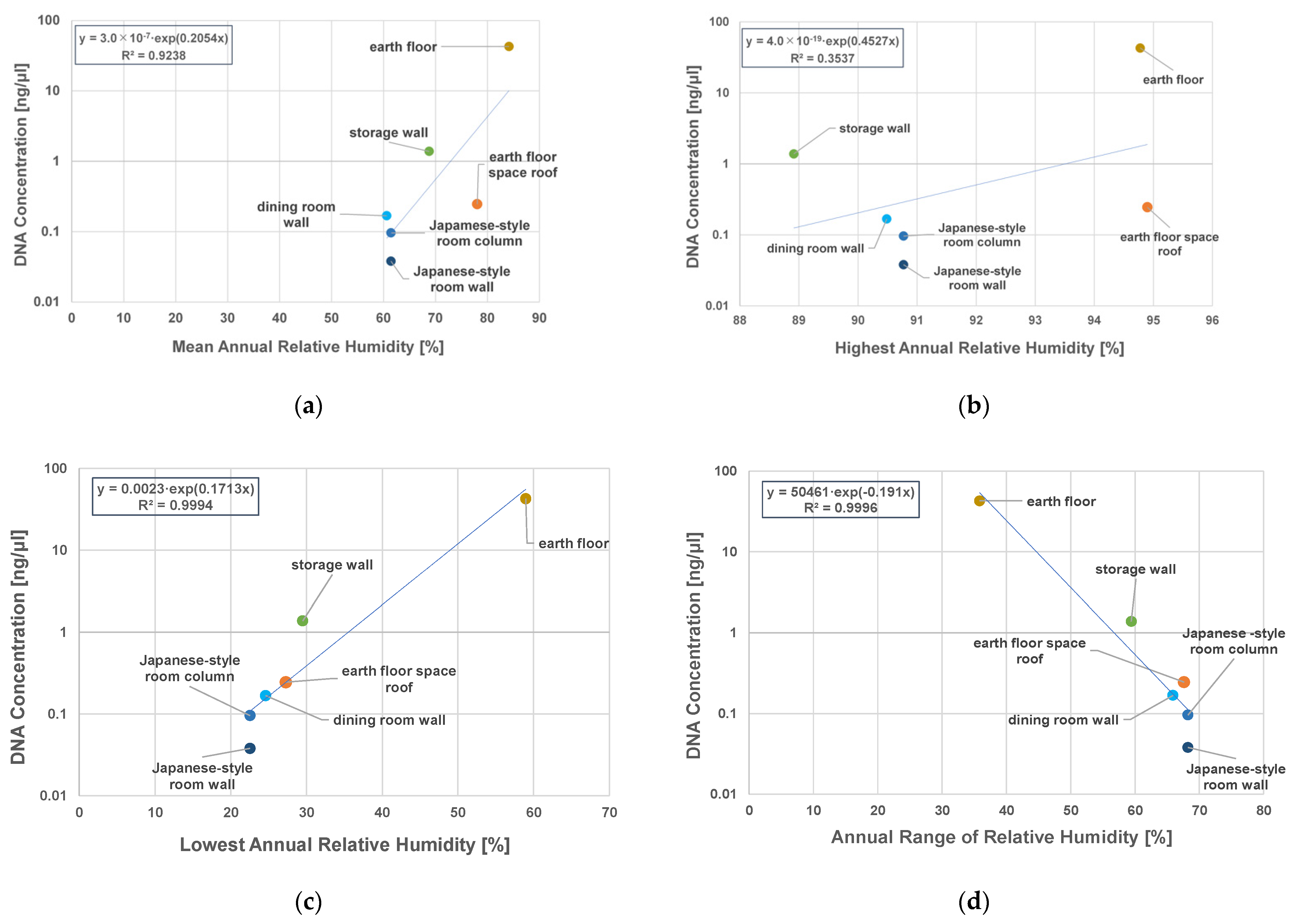
3.3. DNA Concentrations and Relative Humidity of Indoor Surfaces
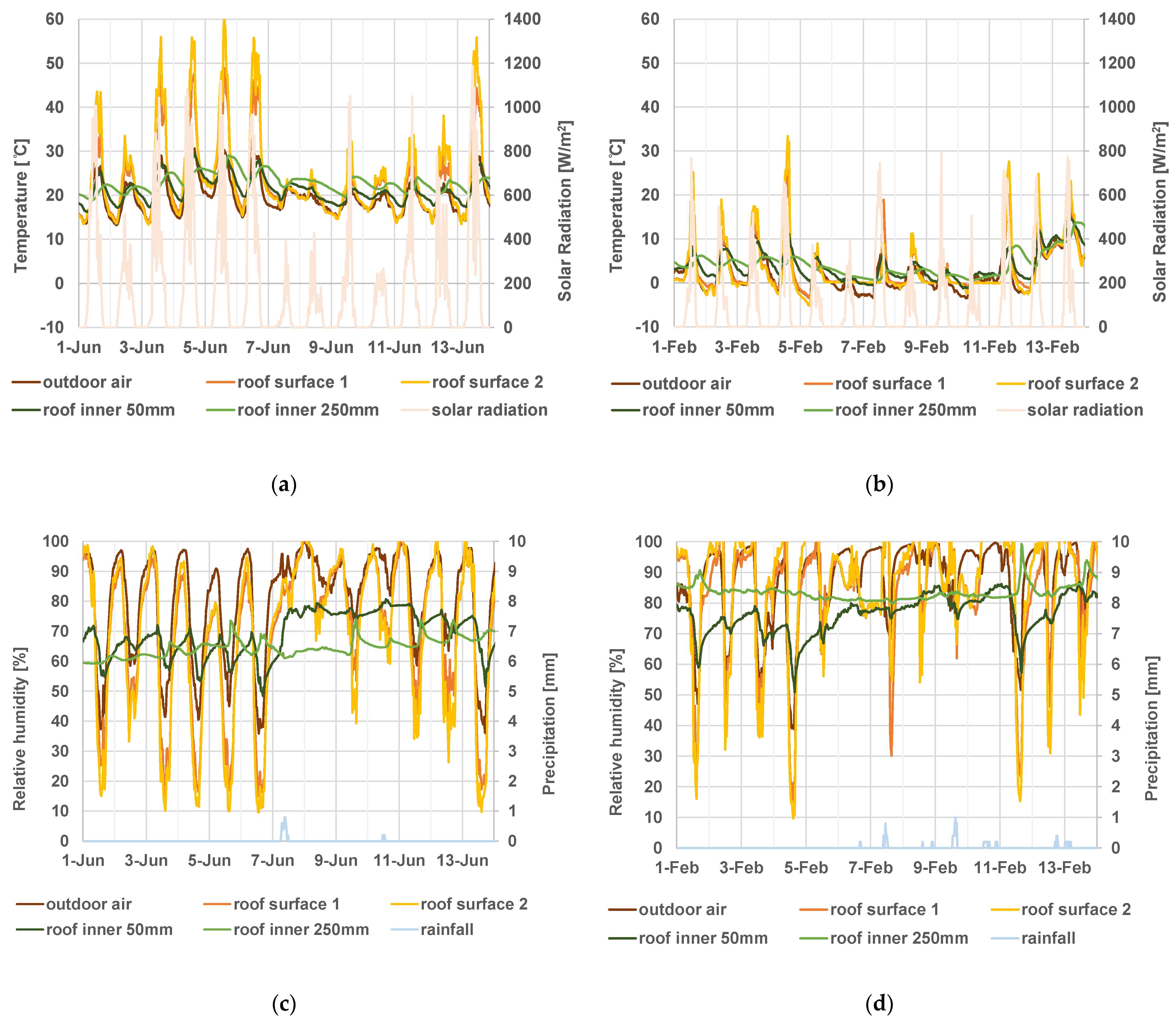
3.4. Thatched Roof Temperature/Humidity
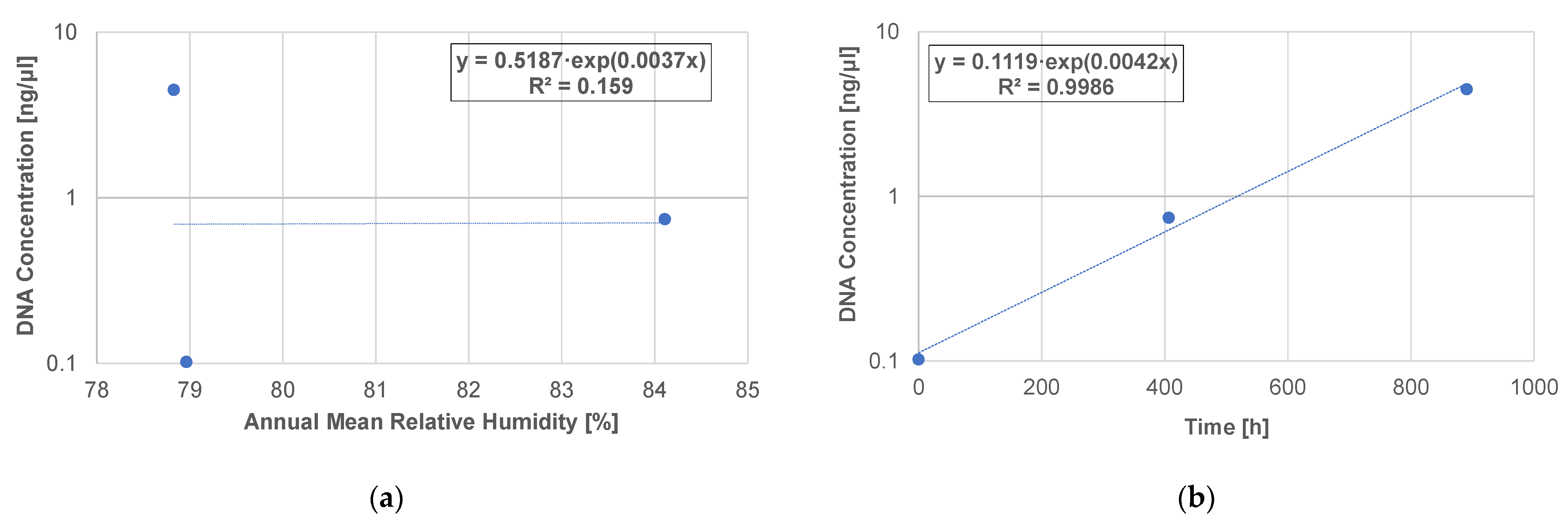
3.5. DNA Concentration and Relative Humidity of the Thatched Roof
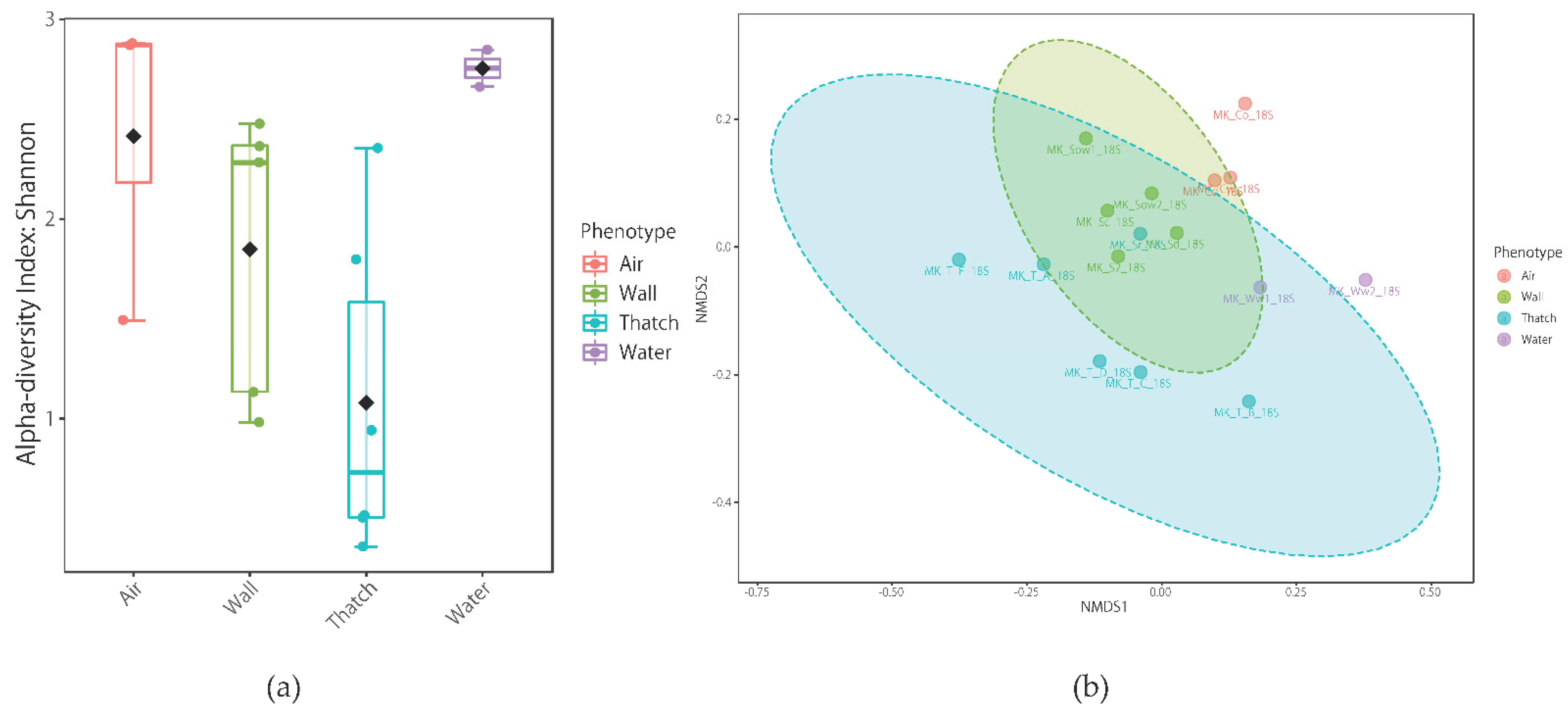
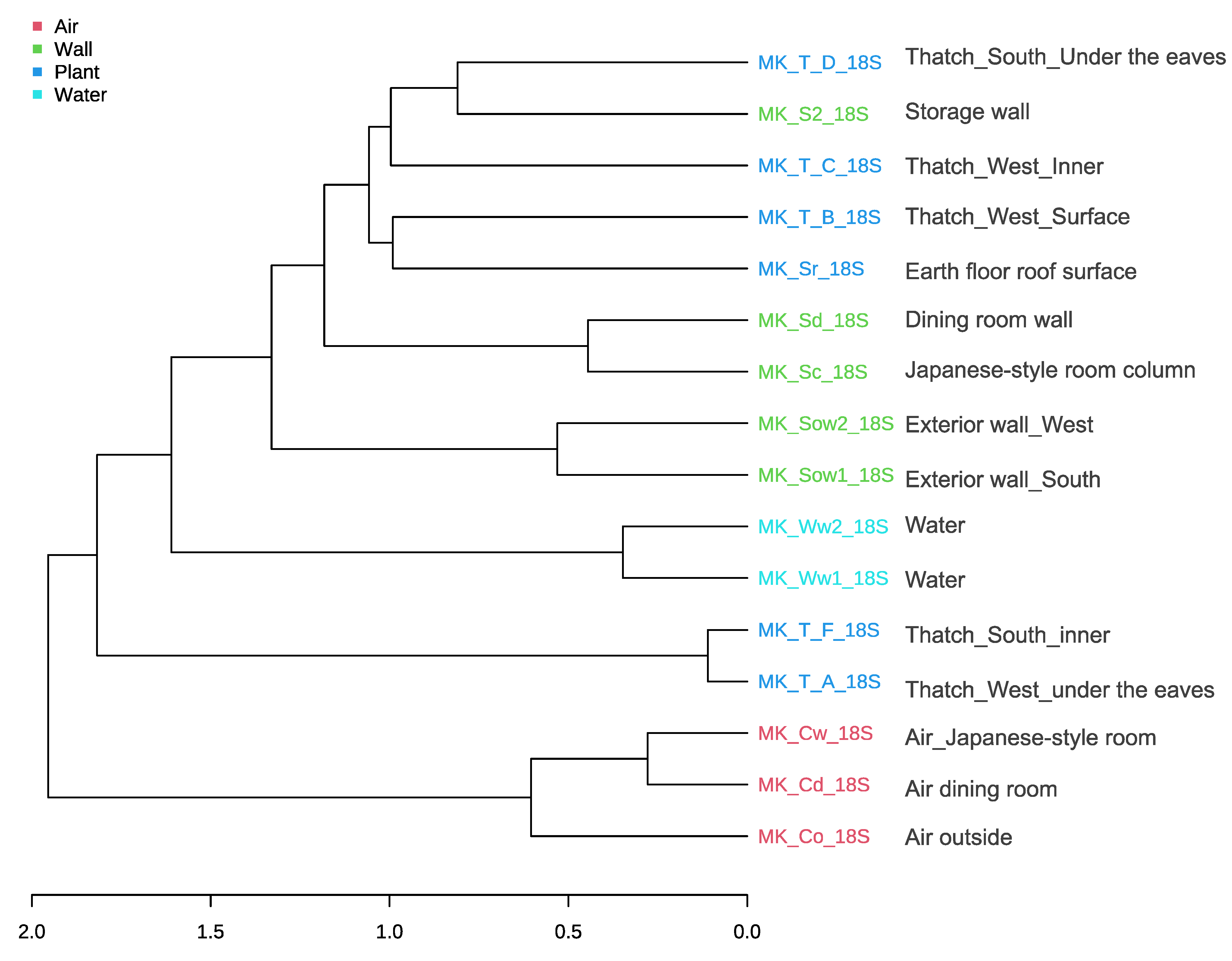
3.6. Sequence Results and Comparison of Microbial Communities
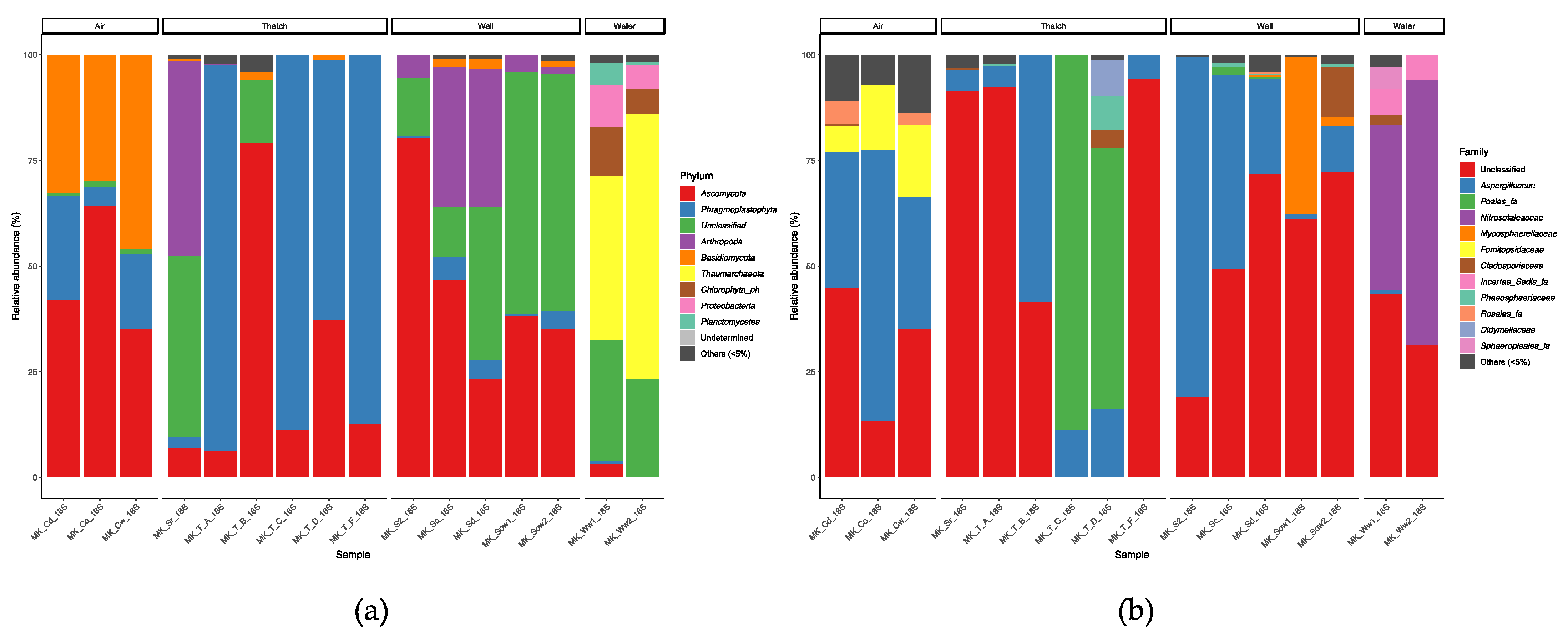
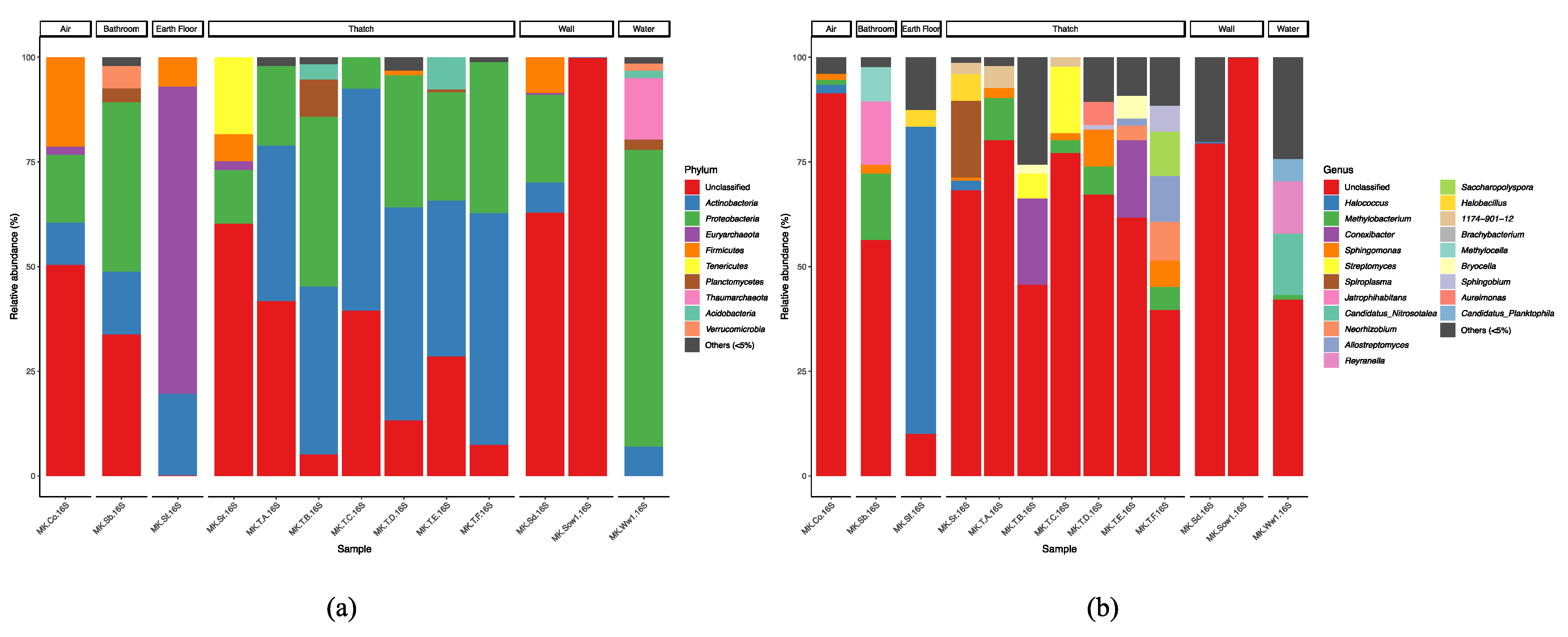
3.7. Taxonomic Composition and Biotic Interactions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Numerical Analysis of Surface Temperature/Humidity
- : gas constant of water vapour () [Pa·m3/kgK]
- : universal gas constant [Pa·m3/kmol·K]
- : molecular mass of water vapour [kg/kmol]
- : absolute temperature [K]
- : relative humidity [–]
- : water vapour pressure [Pa]
- : saturated water vapour pressure [Pa]
- : gas-phase moisture flux [kg/m2·s]
- : liquid-phase moisture flux [kg/m2·s]
- : total water flux [kg/m2·s]
- :density of liquid water [kg/m3]
- : volumetric moisture content [m3/m3]
- : water chemical potential [J/kg]
- : moisture conductivity with respect to water chemical potential gradient (=) [kg/ms(J/kg)]
- : moisture conductivity in the gas phase with respect to the water chemical potential gradient [kg/m s(J/kg)]
- : moisture conductivity in liquid-phase with respect to the moisture chemical potential gradient [kg/m·s·(J/kg)]
- : moisture conductivity with respect to temperature gradient (=) [kg/m·s·K]
- : moisture conductivity in the gas phase with respect to the temperature gradient [kg/m·s·K]
- : moisture conductivity in the liquid phase with respect to the temperature gradient [kg/m·s·K]
- : apparent volumetric specific heat of material which includes moisture [J/m3·K]
- : thermal conductivity [J/m·s·K]
- : heat of phase change from water vapour to liquid water [J/kg]
- : unit vector with downward vertical direction as positive
- : gravitational acceleration [m/s2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Earth Floor | Japanese-Style Room Wall | |
|---|---|---|
| Initial conditions | Spatial side: measured air temperature/humidity in earth floor Bottom of the ground: temperature 17 [°C] (annual mean outdoor air temperature) hydrochemical potential −1.0 [J/kg] | Indoor side: measured air temperature/ humidity in Japanese-style room Outdoor side: measured air temperature/humidity outdoors |
| Boundary conditions | Spatial side: Robin boundary condition Bottom of the ground: Dirichlet boundary condition | Both sides: Robin boundary condition |
| radiative heat transfer coefficient 4.4 [W/m2·K] convective heat transfer coefficient: earth floor space air 9.0 [w/m2·K], indoor air 4.9 [W/m2·K], outdoor air 18.6 [W/m2·K] measured air temperature/humidity The moisture transfer coefficient was determined from the convective heat transfer coefficient using the Lewis relationship. The Lewis number of 0.86 [58] was used. | ||
| Calculation area | depth of the ground 20 [m] | wall thickness 0.5 [m] |
- : combined heat transfer coefficient [W/m2·K] ()
- : radiative heat transfer coefficient [W/m2·K]
- : convective heat transfer coefficient [W/m2·K]
- : moisture transfer coefficient with respect to water vapour pressure gradient [kg/Pa·m2·s]
- : absolute temperature of air and wall surface [K]
- : water vapour pressure of air and wall surface [Pa]
- Subscripts:
- s: surface of the material
- i: i represents the room (r) or the outside air (o)
References
- Klepeis, N.E.; Nelson, W.C.; Ott, W.R.; Robinson, J.P.; Tsang, A.M.; Switzer, P.; Behar, J.V.; Hern, S.C.; Engelmann, W.H. The National Human Activity Pattern Survey (NHAPS): A resource for assessing exposure to environmental pollutants. J. Expo. Sci. Environ. Epidemiol. 2001, 11, 231–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, K. Comparison of a measured fungal index determined using fungal growth and a computed fungal index based on temperature and relative humidity. J. Soc. Indoor Environ. 2006, 9, 17–24. [Google Scholar]
- Sedlbauer, K. Prediction of mould growth by hygrothermal calculation. J. Therm. Envel. Build. Sci. 2002, 25, 321–336. [Google Scholar] [CrossRef]
- Hukka, A.; Viitanen, H.A. A mathematical model of mould growth on wooden material. Wood Sci. Technol. 1999, 33, 475–485. [Google Scholar] [CrossRef]
- Horve, P.F.; Lloyd, S.; Mhuireach, G.A.; Dietz, L.; Fretz, M.; MacCrone, G.; Van Den Wymelenberg, K.; Ishaq, S.L. Building upon current knowledge and techniques of indoor microbiology to construct the next era of theory into microorganisms, health, and the built environment. J. Expo. Sci. Environ. Epidemiol. 2020, 30, 219–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickman, M.; Gravesen, S.; Nordvall, S.L.; Pershagen, G.; Sundell, J. Indoor viable dust-bound microfungi in relation to resi-dential characteristics, living habits, and symptoms in atopic and control children. J. Allergy Clin. Immunol. 1992, 89, 752–759. [Google Scholar] [CrossRef]
- Van Strien, R.T.; Gehring, U.; Belanger, K.; Triche, E.; Gent, J.; Bracken, M.B.; Leaderer, B.P. The influence of air conditioning, humidity, temperature and other household characteristics on mite allergen concentrations in the northeastern United States. Allergy 2004, 59, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Harper, G.J. Airborne micro-organisms: Survival tests with four viruses. Epidemiol. Infect. 1961, 59, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, F.L.; Soergel, M.E.; Straube, D.C. Survival of airborne influenza virus: Effects of propagating host, relative humidity, and composition of spray fluids. Arch. Virol. 1976, 51, 263–273. [Google Scholar] [CrossRef]
- Lowen, A.C.; Mubareka, S.; Steel, J.; Palese, P. Influenza virus transmission is dependent on relative humidity and tem-perature. PLoS Pathog. 2007, 3, e151. [Google Scholar] [CrossRef]
- Stephens, B. What have we learned about the microbiomes of indoor environments? MSystems 2016, 1, e00083-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.I.; Bhangar, S.; Dannemiller, K.C.; Eisen, J.A.; Fierer, N.; Gilbert, J.A.; Green, J.L.; Marr, L.C.; Miller, S.L.; Siegel, J.A.; et al. Ten questions concerning the microbiomes of buildings. Build. Environ. 2016, 109, 224–234. [Google Scholar] [CrossRef] [Green Version]
- National Academies of Sciences, Engineering, and Medicine. Microbiomes of the Built Environment: A Research Agenda for Indoor Microbiology, Human Health, and Buildings; The National Academies Press: Washington, DC, USA, 2017. [Google Scholar]
- Weber, D.J.; Raasch, R.; Rutala, W.A. Nosocomial infections in the ICU: The growing importance of antibiotic-resistant patho-gens. Chest 1999, 115, 34S–41S. [Google Scholar] [CrossRef] [Green Version]
- Namkoong, H.; Kurashima, A.; Morimoto, K.; Hoshino, Y.; Hasegawa, N.; Ato, M.; Mitarai, S. Epidemiology of pulmonary nontuberculous mycobacterial disease. Emerg. Infect. Dis. 2016, 22, 1116–1117. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, K.E.; Demoor, T.; Rauch, M.; Faruqi, A.A.; Jang, S.; Johnson, C.C.; Boushey, H.A.; Zoratti, E.; Ownby, D.; Lukacs, N.W.; et al. House dust exposure mediates gut microbiome Lactobacillus enrichment and airway immune defense against allergens and virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 805–810. [Google Scholar] [CrossRef] [Green Version]
- Kirjavainen, P.V.; Karvonen, A.M.; Adams, R.I.; Täubel, M.; Roponen, M.; Tuoresmäki, P.; Loss, G.; Jayaprakash, B.; Depner, M.; Ege, M.J.; et al. Farm-like indoor microbiota in non-farm homes protects children from asthma development. Nat. Med. 2019, 25, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Lax, S.; Sangwan, N.; Smith, D.; Larsen, P.; Handley, K.M.; Richardson, M.; Guyton, K.; Krezalek, M.; Shogan, B.D.; Defazio, J.; et al. Bacterial colonization and succession in a newly opened hospital. Sci. Transl. Med. 2017, 9, eaah6500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kembel, S.W.; Jones, E.; Kline, J.; Northcutt, D.; Stenson, J.; Womack, A.M.; Bohannan, B.J.; Brown, G.Z.; Green, J.L. Architectural design influences the diversity and structure of the built environment microbiome. ISME J. 2012, 6, 1469–1479. [Google Scholar] [CrossRef] [Green Version]
- King, P.; Pham, L.K.; Waltz, S.; Sphar, D.; Yamamoto, R.T.; Conrad, D.; Taplitz, R.; Torriani, F.; Forsyth, R.A. Longitudinal metagenomic analysis of hospital air identifies clinically relevant microbes. PLoS ONE 2016, 11, e0160124. [Google Scholar]
- Park, D.-U.; Yeom, J.-K.; Lee, W.-J.; Lee, K.-M. Assessment of the levels of airborne bacteria, gram-negative bacteria, and fungi in hospital lobbies. Int. J. Environ. Res. Public Health 2013, 10, 541–555. [Google Scholar] [CrossRef]
- Hospodsky, D.; Yamamoto, N.; Nazaroff, W.W.; Miller, D.; Gorthala, S.; Peccia, J. Characterizing airborne fungal and bacterial concentrations and emission rates in six occupied children’s classrooms. Indoor Air 2015, 25, 641–652. [Google Scholar] [CrossRef]
- Qian, J.; Hospodsky, D.; Yamamoto, N.; Nazaroff, W.W.; Peccia, J. Size-resolved emission rates of airborne bacteria and fungi in an occupied classroom. Indoor Air 2012, 22, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, M.; Masueda, D.; Hokoi, S.; Matsushita, T. Airborne Algal growth on roofs of membrane-structured residences in cold area of Japan. J. Build. Phys. 2020, 45, 113–147. [Google Scholar] [CrossRef]
- Glass, E.M.; Dribinsky, Y.; Yilmaz, P.; Levin, H.; Van Pelt, R.; Wendel, D.; Wilke, A.; Eisen, J.A.; Huse, S.; Shipanova, A.; et al. MIxS-BE: A MIxS extension defining a minimum information standard for sequence data from the built environment. ISME J. 2014, 8, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Fujiyoshi, S.; Tanaka, D.; Maruyama, F. Transmission of airborne bacteria across built environments and its measurement standards: A review. Front. Microbiol. 2017, 8, 2336. [Google Scholar]
- Dannemiller, K.C.; Gent, J.F.; Leaderer, B.P.; Peccia, J. Influence of housing characteristics on bacterial and fungal communities in homes of asthmatic children. Indoor Air 2016, 26, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Lax, S.; Smith, D.P.; Hampton-Marcell, J.; Owens, S.M.; Handley, K.M.; Scott, N.M.; Gibbons, S.M.; Larsen, P.; Shogan, B.D.; Weiss, S.; et al. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Andersen, B.; Frisvad, J.C.; Søndergaard, I.; Rasmussen, I.S.; Larsen, L.S. Associations between fungal species and water-damaged building materials. Appl. Environ. Microbiol. 2011, 77, 4180–4188. [Google Scholar]
- Dannemiller, K.C.; Weschler, C.J.; Peccia, J. Fungal and bacterial growth in floor dust at elevated relative humidity levels. Indoor Air 2017, 27, 354–363. [Google Scholar] [CrossRef]
- Mastumoto, M. Simultaneous heat and moisture transfer in porous wall and analysis of internal condensation. In Energy of Conservation in Heating, Cooling, and Ventilating Buildings: Heat and Mass Transfer Techniques and Alternatives; Hoogendoorn, C.J., Afgan, N.H., Eds.; Hemisphere Publishing Corporation: Washington, DC, USA, 1978; pp. 45–58. [Google Scholar]
- Arai, S.; Kim, H.; Watanabe, T.; Tohya, M.; Suzuki, E.; Ishida-Kuroki, K.; Maruyama, F.; Murase, K.; Nakagawa, I.; Sekizaki, T. Assessment of pig saliva as a Streptococcus suis reservoir and potential source of infection on farms by use of a novel quantitative polymerase chain reaction assay. Am. J. Vet. Res. 2018, 79, 941–948. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Jansson, J.K.; Knight, R. The Earth Microbiome project: Successes and aspirations. BMC Biol. 2014, 12, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarimizu, K.; Fujiyoshi, S.; Kawai, M.; Norambuena-Subiabre, L.; Cascales, E.-K.; Rilling, J.-I.; Vilugrón, J.; Cameron, H.; Vergara, K.; Morón-López, J.; et al. Protocols for Monitoring harmful algal blooms for sustainable aquaculture and coastal fisheries in Chile. Int. J. Environ. Res. Public Health 2020, 17, 7642. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Team DCore. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2007; Available online: http://www.R-project.org (accessed on 18 May 2021).
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Yoshino, H.; Hasegawa, K.; Matsumoto, S.I. Passive cooling effect of traditional Japanese building’s features. Manag. Environ. Qual. 2007, 18, 578–590. [Google Scholar] [CrossRef]
- Szokolay, S. Introduction to Architectural Science, 3rd ed.; Routledge: Abingdon, UK; New York, NY, USA, 2012; pp. 35–37. [Google Scholar]
- Stres, B.; Danevčič, T.; Pal, L.; Fuka, M.M.; Resman, L.; Leskovec, S.; Hacin, J.; Stopar, D.; Mahne, I.; Mandic-Mulec, I. Influence of temperature and soil water content on bacterial, archaeal and denitrifying microbial communities in drained fen grassland soil microcosms. FEMS Microbiol. Ecol. 2008, 66, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Dunn, R.R.; Fierer, N.; Henley, J.B.; Leff, J.W.; Menninger, H.L. Home life: Factors structuring the bacterial diversity found within and between homes. PLoS ONE 2013, 8, e64133. [Google Scholar]
- Kembel, S.W.; Meadow, J.F.; O’Connor, T.K.; Mhuireach, G.; Northcutt, D.; Kline, J.; Moriyama, M.; Brown, G.Z.; Bohannan, B.J.; Green, J.L. Architectural design drives the biogeography of indoor bacterial communities. PLoS ONE 2014, 9, e87093. [Google Scholar] [CrossRef]
- Hsu, T.; Joice, R.; Vallarino, J.; Abu-Ali, G.; Hartmann, E.M.; Shafquat, A.; DuLong, C.; Baranowski, C.; Gevers, D.; Green, J.L.; et al. Urban transit system microbial communities differ by surface type and interaction with humans and the environment. Msystems 2016, 1, e00018-16. [Google Scholar] [CrossRef] [Green Version]
- Rutala, W.A.; Weber, D.J. Surface disinfection: Should we do it? J. Hosp. Infect. 2001, 48, S64–S68. [Google Scholar] [CrossRef]
- Vincent, M.; Hartemann, P.; Engels-Deutsch, M. Antimicrobial applications of copper. Int. J. Hyg. Environ. Health 2016, 219, 585–591. [Google Scholar] [CrossRef]
- Meadow, J.F.; Altrichter, A.E.; Kembel, S.W.; Kline, J.; Mhuireach, G.; Moriyama, M.; Northcutt, D.; O’Connor, T.K.; Womack, A.M.; Brown, G.Z.; et al. Indoor airborne bacterial communities are influenced by ventilation, occupancy, and outdoor air source. Indoor Air 2014, 24, 41–48. [Google Scholar] [CrossRef]
- Fahimipour, A.K.; Hartmann, E.M.; Siemens, A.; Kline, J.; Levin, D.A.; Wilson, H.; Betancourt-Román, C.M.; Brown, G.Z.; Fretz, M.; Northcutt, D.; et al. Daylight exposure modulates bacterial communities associated with household dust. Microbiome 2018, 6, 1–13. [Google Scholar] [CrossRef]
- Vandegrift, R.; Ishaq, S.L.; Kline, J.; Fahimipour, A.; Stenson, J.; Crowley, R.; Wilson, H.; Northcutt, D.; Hartmann, E.; Johnson-Shelton, D.; et al. Shut the front door: Seasonal patterns in window operation drive fungal and bacterial community dissimilarity between indoor and outdoor air. In Proceedings of the 15th Conference of the International Society of Indoor Air Quality and Climate, INDOOR AIR 2018, Philadelphia, PA, USA, 22–27 July 2018. [Google Scholar]
- Gibbons, S.M. The built environment is a microbial wasteland. MSystems 2016, 1, e00033-16. [Google Scholar] [CrossRef] [Green Version]
- Otter, J.A.; French, G.L. Survival of nosocomial bacteria and spores on surfaces and inactivation by hydrogen peroxide vapor. J. Clin. Microbiol. 2009, 47, 205–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, A.; Schwebke, I.; Kampf, G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect. Dis. 2006, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.M.; Eng, R.H.; Padberg, F.T., Jr. Survival of nosocomial pathogenic bacteria at ambient temperature. J. Med. 1996, 27, 293–302. [Google Scholar] [PubMed]
- Gil, F.; Lagos-Moraga, S.; Calderón-Romero, P.; Pizarro-Guajardo, M.; Paredes-Sabja, D. Updates on Clostridium difficile spore biology. Anaerobe 2017, 45, 3–9. [Google Scholar] [CrossRef]
- Ogura, D.; Matsushita, T.; Matsumoto, M. Analysis of heat and moisture behavior in underground space by quasilinearized method—Accuracy range for outdoor climate variation from the reference. In Proceedings of the Building Simulation ’99: Sixth International IBPSA Conference, Kyoto, Japan, 13–15 September 1999; pp. 755–762. [Google Scholar]
- Takada, S.; Uno, Y.; Ota, M. Study on hygrothermal behavior of mud wall—Measurement of sorption isotherm and analysis onindoor temperature and humidity. Summ. AIJ Kinki Chapter Res. Meet. Environ. Eng. 2013, 53, 177–180. (In Japanese) [Google Scholar]
- Eckert, E.R.; Drake, R.M., Jr. Analysis of Heat and Mass Transfer; McGraw Hill: New York, NY, USA, 1972; p. 733. [Google Scholar]













| (a) | ||||
| Annual Mean | Annual Highest | Annual Lowest | Annual Range | |
| outdoor air | 14.1 | 37.3 | −1.5 | 38.8 |
| earth floor space | 15.4 | 33.3 | 2.0 | 31.3 |
| space under the ceiling | 17.3 | 34.0 | 2.4 | 31.6 |
| storage | 18.9 | 34.9 | 5.1 | 29.8 |
| dining room | 21.3 | 34.2 | 6.4 | 27.8 |
| Japanese-style room | 21.7 | 39.6 | 6.4 | 33.2 |
| (b) | ||||
| Annual Mean | Annual Highest | Annual Lowest | Annual Range | |
| outdoor air | 84.4 | 100 | 17.3 | 82.7 |
| earth floor space | 78.0 | 94.9 | 27.3 | 67.6 |
| space under the ceiling | 69.3 | 89.8 | 27.7 | 62.2 |
| storage | 68.8 | 88.9 | 29.5 | 59.4 |
| dining room | 60.6 | 90.5 | 24.6 | 65.9 |
| Japanese-style room | 60.7 | 91.8 | 18.5 | 73.4 |
| Outdoor Air | Roof Surface 1 | Roof Inner 250 mm | |
|---|---|---|---|
| Days | 73 | 185 | 0 |
| Total time | 406.5 | 890.5 | 0 |
| Number of data available | 17,568 | 16,819 | 17,567 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokubo, M.; Fujiyoshi, S.; Ogura, D.; Nakajima, M.; Fujieda, A.; Noda, J.; Maruyama, F. Relationship between the Microbiome and Indoor Temperature/Humidity in a Traditional Japanese House with a Thatched Roof in Kyoto, Japan. Diversity 2021, 13, 475. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100475
Kokubo M, Fujiyoshi S, Ogura D, Nakajima M, Fujieda A, Noda J, Maruyama F. Relationship between the Microbiome and Indoor Temperature/Humidity in a Traditional Japanese House with a Thatched Roof in Kyoto, Japan. Diversity. 2021; 13(10):475. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100475
Chicago/Turabian StyleKokubo, Makoto, So Fujiyoshi, Daisuke Ogura, Makiko Nakajima, Ayako Fujieda, Jun Noda, and Fumito Maruyama. 2021. "Relationship between the Microbiome and Indoor Temperature/Humidity in a Traditional Japanese House with a Thatched Roof in Kyoto, Japan" Diversity 13, no. 10: 475. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100475