
Lessons of 15,000 Years of Human–Wildlife Interaction for Conservation in Patagonia in the 21st Century

1
Instituto de Investigaciones en Biodiversidad y Medioambiente (INIBIOMA), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)-Universidad Nacional del Comahue, Centro de Ecología Aplicada del Neuquén, Ruta 61, km 3, Junín de los Andes, Neuquén 8371, Argentina
2
Wildlife Conservation Society, Argentina Program, Curruhue 395, Junín de los Andes, Neuquén 8371, Argentina
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(12), 633; https://0-doi-org.brum.beds.ac.uk/10.3390/d13120633
Submission received: 21 February 2021
/
Revised: 15 November 2021
/
Accepted: 20 November 2021
/
Published: 2 December 2021
(This article belongs to the Special Issue Humans and Wild Animals: Interactions in Deep Time, Recent History, and Now)
Abstract
:Humans arrived in the Patagonia region of southern South America in the late Pleistocene period, ca. 15,000 years ago. A few centuries later, during a period of rapid warming, the megafauna went extinct in Patagonia, as well as some smaller species, like the guanaco (Lama guanicoe), in the southern portion of the region. As in other regions, extinctions probably occurred due to a combination of effects of climate and direct and indirect impacts of humans on wildlife communities. We reviewed recent archeological and genetic-based discoveries about numbers and distributions of humans and wildlife and their early interactions and used them to draw lessons for current debates among managers and scientists. Recent discoveries, for example, help us understand (1) the population limitation mechanisms and other interactions involving guanacos, livestock, forage, predators, and scavengers; (2) the magnitude of wildlife movements and the need for landscape-level planning for conservation; (3) the importance of indirect effects of human activities on wildlife communities; and (4) the compounded effects of human activities and climate change on wildlife. We believe these lessons drawn from deep time and recent history can help define new priorities for research and management and inform our conservation vision for the 21st century, a period when dramatic climate change impacts will add challenges to a region subject to a century of overgrazing and other anthropogenic pressures.
Keywords:
Patagonia; Pleistocene; Anthropocene; conservation; human–wildlife coexistence; rewilding; guanaco1. Introduction
For millions of years, geological and evolutionary forces, such as the breaking up of Gondwanaland, the rising of the Andes mountains, and the emergence of the Panamanian land bridge, together with changes in climate, governed the evolution of life on the South American continent and the development of faunal communities. Then, approximately 15,000 years ago, Homo sapiens arrived [1] and became an additional force influencing wildlife species and communities. With the exception of Antarctica, the human arrival in South America is the most recent encounter between humans and the wildlife assemblages of an entire continent.
Paleontological, archeological, and paleobiological studies have provided essential insights to guide conservation science and ecosystem management and restoration around the world for decades [2,3,4,5]. Historical perspectives are particularly necessary to understand and conserve long-term processes, such as ecological succession, migration, adaptation, microevolution, and extinction, especially in the context of ongoing and future global challenges to biodiversity, such as climate change [6,7]. Examples of key paleoecological insights for conservation planning include understanding how bottom-up processes of climate change can be distinguished from top-down processes, including anthropogenic impacts on biodiversity [8,9], long-term biotic responses that depend on complex and non-linear processes with thresholds and feedbacks that lead to unexpected surprises, and the effects of global warming in the late Pleistocene era (ca 13,000 to 11,500 years ago) at similar rates and magnitudes to the warming projected for the 21st century [7].
Throughout most of the world, great numbers of large-bodied terrestrial species became extinct during the waning days of the Pleistocene period (50–10,000 years ago) [10]. The primary causes of these extinctions have been debated for decades and hypotheses proposed range from human hunting to climate and habitat changes, disease, and a comet impact [9,11,12,13]. In the Americas, the late Pleistocene extinction of megafauna was severe and occurred at the same time of significant climate changes and human colonization, so the debate on the causes of extinction has been more controversial and persistent than in other continents. While the role of humans versus climate and other causes has been debated for almost 50 years in North America [14,15,16,17], few researchers, especially among archeologists, considered that humans had played a major role in South America until relatively recently (reviewed by [18]). This view has shifted in recent years, mainly due to paleoecological and paleontological studies increasingly assigning a possible major role to humans and concluding that more than one factor may be responsible for most extinctions [6,11,18].
In the last 20 years, we have learned much about interactions between early humans and wildlife in the southernmost Patagonia region of South America, the ecology of Patagonian wildlife, and current impacts of and interactions between humans and wildlife in Patagonia, where human and zooarcheological records are abundant and well preserved (for example: [19,20]). In particular, a wealth of recent research by paleontologists and archeologists published in the last 5 years has given us a better idea of how humans and wildlife coexisted in the region during the late Pleistocene period and throughout the Holocene period (for example: [1,18,19,20,21,22,23,24,25,26]). We believe this new understanding of the history of human and wildlife coexistence can help inform current and future efforts to promote wildlife conservation and coexistence with humans in Patagonia.
In this article, we will integrate the research from these disparate fields to examine lessons of the history of 15,000 years of coexistence between humans and wildlife, relevant to conservation issues we are facing as conservation practitioners in Patagonia in the 21st century. We will focus our analyses on interactions involving guanacos (Lama guanicoe), 80–120 kg camelids well adapted to arid and semiarid environments [27], because they became the dominant ungulate of Patagonian grasslands after the late Pleistocene extinction of the megafauna. We will analyze four lessons that we gleaned from our review of recent paleontological and ecological research in Patagonia: (1) how understanding the shifting top-down and bottom-up population limitation of guanacos can inform contentious guanaco management and grassland conservation decisions today; (2) how understanding the role of large-scale wildlife movements can aid in the design of strategies for coexistence between livestock and wildlife; (3) how understanding the possible indirect effects of humans on the wildlife community since humans arrived can inform conservation targets, goals, and strategies; and (4) the cautionary tale of human impacts on wildlife during a scenario of climate change. Finally, we will discuss how these lessons can guide the redefining of rewilding as a conservation vision that more thoroughly recognizes human–wildlife coexistence in Patagonia.
2. Human and Wildlife History in Patagonia from the Late Pleistocene Era to the Present
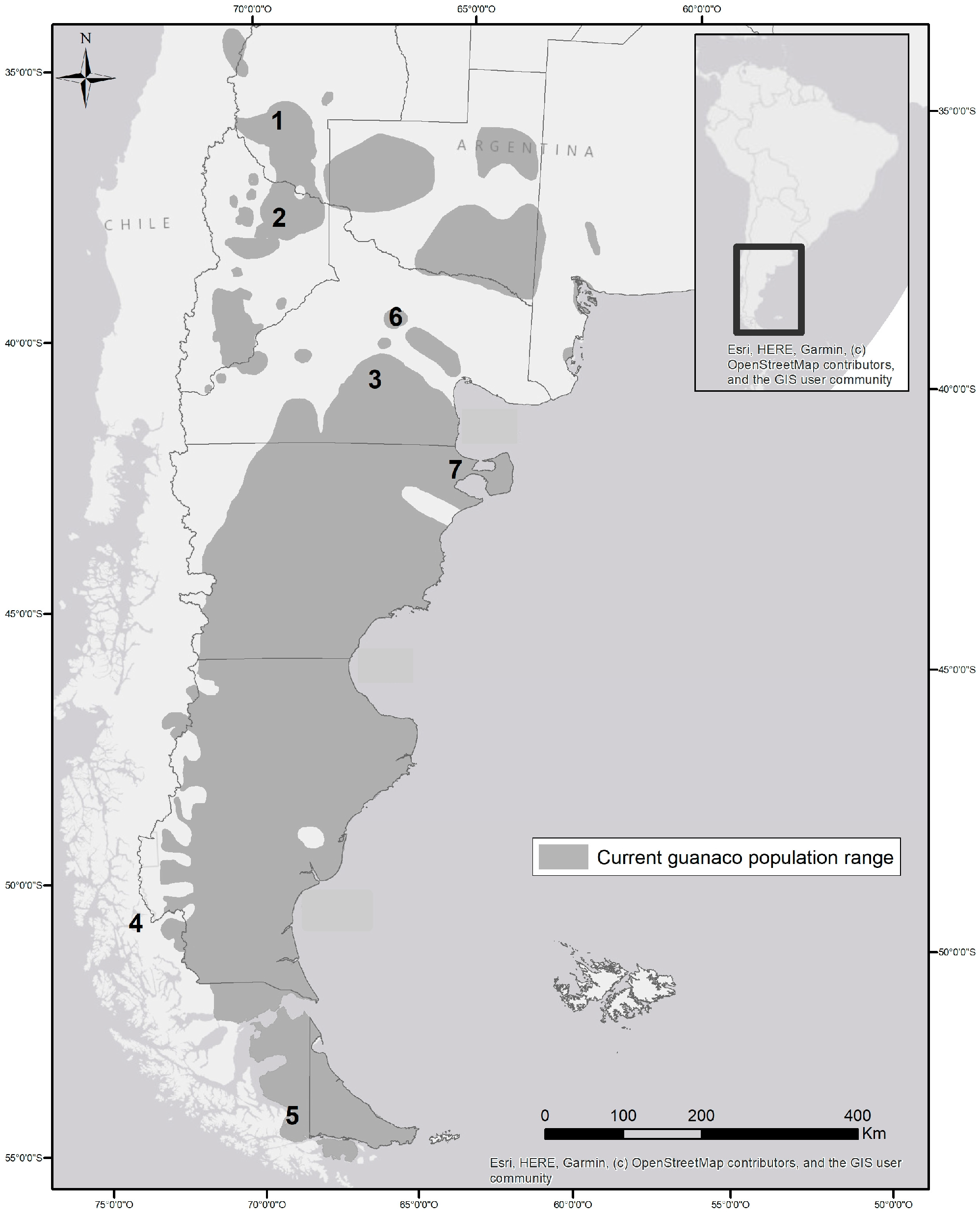
Patagonia is the southernmost portion of the South American continent, about 90% of which is in Argentina and 10% in Chile, covering about one million square kilometers and stretching from approximately 35°24′ S to 55°15′ S, what is now the island of Tierra del Fuego (Figure 1). The last continental region to be colonized by Homo sapiens, recent analyses of radiocarbon dates from archeological sites and mtDNA from ancient and living people have estimated an arrival in Patagonia between 14,500 and 17,000 years ago, very quickly after reaching the continent [1,20]. The fauna these humans encountered included large-bodied herbivores, such as ground sloths (Mylodon darwini), equids (Hippidion saldiasi), and camelids (Lama gracilis and L. guanicoe), and large carnivores including the puma (Puma concolor), a giant jaguar (Panthera onca mesembrina), a sabretooth (Smilodon populator), a bear (Arctotherium tarijense), and a canid (Dusicyon avus) [19,28,29]. Proboscideans did not reach Patagonia, except in the northernmost fringes, and no bovids dispersed into South America, so large-bodied ecosystem engineers were absent and the Patagonian steppe evolved under low herbivory pressure [21]. Humans represented a new predator in the Patagonian ecosystem in the late Pleistocene period, capable of hunting even the largest-bodied herbivores [18]. They appeared to have relied more on the smaller and more abundant guanacos and the even smaller camelid, Lama gracilis, than on megafauna [29]. Guanacos descended from North American camelids that moved into South America 2–3 million years ago, and the genus Lama arose in the early to mid-Pleistocene period, while the guanaco was in Patagonia by at least about 13,600 years ago [30].
Extinctions of large-bodied animals in Patagonia have been more recent and were more devastating than in most regions other than islands, and affected smaller species, including the guanaco [11,19,29]. The guanaco was extirpated from the region by around 12,300 years ago and was later replaced by a genetically distinct northern population that moved in by around 10,500 years ago, which is ancestral to modern guanacos [19]. By the early to middle Holocene period, from 9500–4000 years ago, the guanaco had become the dominant herbivore on the Patagonian steppe. The largest predators were gone, leaving only Dusicyon avus, the culpeo (Lycalopex culpaeus), the puma, and humans ([19]: supplementary Table S2).
Human population growth and distribution appears to have tracked that of guanacos [20,24]. Initially moving in along the Atlantic and Pacific coasts, human population growth at the regional level was slow and steady from about 15,000 to 7000 years ago, then rapidly increased from around 7000 to 1000 years ago, when it stabilized. Like the guanaco, the distribution of this growing human population was not uniform across space and time. After having spread inland prior to 12,000 years ago, human occupation was greatly reduced in the interior after the megafauna and guanaco extinctions and increased again in the mid-Holocene period as guanacos recolonized from the north and their abundance and distribution increased. By the late Holocene period human population density has been estimated at between 5 and 20 individuals/100 km2, thirty times that of the late Pleistocene period [20].
Thus, when Europeans arrived to the continent about 400 years ago, the faunal community they encountered was less than 8000 years old, and humans were a key component of the ecosystem. Both pumas and humans relied principally on guanacos and, to a lesser extent, rheas (Rhea pennata), armadillos (Family Dasypodidae), and huemul deer (Hippocamelus bisulcus) in the ecotone habitats where these species were available [24,28,29]. Although Magellan reported the discovery of the Patagonia region in 1520, people of European descent did not occupy the interior of Patagonia until the late 1800s. Nevertheless, their influence on biodiversity and human culture was strong during this lapse of more than 300 years, as they conquered and colonized what came to be Argentina to the north, as well as the coasts of Chile and Argentine Patagonia.
When western European colonizers first arrived, native populations in the Patagonian region were nomadic hunter–gatherers. Some in the northern and western (Chile) parts of Patagonia quickly adopted more sedentary lifestyles based on livestock they obtained through trade or raiding of livestock of Europeans in nearby areas, losing their dependence on wildlife for meat, shelter, and clothing. The Tehuelche people in southern Patagonia, however, continued their nomadic, hunter–gatherer lifestyle, following movements of the guanaco [35]. Nevertheless, their culture was greatly influenced by the adoption of the horse, which became a vital means of transportation, and was used for hunting, meat, and ceremonial purposes. In addition, feral cattle and other feral livestock provided new prey species. By the time Europeans entered the area, the Tehuelches had already greatly decreased in number, ravished by diseases brought through contact with Europeans and military excursions [36].
The Second Desert Campaign by the Argentine army in 1878 sought to exterminate the remaining tribes and extend Argentine national territory. By 1899 over 20,000 people were killed and many more moved away into urban areas [37], and the Tehuelche lifestyle based on guanaco hunting was extirpated. The vast lands taken over by the Argentine state were distributed among Argentines of European descent and new European immigrants, ushering in a new era of human–wildlife coexistence as these settlers brought in large herds of sheep and introduced wildlife species from their homelands.
Since that time, over the last 100 years, livestock (sheep, and to a lesser extent, cattle and goats), and wild invasive species (first, feral horses and cattle, then European hare (Lepus eurapeus), rabbit (Oryctolagus cuniculus), red deer (Cervus elaphus), and wild swine, (Sus scrofa)) have drastically reshaped Patagonian steppe ecosystems. Grazing by sheep led to severe or very severe desertification of 31% of the steppe [38] and direct competition with native herbivores [39,40]. In combination with unsustainable commercial hunting and indirect impacts, such as disease and apparent competition, populations of guanacos, rheas, and caviomorph rodents in the steppe and huemul in the forest–steppe ecotone collapsed [39,41,42,43,44,45]. Guanacos declined to 2–9 percent of their pre-conquest numbers and were pushed to the more arid and isolated areas [46]. To protect sheep, persecution of pumas through hunting and the use of poison led to their extirpation and that of Andean condors (Vultur gryphus), the largest avian scavenger, from most of the steppe by the 1950s [45,47].
During the last few decades, the number of sheep in Patagonia has declined from its peak of 50 million to approximately 8 million due to decreased productivity caused by overgrazing [48] and low wool prices, though in some areas sheep were replaced by cattle and goats [49]. With the decline in sheep numbers, and the concomitant decline in the rural human population, guanacos, pumas, and other wildlife began to recover in some areas, but some invasive wildlife, such as the red deer, also expanded their distributions. The guanaco recovery has been most dramatic in southern Patagonia, where a large proportion of ranches have been abandoned for two decades or more [50]. The guanaco population range today is continuous in southern Patagonia and still highly fragmented in northern Patagonia, though some high-density populations occur in both areas (Figure 1, Baldi et al.’s unpublished manuscript, [51]). Similarly, pumas have recovered most of their early 20th century distribution in the steppe [52]. Some iconic wildlife species, such as Andean condors, rheas, and Andean cats (Leopardus jacobita), however, have not recovered or have continued their declines in abundance and/or distribution, and the causes for these trends are not well understood [42,47,53].
3. Lesson 1: Shifting Outcomes between Top-Down and Bottom-Up Control among Guanacos, Forage, and Predators
A pervasive debate has dominated the community of Patagonian ranchers, scientists, and conservationists over the last three decades, as guanaco populations have begun to recover in parts of Patagonia despite continued degradation of rangelands. Some ranchers and scientists argue that in addition to high and constant livestock numbers, guanaco recovery is contributing to this ongoing degradation of vegetation in the Patagonian steppe. They hypothesize that the removal of pumas, their top predator in previous decades, and inefficient density-dependent mechanisms of guanacos render them likely to exceed carrying capacity, leading to further range degradation and possible massive mortality events [49,54]. Therefore, they advocate culling and harvesting of locally abundant guanaco populations to protect and improve range condition. Other scientists argue that plant grazing resistance traits support the hypothesis of bottom-up limitation in the Patagonian steppe [55,56] and that guanaco territoriality and resource-defense mechanisms are sufficient to limit populations below numbers predicted by forage availability [57,58]. These authors argue that guanacos are used as scapegoats for degradation caused by livestock numbers that have continued to be elevated locally and livestock grazing heterogeneity that has continued depleting plant resources. Ultimately, the debate is centered, in part, around the relative importance of top-down versus bottom-up processes of population limitation and the evolutionary history of herbivory, predation, and population regulation in Patagonia [21,56,59,60].
Guanaco adaptations to highly variable arid and semiarid environments include morphological, physiological, and behavioral traits, as summarized by Oliva et al. [49]. They have soft foot pads under deeply cleft toes [61], which may reduce trampling [62], and their mouths have split upper lips that draw in food instead of cutting with a deep bite as sheep do. Guanaco and sheep diets overlap but not completely, as guanacos are more generalists, consuming a broad range of grasses and dicotyledonous plants [40,63], and can graze and browse tough, low-quality food during dry periods, reducing intake and decreasing transit time through the intestinal tract to enhance microbial attack of fibrous vegetation [64,65]. Guanaco birth rates can decline dramatically during drought periods [66], leaving more forage available for adults to survive. Finally, guanacos also are habitat generalists in comparison to sheep [67,68], which, in combination with their territorial behavior and resource defense polygyny [57,69], may have allowed them to persist in marginal habitats during periods of intense interspecific competition, both in the late Pleistocene period and the recent past.
In this section, we review the archeological and paleobiological evidence for the opposing hypotheses proposed on guanaco population limitation and density dependence, analyze the prevalence of top-down and bottom-up processes driving guanaco dynamics, and assess the possible impacts of guanaco dynamics on habitat degradation. The recent insights provided by genetics studies into the demographic trajectory of both guanacos and humans over the last 15,000 years provide crucial lessons to guide the recovery of guanacos where they are still in low numbers, as well as the management of abundant guanacos in areas of conflict with livestock.
The initial extinction of the guanaco, along with the Patagonian megafauna, around 12,000 years ago probably resulted from combined effects of human hunting pressure and a period of rapid warming and possibly a retraction of grasslands [19,70]. This extinction event could be evidence of a top-down effect on guanacos, which is surprising given that human density at the end of the Pleistocene period was very low [20]. However, in the late Pleistocene period, guanacos shared the steppe with 15 other large and mega herbivores and five non-human large predators [19,21]. Thus, even if guanacos were abundant enough to be a preferred prey of humans, it is likely that guanaco population density was not high, a demographic condition that increases the probability of being subject to top-down control by predation [71]. In addition, humans were a new and very resourceful predator that relatively suddenly appeared on the scene, adding to the predation pressure of the other large predators. The spatial extent of this extinction of guanacos and megaherbivores was so widespread in the steppe that it led to human abandonment of vast portions of Patagonia‘s interior for a period of up to 2000 years, apparently due to the absence of prey [20,24,72].
Thus, when guanacos began to recolonize the steppe from northern populations, they encountered a habitat with few or no humans. In this ecosystem, 7500 years ago, guanaco populations expanded and increased in numbers for about 6000 years, stabilizing at a high density level 1000 years ago [24] and remaining at that level until the arrival of Europeans. During the Holocene period, as humans increased in numbers, recolonized the interior of Patagonia, and began consuming guanacos as their dominant food source once again [22,23], they did not have the demographic impact they possibly had in the late Pleistocene and early Holocene periods. During the Holocene period, human populations closely tracked the regional increasing population trend of their main prey and stabilized at the same time [20,22]. This demographic relationship was very different from that of guanacos and humans in the late Pleistocene period and usually is indicative of bottom-up regulation [73], as the amount of forage limits the number of herbivores and the number of herbivores limits abundance of predators.
The general lack of demographic impact of hunting by humans on abundant guanaco populations during the late Holocene period, however, may have occurred because climate and habitat conditions remained favorable in most of Patagonia. The exception to this pattern, and possible confirmation that human hunting in combination with climate was a driving force in guanaco dynamics, came recently from findings in northwest Patagonia that contrasted with regional trends described by Moscardi et al. [24]. Abbona et al. [26], also using mDNA analysis, concluded that after the guanaco population in northwest Patagonia had reached the high and seemingly stable level reached in the rest of Patagonia, a marked decline occurred ca. 2500 YBP. Abbona et al. [26] concluded that the decline was likely caused by the temporal overlap between hunting by a large human population and a period of increased aridity that did not occur in the rest of Patagonia and resulted in declining forage resources. The guanaco decline in northwest Patagonia was followed by increased human sedentarism, diet diversification, and subsistence farming around 2000 YBP [25]. Conversely, in the rest of Patagonia, where guanaco numbers remained high, humans continued to be hunter–gatherers and rely on guanacos as their main food source until the arrival of Europeans [24].
Different outcomes of the interaction between human predators and guanacos between the late Pleistocene and the Holocene periods match predictions of predator–prey theory, as well as extensive empirical evidence, because the limiting role of predators, in addition to other factors, depends on the density of the prey in relation to the carrying capacity of the habitat, primarily in simple systems with one main prey and one main predator [71,73]. Predators are able to exert a limiting effect only below a certain threshold density of prey, beyond which prey numbers are limited, often by other mechanisms such as density dependence and food availability. Different impacts of predation on ungulate populations depending on ungulate density levels have been reported for various species, with predation being significant at low but not at high prey density [73,74,75,76]. In the late Pleistocene period, guanaco densities were likely low, and when guanacos recolonized the Patagonian interior in the early Holocene period with high forage availability and without competition from other large herbivores, their high intrinsic rate of population increase may have helped them reach densities above the threshold for limitation by predators before human hunters and pumas were able to rebuild their populations. So while human numbers, after recolonizing the interior, were also able to increase rapidly thanks to increasing guanaco numbers [20], their hunting methods did not appear to allow them to exert a significant top-down limitation again. This is evidenced by the relative stability of their growth curves for around 1000 years before the arrival of Europeans and their livestock and diseases [20,22]. Predation only played a role in helping limit guanacos at high density in the late Holocene in northwest Patagonia [26], but bottom-up limitation was also operating in this area due to the drying climate.
If guanacos had weak density-dependent mechanisms, as Oliva et al. [54] argued, we would expect them to have overshot carrying capacity, have overgrazed vegetation, and experienced population retractions in the late Holocene period in various areas of Patagonia. However, widespread overgrazing and retractions did not occur. Demographic reconstructions show a period of fast increase [24,26,77] and a monotonic approach to a high and stable density level [24], consistent with population dynamics that at high density are limited by food resources and effective density-dependent mechanisms like intraspecific competition and social interactions [73,78].
Finally, during two periods in the Anthropocene age, human–guanaco interactions had two other turning points that are relevant to understand their intertwined dynamics and the implications for future management. Guanaco numbers in Patagonia at the time of the arrival of Europeans had been estimated at around 10 million [79,80]. This number was recently revised to a minimum range of 12–15 million using mitochondrial DNA-based estimations [22,24]. From this level, numbers dropped dramatically until the late 20th century. During this decline, as in their late Pleistocene extinction, guanacos were sharing the range with a large number of competitors, this time domestic sheep rather than a diverse native megafauna. Both interspecific competition and hunting by humans (through direct persecution by ranchers) once again played key roles in the population collapse. Then, in the last 20–30 years as both of these forces were relaxed, due to the sheep decline and hunting restrictions, guanacos have been recovering in Patagonia, though scales of spatial recolonization and recovery in numbers have been more extensive in southern Patagonia [Baldi et al., unpublished manuscript, Figure 1].
Observations throughout Patagonia where sheep were removed and guanacos have recovered, remained at high densities for one to three decades, and been allowed to range freely, show no sign of vegetation overgrazing beyond levels occurring at sheep removal, or population declines [45,57,81,82]. Thus, there is extensive evidence of guanaco persistence at high density without range degradation and resulting population collapses both in the 1000-year period before European arrival and in the present. The only documented case of range degradation possibly caused by guanacos, and a resulting die off, is a population where movements and recruitment resulting from those movements were artificially driven by humans, preventing natural density dependent regulation (see Lesson 2). Conversely, range degradation has continued throughout Patagonia wherever livestock grazing has continued to exceed forage availability in recent decades [54,58]. However, possibly because guanacos are better adapted than sheep to harsh habitat conditions, they have been able to recolonize some degraded areas and coexist, at least in the short term (last decade), with sheep herds in southern Patagonia [54,58].
In summary, the newly available genetic and archeological evidence suggests that the prevalence of top-down and bottom-up processes driving guanaco dynamics have shifted through time in Patagonia, depending primarily on the relative densities of competitors and climate. Before the arrival of Europeans and their livestock, and in numerous populations that have recovered, guanacos have occurred at high densities, without evidence of overgrazing and die-offs, thus displaying effective rather than ineffective density dependence. For conservation in the 21st century, this leads us to the conclusion that guanaco population dynamics, and specifically their density-dependence mechanisms, are not likely responsible for the degradation of the range observed in recent decades, in partial agreement with the arguments of Marino et al. [58]. The current abundance of guanacos in some parts of Patagonia is not due to lack of limitation by predation or deficient population regulation mechanisms. However, even if guanacos did not cause the widespread range degradation, mixed management of abundant sheep and guanaco herds, where they co-occur, may be necessary to facilitate range recovery, as Oliva et al. [49,54] have proposed, especially in the face of projected plant cover and productivity declines for arid Patagonia in coming decades as a result of climate change [J. Gaitan et al., unpublished data].
On the other hand, the evidence summarized suggests simultaneous top-down and bottom-up control of guanacos did indeed occur at a regional scale ca. 12,000 years ago and again ca. 2500 years ago in northwestern Patagonia, both times associated with warmer and drier conditions, likely with dramatic consequences for ecosystems and humans. In areas where guanacos are still in low numbers, they may indeed be limited by pumas supplemented by livestock and invasive species, as was the case when guanacos shared the steppe with numerous other herbivores. We have found that guanacos, in areas where intense competition with livestock and persecution by humans ceased, may escape the limiting effect of puma predation when they reach a threshold density of approximately 8/km2 [45,52].
4. Lesson 2: Large-Scale Movements by Wildlife
Seasonal movements help large herbivores sustain high densities [83] and movements (including seasonal, round-trip displacement, and also population dispersal) are particularly important in semi-arid environments such as the Patagonian steppe, with high year-to-year and spatial variability in precipitation and plant productivity. Nevertheless, guanacos are believed to be mostly sedentary by many ranchers and huemul in the eastern Andean foothills are rarely seen in low elevation, highly productive valleys, and these patterns of limited movements were endorsed by early research that described what may have been mostly recent phenomena caused by livestock husbandry monopolizing the best habitats [84]. In the case of huemul, their near extinction and lack of recovery after direct persecution declined in Argentina in recent years is likely the result primarily of the loss of their prime winter range to livestock and human occupation of the Andean foothills [84]. Recent zooarcheological data [85,86] have implicitly endorsed this crucial role for huemul seasonal movements in the Andes.
The recolonization of southern Patagonia by guanacos from northern Patagonia 12,000 years ago, as evidenced by genetic data [19], indicates a high capacity for long-distance movements and the importance of maintaining connectivity to prevent extinction of small, isolated guanaco populations. Even in recent times, with barriers that include high livestock densities, fences, and persecution by ranchers, according to genetic evidence, guanacos appear to have maintained dispersal over long distances [51].
Several recent archeological studies have provided evidence of seasonal movements by people following guanacos (reviewed by [23,87]), and early historical accounts documented Tehuelche people following guanaco movements [35], suggesting that these movements were once common throughout the steppe. Seasonal movement of people following migratory guanacos, and prey and patch choice for hunting, may have allowed sustainable use of guanacos by abundant mid-Holocene hunters in northern Patagonia [87] and elsewhere in the region.
Widespread habitat exclusion by livestock and fencing of Patagonia since the introduction of sheep [39] are likely responsible for most of the sedentary or limited guanaco movements observed today. Guanaco young sustain high mortality on extensive fencing systems [66]. Nevertheless, some or all individual guanacos recently tracked using radiotelemetry in five of seven Patagonian sites had seasonal migrations, even in areas with highly fragmented ranges (Figure 1, [32,33,34]). Movements tracking forage productivity were most extensive in the two rare sites with few or no fences (up to 85 km annually in one of them, with large year to year variation [31], Baldi et al., unpubl. data, Figure 1). Seasonal movements were absent at the two sites studied that appeared to have highest sheep and fence densities in surrounding lands (sites 6 and 7, Figure 1), although a systematic study across sites is pending and some of the telemetry data are unpublished.
Guanacos are often actively chased out of areas with sheep by ranchers who consider them unwelcome competitors, further limiting their ability to move freely across the range. The potentially dramatic effect of limitation of movements is illustrated by the case of the Cabo dos Bahias Reserve [78], which has been presented as evidence of poor population regulation by guanacos [54]. This is a small reserve surrounded by the Atlantic ocean on one side and by sheep ranches on the others, where ranchers persecute guanacos and effectively chase them into the reserve ([78], R. Baldi, pers. comm.). Unable to move to track variable plant productivity, with forced immigration from neighboring ranches and prevented emigration from the reserve, 60% of guanacos died of starvation in one winter [88] and the population has continued to grow fast and to very high levels in recent years, possibly heading towards another die off [78]. Even if local reproduction had declined due to density dependence, the population was forced to grow artificially due to human impact in neighboring areas [78], from which positive net immigration occurred, highlighting how restriction of natural movement prevents both seasonal use of different habitats and also natural population regulation. In summary, this unusual site, where guanaco numbers have grown well above carrying capacity and shown a die off, appears to be a human-created population sink [89] and a poor example for conclusions on natural population regulation. Conversely, it seems to be a useful site to understand the importance of guanaco movements and their interaction with grazing impacts.
Information about guanaco movement from genetic, archeological, historic, and recent telemetry data supports the need for landscape-level planning for coexistence between guanacos and livestock. Specifically, for example, recovery of bottom-up regulation of guanaco populations that can regain some of their seasonal and dispersal movement areas should be a central conservation goal. Seasonal migration and dispersal corridors would take very different forms in southern or coastal steppe ranches with small altitudinal gradients and in Andean steppes with large environmental gradients. However, in both cases it will require modifications of livestock fence layouts and designs and consensus-driven agreements with ranchers and herders. These agreements may, at the same time, facilitate conversations and adjustments of fence layouts that could reduce heterogeneous livestock grazing that is a key driver of overgrazing [58,90].
5. Lesson 3: Long History of Possibly Strong Indirect Effects of Humans on Wildlife Communities
The strong relationship between humans and guanacos through most of the 15,000 years since humans arrived has possibly had significant indirect impacts on herbivory, predation, apparent competition, and scavenging, and on the abundances and distributions of species fulfilling those roles. Pires et al. [29] used network analysis to explore the potential ramifications of the role of humans during the late Pleistocene and Holocene ages, suggesting that humans may have made ecosystems more connected and also more vulnerable to perturbations. In addition to contributing to extinctions of some megafauna by direct hunting, human impacts may have extended through indirect interactions. For example, if human hunting depleted prey species that were important in the diet of other predators, predation pressure on alternative prey species would have increased, an indirect process termed apparent competition [91].
Where guanacos are abundant today, they are the bulk of the diet of pumas and Andean condors, which likely occurred throughout the Holocene period in Patagonia [29,77,92,93]. A recent genetic study [77] partially confirmed the network analysis of indirect human impacts [29]. As guanaco numbers increased through the Holocene period, Perrig et al. [77] were surprised to find that, unlike humans, puma and Andean condor numbers remained stable and at low levels. Perrig et al. [77] have suggested that the lack of a puma increase could have been a consequence of intense competition with human hunters, but did not extend this effect to condors nor propose an explanation for why condor abundance did not track guanaco abundance. Based on the evidence available, we propose that an indirect effect of humans was as likely for Andean condors as it was for pumas.
In present day Andean steppe habitats with little human disturbance, puma predation on camelids is the main source of food for condors, as condors are able to utilize carcasses after pumas partially consume them [93]. Unlike other continents where carcasses of large-bodied native ungulates provide predators and scavengers with food that may last days, in South America, even where native ungulates are abundant, intact carcasses are scarce as the relatively small carcasses of native ungulates are consumed quickly (i.e., in a few hours when condor groups descend on a carcass [93], A. Novaro, personal observation). Wherever pumas are abundant, on the other hand, most (>70%) guanaco mortality that provides carcasses for scavengers is due to depredation [94,95], even with depredation not exerting a regulatory role on guanacos. Holocene-era human hunters in Patagonia, who, like pumas, were efficient guanaco hunters, reached densities that were higher (5 to 20 humans per 100 km2, or 1 human per 200 guanacos [20,22]) than healthy, self-regulated puma populations (1 to 5 pumas per 100 km2, reviewed by [92]). In contrast to the mostly solitary pumas [96], human hunter–gatherers hunted and lived in groups, and they depended on skins, bones, and other guanaco parts as well as meat [22,35,36], so they probably removed entire carcasses of guanacos from kill sites, leaving little food for condors. Thus, if Holocene human hunting of guanacos indirectly prevented an increase in puma numbers, as Perrig et al. [77] proposed, it seems similarly possible that they did the same to condors, limiting their food availability and extending their indirect effects to the scavenger assemblage.
By inserting themselves into the ecological network of the late Pleistocene period, humans had multiple and extensive impacts on that network [29]. Then, in the last 100+ years, they have inserted numerous new elements to the pauperized network, diversifying and supplementing the prey base for generalist native carnivores and raptors, and resulting in apparent competition with native herbivores in addition to the direct competition through capture of resources [41,44,45]. Understanding the extensive roles of humans through direct and indirect interactions in the past could help us reshape conservation objectives and actions in the present. For example, if condors in the steppe were indeed limited by guanaco carcass availability when guanacos were abundant in the late Holocene period, then future restoration of their populations in the steppe perhaps can be more ambitious in terms of range and numbers [97] as their main native food source recovers in the 21st century. On the other hand, although the puma and culpeo are not new predators, as humans were in the late Pleistocene era, they are now supplemented to a degree that they never were before. Just as pumas were perhaps prevented by humans from increasing in number tracking their principal prey, today, rarer and more specialized carnivores, such as the Andean cat, may be prevented from recovering or even caused to decline due to competition for prey, in addition to the numerous other direct and indirect impacts from human activities they face [41,98]. The ongoing expansion of livestock protection tools in Patagonia, like livestock-guarding dogs, should help reduce supplementation of abundant carnivores, indirectly benefiting rarer carnivores, in addition to helping coexistence between livestock husbandry and carnivores.
6. Lesson 4: Climate Change
We struggle as conservation practitioners with how to mitigate and adapt to the effects of human-induced climate change on wildlife in Patagonia in the 21st century, and we were struck by the parallels and differences with the late Pleistocene scenarios described in recent publications. Humans arrived in Patagonia during a time when significant changes in climate were already impacting habitat distribution and species persistence, abundance, and distribution. The arrival of humans was followed by a warming period that possibly exacerbated their direct and indirect effects as predators on wildlife communities [18,19,23,29,70]. The late Holocene drying period in northwest Patagonia, combined with hunting by abundant humans, which possibly resulted in a marked guanaco decline, in turn caused dramatic changes in human economy and society and their impacts on medium-sized species [25,26]. Once again, we are in a period of climate change today in the Anthropocene era, this time change created by humans, and the direct and indirect impacts of humans are no longer restricted to our role as an apex predator [99]. If our impact was so strong in the last two major periods of climate change when our role was much simpler, our potential impact today is devastating.
Within South America, the current human population and biodiversity of Patagonia are particularly vulnerable to climate change. The recent warming trend in Patagonia is among the clearest globally, with an average temperature increase of 1 degree centigrade in the last 50 years. Average temperatures in Patagonia are projected to continue to increase during the rest of the 21st century (between 1 and 4 degrees more, depending on the model) and rainfall will decline between 10% and 30% in much of the region, resulting in increasing aridity, particularly in the north, center, and west, leading to widespread declines of plant cover in grasslands and shrublands by 2060 [100,101], J. Gaitán unpubl. data]. As the production of forage for livestock and wildlife in the steppe is dependent on the balance between precipitation and temperature, and productivity is already relatively low, the rural way of life and biodiversity will be subjected to increasing levels of stress due to increased aridity and a higher frequency of extreme events. The likely interaction between these predicted changes and the current trend in desertification caused by the last few decades of overgrazing by livestock could be catastrophic.
Fortunately, at the same time that our impact in this current climate change scenario could be much greater than in the late Pleistocene era, we now have knowledge and tools to help us mitigate and adapt to the impacts and interactions among these processes. Livestock grazing levels and distribution can be adapted seasonally to respond to highly variable precipitation and productivity patterns, which can be monitored with combinations of remote sensing and a network of field stations available throughout the steppe [102]. Extensive grasslands and rangelands, like the Patagonian steppe, are increasingly valued globally as possible carbon sinks where there is much potential to improve retention of carbon as well as water by improving practices such as livestock grazing [103]. These changes would help mitigate climate change while addressing the most pressing threat to the steppe habitats, wildlife, and people’s livelihood. Simultaneously, guanacos may be more tolerant than sheep, cattle, and even goats to arid conditions and overgrazed steppe habitats, and their impacts on stressed plants and soils may be less severe. Even with abundant guanaco populations that had recovered in a large area of southern Patagonia abandoned by ranchers in the late 20th century, in part due to overgrazing, plant productivity has been stable or increasing during the last 15 years [81]. A kilogram of guanaco fiber from live-sheared and wild populations under appropriate animal welfare standards may sell for 40 times the price of sheep wool [104]. Tools to implement this live shearing have been developed locally in the last 15 years [66,105]. Thus, one of the few survivors of the Pleistocene megafaunal extinctions may help build a diversified economy that is better adapted to the changes that will occur in the 21st century if allowed to coexist with well managed livestock.
Strategies for protected area planning and implementation in Patagonia also must incorporate the lessons from past impacts of climate changes, humans, and other pressures, and from projected changes in those pressures and in habitat conditions for vulnerable species [106,107]. Most protected areas in the Patagonian steppe and scrub are small and isolated, while Patagonian forest reserves along the Andes are much larger and connected [108]. Increasing protected area size and connectivity will facilitate species migration and rescue effects in the steppe and scrub [3], but efforts to provide connectivity [109] at the scale needed in the steppe will need to be integrated with conservation-oriented management of private ranches and herder lands. An ongoing project led by the Argentine Park Service and supported by local governments, scientists, and NGOs is proposing the creation of new conservation cores and connectivity corridors across the steppe and scrub to help conserve Patagonia’s biodiversity in a changing climate [108].
7. Rewilding versus Coexistence or Inclusive Rewilding?
Seventeen years ago, we collaborated with a group of colleagues from several government and non-governmental institutions to elaborate a vision and strategy for conservation in the Patagonian steppe centered around the idea of rewilding [110]. With only 1% of the steppe effectively protected [111] in mostly small and isolated protected areas, and livestock and invasive species widespread, we focused on the need to establish core areas where ecological processes, such as predation on guanacos by pumas, could be restored to their levels prior to European colonization, and where wildlife connectivity through productive lands prevented those cores from becoming islands. We and many colleagues have worked since then to realize that vision by helping to create and expand reserves and facilitate recovery of wildlife on lands used for livestock and hydrocarbon production to promote connectivity among the reserves.
Our rewilding vision, however, was somewhat naive, as it lacked an understanding of the direct and indirect roles of early Patagonian humans in shaping the ecosystems that we sought to rewild. Although we embraced the human role by naming the network of core reserves and connectivity matrices “Tehuelche landscapes”, apart from the Tehuelche’s extensive reliance on guanaco for subsistence, we did not know much about how the Tehuelche ancestors shaped those wild areas. Apart from an understanding that local people who coexist with wildlife must embrace a rewilding vision to make it a reality, seventeen years ago there were few tested tools to help those people embrace it. The last few years have re-shaped our understanding of the possible direct and indirect roles of people and their interaction with climate in the distant and recent past thanks to interdisciplinary work, and new techniques have been tested or are available for testing that fully embrace coexistence. We believe that the examples we provide above, drawn from some of the recent advances and analyzed in the context of ecological processes and conservation strategies, present a unique opportunity to update a rewilding vision for the Patagonian steppe that incorporates a consideration of the lessons learned on the ecological and evolutionary roles of people–wildlife interactions.
An updated vision for large and habitat-diverse core areas of a wild Patagonia must restore and/or preserve seasonal and dispersal movements of wildlife, as well as other processes, like depredation and scavenging. These ecosystems in core areas must be allowed to evolve and adapt to the dramatic changes that will occur in the 21st century and beyond. Interconnected as much as possible with each other, they will be baselines and sources for future recolonization of species that may go extinct in the matrix between cores. Conservation of core areas may be maximized through connectivity corridors and surrounding mosaics of productive lands where rural human populations, which have been declining rapidly in recent decades, conserve their traditional livelihoods, carrying out ecologically and economically sustainable livestock husbandry while coexisting with teeming wildlife, rangelands, and soils that evolve and adapt as well.
This updated vision for rewilding of Patagonia, which considers the influences of humans since their arrival 15,000 years ago while prioritizing a coexistence strategy that brings Patagonian people of today on board, is consistent with the concept of inclusive rewilding [112]. This concept merges the call for valuing thriving and resilient wildlife assemblages while considering the needs and aspirations of the people that must coexist with wildlife, bringing rewilding and coexistence together. Inclusive rewilding seems like a fitting term for a region where the current rural population of ca. 150,000 people is similar in size to the hunter/gatherer population that was sustained mostly by guanacos 500 years ago. However, these rural people for the most part now see guanacos, pumas, condors, and conservationists as their nemesis. A framework of inclusive rewilding can be a conceptual approach that helps bridge this gap and promotes dialogue, in addition to providing an interdisciplinary perspective, bringing together the valuing of species’ ecological roles and interactions and a strong emphasis on addressing challenges of human–wildlife coexistence.
Author Contributions
Conceptualization, A.J.N. and R.S.W.; writing—original draft preparation, A.J.N. and R.S.W.; writing—review and editing, A.J.N. and R.S.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No original data were collected for this manuscript.
Acknowledgments
We thank Ivan Perez, Diego Rindel, and Luciano Prates for stimulating discussions about their recent discoveries and for sharing new data on the history of humans and wildlife in Patagonia and the editors and two anonymous reviewers for help improving the manuscript. We thank Martin Funes, Ricardo Baldi, Emiliano Donadío, Sebastián Di Martino, Alejandro Vila, Claudio Chehébar, Gabriel Oliva, and many other colleagues in Patagonia and elsewhere for enlightening conversations and joint learning about wildlife conservation and rewilding over the last 30 years. We are extremely grateful to the editors of this issue for inviting our contribution and for their patience. We thank Lara Heidel for help with the figure. We thank Avecita Chicchon of the Gordon and Betty Moore Foundation for donations in memory of Frank Chicchon that allowed one of us to devote the time to work on this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Prates, L.; Politis, G.G.; Perez, S.I. Rapid radiation of humans in South America after the last glacial maximum: A radiocarbon-based study. PLoS ONE 2020, 15, e0236023. [Google Scholar] [CrossRef]
- Dietl, G.P.; Kidwell, S.M.; Brenner, M.; Burney, D.A.; Flessa, K.W.; Jackson, S.T.; Koch, P.L. Conservation paleobiology: Leveraging knowledge of the past to inform conservation and restoration. Annu. Rev. Earth Planet. Sci. 2015, 43, 79–103. [Google Scholar] [CrossRef]
- Graham, R.W. The role of climatic change in the design of biological reserves: The paleoecological perspective. Cons. Biol. 1988, 2, 391–394. [Google Scholar] [CrossRef]
- Louys, J. (Ed.) Paleontology in Ecology and Conservation; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–286. [Google Scholar]
- Wolverton, S.; Lyman, R.L. (Eds.) Conservation Biology and Applied Zooarchaeology; University of Arizona Press: Tucson, AZ, USA, 2012; pp. 1–241. [Google Scholar]
- Rowan, J.; Beaudrot, L.; Franklin, J.; Reed, K.E.; Smail, I.E.; Zamora, A.; Kamilar, J.M. Geographically divergent evolutionary and ecological legacies shape large mammal biodiversity in the global tropics and sub-tropics. Proc. Nat. Acad. Sci. USA 2020, 117, 1559–1565. [Google Scholar] [CrossRef]
- Vegas-Villarrúbia, T.; Rull, V.; Montoya, E.; Safont, E. Quaternary palaeoecology and nature conservation: A general review with examples from the neotropics. Quat. Sci. Rev. 2011, 30, 2361–2388. [Google Scholar] [CrossRef]
- Lyman, R.L. Biodiversity, paleozoology, and conservation biology. In Paleontology in Ecology and Conservation; Louys, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 147–169. [Google Scholar]
- Sandom, C.; Faurby, S.; Sandel, B.; Svenning, J.C. Global late Quaternary megafauna extinctions linked to humans, not climate change. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, P.L.; Barnosky, A.D. Late Quaternary Extinctions: State of the Debate. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 215–250. [Google Scholar] [CrossRef] [Green Version]
- Barnosky, A.D.; Lindsey, E.L. Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quat. Int. 2010, 217, 10–29. [Google Scholar] [CrossRef]
- Firestone, R.B.; West, A.; Kennett, J.P.; Becker, L.; Bunch, T.E.; Revay, Z.S.; Schultz, P.H.; Belgya, T.; Kennett, D.J.; Erlandson, J.M.; et al. Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and the Younger Dryas cooling. Proc. Nat. Acad. Sci. USA 2007, 104, 16016–16021. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, B.M.; Laub, R. Hyperdisease in the late Pleistocene: Validation of an early 20th century hypothesis. Naturwissenschaften 2006, 93, 557–564. [Google Scholar] [CrossRef]
- Martin, P.S. The discovery of America: The first Americans may have swept the Western Hemisphere and decimated its fauna within 1000 years. Science 1973, 179, 969–974. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, L.; Rick, T.; Wolverton, S. The overkill model and its impact on environmental research. Ecol. Evol. 2018, 8, 9683–9696. [Google Scholar] [CrossRef]
- Surovell, T.A.; Pelton, S.R.; Anderson-Sprecher, R.; Myers, A.D. Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna. Proc. Nat. Acad. Sci. USA 2016, 113, 886–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolverton, S. The North American Pleistocene overkill hypothesis and the re-wilding debate. Div. Distr. 2010, 16, 874–876. [Google Scholar] [CrossRef]
- Prates, L.; Perez, S.I. Late Pleistocene South American megafaunal extinctions associated with rise of Fishtail points and human population. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Metcalf, J.L.; Turney, C.; Barnett, R.; Martin, F.; Bray, S.C.; Vilstrup, J.T.; Orlando, L.; Salas-Gismondi, R.; Loponte, D.; Medina, M.; et al. Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation. Sci. Adv. 2016, 2, e1501682. [Google Scholar] [CrossRef] [Green Version]
- Perez, S.I.; Postillone, M.B.; Rindel, D.; Gobbo, D.; Gonzalez, P.N.; Bernal, V. Peopling time, spatial occupation and demography of Late Pleistocene–Holocene human population from Patagonia. Quat. Int. 2016. 425, 214–223. [CrossRef]
- Hernández, F.; Ríos, C.; Perotto-Baldivieso, H.L. Evolutionary history of herbivory in the Patagonian steppe: The role of climate, ancient megafauna, and guanaco. Quat. Sci. Rev. 2019, 220, 279–290. [Google Scholar] [CrossRef]
- Rindel, D.D.; Moscardi, B.F.; Perez, S.I. Historias milenarias de presas y predadores: Guanacos y humanos en Patagonia. Museo 2020, 32, 47–54. [Google Scholar]
- Rindel, D.D.; Moscardi, B.F.; Perez, S.I. The distribution of the guanaco (Lama guanicoe) in Patagonia during Late Pleistocene–Holocene and its importance for prehistoric human diet. Holocene 2021, 31, 644–657. [Google Scholar] [CrossRef]
- Moscardi, B. Human diet evolution in Patagonia was driven by the expansion of Lama guanicoe after megafaunal extinctions. J. Arch. Sci. 2020, 10, 105098. [Google Scholar] [CrossRef]
- Gil, A.F.; Villalba, R.; Franchetti, F.R.; Otaola, C.; Abbona, C.C.; Peralta, E.A.; Neme, G. Between foragers and farmers: Climate change and human strategies in Northwestern Patagonia. Quaternary 2020, 3, 17. [Google Scholar] [CrossRef]
- Abbona, C.C.; Neme, G.; Johnson, J.; Gil, A.; Villalba, R.; Nagaoka, L.; Kim, T.; Wolverton, S. Sustainable harvest or resource depression? Using ancient DNA to study the population dynamics of guanaco in western Argentina during the Holocene. J. Archaeol. Sci. 2021, 129, 105355. [Google Scholar] [CrossRef]
- González, B.A.; Palma, R.E.; Zapata, B.; Marín, J.C. Taxonomic and biogeographical status of guanaco Lama guanicoe (Artiodactyla, Camelidae). Mammal Rev. 2006, 36, 157–178. [Google Scholar] [CrossRef]
- Prevosti, F.J.; Martin, F.M. Paleoecology of the mammalian predator guild of Southern Patagonia during the latest Pleistocene: Ecomorphology, stable isotopes, and taphonomy. Quat. Int. 2013, 305, 74–84. [Google Scholar] [CrossRef]
- Pires, M.M.; Rindel, D.; Moscardi, B.; Cruz, L.R.; Guimarães, P.R., Jr.; dos Reis, S.F.; Perez, S.I. Before, during and after megafaunal extinctions: Human impact on Pleistocene-Holocene trophic networks in South Patagonia. Quat. Sci. Rev. 2020, 250, 106696. [Google Scholar] [CrossRef]
- Sarno, R.J.; Franklin, W.L.; O’Brien, S.J.; Johnson, W.E. rPatterns of mtDNA and microsatellite variation in an island and mainland population of guanacos in southern Chile. Anim. Conserv. Forum 2001, 4, 93–101. [Google Scholar] [CrossRef]
- Bolgeri, M.J. Caracterización de movimientos migratorios en guanacos (Lama guanicoe) y patrones de depredación por pumas (Puma concolor) en la Payunia, Mendoza. Ph.D. Thesis, Universidad Nacional del Comahue, San Carlos de Bariloche, Argentina, 2016. [Google Scholar]
- Ortega, I.M.; Franklin, W.L. Social organization, distribution and movements of a migratory guanaco population in the Chilean Patagonia. Rev. Chil. Hist. Nat. 1995, 68, 489–500. [Google Scholar]
- Moraga, C.A.; Funes, M.C.; Pizarro, J.C.; Briceño, C.; Novaro, A.J. Effects of livestock on guanaco Lama guanicoe density, movements and habitat selection in a forest–grassland mosaic in Tierra del Fuego, Chile. Oryx 2015, 49, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Rey, A.; Novaro, A.J.; Guichon, M.L. Guanaco (Lama guanicoe) mortality by entanglement in wire fences. J. Nat. Conserv. 2011, 20, 280–283. [Google Scholar] [CrossRef]
- Musters, L. A year in Patagonia. J. R. Geogr. Soc. Lond. 1871, 41, 59–77. [Google Scholar] [CrossRef]
- Pero, A. The Tehuelche of Patagonia as chronicled by travelers and explorers of the nineteenth century. In Archaeological and Anthropological Perspectives on the Native Peoples of Pampa, Patagonia, and Tierra del Fuego to the Nineteenth Century; Briones, C., Lanata, J.L., Eds.; Greenwood Publishing Group: Westwood, CT, USA, 2002; pp. 103–119. [Google Scholar]
- Sala, A.; Corach, D. Analysis of admixture and genetic structure of two Native American groups of Southern Argentinean Patagonia. Mol. Biol. Rep. 2014, 41, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, H.F.; Elissalde, N.O.; Gagliardini, D.A.; Milovich, J. Status of desertification in the Patagonian region: Assessment and mapping from satellite imagery. Arid Land Res. Manag. 1998, 12, 95–121. [Google Scholar] [CrossRef]
- Baldi, R.; Albon, S.; Elston, D. Guanacos and sheep: Evidence for continuing competition in arid Patagonia. Oecologia 2001, 129, 561–570. [Google Scholar] [CrossRef]
- Baldi, R.; Pelliza-Sbriller, A.; Elston, D.; Albon, S. High potential for competition between guanacos and sheep in Patagonia. J. Wildl. Manag. 2004, 68, 924–938. [Google Scholar] [CrossRef]
- Barbar, F.; Lambertucci, S.A. Introduced lagomorph produce stronger potential apparent competition in invaded communities than any other species in a similar but native food web. Biol. Invasions 2019, 21, 3735–3740. [Google Scholar] [CrossRef]
- Baldi, R.; Pirronitto, A.; Burgi, M.V.; Antún, M. Abundance Estimates of the Lesser Rhea Rhea pennata pennata in the Argentine Patagonia: Conservation Implications. Front. Ecol. Evol. 2015, 345, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Bonino, N.A. Introduced mammals in Patagonia, southern Argentina: Consequences, problems, and management considerations. In Proceedings of the First International Wildlife Management Congress; The Wildlife Society: Bethesda, MD, USA, 1995; pp. 406–409. [Google Scholar]
- Novaro, A.; Funes, M.; Walker, R.S. Ecological extinction of native prey of a carnivore assemblage in Argentine Patagonia. Biol. Conserv. 2000, 92, 25–33. [Google Scholar] [CrossRef]
- Novaro, A.J.; Walker, R.S. Human-induced changes in the role of carnivores in Patagonia. In Large carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Stenneck, R., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 267–287. [Google Scholar]
- Baldi, R.; De Lamo, D.A.; Failla, M.; Ferrando, P.; Funes, M.C.; Nugent, P.; von Thüngen, J. Plan Nacional de Manejo del Guanaco (Lama guanicoe); Secretaría de Ambiente y Desarrollo Sustentable de la Nación: Buenos Aires, Argentina, 2006; pp. 1–50. [Google Scholar]
- Plaza, P.I.; Lambertucci, S.A. Ecology and conservation of a rare species: What do we know and what may we do to preserve Andean condors? Biol. Conserv. 2020, 251, 108782. [Google Scholar] [CrossRef]
- Golluscio, R.A.; Deregibus, V.A.; Paruelo, J.M. Sustainability and range management in the Patagonian steppes. Ecol. Austral. 1998, 8, 265–284. [Google Scholar]
- Oliva, G.; Paredes, P.; Ferrante, D.; Cepeda, C.; Rabinovich, J. Remotely sensed primary productivity shows that domestic and native herbivores combined are overgrazing Patagonia. J. Appl. Ecol. 2019, 56, 1575–1584. [Google Scholar] [CrossRef]
- Travaini, A.; Zapata, S.C.; Bustamante, J.; Pedrana, J.; Zanón, J.I.; Rodríguez, A. Guanaco abundance and monitoring in Southern Patagonia: Distance sampling reveals substantially greater numbers than previously reported. Zool. Stud. 2015, 54, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Mesas, A.; Baldi, R.; González, B.A.; Burgi, V.; Chávez, A.; Johnson, W.E.; Marín, J.C. Past and recent effects of livestock activity on the genetic diversity and population structure of native guanaco populations of arid Patagonia. Animals 2021, 11, 1218. [Google Scholar] [CrossRef]
- Walker, S.; Novaro, A. The world’s southernmost pumas in Patagonia and the Southern Andes. In Cougar: Ecology and Conservation; Hornocker, M., Negri, S., Eds.; University of Chicago Press: Chicago, IL, USA, 2010; pp. 91–99. [Google Scholar]
- Novaro, A.; Walker, S.; Palacios, R.; Di Martino, S.; Monteverde, M.; Cañadell, S.; Rivas, L.; Cossíos, D. Endangered Andean cat distribution beyond the Andes in Patagonia. Cat News 2010, 53, 8–10. [Google Scholar]
- Oliva, G.; Paredes, P.; Ferrante, D.; Cepeda, C.; Rabinovich, J. Are Patagonia grasslands being overgrazed? A response to Marino et al. (2020). J. Appl. Ecol. 2020, 57, 2399–2405. [Google Scholar] [CrossRef]
- Adler, P.B.; Milchunas, D.G.; Sala, O.E.; Burke, I.C.; Lauenroth, W.K. Plant traits and ecosystem grazing effects: Comparison of US sagebrush steppe and Patagonian steppe. Ecol. Appl. 2005, 15, 774–792. [Google Scholar] [CrossRef] [Green Version]
- Lauenroth, W.K. Guanacos, spiny shrubs and the evolutionary history of grazing in the Patagonian steppe. Ecol. Austral. 1998, 8, 211–215. [Google Scholar]
- Marino, A.; Rodríguez, V.; Pazos, G. Resource-defense polygyny and self-limitation of population density in free-ranging guanacos. Behav. Ecol. 2016, 27, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Marino, A.; Rodríguez, V.; Schroeder, N.M. Wild guanacos as scapegoat for continued overgrazing by livestock across southern Patagonia. J. Appl. Ecol. 2020, 57, 2393–2398. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Cingolani, A.M.; Posse, G.; Collantes, M.B. Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands. J. Appl. Ecol. 2005, 42, 50–59. [Google Scholar] [CrossRef]
- König, H.E.; Skewes, O.; Helmreich, M.; Böck, P. Macroscopic and histological investigation of guanaco footpads (Lama guanicoe, Müller 1776). J. Morphol. 2015, 276, 331–341. [Google Scholar] [CrossRef]
- Duncan, K.; Holdaway, R. Footprint pressures and locomotion of moas and ungulates and their effects on the New Zealand indigenous biota through trampling. N. Z. J. Ecol. 1989, 12, 97–101. [Google Scholar]
- Bonino, N.; Pelliza Sbriller, A. Composición botánica de la dieta del guanaco (Lama guanicoe) en dos ambientes contrastantes de Tiera del Fuego, Argentina. Ecol. Austral. 1991, 1, 97–102. [Google Scholar]
- Fraser, M.D.; Gordon, I.J. The diet of goats, red deer and South American camelids feeding on three contrasting Scottish upland vegetation communities. J. Appl. Ecol. 1997, 34, 668–686. [Google Scholar] [CrossRef]
- San Martin, F.; Bryant, F.C. Nutrition of domesticated South American llamas and alpacas. Small Rumin. Res. 1989, 2, 191–216. [Google Scholar] [CrossRef]
- Rey, A.; Novaro, A.J.; Sahores, M.; Guichon, M.L. Demographic effects of live shearing on a guanaco population. Small Rumin. Res. 2012, 107, 92–100. [Google Scholar] [CrossRef]
- Baldi, R.; Campagna, C.; Saba, S. Abundancia y distribución del guanaco (Lama guanicoe) en el NE del Chubut, Patagonia Argentina. Mastozool. Neotrop. 1997, 4, 5–15. [Google Scholar]
- Iranzo, E.C.; Traba, J.; Acebes, P.; González, B.A.; Mata, C.; Estades, C.F.; Malo, J.E. Niche segregation between wild and domestic herbivores in Chilean Patagonia. PLoS ONE 2013, 8, e59326. [Google Scholar]
- Franklin, W.L. Contrasting socioecologies of South America’s wild camelids: The vicuna and guanaco. In Advances in the Study of Mammalian Behavior; Special Publication No. 7; Eisenberg, J.F., Kleiman, D., Eds.; American Society of Mammalogists: Shippensburg, PA, USA, 1983; pp. 573–629. [Google Scholar]
- Villavicencio, N.A.; Lindsey, E.L.; Martin, F.M.; Borrero, L.A.; Moreno, P.I.; Marshall, C.R.; Barnosky, A.D. Combination of humans, climate, and vegetation change triggered Late Quaternary megafauna extinction in the Última Esperanza region, southern Patagonia, Chile. Ecography 2016, 39, 125–140. [Google Scholar] [CrossRef]
- Holling, C. The components of predation as revealed by a study of small mammal predation of the European pine sawfly. Can. Entomol. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Miotti, L.; Salemme, M.C. When Patagonia was colonized: People mobility at high latitudes during Pleistocene/Holocene transition. Quat. Int. 2003, 109, 95–111. [Google Scholar] [CrossRef]
- Hopcraft, J.G.C.; Olff, H.; Sinclair, A.R.E. Herbivores, resources and risks: Alternating regulation along primary environmental gradients in savannas. Trends Ecol. Evol. 2010, 25, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Messier, F. Ungulate population models with predation: A case study with the North American moose. Ecology 1994, 75, 478–488. [Google Scholar] [CrossRef]
- Ballard, W.B.; Lutz, D.; Keegan, T.W.; Carpenter, L.H.; de Vos, J.C., Jr. Deer-predator relationships: A review of recent North American studies with emphasis on mule and black-tailed deer. Wildl. Soc. Bull. 2001, 29, 99–115. [Google Scholar]
- Benson, J.F.; Loveless, K.M.; Rutledge, L.Y.; Patterson, B.R. Ungulate predation and ecological roles of wolves and coyotes in eastern North America. Ecol. Appl. 2017, 27, 718–733. [Google Scholar] [CrossRef] [Green Version]
- Perrig, P.L.; Fountain, E.D.; Lambertucci, S.A.; Pauli, J.N. Demography of avian scavengers after Pleistocene megafaunal extinction. Sci. Rep. 2019, 9, 9680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, A.; Pascual, M.; Baldi, R. Ecological drivers of guanaco recruitment: Variable carrying capacity and density dependence. Oecologia 2014, 175, 1189–1200. [Google Scholar] [CrossRef]
- Cabrera, A.; Yepes, J. Mamíferos Sudamericanos: Vida, Costumbres y Descripción; Ediar, Compañía Argentina de Editores: Buenos Aires, Argentina, 1940. [Google Scholar]
- Torres, H. Guanaco: Distribución y Conservación del Guanaco; IUCN/CSE Grupo Especialista en Camélidos Sudamericanos: London, UK, 1985; Informes Especial No. 2. [Google Scholar]
- Gaitán, J.J.; Bran, D.E.D.; Azcona, C. Trend of NDVI in the period 2000–2014 as indicator of land degradation in Argentina: Advantages and limitations. AgriScientia 2015, 32, 83–93. [Google Scholar] [CrossRef]
- Pazos, G.E.; Rodriguez, M.V.; Blanco, P.D. Vegetación terrestre: Descripción, monitoreo y relación con el clima y los herbívoros. In Reserva de Vida Silvestre San Pablo de Valdés: 10 Años Conservando el Patrimonio Natural y Cultural de la Península Valdés, Patagonia Argentina; Udrizar-Sauthier, D.E., Pazos, G.E., Arias, A., Eds.; Fundación Vida Silvestre Argentina-CONICET: Buenos Aires, Argentina, 2017; pp. 73–97. [Google Scholar]
- Milner-Gulland, E.J.; Fryxell, J.M.; Sinclair, A.R. Animal Migration: A Synthesis; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Serret, A. El Huemul: Fantasma de la Patagonia; Zagier & Urruty Publication: Buenos Aires, Argentina, 2001. [Google Scholar]
- Fernández, P.M.; Cruz, I.; Belardi, J.B.; De Nigris, M.; Muñoz, A.S. Human predation and natural history of huemul (Cervidae; Hippocamelus bisulcus Molina) in Patagonia: A zooarchaeological analysis. J. Ethnobiol. 2015, 35, 472–498. [Google Scholar] [CrossRef]
- Tessone, A.; Fernández, P.; Fernández, N.; De Nigris, M. Variaciones δ13C y δ15N en huemul (Hippocamelus bisulcus) durante el Holoceno en Cerro Casa de Piedra, Santa Cruz, Argentina. Implicancias para el estudio de su distribución pasada. Intersecc. Antropol. 2020, 21, 5–16. [Google Scholar] [CrossRef]
- Wolverton, S.; Otaola, C.; Neme, G.; Giardina, M.; Gil, A. Patch choice, landscape ecology, and foraging efficiency: The zooarchaeology of late Holocene foragers in Western Argentina. J. Ethnobiol. 2015, 35, 499–518. [Google Scholar] [CrossRef]
- Beldomenico, P.M.; Uhart, M.; Bono, M.F.; Marull, C.; Baldi, R.; Peralta, J.L. Internal parasites of free-ranging guanacos from Patagonia. Vet. Parasitol. 2003, 118, 71–77. [Google Scholar] [CrossRef]
- Pulliam, H.R. Sources, sinks, and population regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Oñatibia, G.R.; Aguiar, M.R. Paddock size mediates the heterogeneity of grazing impacts on vegetation. Rangel. Ecol. Manag. 2018, 71, 470–480. [Google Scholar] [CrossRef]
- Holt, R.D.; Bonsall, M.B. Apparent competition. Ann. Rev. Ecol. Evol. Syst. 2017, 48, 447–471. [Google Scholar] [CrossRef]
- Gelin, M.L.; Branch, L.C.; Thornton, D.H.; Novaro, A.J.; Gould, M.J.; Caragiulo, A. Response of pumas (Puma concolor) to migration of their primary prey in Patagonia. PLoS ONE 2017, 12, e0188877. [Google Scholar] [CrossRef] [Green Version]
- Perrig, P.L.; Donadio, E.; Middleton, A.D.; Pauli, J.N. Puma predation subsidizes an obligate scavenger in the high Andes. J. Appl. Ecol. 2017, 54, 846–853. [Google Scholar] [CrossRef]
- Bank, M.S.; Sarno, R.J.; Campbell, N.K.; Franklin, W.L. Predation of guanacos (Lama guanicoe) by southernmost mountain lions (Puma concolor) during a historically severe winter in Torres del Paine National Park, Chile. J. Zool. 2002, 258, 215–222. [Google Scholar] [CrossRef]
- Bolgeri, M.J.; Novaro, A.J. Variación espacial en la depredación por puma (Puma concolor) sobre guanacos (Lama guanicoe) en La Payunia, Mendoza, Argentina. Mastozool. Neotrop. 2015, 22, 255–264. [Google Scholar]
- Seidensticker, J.C.; Hornocker, M.G.; Wiles, W.V.; Messick, J.P. Mountain lion social organization in the Idaho Primitive Area. Wildl. Monogr. 1973, 35, 3–60. [Google Scholar]
- Perrig, P.L.; Lambertucci, S.A.; Cruz, J.; Alarcón, P.A.; Plaza, P.I.; Middleton, A.D.; Blanco, G.; Sánchez-Zapata, J.A.; Donázar, J.A.; Pauli, J.N. Identifying conservation priority areas for the Andean condor in southern South America. Biol. Conserv. 2020, 243, 108494. [Google Scholar] [CrossRef]
- Walker, S.; Funes, M.; Heidel, L.; Palacios, R.; Novaro, A. The Endangered Andean cat and fracking in Patagonia. Oryx 2013, 48, 13–15. [Google Scholar] [CrossRef] [Green Version]
- Darimont, C.T.; Fox, C.H.; Bryan, H.M.; Reimchen, T.E. The unique ecology of human predators. Science 2015, 349, 858–860. [Google Scholar] [CrossRef]
- Barros, V.; Vera, C.; Agosta, E.; Araneo, D.; Camilloni, I.; Carril, A.F.; Doyle, M.E.; Frumento, O.; Nuñez, M.; Ortiz de Zárate, M.I.; et al. Tercera Comunicación Nacional Sobre Cambio Climático. Cambio Climático en Argentina; Tendencias y Proyecciones; Secretaría de Ambiente y Desarrollo Sustentable de la Nación: Buenos Aires, Argentina, 2014. [Google Scholar]
- Pessasq, N.; Flaherty, S.; Solmon, S.; Pascual, M. Climate change in northern Patagonia: Critical decrease in water resources. Theor. Appl. Climatol. 2020, 140, 807–822. [Google Scholar] [CrossRef]
- Oliva, G.; Dos Santos, E.; Sofía, O.; Umaña, F.; Massara, V.; Martínez, G.G.; Caruso, C.; Cariac, G.; Echevarría, D.; Fantozzi, A. The MARAS dataset, vegetation and soil characteristics of dryland rangelands across Patagonia. Sci. Data 2020, 7, 327. [Google Scholar] [CrossRef]
- Bossio, D.A.; Cook-Patton, S.C.; Ellis, P.W.; Fargione, J.; Sanderman, J.; Smith, P.; Wood, S.; Zomer, R.J.; von Unger, M.; Emmer, I.M.; et al. The role of soil carbon in natural climate solutions. Nat. Sustain. 2020, 3, 391–398. [Google Scholar] [CrossRef]
- Baldi, R.; Novaro, A.; Funes, M.; Walker, S.; Ferrando, P.; Failla, M.; Carmanchahi, P. Guanaco management in Patagonian rangelands: A conservation opportunity on the brink of collapse. In Wild Rangelands: Conserving Wildlife while Maintaining Livestock in Semi-Arid Ecosystems; du Toit, J., Kock, R., Deutsch, J., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2010; pp. 266–290. [Google Scholar]
- Carmanchahi, P.; Schroeder, N.; Bolgeri, M.; Walker, R.S.; Funes, M.; Berg, J.; Taraborelli, P.; Ovejero, R.; Gregorio, P.; Moreno, P.; et al. Live-shearing effects on population parameters and movements in sedentary and migratory populations of guanacos. Oryx 2014, 49, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Bonino, M.F.; Azócar, D.L.M.; Schulte, J.A.; Cruz, F.B. Climate change and lizards: Changing species’ geographic ranges in Patagonia. Region. Environ. Chang. 2015, 15, 1121–1132. [Google Scholar] [CrossRef]
- Lazo-Cancino, D.; Rivera, R.; Paulsen-Cortez, K.; González-Berríos, N.; Rodríguez-Gutiérrez, R.; Rodriguez-Serrano, E. The impacts of climate change on the habitat distribution of the vulnerable Patagonian-Fueguian species Ctenomys magellanicus (Rodentia, Ctenomyidae). J. Arid Environ. 2020, 173, 104016. [Google Scholar] [CrossRef]
- Chehébar, C.; Novaro, A.; Iglesias, G.; Walker, S.; Funes, M.; Tammone, M.; Didier, K. Identificación de Áreas de Importancia Para la Biodiversidad en la Estepa y Monte de Patagonia; Administración de Parques Nacionales, Wildlife Conservation Society, and the Nature Conservancy: San Carlos de Bariloche, Argentina, 2013; pp. 1–112. [Google Scholar]
- Hilty, J.; Worboys, G.L.; Keeley, A.; Woodley, S.; Lausche, B.; Locke, H.; Carr, M.; Pulsford, I.; Pittock, J.; White, J.W.; et al. Guidelines for Conserving Connectivity through Ecological Networks and Corridors; Best Practice Protected Area Guidelines Series No. 30; IUCN: Gland, Switzerland, 2020. [Google Scholar]
- Walker, S.; Novaro, A.J.; Funes, M.; Baldi, R.; Chehébar, C.; Ramilo, E.J.; Ayesa, J.A.; Bran, D.M.; Vila, A.; Bonino, N.A. Rewilding Patagonia: In the wild Patagonia reserve network, guanacos, choiques, and pumas will roam free. Wild Earth 2005, 15, 32–37. [Google Scholar]
- Secretaria de Ambiente y Desarrollo Sustentable. Informe Nacional Ambiente y Áreas Protegidas de la Argentina 2008–2018; Presidencia de la Nación y Administración de Parques Nacionales: Buenos Aires, Argentina, 2019; pp. 1–60. [Google Scholar]
- Jørgensen, D. Rethinking rewilding. Geoforum 2015, 65, 482–488. [Google Scholar] [CrossRef]

{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Novaro, A.J.; Walker, R.S. Lessons of 15,000 Years of Human–Wildlife Interaction for Conservation in Patagonia in the 21st Century. Diversity 2021, 13, 633. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120633
AMA Style
Novaro AJ, Walker RS. Lessons of 15,000 Years of Human–Wildlife Interaction for Conservation in Patagonia in the 21st Century. Diversity. 2021; 13(12):633. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120633
Chicago/Turabian StyleNovaro, Andrés J., and Rebecca Susan Walker. 2021. "Lessons of 15,000 Years of Human–Wildlife Interaction for Conservation in Patagonia in the 21st Century" Diversity 13, no. 12: 633. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120633
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.