
Microclimatic Influences on the Abundance of Three Non-Troglobiont Species



1
Department of Life, Health and Environmental Sciences (MeSVA), University of L’Aquila, 67100 L’Aquila, Italy
2
Department of Environmental Sciences and Policy, Università degli Studi di Milano, 20133 Milan, Italy
3
Natural Oasis, 59100 Prato, Italy
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(3), 186; https://0-doi-org.brum.beds.ac.uk/10.3390/d16030186
Submission received: 7 February 2024
/
Revised: 11 March 2024
/
Accepted: 14 March 2024
/
Published: 19 March 2024
(This article belongs to the Section Biodiversity Loss & Dynamics)
Abstract
:Subterranean environments are often characterized by a natural gradient of microclimatic conditions and trophic resources, showing a higher trophic availability and a lower microclimatic stability in the shallowest area (close to the cave entrance), while the opposite occurs in the deepest sections. The shallowest areas of subterranean environments (e.g., the entrance and twilight zone, Mesovoid Shallow Substratum) act as ecotones between the surface habitats and the deep areas, creating a particular habitat which can be exploited by numerous species with different degrees of adaptation to subterranean environments. Species living in these ecotones may hold a key role in sustaining the entire ecosystem, as they are likely one of the major drivers of allochthonous organic matter. Indeed, these species are usually facultative cave-dwellers, meaning that they are able to exit and forage on the surface. Once these species are back inside the cave, they provide the local community with different typologies of organic matter (e.g., feces, eggs), which represent one of the most important sources of organic carbon. Therefore, studying which ecological features may exert significant effects on the abundance of these species may be of great help in understanding the ecosystem dynamics and the functional role of each species. In this study we analyzed the data collected through a year-round monitoring program, aiming to assess the potential effects that both abiotic and biotic features may have on the abundance of three facultative cave species. We focused on seven caves located in Monte Albo (Sardinia, Italy). The cave environments were divided into 3-meter sectors, and within each cave sector, microclimatic and biological data were seasonally recorded. We focused on the following facultative cave species: the spiders Metellina merianae and Tegenaria sp. and the snail Oxychilus oppressus. Different relationships were observed between the ecological features and the abundance of the three species. The two spiders were more abundant in warmer cave sectors closer to the cave entrance, especially the M. merianae. On the other hand, the snail tended to be more abundant farther from the cave entrance and in more illuminated cave sectors, probably because sunlight promotes the abundance of some of its trophic resources (e.g., lichens, vegetation). Furthermore, O. oppressus was the only species whose abundance and cave distribution was significantly affected by seasonality. This study provides useful and novel information to understand the population dynamics of facultative cave species and their role in subterranean ecosystems.
1. Introduction
Subterranean habitats are characterized by peculiar environmental features, where darkness and relatively stable microclimatic conditions are among the most common ones [1]. These conditions mainly occur in the deepest cave sections (i.e., far from the cave entrance which connects the subterranean environment with the surface), where the influences of external climatic conditions are insignificant [2]. On the other hand, external influences such as sunlight, wind, and rain, can strongly affect the environmental conditions near the cave entrance, making them more similar to those found in surface habitats [3,4]. Once they penetrate the subterranean environment, these external influences gradually lose their strength the deeper they go, causing the occurrence of a natural microclimatic gradient extending from the cave entrance (high microclimatic fluctuations resembling conditions on the surface) to the deepest cave sections (high microclimatic stability with an air temperature similar to the annual average air temperature outside the cave) [5]. Along with microclimatic conditions, the trophic supply further contributes to characterizing subterranean habitats. The general lack of sunlight in caves prevents the survival of plants (with a few exceptions for more illuminated cave sections where some species, particularly Cryptogams plant, can be stable residents) [6]; the lack of primary producers drastically contributes to reducing the quantity of organic matter available to local communities [1]. Subterranean habitats are therefore strongly dependent on the external inputs of organic matter, which are more consistent near the cave entrance and almost absent in the deepest cave sections [7,8,9]. Indeed, around the cave entrance, inputs of organic matter originating from the surface environment can be driven not only by species able to exit and forage on the surface [8,10] but also through atmospheric agents such as wind, gravity, and water [1]. On the contrary, organic matter in the deepest areas mostly depends on animals’ inputs and, in lesser amounts, can also be transported by water [7,9,11,12]. Consequently, considering both abiotic and biotic gradients, subterranean environments can be divided into two distinct ecological zones: the deep subterranean environment, which includes the deepest cave sections characterized by high microclimatic stability and a low availability of organic matter, and the surface–subterranean interface, which embraces the shallowest areas characterized by low microclimatic stability and a higher availability of organic matter [5].
The subterranean realm is populated by numerous species, most of which are still unknown [13]. Species that inhabit subterranean environments may show an array of behavioral, physiological, and morphological adaptations allowing them to cope with the ecological features characterizing such environments [1]. For example, the lack of sunlight promoted eye reduction, the elongation of appendages and the loss of pigmentation [14,15], the loss of a circadian rhythm [16,17], and intraspecific interactions which not rely on visual cues [18,19]. Morphological adaptations are the easiest to spot, and this likely promoted their historical use for categorizing subterranean-dwelling species, distinguishing between those adapted to live in subterranean environments (i.e., species showing morphological adaptations) and those which are not (lacking evident adaptive traits) [20]. However, in recent years there has been an increase in our awareness of other less apparent adaptations (e.g., [21,22,23]), making them equally important for classifying subterranean species. The current widely used classification of cave species identifies three main groups according to their degree of adaptation to subterranean environments and their ability to complete their life cycle therein [5,10]. Troglobionts are the most adapted species which can only live and reproduce in subterranean environments; they can exit only under exceptional conditions [24,25]. Troglophiles are facultative cave species which show some adaptations to subterranean environments, where they can reproduce and have stable populations. Trogloxenes are the least adapted species (they mostly lack any adaptations) that occasionally use subterranean environments and are mostly related to the shallowest cave sections.
Considering the intrinsic and objective difficulty in their exploration [26,27,28], the knowledge of subterranean habitats, and of the inhabiting species, is still far from satisfactory. Most researchers’ efforts have been given to the study of the species showing the highest degree of adaptation to subterranean environments (e.g., [18,29,30]), while studies on apparently less adapted ones are still limited [31,32,33]. Despite probably being less “attractive” compared to the troglobionts mostly living in the deepest cave areas, many species living in the transition zone between the surface and the deep subterranean environments (e.g., cave twilight zone, Mesovoid Shallow Substratum; [3]) may hold a strategic role in the sustainment of the entire subterranean community. Indeed, considering their ability to exit caves, these species often forage in surface environments, where the trophic supply is higher [1,34], and then go back to their subterranean refuges where they enrich the environment with their inputs (e.g., feces, eggs) [8,35], representing important drivers of allochthonous organic matter that can be crucial for sustaining the entire cave community [36,37,38].
With this study we aim to evaluate which and to what extent selected ecological conditions drive the abundance of troglophile species inhabiting selected caves under study. The study of the abundance of non-obligate subterranean species can provide important information about multiple species’ life traits such as their phenology, reproduction, or interactions with other species [39,40,41,42], serving as insights into their partial adaptations to the subterranean environment [43]. Furthermore, studying the phenology of these species can provide important information allowing us to understand the dynamics of subterranean communities; for example, assessing when troglophiles foraging activity is the highest may help in understanding in which period the supply of organic matter is the highest [12]. Nonetheless, identifying the cave sections in which these species are usually more abundant would allow us to hypothesize the potential distribution of the imported organic matter [44]. The three species considered here are two spiders, Metellina merianae (Scopoli, 1763) and Tegenaria sp. Latreille, 1804, and a snail, Oxychilus oppressus (Shuttleworth,1877), inhabiting the caves of a specific area of the island of Sardinia (Italy). The two spider species are commonly found close to the cave entrance and in the twilight zone of most of the caves in Europe [45]. In a study on the occupancy of these species in a group of caves in Tuscany (Italy), it was found that both microclimatic and cave morphological features contribute differently to the prediction of the presence/absence of these species in cave sectors [46]. On the other hand, studies considering their abundance were never performed. The snail O. oppressus represents an endemic species of Sardinia (Italy) [47] and no studies on its ecology exist. Knowing the ecological factors that affect the abundance of facultative cave species may be of high importance due to their fundamental role in sustaining the subterranean communities [1]. Indeed, such data can provide useful information allowing us to quantify their contribution to supplying allochthonous organic matter, and where such deposits can be more abundant.
2. Materials and Methods
We analyzed the dataset published by Lunghi et al. [48], which provides data on the abundance of different troglophile species recorded in seven natural caves in Monte Albo (Sardinia, Italy). Cave surveys were performed throughout the year, from autumn 2015 to summer 2016. During each season pairs of surveys were performed during the daytime (9 AM–6 PM) with a maximum gap of seven days; this guaranteed that the criteria of population closure were met (i.e., no immigration/emigration and no birth occurring in the meantime), and that a wide divergence of climatic conditions could be avoided [48,49]. This allowed us to calculate the species detection probability. Within each cave, the cave passage was divided into 3 m long sectors (hereafter, cave sector); at the end of each sector, abiotic and biotic data were collected following a standardized protocol that also considered the occurrence of air fluxes [2,48]. At the end of each cave sector, air temperature (°C) and relative humidity (%) were measured at the ground level and at 2.5 m of height (or at the ceiling if lower); data were then merged to obtain the average values for each cave sector. Illuminance (lux) was also recorded at the end of each cave sector. Two measurements, one from the most illuminated and one from the darkest place, were performed; data were then averaged. Within each cave sector, species were searched for and counted through Visual Encounter Surveys [50] adopting a standardized sampling effort of 7.5 min/sector.
To estimate species detection probability, we built three models in the Unmarked R package [51]: one with sector depth as the covariate, one with survey season as the covariate, one without covariates. We used these covariates to assess whether the species detection probability varied along the cave (i.e., from the entrance to the deepest cave section) and/or was affected by seasonality (i.e., due to the different activity of individuals). We ranked models following the Akaike information criterion (AIC); the best model is indicated with the lowest AIC value [52,53]. The Analysis of Variance (ANOVA) was used to assess whether the range of depths and microclimatic conditions (air temperature, humidity, and illuminance) at which the studied species occurred the most significantly led to differences. For each species, we used the averaged microclimatic data recorded within cave sectors throughout the year. The pairwise comparisons were assessed using Tukey’s post hoc test. We then applied Generalized Linear Mixed Models (GLMMs) in lme4 package [54] to assess the effects of both abiotic and biotic features on the abundance of the three troglophile species. We used the observed abundance of the studied species as a dependent variable, considering it as a proxy of the species’ true abundance [55,56]. Independent variables were: sector depth (i.e., linear distance from cave entrance), the season of survey, and three microclimatic variables (average temperature, humidity, and illuminance). The interaction between season and sector depth was added as an additional independent variable, as it was observed that troglophile species occupy different cave sections seasonally in search of the most suitable microclimatic conditions [4]. We also added the presence of crane fly Diptera (probably genus Limonia Meigen, 1803) as an independent variable, as this species usually represents the most abundant food resource for many predators and scavengers [57,58,59]. Due to the repeated surveys performed throughout the year, caves and sectors’ identities were included as random factors. The likelihood ratio test was used to test the significance of variables in lmerTest package [60]. Analyses were performed in the R environment [53].
3. Results
A total of 863 surveys were performed during the study year (autumn = 179, winter = 179, spring = 326, summer = 179), collecting 331 observations of the studied species (Metellina merianae = 165, Oxychilus oppressus = 93, Tegenaria sp. = 73). The species generally showed a low detection probability (Table 1).
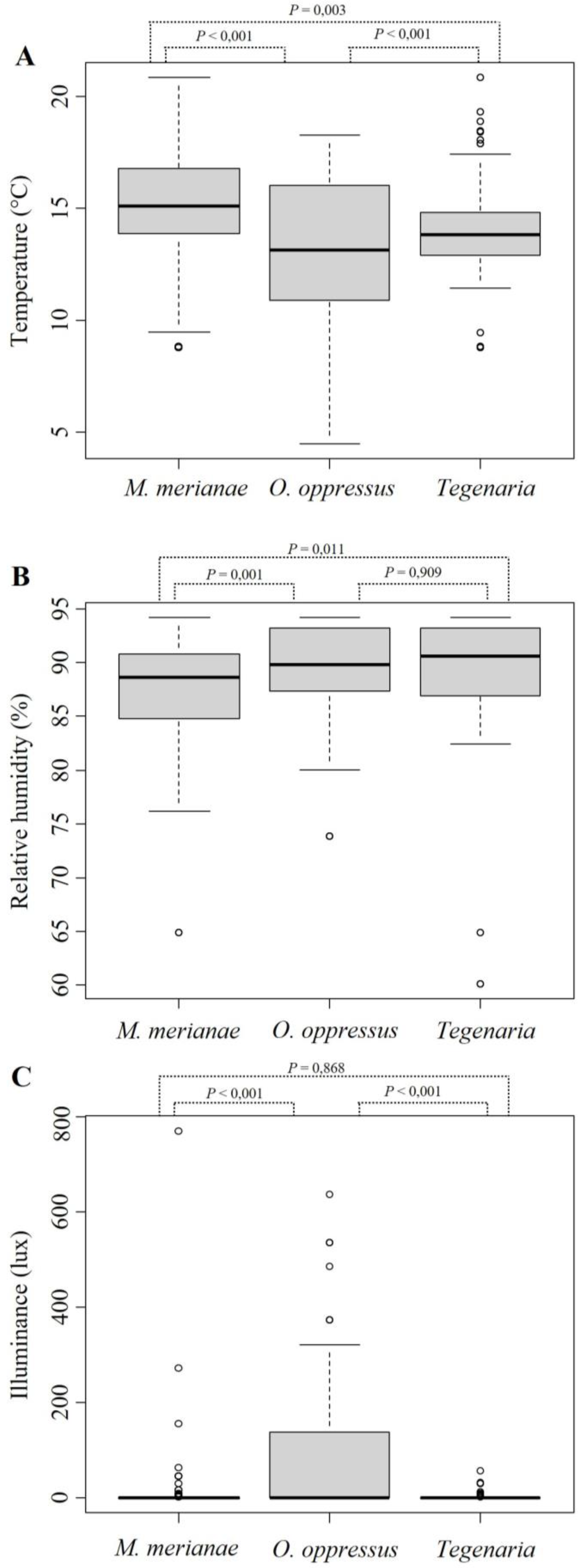
The average annual microclimatic conditions of the monitored caves were the following: temperature (°C), average 13.9, min 4.3, max 23.8; relative humidity (%), average 90, min 55.9, max 94.9; illuminance (lux); average 166.9, min 0, max 42,322. There were significant differences in the temperatures of the sectors occupied by the three species (df = 2, F = 32.02, p < 0.001); M. merianae was present in warmer sectors compared to the other two species (Figure 1A). The values for the relative air humidity in the occupied sectors significantly differed between the three species (df = 2, F = 7.86, p < 0.001); M. merianae was present in drier sectors compared to the other two species (Figure 1B). There were significant differences in the illuminance of the sectors occupied by the three species (df = 2, F = 29.46, p < 0.001); O. oppressus was present in more illuminated sectors compared to the other two species (Figure 1C).
The studied species showed different abundance patterns (df = 2, F = 5.18, p = 0.006). M. meriane was more abundant closer to the cave entrance (average cave sector depth = 18.35 m, min = 3, max = 63), followed by Tegenaria spiders (average cave sector depth = 24.16 m, min = 3, max = 72) and by O. oppressus (average cave sector depth = 28.94 m, min = 3, max = 75).
The abundance of M. merianae was affected by the cave sector’s depth (F1,207.11 = 27.50, p < 0.001), by the presence of diptera (F1,781.91 = 10.69, p = 0.001), by the sector’s humidity (F1,622.27 = 6.61, p = 0.01), and by the sector’s illuminance (F1,242.09 = 6.75, p = 0.01). This species was more abundant in drier (estimate = −0.5, ±SE = 0.2) and less illuminated (estimate = −0.02, SE = 0.007) sectors close to the cave entrance (estimate = −0.004, ±SE = 0.001) with a greater presence of crane flies (estimate = 0.095, ±SE = 0.029).
The abundance of O. oppressus was affected by the cave sector’s depth (F1,230.40 = 7.72, p = 0.006), by the sector’s humidity (F1,426.37 = 7.12, p = 0.008) and illuminance (F1,194.16 = 19.21, p < 0.001), by season (F3,399.99 = 6.22, p < 0.001), and by the interaction between the depth and the season (F3,680.48 = 5.44, p = 0.001). The snail was overall more abundant in drier (estimate = −0.42, ±SE = 0.157) and more illuminated (estimate = 0.022, SE = 0.005) sectors close to the cave entrance (estimate = −0.001, ±SE = 0.001). Furthermore, the species was more abundant in autumn, which is the season in which individuals occupy sectors closer to the cave entrance.
The abundance of Tegenaria spiders was only affected by the cave sector’s depth (F1,205.3 = 8.90, p = 0.003); this spider was more abundant in sectors close to the cave entrance (estimate = −0.003, ±SE = 0.001).
4. Discussion
The analysis performed here identified that different ecological factors affect the presence and abundance of the three facultative cave species. Our results highlighted a higher abundance of the studied species within the shallowest cave sections, those representing the interface between surface habitats and the deepest cave sections. The species’ detectability was generally low, being lower for the spider species compared to the snail. This may be because spiders immediately retreat into cracks in walls or inside their webs (i.e., funnel webs) when they are disturbed and elude operator sight. Indeed, during cave surveys it is not rare to observe just the spider web with no trace of its owner. Similarly, analyses performed on a further syntopic species (i.e., from the same caves considered in this study), the tetragnathid Meta bourneti Simon, 1922, also highlighted a relatively low dp for that spider [41]. Interestingly, spiders from mainland Italy (M. merianae and M. menardi) showed a much higher detection probability [more than twice that observed here; 46], making us wonder about the reasons for such a low detection probability being observed in the caves included in this study. In the future, studies aiming to assess the causes of such divergence would shed light on these puzzling results, also evaluating whether such a low detection probability is only related to the studied area of Monte Albo massif or if it is a characteristic of the species distributed across Sardinia Island. Furthermore, we cannot exclude that the low detection probability observed here is related to the lower population density, which may provide insights into potential conservation issues [61]. Unfortunately, there are no similar studies for Oxychilus snails yet, hampering any possible parallelism for this species. Further studies are therefore needed to assess whether Oxychilus snails from Sardinia also show lower detection probabilities compared to congeneric species from mainland Italy.
Considering the data obtained from occupied cave sectors we were able to identify differences in the range of microclimatic conditions experienced by the studied species. Our results highlighted the possibility that the three cave-dwelling species considered here differentiate their distribution according to environmental features characterizing the cave sections between the entrance and deeper areas. According to our results, each species occupied sections with microclimatic conditions corresponding to the species’ ecological preference (Figure 1), modulating its functional contribution to the cave community [7]. The spider M. merianae seems to tolerate harsher microclimatic conditions (i.e., higher temperatures and lower humidity) compared to the other two species (Figure 1). This could be due to a lack (or limited) of adaptation to the subterranean environment [29,43], a characteristic that increases the chances of the species to forage in the shallowest cave section and in the surface environment, where prey availability is higher [1,8]. Are, therefore, M. merianae spiders able to supply more allochthonous organic matter compared to the other two species? Is the species’ contribution higher and/or limited to the shallowest areas of the caves? On the other hand, the abundance of Tegenaria spiders was not affected by any of the considered microclimatic features, but were just more abundant close to the cave entrance. Differently from what occurred for M. merianae, the abundance of crane flies did not significantly affect the abundance of Tegenaria spiders. This interesting information allows us to hypothesize that these spiders probably prefer different typologies of prey compared to M. merianae, being, therefore, more specialized in capturing non-flying prey [62,63]. This hypothesis needs to be confirmed. The snail O. oppressus was the species that better tolerated colder temperatures, being observed in cave sectors in which the temperature goes below 5 °C (Figure 1A). This characteristic probably expanded the suitable cave area for the species, allowing its presence also in deep cave sections beyond the transition zone. These first insights should be implemented in the future with studies quantifying the specific contribution of each species to the sustainment of the subterranean community within the inhabited ecological zone. Studying the distribution and activity patterns of subterranean species that forage in the surface environment allows us to correlate their abundance with the quantity and distribution of allochthonous organic matter within cave environments [7,64].
The abundance of the three studied species decreased with increasing depth, suggesting their greatest presence within the first few meters of the surface–cave ecotone [46]. This interpretation is supported by the significantly higher abundances of the species observed in drier cave sectors, particularly for the spider M. merianae and the snail O. oppressus (for the latter also in more illuminated cave sectors), being microclimatic conditions that usually characterize the cave sections close to the cave entrance [4]. However, the occupancy and the abundance of the studied species were differently affected by the ecological factors. Combining the use of webs with an active hunting strategy, some cave spiders are able to prey upon a wide number of species entering the cave [45,65,66]. Residuals of consumed prey can be at the disposal for scavengers, therefore representing an important source of organic material for the subterranean community. This is particularly true for the tetragnathid M. merianae, whose abundance was positively influenced by the presence of Limonia dipterans. Crane flies are usually abundant in the first few meters inside the caves [57,67], where they represent one of the most frequent prey for tetragnathid spiders (genera Meta C. L. Koch, 1835 and Metellina Chamberlin and Ivie, 1941; [66]) and other cave predators [68]. However, proper studies on the diet of Tegenaria spiders in caves are lacking, hampering our ability to evaluate whether our results reflect a generally low contribution of crane flies to the diet of representatives of this genus.
The abundance of the snail was higher in cave sectors farther from the cave entrance compared to the two spider species. This might be due to the different foraging strategies adopted by these species. Indeed, although spiders are strictly predators, Oxychilus snails can modulate their foraging strategy (from being predators to scavengers; [59]), being able to exploit habitats characterized by different trophic resources [1,58]. For example, for cave-dwelling snails, vegetation and lichens represent a consistent trophic resource [69]; this may be also the case for O. oppressus. Indeed, the species tended to be more abundant in more illuminated cave sectors, an environmental characteristic which is essential for these autotrophic organisms. Nonetheless, snails may be characterized by a lower metabolic rate compared to spiders, a characteristic which allows them to exploit food-deprived cave sections [21]. The abundance of the snail was the only affected by seasonality; compared to the other seasons, in autumn the species’ abundance increased towards the cave entrance. It has been observed in another Oxychilus species that its highest reproductive activity is reached between May and November [70]. The climatic conditions which occur in Sardinia can be particularly harsh for O. oppressus (too hot and dry) in late spring and summer [48], forcing the species to concentrate its reproductive activity in autumn.
5. Conclusions
This research provides useful and novel information about the relationship between multiple environmental factors and the population dynamics of a heterogeneous group of three facultative cave species. By analyzing the abundance of these species we identified a divergence in their distribution along the ecotonal zone that connects the surface environments with the deep cave sections. This divergence was affected by a different preference for microclimatic features, but also by a divergent distribution of the preferred trophic resources. The insights provided by our study should be used to deepen our knowledge about the role of non-troglobiont species in the cave community and in the overall subterranean ecosystem. This should also stimulate more interest in less adapted, and therefore usually neglected, cave species, allowing researchers to shed light on several unknown cave community dynamics. Our study provides the basis for further studies on a neglected, but rich in perspectives, subterranean ecotonal areas connecting the surface and deep subterranean environments.
Author Contributions
E.L. conceived the idea and performed the analyses; E.L. and L.C. drafted the manuscript. E.L., L.C. and R.M. have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Analized data can be retrieved from [48].
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats, 2nd ed.; Oxford University Press: New York, NY, USA, 2019. [Google Scholar]
- Badino, G. Underground Meteorology—“What’s the Weather Underground”? Acta Carsologica 2010, 39, 427–448. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats: Ecology, Evolution, and Conservation; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Seasonal Variation in Microhabitat of Salamanders: Environmental Variation or Shift of Habitat Selection? PeerJ 2015, 3, e1122. [Google Scholar] [CrossRef]
- Mammola, S. Finding Answers in the Dark: Caves as Models in Ecology Fifty Years after Poulson and White. Ecography 2019, 42, 1331–1351. [Google Scholar] [CrossRef]
- Monro, A.K.; Bystriakova, N.; Fu, L.; Wen, F.; Wei, Y. Discovery of a Diverse Cave Flora in China. PLoS ONE 2018, 13, e0190801. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.L.; Martins, R.P. Trophic Structure and Natural History of Bat Guano Invertebrate Communities, with Special Reference to Brazilian Caves. Trop. Zool. 1999, 12, 231–252. [Google Scholar] [CrossRef]
- Lavoie, K.H.; Helf, K.L.; Poulson, T.L. The Biology and Ecology of North American Cave Crickets. J. Cave Karst Stud. 2007, 69, 114–134. [Google Scholar]
- Lunghi, E.; Valle, B.; Guerrieri, A.; Bonin, A.; Cianferoni, F.; Manenti, R.; Ficetola, G.F. Complex Patterns of Environmental DNA Transfers from Surface to Subterranean Soils Revealed by Analyses of Cave Insects and Springtails. Sci. Total Environ. 2022, 826, 154022. [Google Scholar] [CrossRef] [PubMed]
- Howarth, F.G.; Moldovan, O.T. The Ecological Classification of Cave Animals and Their Adaptations. In Cave Ecology; Moldovan, O.T., Kováč, Ľ., Halse, S., Eds.; Springer: Berlin, Germany, 2018; pp. 41–67. [Google Scholar]
- Fenolio, D.B.; Niemiller, M.L.; Bonett, R.M.; Graening, G.O.; Collier, B.A.; Stout, J.F. Life History, Demography, and the Influence of Caveroosting Bats on a Population of the Grotto Salamander (Eurycea Spelaea) from the Ozark Plateaus of Oklahoma (Caudata: Plethodontidae). Herpetol. Conserv. Biol. 2014, 9, 394–405. [Google Scholar]
- Simon, K.S.; Pipan, T.; Culver, D.C. A Conceptual Model of the Flow and Distribution of Organic Carbon in Caves. J. Cave Karst Stud. 2007, 69, 279–284. [Google Scholar]
- Ficetola, G.F.; Canedoli, C.; Stoch, F. The Racovitzan Impediment and the Hidden Biodiversity of Unexplored Environments. Conserv. Biol. 2019, 33, 214–216. [Google Scholar] [CrossRef]
- Bilandžija, H.; Ćetković, H.; Jeffery, W.R. Evolution of Albinism in Cave Planthoppers by a Convergent Defect in the First Step of Melanin Biosynthesis. Evol. Dev. 2012, 14, 196–203. [Google Scholar] [CrossRef]
- Gonzalez, B.C.; Worsaae, K.; Fontaneto, D.; Martínez, A. Anophthalmia and Elongation of Body Appendages in Cave Scale Worms (Annelida: Aphroditiformia). Zool. Scr. 2017, 47, 106–121. [Google Scholar] [CrossRef]
- Mack, K.L.; Jaggard, J.B.; Persons, J.L.; Roback, E.Y.; Passow, C.N.; Stanhope, B.A.; Ferrufino, E.; Tsuchiya, D.; Smith, S.E.; Slaughter, B.D.; et al. Repeated Evolution of Circadian Clock Dysregulation in Cavefish Populations. PLoS Genet. 2021, 17, e1009642. [Google Scholar] [CrossRef]
- Pasquali, V.; Sbordoni, V. High Variability in the Expression of Circadian Rhythms in a Cave Beetle Population. Biol. Rhythm. Res. 2014, 45, 925–939. [Google Scholar] [CrossRef]
- Hyacinthe, C.; Attia, J.; Rétaux, S. Evolution of Acoustic Communication in Blind Cavefish. Nat. Commun. 2019, 10, 4231. [Google Scholar] [CrossRef]
- Plath, M.; Parzefall, J.; Körner, K.E.; Schlupp, I. Sexual Selection in Darkness? Female Mating Preferences in Surface- and Cave-Dwelling Atlantic Mollies, Poecilia mexicana (Poeciliidae, Teleostei). Behav. Ecol. Sociobiol. 2004, 55, 596–601. [Google Scholar] [CrossRef]
- Christiansen, K. Proposition Pour La Classification Des Animaux Cavernicoles. Spelunca 1962, 2, 76–78. [Google Scholar]
- Hervant, F. Starvation in Subterranean Species versus Surface-Dwelling Species: Crustaceans, Fish, and Salamanders. In Comparative Physiology of Fasting, Starvation, and Food Limitation; McCue, M.D., Ed.; Springer: Berlin, Germany, 2012; pp. 91–102. [Google Scholar]
- Lunghi, E.; Bilandžija, H. Longevity in Cave Animals. Front. Ecol. Evol. 2022, 10, 874123. [Google Scholar] [CrossRef]
- Lunghi, E.; Mammola, S.; Martinez, A.; Hesselberg, T. Behavioural Adjustments Enable the Colonization of Subterranean Environments. Zool. J. Linn. Soc. Lond. 2023, zlad133. [Google Scholar] [CrossRef]
- Kirn, T. Finds of Washed-out Proteus from the Pivka Intermittent Lakes and the Pivka River. Nat. Slov. 2018, 20, 51–56. [Google Scholar] [CrossRef]
- Manenti, R.; Di Nicola, M.R.; Zampieri, V.; Grassi, G.; Creanza, T.; Mauri, E.; Ficetola, G.F.; Barzaghi, B. Wandering Outside of the Styx: Surface Activity of an Iconic Subterranean Vertebrate, the Olm (Proteus anguinus). Ecology 2024, 105, e4252. [Google Scholar] [CrossRef] [PubMed]
- Buzzacott, P.L.; Zeigler, E.; Denoble, P.; Vann, R. American Cave Diving Fatalities 1969–2007. Int. J. Aquat. Res. Educ. 2009, 3, 162–177. [Google Scholar] [CrossRef]
- Iliffe, T.M.; Bowen, C. Scientific Cave Diving. Mar. Technol. Soc. J. 2001, 35, 36–41. [Google Scholar] [CrossRef]
- Zagmajster, M.; Culver, D.C.; Christman, M.C.; Sket, B. Evaluating the Sampling Bias in Pattern of Subterranean Species Richness: Combining Approaches. Biodivers. Conserv. 2010, 19, 3035–3048. [Google Scholar] [CrossRef]
- Rizzo, V.; Sánchez-Fernández, D.; Fresneda, J.; Cieslak, A.; Ribera, I. Lack of Evolutionary Adjustment to Ambient Temperature in Highly Specialized Cave Beetles. BMC Evol. Biol. 2015, 15, 10. [Google Scholar] [CrossRef]
- Fenolio, D.B.; Graening, G.O.; Collier, B.A.; Stout, J.F. Coprophagy in a Cave-Adapted Salamander; the Importance of Bat Guano Examined through Nutritional and Stable Isotope Analyses. Proc. R. Soc. B 2006, 273, 439–443. [Google Scholar] [CrossRef]
- Guillaume, O. Surface Newt Calotriton Asper Acclimation to Cave Conditions Improved Their Foraging Ability in Darkness. Front. Ecol. Evol. 2022, 10, 1057023. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Do Cave Features Affect Underground Habitat Exploitation by Non-Troglobite Species? Acta Oecologica 2014, 55, 29–35. [Google Scholar] [CrossRef]
- Lunghi, E. Occurrence of the Black Lace-Weaver Spider, Amaurobius Ferox, in Caves. Acta Carsologica 2020, 49, 119–124. [Google Scholar] [CrossRef]
- Venarsky, M.P.; Huntsman, B.M.; Huryn, A.D.; Benstead, J.P.; Kuhajda, B.R. Quantitative Food Web Analysis Supports the Energy Limitation Hypothesis in Cave Stream Ecosystems. Oecologia 2014, 176, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Mulargia, M.; Veith, M.; Corti, C.; Ficetola, G.F. Environmental Suitability Models Predict Population Density, Performance and Body Condition for Microendemic Salamanders. Sci. Rep. 2018, 8, 7527. [Google Scholar] [CrossRef]
- Barzaghi, B.; Ficetola, G.F.; Pennati, R.; Manenti, R. Biphasic Predators Provide Biomass Subsidies in Small Freshwater Habitats: A Case Study of Spring and Cave Pools. Freshw. Biol. 2017, 62, 1637–1644. [Google Scholar] [CrossRef]
- Chapin, K.J. Guano Deposition Predicts Top Predator (Amblypygi: Phrynidae) Abundance in Subtropical Caves. J. Arachnol. 2019, 47, 385–388. [Google Scholar] [CrossRef]
- Salgado, S.S.; Motta, P.C.; De Souza Aguiar, L.M.; Nardoto, G.B. Tracking Dietary Habits of Cave Arthropods Associated with Deposits of Hematophagous Bat Guano: A Study from a Neotropical Savanna. Austral Ecol. 2014, 39, 560–566. [Google Scholar] [CrossRef]
- Acevedo, P.; Escudero, M.A.; Muńoz, R.; Gortázar, C. Factors Affecting Wild Boar Abundance across an Environmental Gradient in Spain. Acta Theriol. 2006, 51, 327–336. [Google Scholar] [CrossRef]
- Amarasekare, P.; Coutinho, R.M. Effects of Temperature on Intraspecific Competition in Ectotherms. Am. Nat. 2014, 184, E50–E65. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E. Ecology and Life History of Meta bourneti (Araneae: Tetragnathidae) from Monte Albo (Sardinia, Italy). PeerJ 2018, 6, e6049. [Google Scholar] [CrossRef]
- Lunghi, E.; Bruni, G.; Ficetola, G.F.; Manenti, R. Is the Italian Stream Frog (Rana italica Dubois, 1987) an Opportunistic Exploiter of Cave Twilight Zone? Subterr. Biol. 2018, 25, 49–60. [Google Scholar] [CrossRef]
- Lipovšek, S.; Leitinger, G.; Janžekovič, F.; Kozel, P.; Dariš, B.; Perc, M.; Devetak, D.; Weiland, N.; Novak, T. Towards Understanding Partial Adaptation to the Subterranean Habitat in the European Cave Spider, Meta menardi: An Ecocytological Approach. Sci. Rep. 2019, 9, 9121. [Google Scholar] [CrossRef]
- Campbell Grant, E.H.; Lowe, W.H.; Fagan, W.F. Living in the Branches: Population Dynamics and Ecological Processes in Dendritic Networks. Ecol. Lett. 2007, 10, 165–175. [Google Scholar] [CrossRef]
- Mammola, S.; Isaia, M. Spiders in Cave. Proc. R. Soc. B 2017, 284, 20170193. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Cave Features, Seasonality and Subterranean Distribution of Non-Obligate Cave Dwellers. PeerJ 2017, 5, e3169. [Google Scholar] [CrossRef] [PubMed]
- Bank, R.A.; Neubert, E. MolluscaBase. Checklist of the Land and Freshwater Gastropoda of Europe. Available online: https://www.marinespecies.org/traits./aphia.php?p=sourcedetails&id=279050 (accessed on 1 March 2024).
- Lunghi, E.; Corti, C.; Mulargia, M.; Zhao, Y.; Manenti, R.; Ficetola, G.F.; Veith, M. Cave Morphology, Microclimate and Abundance of Five Cave Predators from the Monte Albo (Sardinia, Italy). Biodivers. Data J. 2020, 8, e48623. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling. Inferring Patterns and Dynamics of Species Occurrence; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- Crump, M.L.; Scott, N.J. Visual Encounter Surveys. In Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians; Heyer, W.R., Donnelly, M.A., McDiarmid, R.W., Hayek, L.C., Foster, M.S., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1994; pp. 84–92. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference. Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria.
- Douglas, B.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barke, R.J.; Schofield, M.R.; Link, W.A.; Sauer, J.R. On the Reliability of N-Mixture Models for Count Data. Biometrics 2017, 74, 369–377. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Barzaghi, B.; Melotto, A.; Muraro, M.; Lunghi, E.; Canedoli, C.; Lo Parrino, E.; Nanni, V.; Silva-Rocha, I.; Urso, A.; et al. N-Mixture Models Reliably Estimate the Abundance of Small Vertebrates. Sci. Rep. 2018, 8, 10357. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Ficetola, G.F.; Zhao, Y.; Manenti, R. Are the Neglected Tipuloidea Crane Flies (Diptera) an Important Component for Subterranean Environments? Diversity 2020, 12, 333. [Google Scholar] [CrossRef]
- Manenti, R.; Lunghi, E.; Ficetola, G.F. Distribution of Spiders in Cave Twilight Zone Depends on Microclimatic Features and Trophic Supply. Invertebr. Biol. 2015, 134, 242–251. [Google Scholar] [CrossRef]
- Curry, P.A.; Yeung, N.W. Predation on Endemic Hawaiian Land Snails by the Invasive Snail Oxychilus alliarius. Biodivers. Conserv. 2013, 22, 3165–3169. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, B.; Christensen, H.B. lmerTest: Tests in Linear Mixed Effects Models; R Package, Version 2.0–2.9; 2016. Available online: www.r-project.org (accessed on 10 December 2023).
- Ficetola, G.F.; Romano, A.; Salvidio, S.; Sindaco, R. Optimizing Monitoring Schemes to Detect Trends in Abundance over Broad Scales. Anim. Conserv. 2017, 21, 221–231. [Google Scholar] [CrossRef]
- Pekár, S.; Líznarová, E.; Řezáč, M. Suitability of woodlice prey for generalist and specialist spider predators: A comparative study. Ecol. Entomol. 2016, 41, 123–1300. [Google Scholar] [CrossRef]
- Nyffeler, M.; Moor, H.; Foelix, R.F. Spiders feeding on earthworms. J. Arachnol. 2001, 29, 119–124. [Google Scholar] [CrossRef]
- Ferreira, R.L.; Mendes Martins, V.; Arantes Paixão, E.; Souza Silva, M. Spatial and Temporal Fluctuations of the Abundance of Neotropical Cave-Dwelling Moth Hypena sp. (Noctuidae, Lepidoptera) Influenced by Temperature and Humidity. Subterr. Biol. 2015, 16, 47–60. [Google Scholar] [CrossRef]
- Hesselberg, T.; Simonsen, D.; Juan, C. Do Cave Orb Spiders Show Unique Behavioural Adaptations to Subterranean Life? A Review of Evidence from Cave Orb Spiders. Behaviour 2019, 156, 969–996. [Google Scholar] [CrossRef]
- Novak, T.; Tkavc, T.; Kuntner, M.; Arnett, A.E.; Lipovšek Delakorda, S.; Perc, M.; Janžekovič, F. Niche Partitioning in Orbweaving Spiders Meta menardi and Metellina merianae (Tetragnathidae). Acta Oecologica 2010, 36, 522–529. [Google Scholar] [CrossRef]
- Novak, T.; Sambol, J.; Janžekovič, F. Faunal Dynamics in the Železna Jama Cave. Acta Carsologica 2004, 33, 249–267. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What Shapes the Trophic Niche of European Plethodontid Salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef] [PubMed]
- Manenti, R. Dry stone walls favour biodiversity: A case-study from the Appennines. Biodiv. Conserv. 2014, 23, 1879–1893. [Google Scholar] [CrossRef]
- Ferreira, A.F.; Martins, A.M.d.F.; da Cunha, R.T.; Melo, P.J.; Rodrigues, A.R. Reproductive Biology of Oxychilus (Atlantoxychilus) Spectabilis (Milne-Edwards, 1885) (Gastropoda: Pulmonata): A Gametogenic Approach. Arquipélago Life Mar. Sci. 2013, 30, 11–17. [Google Scholar]
Figure 1.
Differences in microclimatic conditions experienced by the studied species. Boxplots show the species’ range preferences for (A) temperature, (B) humidity, and (C) illuminance recorded within occupied cave sectors. The diagonal bar inside the box represents the median, dots indicate outliers. Significance of pairwise comparisons (Tukey’s HSD) is also shown.
Figure 1.
Differences in microclimatic conditions experienced by the studied species. Boxplots show the species’ range preferences for (A) temperature, (B) humidity, and (C) illuminance recorded within occupied cave sectors. The diagonal bar inside the box represents the median, dots indicate outliers. Significance of pairwise comparisons (Tukey’s HSD) is also shown.


{kind=link}
Table 1.
Species detection probabilities. For each studied species, the table shows the three models ranked by AIC criterion (the best one in bold), along with the effect of the relative covariate and the estimation (±SE) of species detection probability.
Table 1.
Species detection probabilities. For each studied species, the table shows the three models ranked by AIC criterion (the best one in bold), along with the effect of the relative covariate and the estimation (±SE) of species detection probability.
| Species | Model | Type of Covariate | AICc | Covariate Effect (±SE) | Detection Probability (±SE) |
|---|---|---|---|---|---|
| Metellina merianae | 1 | None | 879.14 | ||
| 2 | Sector depth | 853.59 | −0.05 (±0.01) | 0.195 (±0.02) | |
| 3 | Season | 864.93 | |||
| Tegenaria sp. | 1 | None | 786.11 | ||
| 2 | Sector depth | 775.8 | −0.02 (±0.01) | 0.209 (±0.04) | |
| 3 | Season | 785.24 | |||
| Oxychilus oppressus | 1 | None | 842.68 | 0.358 (±0.05) | |
| 2 | Sector depth | 843.19 | |||
| 3 | Season | 845.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Coppari, L.; Manenti, R.; Lunghi, E. Microclimatic Influences on the Abundance of Three Non-Troglobiont Species. Diversity 2024, 16, 186. https://0-doi-org.brum.beds.ac.uk/10.3390/d16030186
AMA Style
Coppari L, Manenti R, Lunghi E. Microclimatic Influences on the Abundance of Three Non-Troglobiont Species. Diversity. 2024; 16(3):186. https://0-doi-org.brum.beds.ac.uk/10.3390/d16030186
Chicago/Turabian StyleCoppari, Luca, Raoul Manenti, and Enrico Lunghi. 2024. "Microclimatic Influences on the Abundance of Three Non-Troglobiont Species" Diversity 16, no. 3: 186. https://0-doi-org.brum.beds.ac.uk/10.3390/d16030186
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.