
Geographical Distribution and Relationship with Environmental Factors of Paphiopedilum Subgenus Brachypetalum Hallier (Orchidaceae) Taxa in Southwest China

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Study Area
2.3. Data Collection
2.3.1. Distribution of Target Orchids
2.3.2. Environmental and Climatic Data
2.4. Data Analyses
2.4.1. Differences of Environmental Factors among Species Distribution Sites
2.4.2. Relationship between Species Distribution and Environmental Factors
3. Results
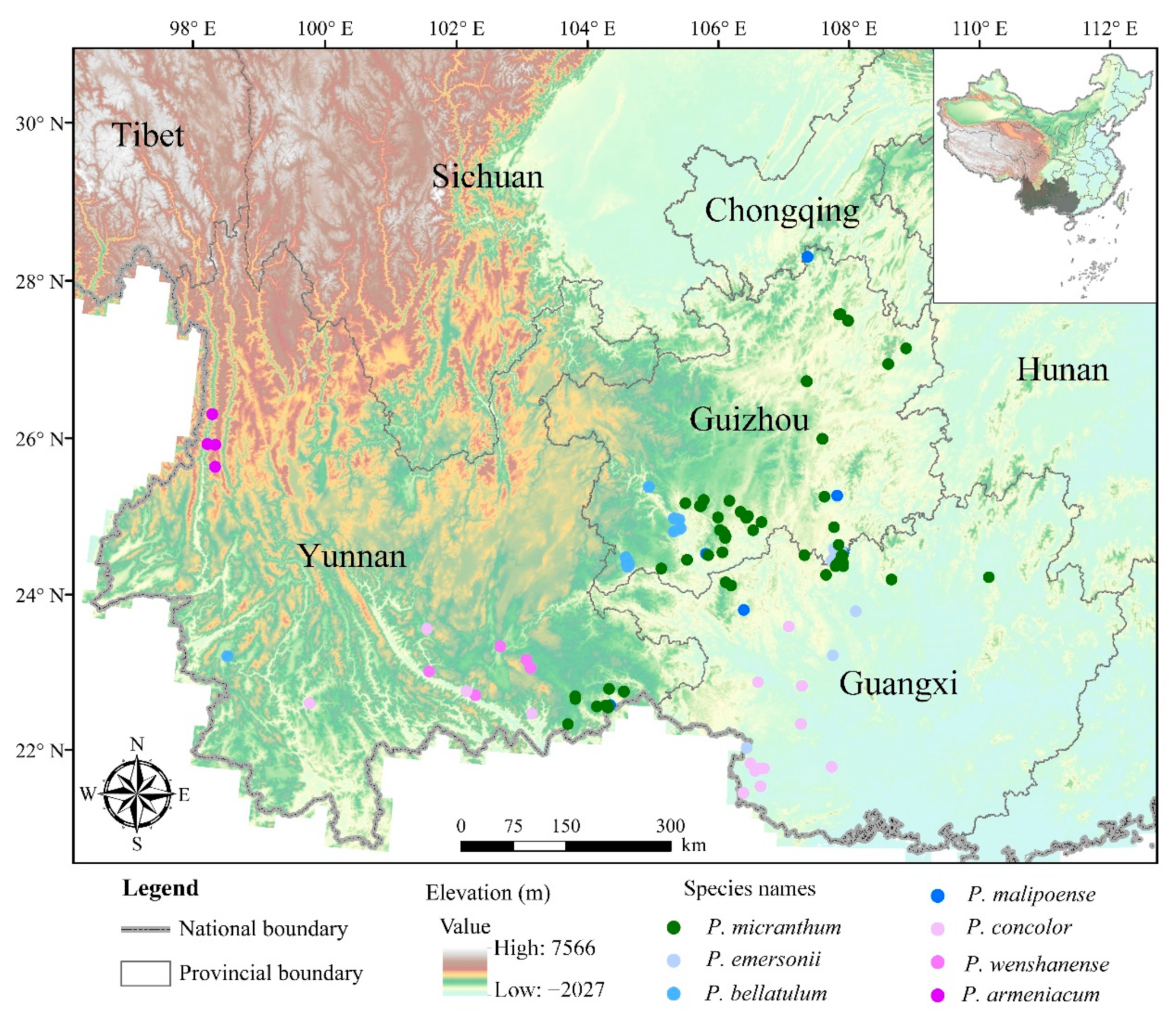
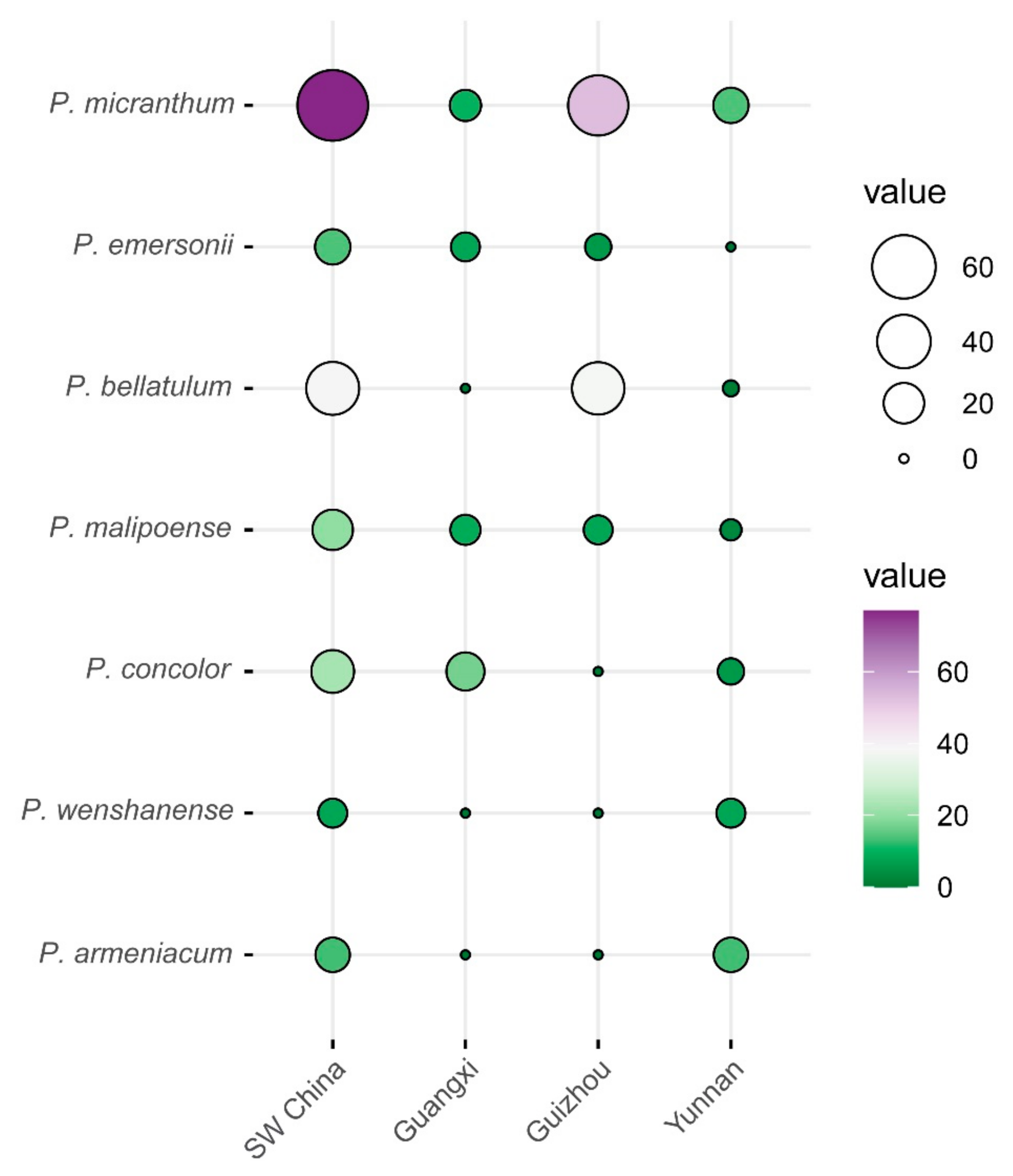
3.1. Distribution Pattern of Brachypetalum Species in Southwest China
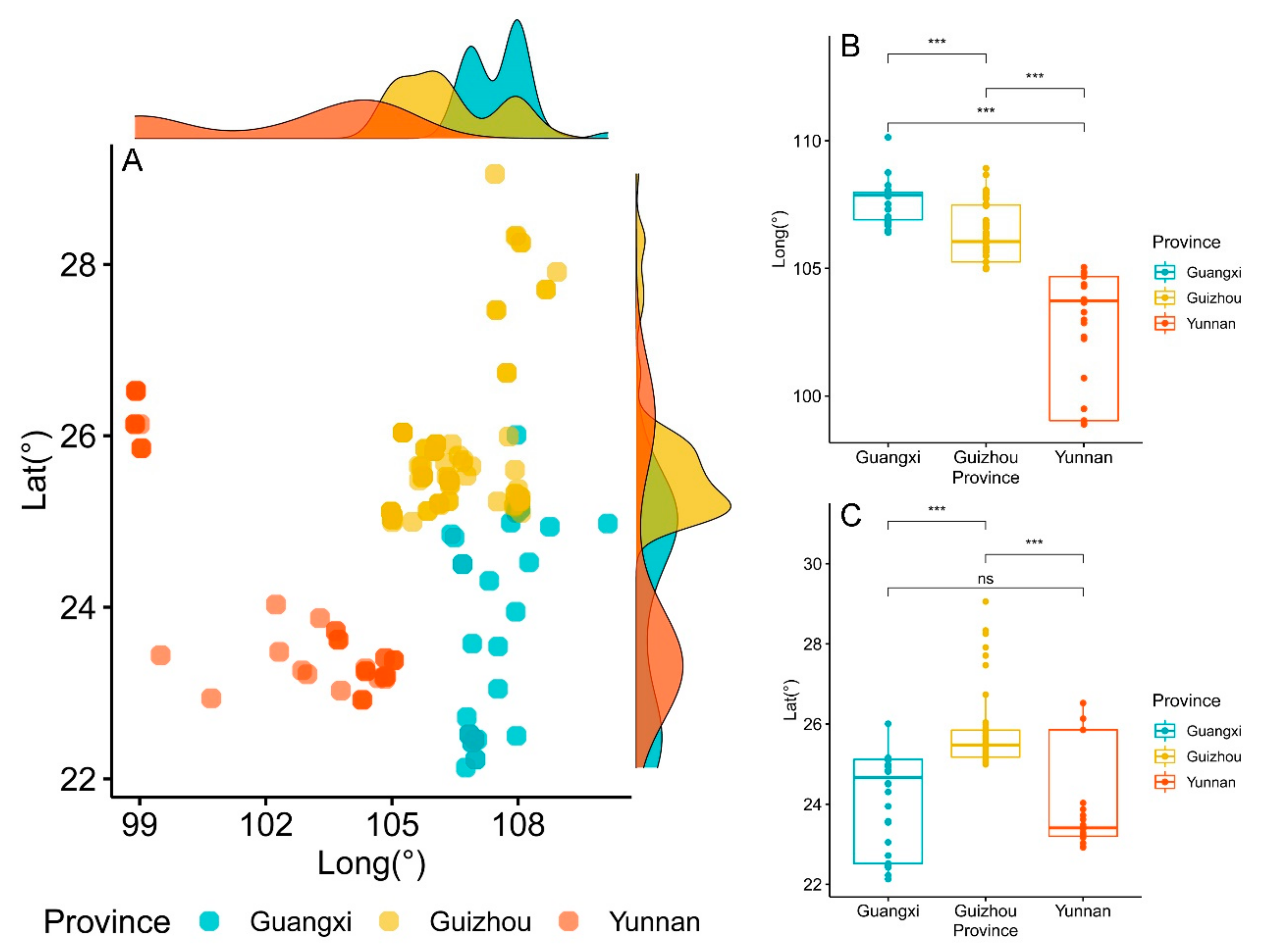
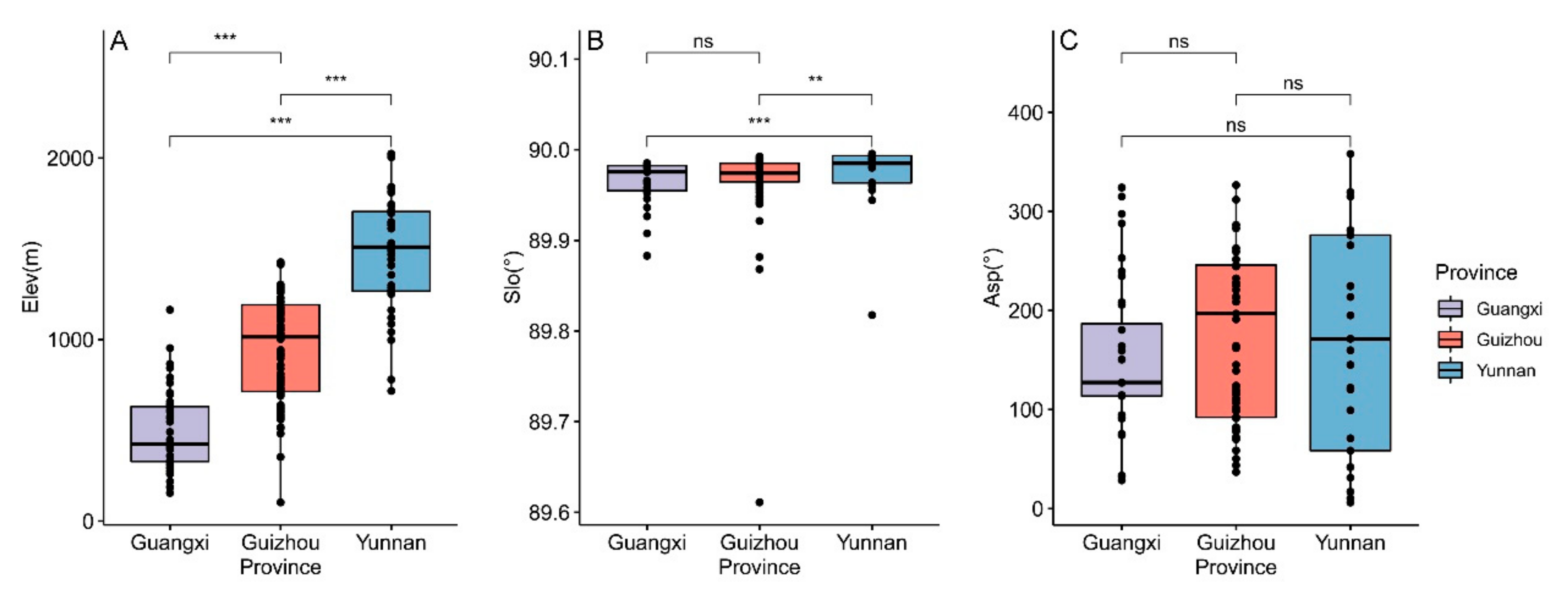
3.2. Differences in Geographical Distribution and Growth Environment of Brachypetalum Species
3.2.1. Differences in Topographic Factors
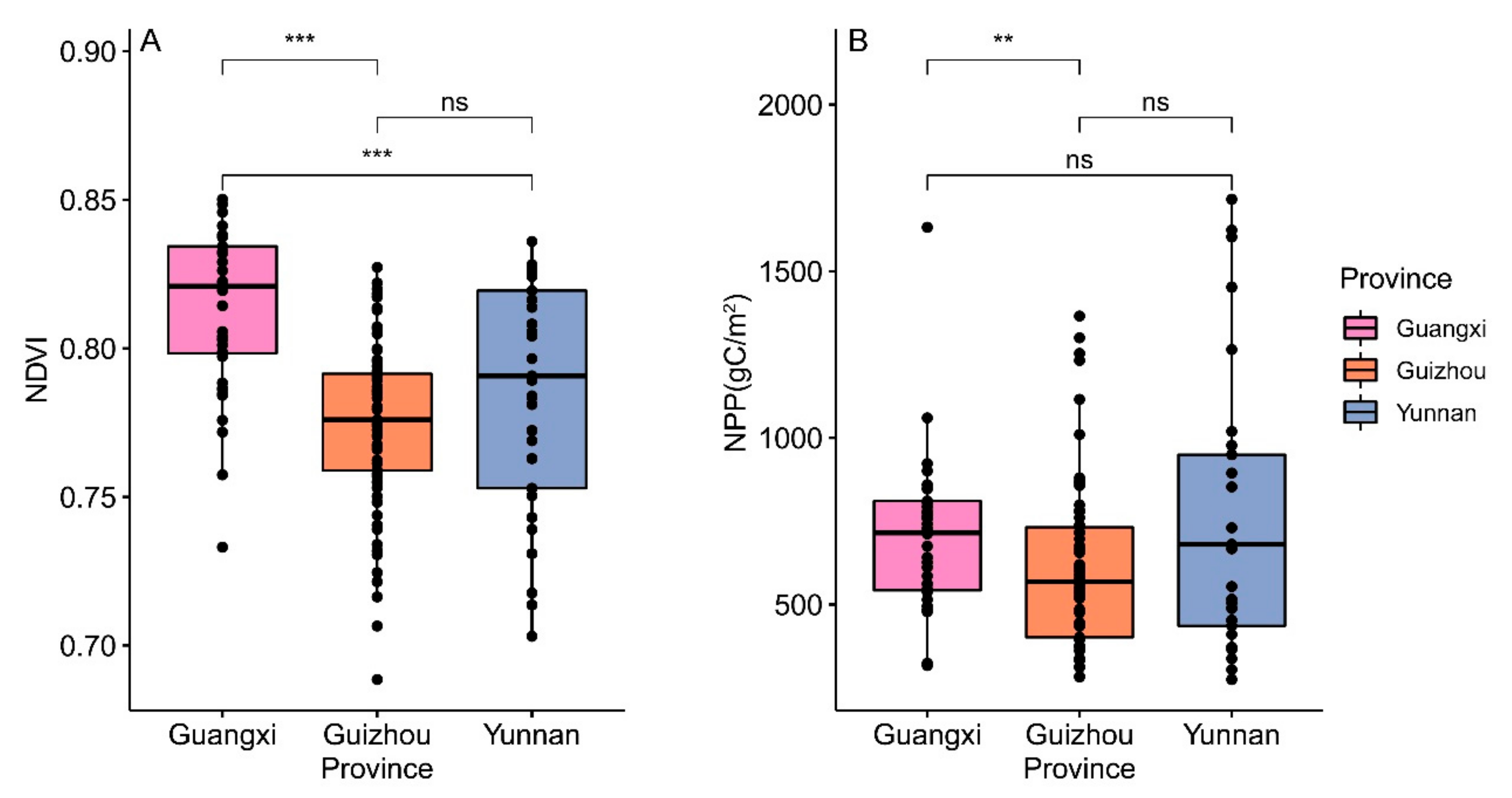
3.2.2. Differences in Productive Factors
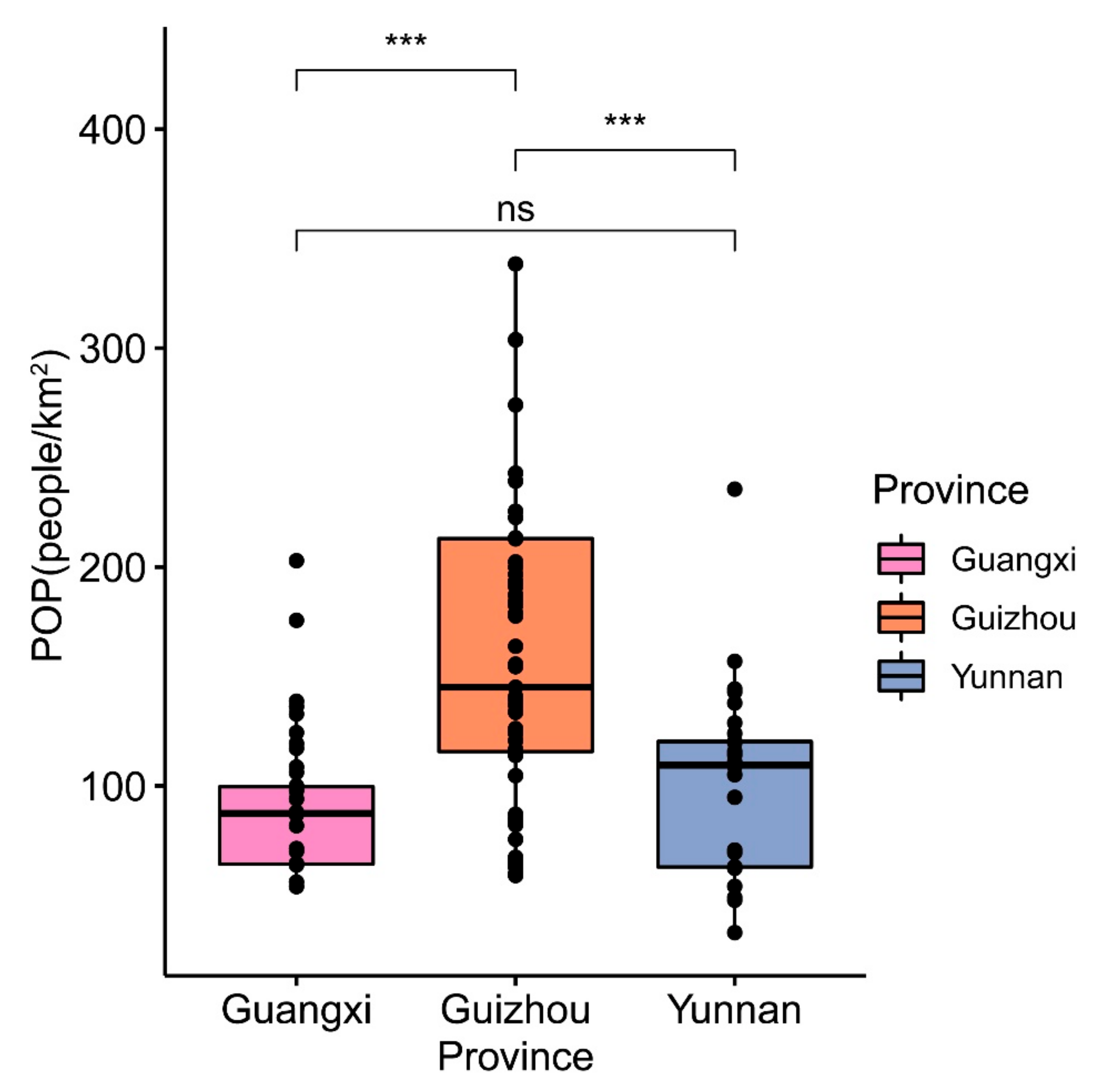
3.2.3. Differences in Human Interference Factors
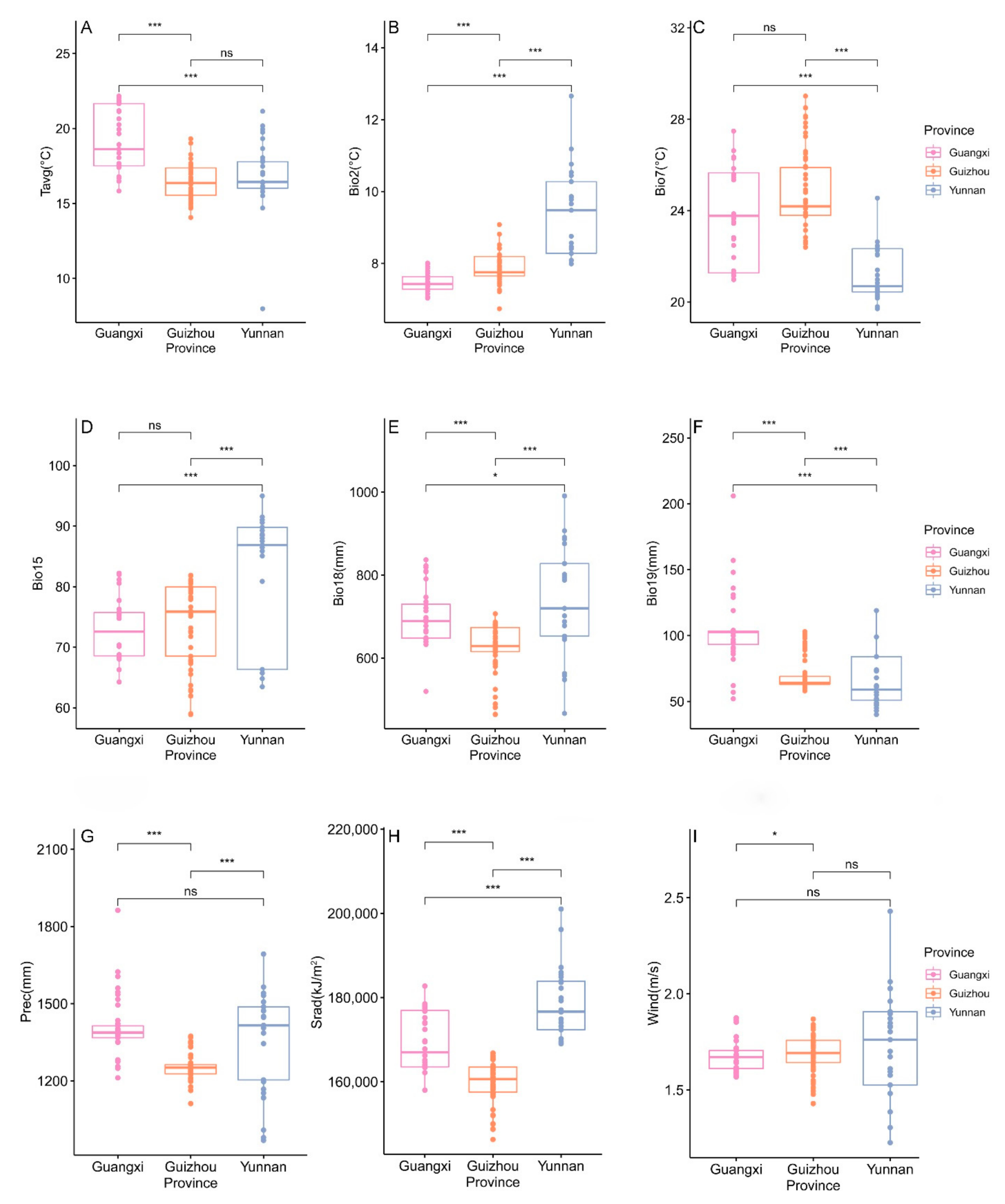
3.2.4. Differences in Climatic Factors
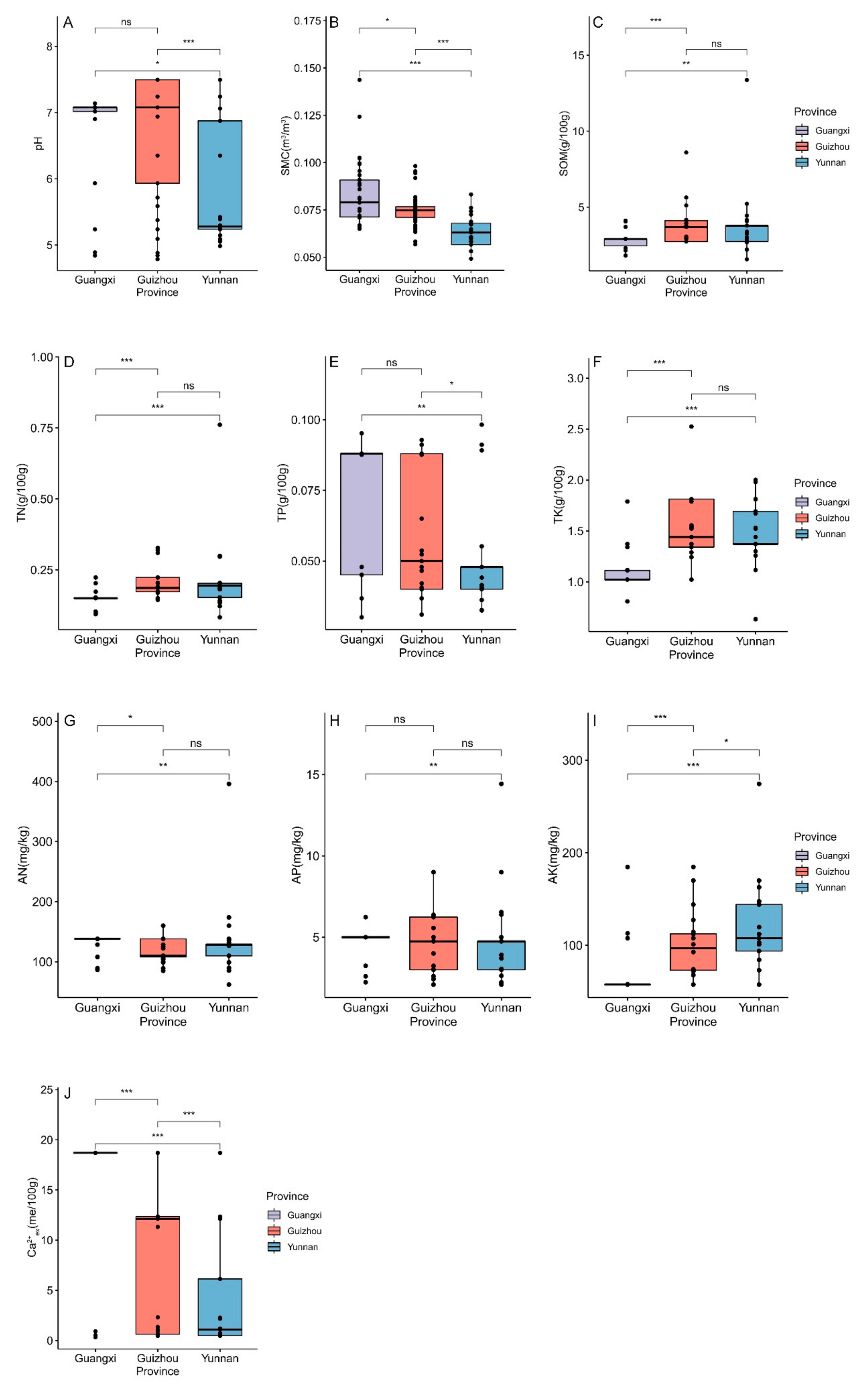
3.2.5. Differences in Edaphic Factors
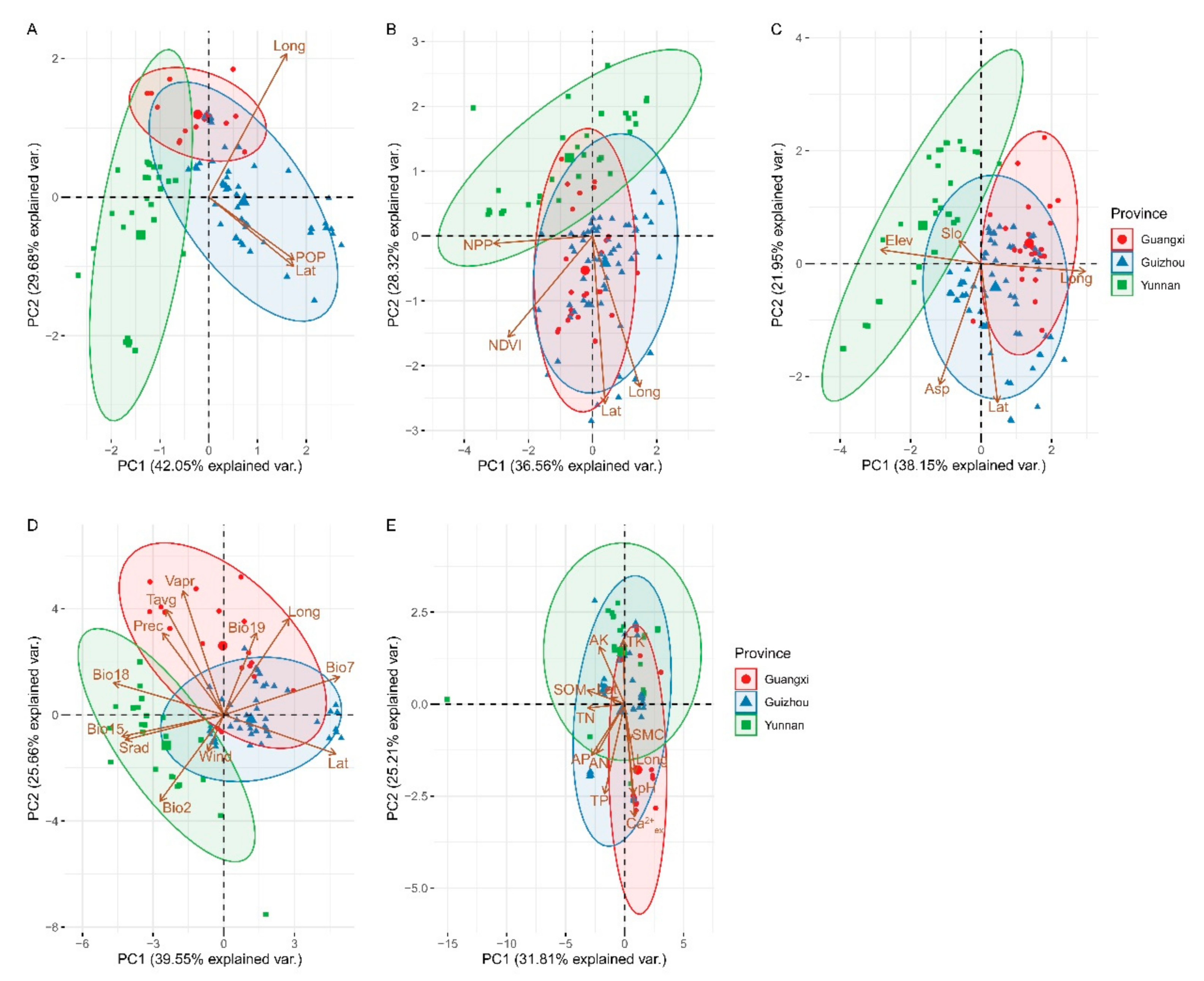
3.3. Environmental Factors Related to Distribution Patterns of Brachypetalum Species
4. Discussion
4.1. Distribution Patterns of Brachypetalum Orchids in Southwest China
4.2. Relationships between Environmental Factors and the Distribution of Brachypetalum Orchids in Southwest China
4.3. Conservation of Brachypetalum Orchids in Southwest China
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niu, Z.T.; Xue, Q.Y.; Zhu, S.Y.; Sun, J.; Liu, W.; Ding, X.Y. The complete plastome sequences of four orchid species: Insights into the evolution of the orchidaceae and the utility of plastomic mutational hotspots. Front. Plant Sci. 2017, 8, 715. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.J.; Yan, Y.J.; Tian, Y.; Li, J.S.; He, J.S.; Tang, Z.Y. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Gaskett, A.C.; Gallagher, R.V. Orchid diversity: Spatial and climatic patterns from herbarium records. Ecol. Evol. 2018, 8, 11235–11245. [Google Scholar] [CrossRef] [Green Version]
- Cribb, P.J.; Kell, S.P.; Dixon, K.W.; Barrett, R.L. Orchid conservation: A global perspective. In Orchid Conservation; Natural History Publications: Kota Kinabalu, Malaysia, 2003; pp. 1–24. [Google Scholar]
- Guo, Z.L.; Wang, L.F. The horizontal distribution pattern of Orchidaceae in China along latitude and longitude. J. Biol. 2013, 30, 49–53. [Google Scholar]
- Shi, X.W.; Zhang, L.; Zhang, J.J.; Ouyang, Z.Y.; Xiao, Y. Priority area of biodiversity conservation in Southwest China. Chin. J. Ecol. 2018, 37, 3721–3728. [Google Scholar]
- Medhi, R.P.; Chakrabarti, S. Traditional knowledge of NE people on conservation of wild orchids. Indian J. Tradit. Knowl. 2009, 8, 11–16. [Google Scholar]
- Liu, H.; Jacquemyn, H.; He, X.Y.; Chen, W.; Huang, Y.Q.; Yu, S.; Lu, Y.P.; Zhang, Y. The impact of human pressure and climate change on the habitat availability and protection of Cypripedium (Orchidaceae) in Northeast China. Plants 2021, 10, 84. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Kumaria, S.; Job, N.; Tandon, P. En-masse production of elite clones of Dendrobium crepidatum: A threatened, medicinal orchid used in Traditional Chinese Medicine (TCM). J. Appl. Res. Med. Aromat. Plants 2016, 3, 168–176. [Google Scholar] [CrossRef]
- Zhang, Q.; Qi, C.; Wang, H.; Xiao, X.F.; Zhuang, Y.; Gu, S.J.; Zhou, Y.S.; Wang, L.; Yang, H.J.; Xu, W.L. Biocompatible and degradable Bletilla striata polysaccharide hemostasis sponges constructed from natural medicinal herb Bletilla striata. Carbohydr. Polym. 2019, 226, 115304. [Google Scholar] [CrossRef]
- Pillon, Y.; Chase, M. Taxonomic exaggeration and its effects on orchid conservation. Conserv. Biol. 2006, 21, 263–265. [Google Scholar] [CrossRef]
- Zhang, F.P.; Huang, J.L.; Zhang, S.B. Trait evolution in the slipper orchid Paphiopedilum (Orchidaceae) in China. Plant Signal. Behav. 2016, 11, e1149668. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.J.; Chen, S.C.; Phillip, J.C. Flora of China; Science Press: Beijing, China, 2009; Volume 25, pp. 33–44. [Google Scholar]
- Li, Z.Y.; Wu, Y.L.; Peng, K. Micro-morphological characters of leaf epidermis of ten species in genus Paphiopedilum. Bull. Bot. Res. 2014, 34, 723–729. [Google Scholar]
- Shi, J.Z.; Chen, H.; An, M.T.; Zhang, Y.; Ye, C.; Wu, J.Y. Analyses on distribution characteristics and protection effect of wild Paphiopedilum in Guizhou Province. Guihaia 2021. Available online: https://kns.cnki.net/kcms/detail/45.1134.Q.20210324.1433.012.html (accessed on 29 July 2021).
- Guan, Z.J.; Zhang, S.B.; Guan, K.Y.; Li, S.Y.; Hu, H. Leaf anatomical structures of Paphiopedilum and Cypripedium and their adaptive significance. J. Plant Res. 2011, 124, 289–298. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, Z.L.; Li, S.Y.; Hu, H.; Huang, J.L. Mycorrhizal specificity, preference, and plasticity of six slipper orchids from South Western China. Mycorrhiza 2010, 20, 559–568. [Google Scholar] [CrossRef]
- Evonne, T. Conversion of RAPD Markers to Co-Dominant Based Sequence Characterized Amplified Region (SCAR) Markers in Three Paphiopedilum Species; Universiti Malaysia Sabah: Kota Kinabalu, Malaysia, 2007; pp. 1–2. [Google Scholar]
- Lee, Y.I. The asymbiotic seed germination of six Paphiopedilum species in relation to the time of seed collection and seed pretreatment. Acta Hortic. 2007, 755, 381–386. [Google Scholar] [CrossRef]
- Chochai, A.; Leitch, I.J.; Ingrouille, M.J.; Fay, M.F. Molecular phylogenetics of Paphiopedilum (Cypripedioideae; Orchidaceae) based on nuclear ribosomal ITS and plastid sequences. Bot. J. Linnean Soc. 2012, 170, 176–196. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.C.; Liao, P.C.; Ko, Y.Z.; Chen, C.H.; Chiang, Y.C. Phylogeny and historical biogeography of Paphiopedilum Pfitzer (Orchidaceae) based on nuclear and plastid DNA. Front. Plant Sci. 2020, 11, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.J.; Zhu, G.F.; Lü, F.B.; Zhang, X.; Wang, B.Q. Studies on the karyotypes of eight species of Paphiopedilum subgenus Brachypetalum. Acta Hortic. Sin. 2006, 33, 1015–1020. [Google Scholar]
- Liu, Z.J.; Chen, S.C. Paphiopedilum angustatum, a new orchid from Yunnan, China. Acta Phytotaxon. Sin. 2000, 38, 464–466. [Google Scholar]
- Liu, Z.J.; Zhang, J.Y. Paphiopedilum singchii sp. nov., an addition to the subgenus Brachypetalum of Paphiopedilum (Orchidaceae). Acta Phytotaxon. Sin. 2000, 38, 467–470. [Google Scholar]
- Juiling, S.; Leon, S.K.; Jumian, J.; Tsen, S.; Lee, Y.L.; Khoo, E.; Sugau, J.B.; Nilus, R.; Pereira, J.T.; Damit, A.; et al. Conservation assessment and spatial distribution of endemic orchids in Sabah, Borneo. Nat. Conserv. Res. 2020, 5, 136–144. [Google Scholar] [CrossRef]
- Štípková, Z.; Kindlmann, P. Orchid extinction over the last 150 years in the Czech Republic. Diversity 2021, 13, 78. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Lievens, B.; Wiegand, T. Spatial variation in below-ground seed germination and divergent mycorrhizal associations correlate with spatial segregation of three co-occurring orchid species. J. Ecol. 2012, 100, 1328–1337. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Rajaram, M.C.; Yong, C.S.Y.; Go, R. The differences between Paphiopedilum barbatum (Lindl.) and Papiopedilum callosum (Rchb.f.) Stein var. sublaeve (Rchb.f.) P.J. Cribb. In Proceedings of the Malaysia International Biology Symposium 2016, Putrajaya, Malaysia, 26–27 October 2016; p. 26. [Google Scholar]
- Liu, Z.J.; Liu, K.W.; Chen, L.J.; Lei, S.P.; Li, L.Q.; Shi, X.C.; Huang, L.Q. Conservation ecology of endangered species Paphiopedilum armeniacum (Orchidaceae). Acta Ecol. Sin. 2006, 26, 2791–2800. [Google Scholar] [CrossRef]
- Wang, M.N.; Li, S.Z.; Chen, L.J.; Li, J.; Li, L.Q.; Rao, W.H.; Liu, H.; Chen, J.B.; Ren, H. Conservation and reintroduction of the rare and endangered orchid Paphiopedilum armeniacum. Ecosyst. Health Sustain. 2021, 7, 1903817. [Google Scholar] [CrossRef]
- Li, Z.Y.; Li, M.Y. Study on phenotypic variation of Paphiopedilum micranthum population. J. Plant Genet. Resour. 2015, 16, 765–771. [Google Scholar]
- National Forestry and Grassland Administration, Ministry of Agriculture and Rural Affairs of the People’s Republic of China. The National Key Protected Wild Plants List. 2021. Available online: http://www.gov.cn/zhengce/zhengceku/2021-09/09/content_5636409.htm (accessed on 23 September 2021).
- Wang, B.S.; Mao, J.F.; Zhao, W.; Wang, X.R. Impact of geography and climate on the genetic differentiation of the subtropical pine Pinus yunnanensis. PLoS ONE 2013, 8, e67345. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, G.H.; He, J.X.; Yu, G.C. Factors about shallow biogenic gas reservoir and evidence of its migration and accumulation in the Qujing basin. Nat. Gas Geosci. 2007, 18, 673–677. [Google Scholar]
- Zhang, Y.; Shi, X.Z.; Yu, D.S.; Wang, H.J.; Zhao, Y.C.; Sun, W.X. Effects of the linkage between spatial data and attribute data on estimates of soil organic carbon. Adv. Earth Sci. 2008, 23, 840–847. [Google Scholar]
- Du, H.; Hu, F.; Zeng, F.P.; Wang, K.L.; Peng, W.X.; Zhang, Z.X.; Song, T.Q. Spatial distribution of tree species in evergreen-deciduous broadleaf karst forests in southwest China. Sci. Rep. 2017, 7, 15664. [Google Scholar] [CrossRef] [Green Version]
- França, S.; Cabral, H.N. Predicting fish species distribution in estuaries: Influence of species’ ecology in model accuracy. Estuar. Coast. Shelf Sci. 2016, 180, 11–20. [Google Scholar] [CrossRef]
- Ye, P.C.; Zhang, G.F.; Zhao, X.; Chen, H.; Si, Q.; Wu, J.Y. Potential geographical distribution and environmental explanations of rare and endangered plant species through combined modelling: A case study of Northwest Yunnan, China. Ecol. Evol. 2021, 11, 13052–13067. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Xu, X.L. China Population Spatial Distribution Kilometer Grid Dataset; Data Registration and Publishing System of Resource and Environmental Science Data Center, Chinese Academy of Sciences: Beijing, China, 2017. [Google Scholar] [CrossRef]
- Xu, X.L. China Annual Normalized Difference Vegetation Index (NDVI) Spatial Distribution Dataset; Data Registration and Publishing System of Resource and Environmental Science Data Center, Chinese Academy of Sciences: Beijing, China, 2018. [Google Scholar] [CrossRef]
- Mao, K. SMC Dataset: Soil Moisture in China Dataset (2002–2018); National Tibetan Plateau Data Center: Beijing, China, 2021. [Google Scholar] [CrossRef]
- Meng, X.; Mao, K.; Meng, F.; Shi, J.; Zeng, J.; Shen, X.; Cui, Y.; Jiang, L.; Guo, Z. A fine-resolution soil moisture dataset for China in 2002–2018. Earth Syst. Sci. Data 2021, 13, 3239–3261. [Google Scholar] [CrossRef]
- Shangguan, W.; Dai, Y.J.; Liu, B.Y.; Zhu, A.X.; Duan, Q.Y.; Wu, L.Z.; Ji, D.Y.; Ye, A.Z.; Yuan, H.; Zhang, Q.; et al. A China dataset of soil properties for land surface modeling. J. Adv. Model. Earth Syst. 2013, 5, 212–224. [Google Scholar] [CrossRef]
- Dai, Y.; Shangguan, W. Dataset of Soil Properties for Land Surface Modeling over China; National Tibetan Plateau Data Center: Beijing, China, 2019. [Google Scholar] [CrossRef]
- LaMorte, W.W. Mann Whitney U Test (Wilcoxon Rank Sum Test). Boston University School of Public Health, 4 May 2017. Available online: https://sphweb.bumc.bu.edu/otlt/MPH-Modules/BS/BS704_Nonparametric/BS704_Nonparametric4.html (accessed on 28 September 2021).
- Kassambara, A. R Package ‘Ggpubr’ Version 0.4.0: ‘Ggplot2’ Based Publication Ready Plots. 2020. Available online: https://cran.r-project.org/web/packages/ggpubr/ggpubr.pdf (accessed on 25 August 2021).
- Ding, C.; He, X.F. K-means clustering via principal component analysis. In Proceedings of the Twenty-First International Conference on Machine Learning ACM, Banff, AB, Canada, 4–8 July 2004. [Google Scholar]
- Kassambara, A. Practical Guide to Principal Component Methods in R. 2017. Available online: https://www.bibsonomy.org/bibtex/25f79c00df4524ae5b9455539b6b00c0f/asalber (accessed on 22 November 2021).
- Tsiftsis, S.; Tsiripidis, I.; Karagiannakidou, V.; Alifragis, D. Niche analysis and conservation of the orchids of east Macedonia (NE Greece). Acta Oecol. Int. J. Ecol. 2008, 33, 27–35. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Du, H.D.; Jin, X.H.; Ma, K.P. Species diversity and geographic distribution of wild Orchidaceae in China. Chin. Sci. Bull. 2015, 60, 179–188. [Google Scholar]
- Tang, Z.; Wang, Z.; Zheng, C.; Fang, J. Biodiversity in China’s mountains. Front. Ecol. Environ. 2006, 4, 347–352. [Google Scholar] [CrossRef]
- Wittlinger, L.; Petrikovičová, L. Phytogeographical analysis and ecological factors of the distribution of Orchidaceae taxa in the Western Carpathians (Local study). Plants 2021, 10, 588. [Google Scholar] [CrossRef]
- Ye, P.C.; Zhang, G.F.; Wu, J.Y. Hotspots and conservation gaps: A case study of key higher plant species from Northwest Yunnan, China. Glob. Ecol. Conserv. 2020, 23, e01005. [Google Scholar] [CrossRef]
- Bell, T.J.; Bowles, M.L.; Zettler, L.W.; Pollack, C.A.; Ibberson, J.E. Environmental and management effects on demographic processes in the U.S. threatened Platanthera leucophaea (Nutt.) Lindl. (Orchidaceae). Plants 2021, 10, 1308. [Google Scholar] [CrossRef]
- Chapagain, D.J.; Meilby, H.; Baniya, C.B.; Budha-Magar, S.; Ghimire, S.K. Illegal harvesting and livestock grazing threaten the endangered orchid Dactylorhiza hatagirea (D. Don) Soó in Nepalese Himalaya. Ecol. Evol. 2021, 11, 6672–6687. [Google Scholar] [CrossRef]
- Pettorelli, N.; Ryan, S.; Mueller, T.; Bunnefeld, N.; Jędrzejewska, B.; Lima, M.; Kausrud, K. The Normalized Difference Vegetation Index (NDVI): Unforeseen successes in animal ecology. Clim. Res. 2011, 46, 15–27. [Google Scholar] [CrossRef]
- DeLucia, E.H.; Gomez-Casanovas, N.; Greenberg, J.A.; Hudiburg, T.W.; Kantola, I.B.; Long, S.P.; Miller, A.D.; Ort, D.R.; Parton, W.J. The theoretical limit to plant productivity. Environ. Sci. Technol. 2014, 48, 9471–9477. [Google Scholar] [CrossRef]
- Johnson, S.D.; Morita, S. Lying to Pinocchio: Floral deception in an orchid pollinated by long-proboscid flies. Bot. J. Linnean Soc. 2010, 152, 271–278. [Google Scholar] [CrossRef]
- Pansarin, E.R.; Pansarin, L.M. Reproductive biology of Trichocentrum pumilum: An orchid pollinated by oil-collecting bees. Plant Biol. 2011, 13, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.M.; Toft, R.J.; Gaskett, A.C. Pollination and insect visitors to the putatively brood-site deceptive endemic spurred helmet orchid, Corybas cheesemanii. N. Z. J. Bot. 2013, 51, 155–167. [Google Scholar] [CrossRef]
- Ding, Z.F.; Liang, J.C.; Hu, Y.M.; Zhou, Z.X.; Sun, H.B.; Liu, L.N.; Liu, H.J.; Hu, H.J.; Si, X.F. Different responses of avian feeding guilds to spatial and environmental factors across an elevation gradient in the central Himalaya. Ecol. Evol. 2019, 9, 4116–4128. [Google Scholar] [CrossRef] [Green Version]
- Tsiftsis, S.; Štípková, Z.; Kindlmann, P. Role of way of life, latitude, elevation and climate on the richness and distribution of orchid species. Biodivers. Conserv. 2019, 28, 75–96. [Google Scholar] [CrossRef]
- Michael, F.; Chase, M.W. Orchid biology: From Linnaeus via Darwin to the 21st century. Ann. Bot. 2009, 104, 359–364. [Google Scholar]
- He, Y.L.; Li, X.Z.; Craft, C.; Ma, Z.G.; Sun, Y.G. Relationships between vegetation zonation and environmental factors in newly formed tidal marshes of the Yangtze River estuary. Wetl. Ecol. Manag. 2011, 19, 341–349. [Google Scholar] [CrossRef]
- Diez, J.M. Hierarchical patterns of symbiotic orchid germination linked to adult proximity and environmental gradients. J. Ecol. 2007, 95, 159–170. [Google Scholar] [CrossRef]
- Figura, T.; Weiser, M.; Ponert, J. Orchid seed sensitivity to nitrate reflects habitat preferences and soil nitrate content. Plant Biol. 2020, 22, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Hrivnák, M.; Slezák, M.; Galvánek, D.; Vlčko, J.; Belanová, E.; Rízová, V.; Senko, D.; Hrivnák, R. Species richness, ecology, and prediction of orchids in Central Europe: Local-scale study. Diversity 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Brewbaker, J.L.; Kwack, B.H. The essential role of calcium ion in pollen germination and pollen tube growth. Am. J. Bot. 1963, 50, 859–865. [Google Scholar] [CrossRef]
- Izuddin, M.; Yam, T.W.; Webb, E.L. Germination niches and seed persistence of tropical epiphytic orchids in an urban landscape. J. Plant Res. 2019, 132, 383–394. [Google Scholar] [CrossRef]
- Štípková, Z.; Kindlmann, P. Factors determining the distribution of orchids–A review with examples from the Czech Republic. Eur. J. Environ. Sci. 2021, 11, 21–30. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Red List Level | Protected Level | CITES Appendix | PSESP |
|---|---|---|---|---|
| Paphiopedilum micranthum | CR | II | I | NO |
| Paphiopedilum emersonii | CR | I | I | YES |
| Paphiopedilum bellatulum | EN | I | I | NO |
| Paphiopedilum malipoense | EN | I | I | NO |
| Paphiopedilum concolor | EN | I | I | NO |
| Paphiopedilum wenshanense | CR | I | I | YES |
| Paphiopedilum armeniacum | EN | I | I | YES |
| Classification | Variable (Unit) | Abbreviation | Resolution | Time Period | Data Source | Reference |
|---|---|---|---|---|---|---|
| Geographic | Longitude (°) | Long | None | None | Field investigation | None |
| Geographic | Latitude (°) | Lat | None | None | Field investigation | None |
| Topographic | Elevation (m) | Elev | 2.5 min | None | WorldClim version 2.1 climate data | [40] |
| Topographic | Slope (°) | Slo | 2.5 min | None | Convert from elevation data | None |
| Topographic | Aspect (°) | Asp | 2.5 min | None | Convert from elevation data | None |
| Climate | Average Temperature (°C) | Tavg | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Precipitation (mm) | Prec | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Solar Radiation (kJ/m2) | Srad | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Wind Speed (m/s) | Wind | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Mean Diurnal Range (℃) | Bio2 | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Annual Temperature Range (℃) | Bio7 | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Precipitation Seasonality | Bio15 | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Precipitation of the Warmest Quarter (mm) | Bio18 | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Climate | Precipitation of Coldest Quarter (mm) | Bio19 | 2.5 min | 1970–2000 | WorldClim version 2.1 climate data | [40] |
| Anthropogenic | Population Density (people/km2) | POP | 1 km2 | 1990–2015 | Resource and Environment Science and Data Center | [41] |
| Productivity | Normalized Difference Vegetation Index | NDVI | 1 km2 | 1998–2019 | Resource and Environment Science and Data Center | [42] |
| Productivity | Net Primary Production (gC/m2) | NPP | 1 km2 | 2000–2010 | Resource and Environment Science and Data Center | None |
| Soil | Soil Moisture (m3/m3) | SMC | 0.05 degrees | 2002–2018 | National Tibetan Plateau Data Center | [43,44] |
| Soil | pH Value | pH | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Soil Organic Matter (g/100 g) | SOM | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Total N (g/100 g) | TN | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Total P (g/100 g) | TP | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Total K (g/100 g) | TK | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Alkali-hydrolysable N (mg/kg) | AN | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Available P (mg/kg) | AP | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Available K (mg/kg) | AK | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
| Soil | Exchangeable Ca2+ (me/100 g) | Ca2+ex | 30 s | None | National Tibetan Plateau Data Center | [45,46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, P.; Wu, J.; An, M.; Chen, H.; Zhao, X.; Jin, X.; Si, Q. Geographical Distribution and Relationship with Environmental Factors of Paphiopedilum Subgenus Brachypetalum Hallier (Orchidaceae) Taxa in Southwest China. Diversity 2021, 13, 634. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120634
Ye P, Wu J, An M, Chen H, Zhao X, Jin X, Si Q. Geographical Distribution and Relationship with Environmental Factors of Paphiopedilum Subgenus Brachypetalum Hallier (Orchidaceae) Taxa in Southwest China. Diversity. 2021; 13(12):634. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120634
Chicago/Turabian StyleYe, Pengcheng, Jianyong Wu, Mingtai An, Hui Chen, Xiao Zhao, Xiaohua Jin, and Qin Si. 2021. "Geographical Distribution and Relationship with Environmental Factors of Paphiopedilum Subgenus Brachypetalum Hallier (Orchidaceae) Taxa in Southwest China" Diversity 13, no. 12: 634. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120634