
Genetic Diversity of Oxytropis Species from the Center of the Genus Origin: Insight from Molecular Studies

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
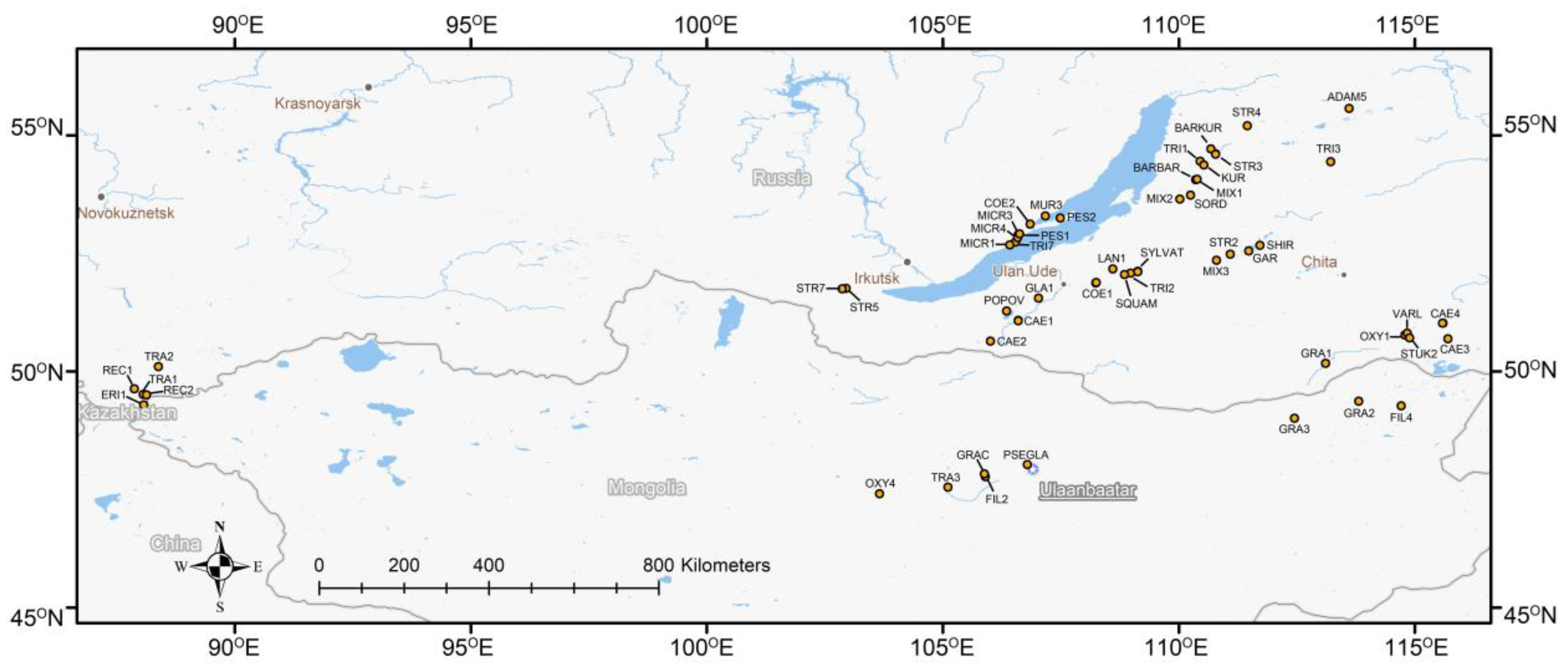
2.1. Taxon Sampling, DNA Extraction, and Sequencing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
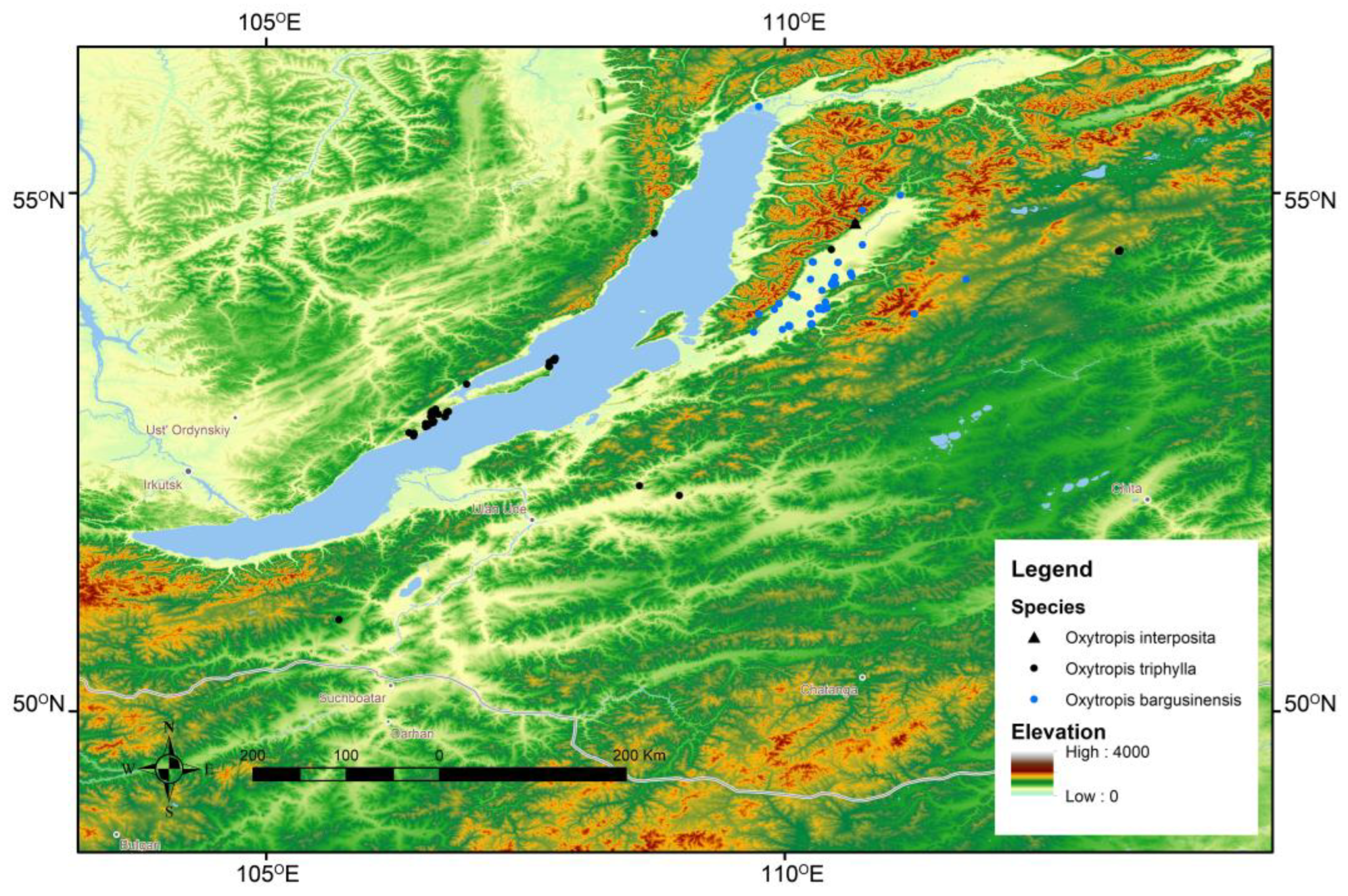{kind=link}
| Subgenus, Section, Species [Reference], 2n [Reference], The Location of the Population (No. of Samples) | Latitude, Longitude | Population Code | Genetic Diversity (SD) | Genbank Accession Numbers | |||
|---|---|---|---|---|---|---|---|
| Haplotype Diversity | Nucleotide Diversity | Psba-Trnh | Trnl-Trnf | Trns-Trng | |||
| Subgenus Tragacanthoxytropis Vass. Section Hystrix Bunge | |||||||
| O. tragacanthoides Fisch. ex DC. [26], 2n = 16, 32 [15] | |||||||
| 1. Russia, Altai Mts., Ukok Plateau, Zhumaly River (10) | 49.51°, 88.06° | TRA1 | 0.867 (0.107) | 0.0048 (0.0027) | MW172222-MW172234 | MW177548-MW177560 | MW177535-MW177547 |
| 2. Russia, Altai Mts., near the village Chagan-Uzun (5) | 50.10°, 88.38° | TRA2 | 0.700 (0.218) | 0.0008 (0.0006) | |||
| 3. Mongolia, Central Aimag, near the Öndörshireet sum (5) | 47.55°, 105.11° | TRA3 | 0.800 (0.164) | 0.0005 (0.0004) | |||
| Subgenus Phacoxytropis Bunge Section Janthina Bunge | |||||||
| O. coerulea (Pall.) DC. [26], 2n = 32 [36] | |||||||
| 1. Russia, Buryatia, the village Zaigraevo (5) | 51.87°, 108.24° | COE1 | 0.900 (0.161) | 0.0017 (0.0012) | LR898256-LR898269 | LR898302-LR898315 | LR898413-LR898426 |
| 2. Russia, Irkutsk Region, near the village Sarma (12) | 53.12°, 106.85° | COE2 | 0.970 (0.044) | 0.0046 (0.0025) | |||
| O. filiformis DC. [26], 2n = 16 [15] | |||||||
| 1. Mongolia, Central Aimag, near the Argalant sum (5) | 47.77°, 105.90° | FIL2 | 1.000 (0.126) | 0.0048 (0.0031) | LR898271, LR898272, LR898275-LR898279, LR898281-LR898283 | LR898317, LR898318, LR898321-LR898325, LR898327-LR898329 | LR898428, LR898429, LR898432-LR898436, LR898438-LR898440 |
| 2. Mongolia, Eastern Aimag, near the Gurvanzagal sum (9) | 49.27°, 114.71° | FIL4 | 0.722 (0.159) | 0.0012 (0.0008) | |||
| Section Mesogaea Bunge | |||||||
| O. glabra (Lam.) DC. [26], 2n = 16 [15] | |||||||
| 1. Russia, Buryatia, the village Orongoy (6) | 51.55°, 107.03° | GLA1 | 0.333 (0.215) | 0.0001 (0.0002) | LR898299, LR898300 | LR898345, LR898346 | LR898456, LR898457 |
| Subgenus Oxytropis ex genere Oxytropis DC. Section Orobia Bunge | |||||||
| O. adamsiana (Trautv.) Jurtzev [29], 2n = 32 [15] | |||||||
| 1. Russia, Buryatia, the village Uakit (10) | 55.57°, 113.61° | ADAM5 | 0.956 (0.059) | 0.0047 (0.0027) | OV260617-OV260624 | OV260717-OV260724 | OV260806-OV260813 |
| O. popoviana Peschkova, 2n = 16, 32 [36] | |||||||
| 1. Russia, Buryatia, the village Baraty (5) | 51.28°, 106.35° | POPOV | 0.800 (0.164) | 0.0009 (0.0007) | ON567429-ON567431 | ON567410-ON567412 | ON567391-ON567393 |
| O. recognita Bunge, 2n = 32 [15] | |||||||
| 1. Russia, Altai Mts., Jazator River (10) | 49.63°, 87.87° | REC1 | 0.822 (0.097) | 0.0059 (0.0033) | ON567432-ON567443 | ON567413-ON567424 | ON567394-ON567405 |
| 2. Russia, Altai Mts., Ukok Plateau (8) | 49.50°, 88.13° | REC2 | 1.000 (0.062) | 0.0020 (0.0013) | |||
| O. sordida (Willd.) Pers. [25], 2n = 48, 64 [15] | |||||||
| 1. Russia, Buryatia, the village Ina (8) | 53.73°, 110.25° | SORD | 0.786 (0.151) | 0.0008 (0.0005) | LS991870-LS991874 | LS991896-LS991900 | LS991922-LS991926 |
| O. strobilacea Bunge [29], 2n = 16, 32, 48, 64 [15] | |||||||
| 1. Russia, Buryatia, the village Zaigraevo (5) | 51.88°, 108.25° | STR1 | 0.800 (0.164) | 0.0009 (0.0007) | OV260579-OV260605, OV260608-OV260614 | OV260679-OV260705, OV260708-OV260714 | OV260768-OV260794, OV260797-OV260803 |
| 2. Russia, Buryatia, the village Komsomol’skoe (10) | 52.48°, 111.09° | STR2 | 0.867 (0.107) | 0.0016 (0.0010) | |||
| 3. Russia, Buryatia, the village Maisk (15) | 54.61°, 110.78° | STR3 | 0.952 (0.040) | 0.0032 (0.0018) | |||
| 4. Russia, Buryatia, tract Ukshaki (5) | 55.20°, 111.45° | STR4 | 1.000 (0.127) | 0.0042 (0.0027) | |||
| 5. Russia, Buryatia, the village Tory (6) | 51.76°, 102.95° | STR5 | 0.600 (0.215) | 0.0013 (0.0009) | |||
| 6. Russia, Buryatia, the village Zun-Murino (11) | 51.75°, 102.87° | STR7 | 0.964 (0.051) | 0.0025 (0.0015) | |||
| O. sylvatica (Pall.) DC. [22], 2n = 16, 32 [15] | |||||||
| 1. Russia, Buryatia, the village Udinsk (12) | 52.12°, 109.13° | SYLVAT | 0.818 (0.084) | 0.0008 (0.0006) | LT856560-LT856562 | LT856573-LT856575 | LT856586-LT856588 |
| Section Xerobia Bunge | |||||||
| O. caespitosa (Pall.) Pers. [27], 2n = 48 [37], 64 [15] | |||||||
| 1. Russia, Buryatia, near the village Novoselenginsk (11) | 51.08°, 106.60° | CAE1 | 0.327 (0.153) | 0.0001 (0.0001) | LR861115-LR861122 | LR828424-LR828431 | LR828468-LR828475 |
| 2. Russia, Buryatia, near the village Dyrestuy (9) | 50.64°, 106.01° | CAE2 | 0.000 (0.000) | 0.0000 (0.0000) | |||
| 3. Russia, Transbaikalia, near the village Kusocha (5) | 50.69°, 115.70° | CAE3 | 0.700 (0.218) | 0.0030 (0.0020) | |||
| 4. Russia, Transbaikalia, near the village Tsugol (5) | 51.02°, 115.60° | CAE4 | 0.700 (0.218) | 0.0017 (0.0012) | |||
| O. eriocarpa Bunge [27], 2n = 16 [15], 24 + 2–3B [38] | |||||||
| 1. Russia, Altai Mts., Ukok Plateau, Zhumaly River (7) | 49.29°, 88.07° | ERI1 | 0.893 (0.111) | 0.0090 (0.0051) | LR861754-LR861759 | LR861762-LR861767 | LR861770-LR861775 |
| O. grandiflora (Pall.) DC. [27], 2n = 48 [39] | |||||||
| 1. Russia, Transbaikalia, near the village Bytev (5) | 50.17°, 113.11° | GRA1 | 0.900 (0.161) | 0.0046 (0.0029) | LR861123-LR861131 | LR828432-LR828440 | LR828476-LR828484 |
| 2. Mongolia, Dornod Province, near the Bayandun sum (7) | 49.37°, 113.81° | GRA2 | 0.857 (0.137) | 0.0035 (0.0021) | |||
| 3. Mongolia, Dornod Province, near the Bayan-Uul sum (9) | 49.01°, 112.45° | GRA3 | 0.667 (0.132) | 0.0043 (0.0024) | |||
| O. mixotriche Bunge [27], 2n—no data | |||||||
| 1. Russia, Buryatia, near the village Urzhil (5) | 54.07°, 110.39° | MIX1 | 0.400 (0.237) | 0.0002 (0.0002) | LR861724 -LR861733 | LR861734 -LR861743 | LR861744 -LR861753 |
| 2. Russia, Buryatia, near the village Suvo (7) | 53.65°, 110.02° | MIX2 | 0.851 (0.137) | 0.0053 (0.0031) | |||
| 3. Russia, Buryatia, near the village Mozhayka (5) | 52.35°, 110.80° | MIX3 | 1.000 (0.126) | 0.0019 (0.0013) | |||
| O. peschkovae M. Pop. [27], 2n = 48 [37] | |||||||
| 1. Russia, Irkutsk Region, near the Gyzgi-Nur Lake (9) | 52.91°, 106.63° | PES1 | 0.972 (0.064) | 0.0082 (0.0046) | LR861691-LR861701 | LR861702-LR861712 | LR861713-LR861723 |
| 2. Russia, Irkutsk Region, Olkhon Island, near the village Khuzhir (11) | 53.25°, 107.49° | PES2 | 0.345 (0.172) | 0.0016 (0.0009) | |||
| O. triphylla (Pall.) DC. [27], 2n = 16 [36,40] | |||||||
| 1. Russia, Buryatia, near the village Sakhuli (12) | 54.45°, 110.45° | TRI1 | 0.924 (0.057) | 0.0018 (0.0011) | LR861134-LR861149, LR861154-LR861158 | LR828443-LR828458, LR828463-LR828467 | LR828487-LR828502, LR828507-LR828511 |
| 2. Russia, Buryatia, near the village Udinsk (6) | 52.08°, 108.98° | TRI2 | 0.800 (0.172) | 0.0022 (0.0014) | |||
| 3. Russia, Buryatia, near the village Bagdarin (10) | 54.44°, 113.22° | TRI3 | 0.844 (0.103) | 0.0021 (0.0013) | |||
| 4. Russia, Irkutsk Region, Begul Bay (6) | 52.74°, 106.54° | TRI7 | 1.000 (0.096) | 0.0025 (0.0016) | |||
| Section Polyadena Bunge | |||||||
| O. glandulosa Turcz. [23], 2n = 32 [15] | |||||||
| 1. Russia, Buryatia, near the village Argada (10) | 54.37°, 110.53° | KUR | 0.911 (0.077) | 0.0059 (0.0033) | LT732668-LT732678 | LT732657-LT732667 | LT732646-LT732656 |
| 2. Russia, Buryatia, near the village Urzhil (14) | 54.07°, 110.39° | BAR | 0.703 (0.101) | 0.0036 (0.0020) | |||
| 3. Russia, Buryatia, near the village Shiringa (15) | 52.67°, 111.72° | SHIR | 0.133 (0.112) | 0.0002 (0.0002) | |||
| 4. Russia, Buryatia, near the village Garam (10) | 52.55°, 111.48° | GAR | 0.356 (0.159) | 0.0006 (0.0004) | |||
| O. microphylla (Pall.) DC. [28], 2n—no data | |||||||
| 1. Russia, Irkutsk Region, near the village Ozera (9) | 52.68°, 106.42° | MICR1 | 0.000 (0.000) | 0.0000 (0.0000) | MN199988-MN199990 | MN200000-MN200003 | MN200014-MN200016 |
| 2. Russia, Irkutsk Region, near the Gyzgi-Nur Lake (6) | 52.91°, 106.63° | MICR2 | 0.600 (0.215) | 0.0007 (0.0005) | |||
| 3. Russia, Irkutsk Region, near the Gurbi-Nur Lake (11) | 52.89°, 106.60° | MICR3 | 0.182 (0.144) | 0.0001 (0.0001) | |||
| 4. Russia, Irkutsk Region, near the Namish-Nur Lake (11) | 52.83°, 106.59° | MICR4 | 0.182 (0.144) | 0.0001 (0.0001) | |||
| O. muricata (Pall.) DC. [28], 2n = 16, 32 [15] | |||||||
| 1. Russia, Irkutsk Region, near the Gyzgi-Nur Lake (9) | 52.91°, 106.63° | MUR1 | 0.417 (0.191) | 0.0009 (0.0006) | MN199983-MN199987 | MN199996-MN200000 | MN200009-MN200013 |
| 2. Russia, Irkutsk Region, near the village Sarma (8) | 53.12°, 106.85° | MUR2 | 0.250 (0.180) | 0.0007 (0.0005) | |||
| 3. Russia, Irkutsk Region, near the Oto-Khushun Cape (13) | 53.29°, 107.17° | MUR3 | 0.692 (0.075) | 0.0014 (0.0008) | |||
| O. pseudoglandulosa Gontsch. ex Grub. [28], 2n—no data | |||||||
| 1. Mongolia, near the Ulan-Bator (7) | 48.03°, 106.79° | PSEGLA | 0.809 (0.130) | 0.0011 (0.0007) | MN199991-MN199994 | MN200004-MN200007 | MN200017-MN200020 |
| O. varlakovii Serg. [22,28], 2n = 24 + 1B [40] | |||||||
| 1. Russia, Transbaikalia, near the Nozhii Lake (5) | 50.80°, 114.83° | VARL | 0.700 (0.218) | 0.0003 (0.0003) | LT856563-LT856565 | LT856576-LT856578 | LT856589-LT856591 |
| Section Leucopodia Bunge | |||||||
| O. squammulosa DC., 2n = 16 [38] | |||||||
| 1. Russia, Buryatia, Uda River (5) | 52.05°, 108.85° | SQUAM | 0.900 (0.161) | 0.0022 (0.0015) | ON567444-ON567447 | ON567425-ON567428 | ON567406-ON567409 |
| Section Verticillares DC. | |||||||
| O. bargusinensis Peschkova [22], 2n—no data | |||||||
| 1. Russia, Buryatia, Alla River (15) | 54.71°, 110.68° | BARKUR | 0.762 (0.096) | 0.0011 (0.0007) | LT856473-LT856488 | LT856506-LT856521 | LT856539-LT856554 |
| 2. Russia, Buryatia, near the village Urzhil (15) | 54.06°, 110.36° | BARBAR | 0.895 (0.070) | 0.0016 (0.0009) | |||
| O. gracillima Bunge [24], 2n—no data | |||||||
| 1. Mongolia, Central Aimag, near the Argalant sum (5) | 47.83°, 105.88° | GRAC | 1.000 (0.126) | 0.0042 (0.0027) | MH174938, LT996058-LT996061 | LT996062-LT996066 | LT996067-LT996071 |
| O. interposita Sipl. [22], 2n = 24 [40] | |||||||
| 1. Russia, Buryatia, Alla River (14) | 54.71°, 110.68° | INTKUR | 0.769 (0.075) | 0.0015 (0.0009) | LT856489-LT856493 | LT856522-LT856526 | LT856555-LT856559 |
| O. lanata (Pall.) DC. [24], 2n = 16 [15] | |||||||
| 1. Russia, Buryatia, near the village Unegetei (20) | 52.17°, 108.60° | LAN1 | 0.895 (0.051) | 0.0010 (0.0006) | LT994841-LT994858 | LT994895-LT994912 | LT994949-LT994966 |
| 2. Russia, Buryatia, near the village Urzhil (15) | 54.07°, 110.39° | LAN2 | 0.895 (0.052) | 0.0010 (0.0006) | |||
| 3. Mongolia, Central Aimag, near the Argalant sum (5) | 47.83°, 105.88° | LAN3 | 0.900 (0.161) | 0.0017 (0.0012) | |||
| O. myriophylla (Pall.) DC [24], 2n = 16 [15] | |||||||
| 1. Russia, Transbaikalia, near the village Tsugol (5) | 51.02°, 115.59° | MYR1 | 0.700 (0.218) | 0.0003 (0.0003) | LT994859-LT994866 | LT994913-LT994920 | LT996002-LT996009 |
| 2. Russia, Buryatia, near the village Udinsk (21) | 52.11°, 109.13° | MYR2 | 0.767 (0.057) | 0.0007 (0.0005) | |||
| 3. Russia, Buryatia, the village Komsomol’skoe (8) | 52.48°, 111.09° | MYR3 | 0.821 (0.101) | 0.0008 (0.0005) | |||
| O. oxyphylla (Pall.) DC. [24], 2n = 16 [15] | |||||||
| 1. Russia, Transbaikalia, near the Nozhii Lake (5) | 50.77°, 114.79° | OXY1 | 0.900 (0.161) | 0.0007 (0.0005) | LT994867-LT994891 | LT994921-LT994945 | LT996010-LT996034 |
| 2. Russia, Buryatia, near the village Novoselenginsk (16) | 51.07°, 106.60° | OXY2 | 0.958 (0.036) | 0.0023 (0.0013) | |||
| 3. Russia, Buryatia, the village Komsomol’skoe (8) | 52.48°, 111.09° | OXY3 | 0.964 (0.077) | 0.0045 (0.0026) | |||
| 4. Mongolia, Bulgan Aimag, near the Rashaant sum (4) | 47.41°, 103.66° | OXY4 | 1.000 (0.177) | 0.0042 (0.0029) | |||
| O. selengensis Bunge [24], 2n = 32 [15] | |||||||
| 1. Russia, Buryatia, near the village Unegetei (18) | 52.17°, 108.60° | SEL | 0.634 (0.127) | 0.0009 (0.0006) | MH174937, LT996052-LT996057 | LT996045-LT996051 | LT996038-LT996044 |
| O. stukovii Palibin [24], 2n = 16 [40] | |||||||
| 1. Russia, Transbaikalia, near the Nozhii Lake (4) | 50.81°, 114.84° | STUK1 | 0.833 (0.222) | 0.0005 (0.0005) | LT994892-LT994894 | LT994946-LT994948 | LT996035-LT996037 |
| 2. Russia, Transbaikalia, near the Kunkur Lake (10) | 50.71°, 114.89° | STUK2 | 0.711 (0.117) | 0.0004 (0.0003) | |||
| O. tompudae M. Pop. [22], 2n—no data | |||||||
| 1. Russia, Buryatia, the village Maisk (8) | 54.60°, 110.78° | TOMP | 0.893 (0.111) | 0.0026 (0.0016) | LT856566-LT856571 | LT856579-LT856584 | LT856592-LT856597 |
2.2. Sequence Alignment and Phylogenetic Analyses
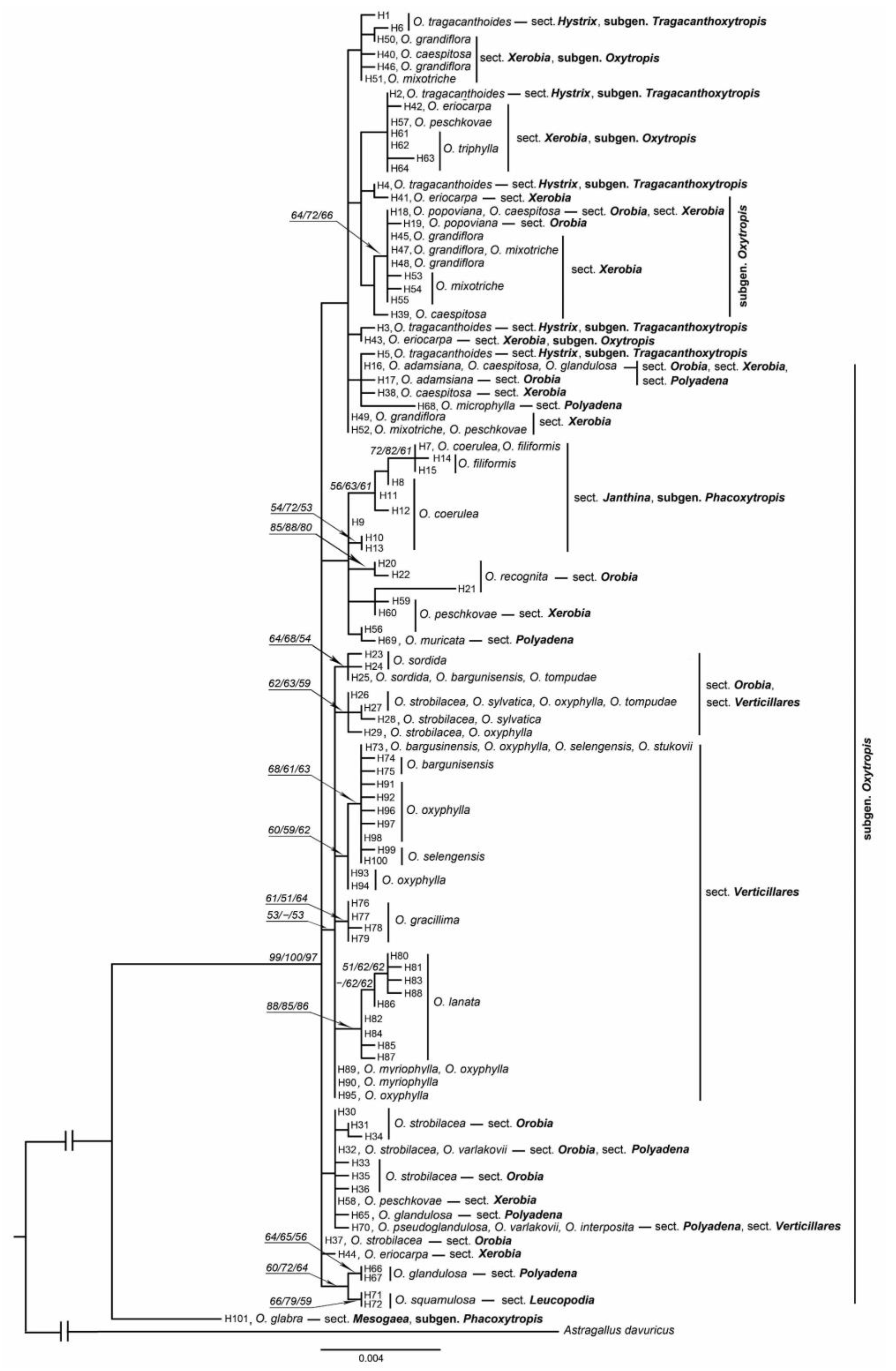
3. Results
4. Discussion
4.1. Morphological Differences among Oxytropis Species
4.2. Genetic Diversity of the Populations of Oxytropis Species
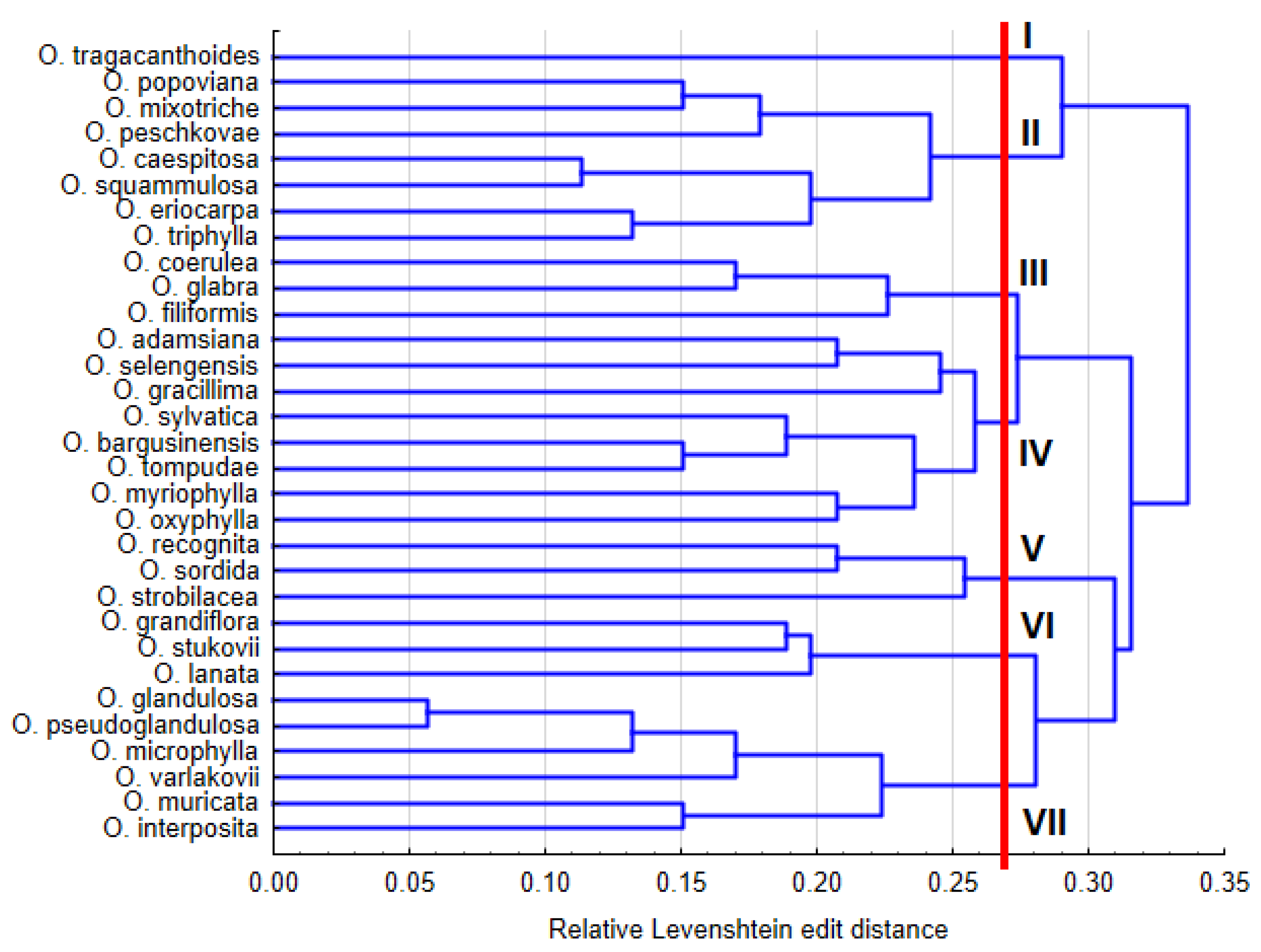
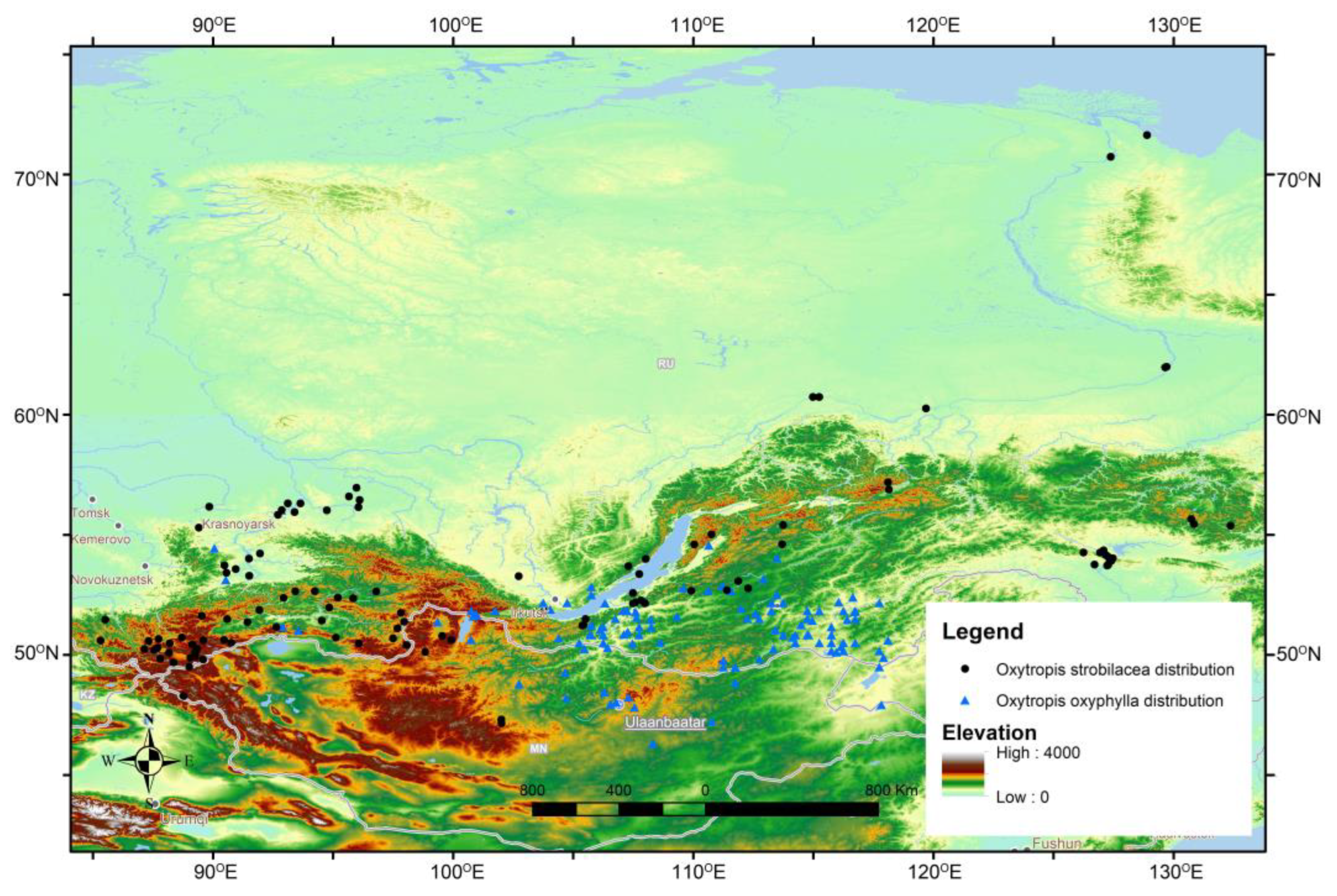
4.3. Phylogenetic Relationships of Oxytropis Species within Southern Siberia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, X.; Welsh, S.L.; Ohashi, H. Oxytropis. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2010; Volume 10, pp. 453–500. [Google Scholar]
- Malyshev, L.I. Phenetics of the Subgenera and Sections in the Genus Oxytropis DC. (Fabaceae) Bearing on Ecology and Phylogeny. Contemp. Probl. Ecol. 2008, 1, 440–444. [Google Scholar] [CrossRef]
- Polozhii, A.V. On the Problem of the Origin and Evolution of the Genus Oxytropis (Fabaceae). Bot. Zhurnal 2003, 88, 55–59. (In Russian) [Google Scholar]
- Shavvon, R.S.; Kazempour-Osaloo, S.; Maassoumi, A.A.; Moharrek, F.; Karaman Erkul, S.; Lemmon, A.; Lemmon, E.M.; Michalak, I.; Muellner-Riehl, A.N.; Favre, A. Increasing Phylogenetic Support for Explosively Radiating Taxa: The Promise of High-throughput Sequencing for Oxytropis (Fabaceae). J. Syst. Evol. 2017, 55, 385–404. [Google Scholar] [CrossRef]
- Kas’yanova, L.N.; Azovskii, M.G. Vegetation of Dune Sands of Olkhon Island on Baikal and Some Questions Concerning its Protection. Geogr. Nat. Res. 2011, 32, 248–253. [Google Scholar]
- Boikov, T.G.; Sutkin, A.V. Ecophytocenotic Features of Vicia tsydenii Malysch. in Southern Transbaikalia. Rus. J. Ecol. 2012, 5, 340–346. [Google Scholar]
- Plennik, R.Y. Morphologicheskaya Evolutsia Bobovyh Yugo-Vostochnogo Altaya [Morphological Evolution of Legumes from the South-Eastern Altai]; Nauka: Novosibirsk, Russia, 1976. (In Russian) [Google Scholar]
- Blinova, K.F.; Sakanyan, E.I. Species of Oxytropis DC. Used in Tibetan Medicine and Their Flavonoid Composition. Rast. Resur. 1986, 22, 266–272. (In Russian) [Google Scholar]
- Amirkhanova, A.S.; Ustenova, G.O. Review of the Current Status of Study Oxytropis. Asian J. Pharm. Clin. Res. 2018, 11, 50–55. [Google Scholar]
- Povydysh, M.N.; Bobyleva, N.S.; Bityukova, N.V. Genus 29. Oxytropis DC. In Plant Resources of Russia: Wild Flowering Plants, Their Component Composition, and Biological Activity; Budantsev, A.L., Ed.; KMK, SPb.: Moscow, Russia, 2010; Volume 3, pp. 65–69. (In Russian) [Google Scholar]
- Li, M.X.; Lan, Z.H.; Wei, L.L.; Zhang, W.J.; Zhang, R.X.; Jia, Z.P. Phytochemical and Biological Studies of Plants from the Genus Oxytropis. Rec. Nat. Prod. 2012, 6, 1–20. [Google Scholar]
- Polozhii, A.V. Oxytropis DC. In Flora Sibiriae. Fabaceae (Leguminosae); Polozhii, A.V., Malyshev, L.I., Eds.; Nauka: Novosibirsk, Russia, 1994; Volume 9, pp. 74–151. (In Russian) [Google Scholar]
- Peshkova, G.A. Florogeneticheskiy Analiz Stepnoy Floryi Gor Yuzhnoy Sibiri [Florogenetic Analysis of the Steppe Flora of the South Siberia]; Nauka: Novosibirsk, Russia, 2001. (In Russian) [Google Scholar]
- Polozhii, A.V. Oxytropis (Fabaceae) Species in the Mountain Flora of South Siberia. Bot. Zhurnal 1995, 80, 58–66. (In Russian) [Google Scholar]
- Malyshev, L.I. Diversity of the Genus Oxytropis in the Asian part of Russia. Turczaninowia 2008, 11, 5–141. (In Russian) [Google Scholar]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly Variable Chloroplast Markers for Evaluating Plant Phylogeny at Low Taxonomic Levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar]
- Kress, W.J. Plant DNA Barcodes: Applications Today and in the Future. J. Syst. Evol. 2017, 55, 291–307. [Google Scholar]
- Shneyer, V.S.; Rodionov, A.V. Plant DNA Barcodes. Biol. Bull. Rev. 2019, 9, 295–300. [Google Scholar]
- Shaw, J.; Lickey, E.B.; Beck, J.T.; Farmer, S.B.; Liu, W.; Miller, J.; Siripun, K.C.; Winder, C.T.; Schilling, E.E.; Small, R.L. The Tortoise and the Hare II: Relative Utility of 21 Noncoding Chloroplast DNA Sequences for Phylogenetic Analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar]
- Kim, S.-C.; Crawford, D.J.; Jansen, R.K.; Santos-Guerra, A. The Use of a Non-coding Region of Chloroplast DNA in Phylogenetic Studies of the Subtribe Sonchinae (Asteraceae: Lactuceae). Plant Syst. Evol. 1999, 215, 85–99. [Google Scholar]
- Tekpinar, A.; Karaman Erkul, S.; Aytac, Z.; Kaya, Z. Phylogenetic Relationships among Native Oxytropis Species in Turkey Using trnL intron, trnL–F IGS, and trnV intron cpDNA Regions. Turk. J. Bot. 2016, 40, 472–479. [Google Scholar]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Sandanov, D.V. Modern State of Populations of Endemic Oxytropis Species from Baikal Siberia and Their Phylogenetic Relationships Based on Chloroplast DNA Markers. Russ. J. Genet. 2018, 54, 805–815. [Google Scholar] [CrossRef]
- Kholina, A.; Kozyrenko, M.; Artyukova, E.; Sandanov, D.; Selyutina, I.; Chimitov, D. Plastid DNA Variation of the Endemic Species Oxytropis glandulosa Turcz. (Fabaceae). Turk. J. Bot. 2018, 42, 38–50. [Google Scholar] [CrossRef]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Sandanov, D.V. The Divergence of Oxytropis Species of Section Verticillares (Fabaceae) of the Steppe Flora of Baikal Siberia Based on Chloroplast DNA Sequence Data. Russ. J. Genet. 2019, 55, 701–710. [Google Scholar] [CrossRef]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Andrianova, E.A.; Sandanov, D.V.; Selyutina, I.Y.; Khoreva, M.G. Genetic Diversity and Phylogenetic Relationships of Oxytropis evenorum (Fabaceae) Based on Data of Sequencing of the Intergenic Spacers of the Chloroplast DNA. Bull. NESC 2019, 2, 117–125. (In Russian) [Google Scholar]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Koldaeva, M.N.; Sandanov, D.V.; Selyutina, I.Y. Phylogenetic Relationships of the Species of Asian Russia of the Subgenera Phacoxytropis and Tragacanthoxytropis Genus Oxytropis Based on the Polymorphism of Markers of the Chloroplast and Nuclear Genomes. Russ. J. Genet. 2021, 57, 1042–1056. [Google Scholar]
- Kholina, A.; Kozyrenko, M.; Artyukova, E.; Sandanov, D.; Selyutina, I. Genetic Diversity of Oxytropis Section Xerobia (Fabaceae) in One of the Centres of Speciation. Genetica 2021, 149, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Pozdnyakova, T.E. Variability of Chloroplast DNA of Oxytropis Section Polyadena (Fabaceae) in Asian Russia: Population Analysis and Phylogenetic Relationships. Biol. Bull. 2021, 48, 16–25. [Google Scholar] [CrossRef]
- Kholina, A.B.; Artyukova, E.V.; Yakubov, V.V.; Khoreva, M.G.; Mochalova, O.A.; Sandanov, D.V.; Selyutina, I.Y. Genetic Divergence of Closely Related Species Oxytropis strobilacea, O. adamsiana, O. vassilczenkoi of the series Strobilacei of the section Orobia (Fabaceae) from Asian Russia. Biol. Bull. 2023, 50, 15–25. [Google Scholar]
- Sandanov, D.V.; Dugarova, A.S.; Brianskaia, E.P.; Selyutina, I.Y.; Makunina, N.I.; Dudov, S.V.; Chepinoga, V.V.; Wang, Z. Diversity and Distribution of Oxytropis DC. (Fabaceae) Species in Asian Russia. Biodivers. Data J. 2022, 10, e78666. [Google Scholar]
- Ulziikhutag, N. Legumes of Mongolia (Taxonomy, Ecology, Geography, Phylogeny and Economic Value); Ulanbaatar, Mongolia, 2003. (In Russian) [Google Scholar]
- Levenshtein, V.I. Binary Code Capable of Correcting Deletions, Insertions and Reversals. Sov. Phys.-Dokl. Cybern. Control Theory 1966, 10, 707–710. [Google Scholar]
- Kohonen, T. Self-Organizing Maps, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Anenkhonov, O.A.; Korolyuk, A.Y.; Sandanov, D.V.; Liu, H.; Zverev, A.A.; Guo, D. Soil-moisture Conditions Indicated by Field-layer Plants Help Identify Vulnerable Forests in the Forest-steppe of Semi-arid Southern Siberia. Ecol. Indic. 2015, 57, 196–207. [Google Scholar]
- Zverev, A.A. Methodological Aspects of Indicator Values Use in Biodiversity Analysis. Contemp. Probl. Ecol. 2020, 13, 321–332. [Google Scholar] [CrossRef]
- Krivenko, D.A.; Kotseruba, V.V.; Kazanovsky, S.G.; Verkhozina, A.V.; Stepanov, A.V. Fabaceae. IAPT/IOPB Chromosome Data 11. Taxon 2011, 60, 1222. [Google Scholar]
- Krivenko, D.A.; Kazanovsky, S.G.; Vinogradova, Y.K.; Verkhozina, A.V.; Knyazev, M.S.; Murtazaliev, R.A. Fabaceae (Leguminosae). IAPT/IOPB Chromosome Data 26. Taxon 2017, 66, 1492. [Google Scholar]
- An’kova, T.V.; Shaulo, D.N. Fabaceae. IAPT/IOPB Chromosome Data 14. Taxon 2012, 61, 1336. [Google Scholar]
- Krivenko, D.A.; Kazanovsky, S.G.; Verkhozina, A.V.; Chernova, O.D.; Dymshakova, O.S.; Turskaya, A.L. Fabaceae. IAPT/IOPB Chromosome Data 15. Taxon 2013, 62, 1077. [Google Scholar]
- Konichenko, E.S.; Selyutina, I.Y.; Dorogina, O.V. Fabaceae. IAPT/IOPB Chromosome Data 14. Taxon 2012, 61, 1338–1339. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal Primers for Amplification of Three Non-coding Regions of Chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar]
- Bonfeld, J.K.; Smith, K.F.; Staden, R. A New DNA Sequence Assembly Program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods): Version 4.04; Sinauer Associates Inc.: Sunderland, MA, USA, 2003. [Google Scholar]
- Posada, D.; Crandall, K.A. Modeltest: Testing the Model of DNA Substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar]
- Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Sandanov, D.V.; Andrianova, E.A. Phylogenetic Relationships of the Species of Oxytropis DC. subg. Oxytropis and Phacoxytropis (Fabaceae) from Asian Russia Inferred from the Nucleotide Sequence Analysis of the Intergenic Spacers of the Chloroplast Genome. Russ. J. Genet. 2016, 52, 780–793. [Google Scholar]
- Chimitov, D.G. Taxonomic Diversity of the Genus Oxytropis Section Polyadena (Fabaceae) in Baikal Siberia. Novit. Syst. Plant. Vasc. 2022, 53, 43–57. (In Russian) [Google Scholar] [CrossRef]
- Selyutina, I.Y.; Sandanov, D.V.; Chimitov, D.G. Demographic Structure Analysis of Rare Species Oxytropis glandulosa Bunge (Fabaceae) in Republic of Buryatia. BSU Bull. Biol. Geography 2016, 2–3, 168–172. (In Russian) [Google Scholar]
- Selyutina, I.Y.; Sandanov, D.V. Demographic Structure and Population Size of Rare Endemic Oxytropis species from Priol’khonye Steppe. Rastit. Mir Aziat. Ross. 2018, 1, 14–23. (In Russian) [Google Scholar]
- Kholina, A.; Kozyrenko, M.; Artyukova, E.; Yakubov, V.; Khoreva, M.; Andrianova, E.; Mochalova, O.; Sandanov, D. Phylogenetic Relationships of Oxytropis Section Gloeocephala from Northeast Asia Based on Sequencing of the Intergenic Spacers of cpDNA and ITS nrDNA. Genetica 2022, 150, 117–128. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W. Effects of Life History Traits on Genetic Diversity in Plant Species. Philos. Trans. R. Soc. Lond. B 1996, 351, 1291–1298. [Google Scholar]
- Gitzendanner, M.A.; Soltis, P.S. Patterns of Genetic Variation in Rare and Widespread Plant Congeners. Am. J. Bot. 2000, 87, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Nybom, H. Comparison of Different Nuclear DNA Markers for Estimating Intraspecific Genetic Diversity in Plants. Mol. Ecol. 2004, 137, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, Y.G.S.; Vitorino, L.C.; de Souza, U.J.B.; Bessa, L.A. Recent Trends in Research on the Genetic Diversity of Plants: Implications for Conservation. Diversity 2019, 11, 62. [Google Scholar] [CrossRef]
- Blanco-Pastor, J.L.; Fernández-Mazuecos, M.; Coello, A.J.; Pastor, J.; Vargas, P. Topography Explains the Distribution of Genetic Diversity in One of the Most Fragile European Hotspots. Divers. Distrib. 2019, 25, 74–89. [Google Scholar] [CrossRef]
- De Kort, H.; Prunier, J.G.; Ducatez, S.; Honnay, O.; Baguette, M.; Stevens, V.M.; Blanchet, S. Life History, Climate and Biogeography Interactively Affect Worldwide Genetic Diversity of Plant and Animal Populations. Nat. Commun. 2021, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Wambulwa, M.C.; Luo, Y.-H.; Zhu, G.-F.; Milne, R.; Wachira, F.N.; Wu, Z.-Y.; Wang, H.; Gao, L.-M.; Li, D.-Z.; Liu, J. Determinants of Genetic Structure in a Highly Heterogeneous Landscape in Southwest China. Front. Plant Sci. 2022, 13, 779989. [Google Scholar] [CrossRef]
- Plenk, K.; Willner, W.; Demina, O.N.; Höhn, M.; Kuzemko, A.; Vassilev, K.; Kropf, M. Phylogeographic Evidence for Long-term Persistence of the Eurasian steppe Plant Astragalus onobrychis in the Pannonian Region (Eastern Central Europe). Flora 2020, 264, 151555. [Google Scholar] [CrossRef]
- Shepherd, L.D.; Lange, P.J.; Perrie, L.R.; Heenan, P.B. Chloroplast Phylogeography of New Zealand Sophora trees (Fabaceae): Extensive Hybridization and Widespread Last Glacial Maximum Survival. J. Biogeogr. 2017, 44, 1640–1651. [Google Scholar] [CrossRef]
- Wang, H.; Liu, P.-L.; Li, J.; Yang, H.; Li, Q.; Chang, Z.-Y. Why More Leaflets? The Role of Natural Selection in Shaping the Spatial Pattern of Leaf-shape Variation in Oxytropis diversifolia (Fabaceae) and Two Close Relatives. Front. Plant Sci. 2021, 12, 681962. [Google Scholar] [CrossRef]
- Plenk, K.; Bardy, K.; Höhn, M.; Thiv, M.; Kropf, M. No Obvious Genetic Erosion, but Evident Relict Status at the Westernmost Range Edge of the Pontic-Pannonian Steppe Plant Linum flavum L. (Linaceae) in Central Europe. Ecol. Evol. 2017, 7, 6527–6539. [Google Scholar] [CrossRef]
- Zheng, H.-Y.; Guo, X.-L.; Price, M.; He, X.-J.; Zhou, S.-D. Effects of Mountain Uplift and Climatic Oscillations on Phylogeography and Species Divergence of Chamaesium (Apiaceae). Front. Plant Sci. 2021, 12, 673200. [Google Scholar] [CrossRef]
- Salomón, L.; Nicola, M.V.; Kandziora, M.; Kolář, F.; Sklenář, P. Center of Origin and Evolutionary History in the High Andean Genus Oritrophium (Astereae, Asteraceae). Alp. Bot. 2021, 132, 123–139. [Google Scholar] [CrossRef]
- Namzalov, B.B. Baikal Phytogeographic Node as the Newest Center of Endemism of Inner Asia. Contemp. Probl. Ecol. 2009, 2, 341–347. [Google Scholar] [CrossRef]
- Jones, M.R.; Winkler, D.E.; Massatti, R. The Demographic and Ecological Factors Shaping Diversification among Rare Astragalus Species. Divers. Distrib. 2021, 27, 1407–1421. [Google Scholar] [CrossRef]
- Cortés, A.J.; Garzón, L.N.; Valencia, J.B.; Madriñán, S. On the Causes of Rapid Diversification in the Páramos: Isolation by Ecology and Genomic Divergence in Espeletia. Front. Plant Sci. 2018, 9, 1700. [Google Scholar] [CrossRef]
- Sampson, J.; Byrne, M. Genetic Differentiation among Subspecies of Banksia nivea (Proteaceae) Associated with Expansion and Habitat Specialization. Diversity 2022, 14, 98. [Google Scholar] [CrossRef]
- Özüdoğru, B.; Karacaoğlu, Ç.; Akaydın, G.; Erik, S.; Mummenhoff, K.; Sağlam, I.K. Ecological Specialization Promotes Diversity and Diversification in the Eastern Mediterranean Genus Ricotia (Brassicaceae). J. Syst. Evol. 2022, 60, 331–343. [Google Scholar] [CrossRef]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The Frequency of Polyploid Speciation in Plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. Ancient WGD Events as Drivers of Key Innovations in Angiosperms. Curr. Opin. Plant Biol. 2016, 30, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Van de Peer, Y.; Mizrachi, E.; Marchal, K. The Evolutionary Significance of Polyploidy. Nat. Rev. Genet. 2017, 18, 411–424. [Google Scholar] [CrossRef]
- Martin, E.; Karaman Erkul, S.; Aytaç, Z. Karyological Studies on Oxytropis (Fabaceae) from Turkey. Caryologia 2015, 68, 357–362. [Google Scholar] [CrossRef]
- Liu, Y.-H.; Meng, Y.; Yang, Y.-H.; Yang, Y.-P. Chromosome Numbers and Karyotypes of Six Oxytropis Species (Fabaceae) from the Qinghai Tibetan Plateau, China. Plant Divers. Res. 2011, 33, 423–431. (In Chinese) [Google Scholar]
- Yurtsev, B.A. Oxytropis DC. In Arctic Flora of the Soviet Union; Yurtsev, B.A., Ed.; Nauka: Leningrad, Russia, 1986; pp. 61–146. (In Russian) [Google Scholar]
- Yurtsev, B.A. Main Directions in the Evolution of Oxytropis Species Belonging to the Section Baikalia Bge. Bot. Zhurnal 1964, 49, 634–648. (In Russian) [Google Scholar]
- Vassilczenko, I.T. On the Problem of the Genesis of the Genus Oxytropis DC. Bot. Zhurnal 1965, 50, 313–323. (In Russian) [Google Scholar]
- Xu, Z.; Zhang, M.-L.; Cohen, J.I. Phylogeographic History of Atraphaxis Plants in Arid Northern China and the Origin of A. bracteata in the Loess Plateau. PLoS ONE 2016, 11, e0163243. [Google Scholar] [CrossRef]
- Segatto, A.L.A.; Reck-Kortmann, M.; Turchetto, C.; Freitas, L.B. Multiple Markers, Niche Modelling, and Bioregions Analyses to Evaluate the Genetic Diversity of a Plant Species Complex. BMC Evol. Biol. 2017, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ma, W.-X.; He, X.; Zhao, T.-T.; Yang, X.-H.; Wang, L.-J.; Ma, Q.-H.; Liang, L.-S.; Wang, G.-X. Species Divergence and Phylogeography of Corylus heterophylla Fisch Complex (Betulaceae): Inferred from Molecular, Climatic and Morphological Data. Mol. Phylogenet. Evol. 2022, 168, 107413. [Google Scholar] [CrossRef] [PubMed]
- Bobrov, E.G. Introgressive Hybridization in the Flora of Baikal Siberia. Bot. Zhurnal 1961, 46, 313–327. (In Russian) [Google Scholar]
- Orthia, L.A.; Crisp, M.D.; Cook, L.G.; de Kok, R.P.J. Bush Pea: A Rapid Radiation with no Support for Monophyly of Pultenaea (Fabaceae: Mirbelieae). Aust. Syst. Bot. 2005, 18, 133–147. [Google Scholar] [CrossRef]
- Drummond, C.S.; Eastwood, R.J.; Miotto, S.T.S.; Hughes, C.E. Multiple Continental Radiations and Correlates of Diversification in Lupinus (Leguminosae): Testing for Key Innovation with Incomplete Taxon Sampling. Syst. Biol. 2012, 61, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, A.; Maassoumi, A.A.; Rahiminejad, M.R.; Brassac, J.; Blattner, F.R. Molecular Phylogeny and Divergence Times of Astragalus Section Hymenostegis: An Analysis of a Rapidly Diversifying Species Group in Fabaceae. Sci. Rep. 2017, 7, 14033. [Google Scholar] [CrossRef]
- Zhao, X.-L.; Gao, X.-F.; Zhu, Z.-M.; Gao, Y.-D.; Xu, B. The Demographic Response of a Deciduous Shrub (the Indigofera bungeana complex, Fabaceae) to the Pleistocene Climate Changes in East Asia. Sci. Rep. 2017, 7, 697. [Google Scholar] [CrossRef]
- Bartha, L.; Dragos, N.; Molnar, A.; Sramko, G. Molecular Evidence for Reticulate Speciation in Astragalus (Fabaceae) as Revealed by a Case Study from Sect. Dissitiflori. Botany 2013, 91, 702–714. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.N. The Taxonomical Study of Oxytropis DC. in Inner Mongolia. Master’s Thesis, Inner Mongolia Normal University, Inner Mongolia, China, 2005. [Google Scholar]
- Zhao, X.; Hou, Q.; Du, M.; Zhang, H.; Jia, L.; Zhang, Z.; Ma, Z.; Sun, K. Micromorphological Leaf Epidermal Traits as Potential Taxonomic Markers for Infrageneric Classification of Oxytropis (Fabaceae). PhytoKeys 2022, 201, 51–76. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandanov, D.V.; Kholina, A.B.; Kozyrenko, M.M.; Artyukova, E.V.; Wang, Z. Genetic Diversity of Oxytropis Species from the Center of the Genus Origin: Insight from Molecular Studies. Diversity 2023, 15, 244. https://0-doi-org.brum.beds.ac.uk/10.3390/d15020244
Sandanov DV, Kholina AB, Kozyrenko MM, Artyukova EV, Wang Z. Genetic Diversity of Oxytropis Species from the Center of the Genus Origin: Insight from Molecular Studies. Diversity. 2023; 15(2):244. https://0-doi-org.brum.beds.ac.uk/10.3390/d15020244
Chicago/Turabian StyleSandanov, Denis V., Alla B. Kholina, Marina M. Kozyrenko, Elena V. Artyukova, and Zhiheng Wang. 2023. "Genetic Diversity of Oxytropis Species from the Center of the Genus Origin: Insight from Molecular Studies" Diversity 15, no. 2: 244. https://0-doi-org.brum.beds.ac.uk/10.3390/d15020244