3.1. Systematic Paleontology
Order Odonata Fabricius, 1793
Clade Isophlebioptera Bechly, 1996
Superfamily Isophlebioidea Handlirsch, 1906
Family Euthemistidae Pritykina, 1968
Type genus: Euthemis Pritykina, 1968; other genera included Sinoeuthemis Li, Nel, and Ren, 2013 and Kidaneuthemis gen. nov.
Emended familial diagnosis. Forewing: IR1 long, basally zigzagged, then smooth and slightly curved distally; postdiscoidal area extremely narrow, with one row of cells; areas between RP2 and IR2, IR2 and RP3/4, RP3/4 and MA, as well as between MA and MP before N separated by only one row of cells apart; area between RP3/4 and MA widen distally, while area between MA and MP slightly narrowed distally.
Other characteristics as in the diagnosis of Pritykina [
3] and Li et al. [
2]: several long intercalary veins between RP1 and IR1, IR1 and RP2, as well as between RP2 and IR2, and between IR2 and RP3/4 (intercalaries parallel to main longitudinal veins without apparent origin on them but originating in cross-venation); extremely narrow postdiscoidal area; not petiolate; numerous secondary antenodal crossveins between anterior wing margin and ScP distal of Ax2; discoidal cells opened in forewing and closed in hind wing; hind wing subdiscoidal area transverse, posteriorly closed, short and broad, with vein CuAb making a strong angle with AA; hind wing ‘gaff’ not very long; RP2 aligned with subnodus; crossveins in hind wing postdiscoidal space are not very long and not oblique.
Genus Kidaneuthemis gen. nov. Liu, Ren et Yang (契丹优美蜓属)
urn:lsid:zoobank.org:act:A4D5A72C-22F1-4BF8-A335-210514B379AA
Type species. Kidaneuthemis ningchengensis gen. et sp. nov. Liu, Ren et Yang
Etymology. The generic name is derived from the Latin “Kidani” (from which the locality of Ningcheng was once the capital of the Liao Dynasty, founded by the Khitan people) and Euthemis (type genus of this family).
Diagnosis. Fore- and hind wings falcate, slender, and longer than 100 mm. Forewing characteristics include (1) Pt short; (2) IR1 long, basally zigzagged, then smooth curved distally; (3) numerous intercalary veins between IRl and RP1, IR1 and RP2, RP2 and IR2, IR2 and RP3/4, as well as RP3/4 and MA, and these areas widened distally; (4) area between MA and MP narrows slightly distally, spaced three rows of cells; (5) discoidal space basally opened; (6) 10 near-parallel intercalary cross-veins in area between the MP and the wing margin; (7) CuAa short, divided into three branches. Hind wing: (1) Pt elongate; (2) IR1 long, basally slightly zigzagged, smooth, and curved distally; (3) presence of four cross-veins in area between the base of RP2 and the base of RP3/4; (4) discoidal cell basally closed; (5) lots of polygonal groups of cells separated by veinlets and intercalary veins between RPl and IR1, IR1 and RP2, RP2 and IR2, IR2 and RP3/4, as well as RP3/4 and MA; (6) cubital area broad, CuAa with three posterior branches; (7) not less than nine longer intercalary cross-veins between the MP and the wing margin. Body: ovipositor weakly curved and elongated, expanding beyond the end of the abdomen.
Kidaneuthemis ningchengensis gen. et sp. nov. Liu, Ren et Yang (宁城契丹优美蜓)
urn:lsid:zoobank.org:act:D5433533-0742-43E2-89DC-C7520E5D23BD
Etymology. The specific epithet is from the locality “Ningcheng county”, where the holotype yielded.
Diagnosis. As for the genus.
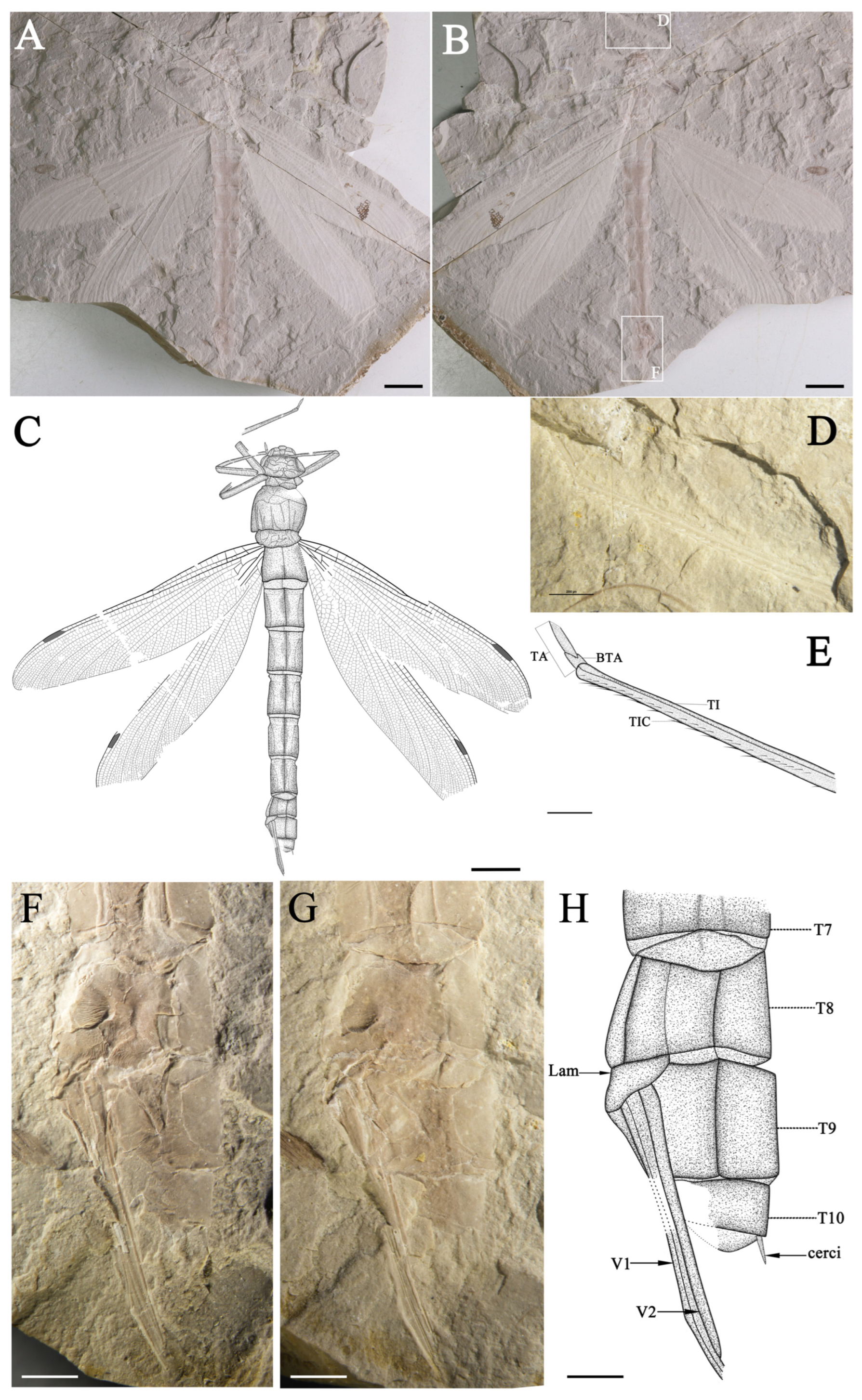
Holotype. No. NCG012-1, No. NCG012-2 (part and counterpart) Female. Imprint of a relatively complete specimen. Fore- and hind wings preserved almost intact with slight overlapped (
Figure 1A–C and
Figure 2).
Type locality and horizon. Jiulongshan Formation, Middle Jurassic (late Callovian); Daohugou Village, Wuhua Township, Ningcheng County, Inner Mongolia, China.
Description
Holotype. Female (hind wing anal margin rounded, preserved with ovipositor). Imprint of a relatively complete specimen. The insect carapace retains the head, thorax, two forelegs, one midleg and one hindleg, the abdomen, the female ovipositor, and the cerci attached to the most terminal abdominal segment. Fore- and hind wings preserved almost intact, with wing bases slightly overlapped.
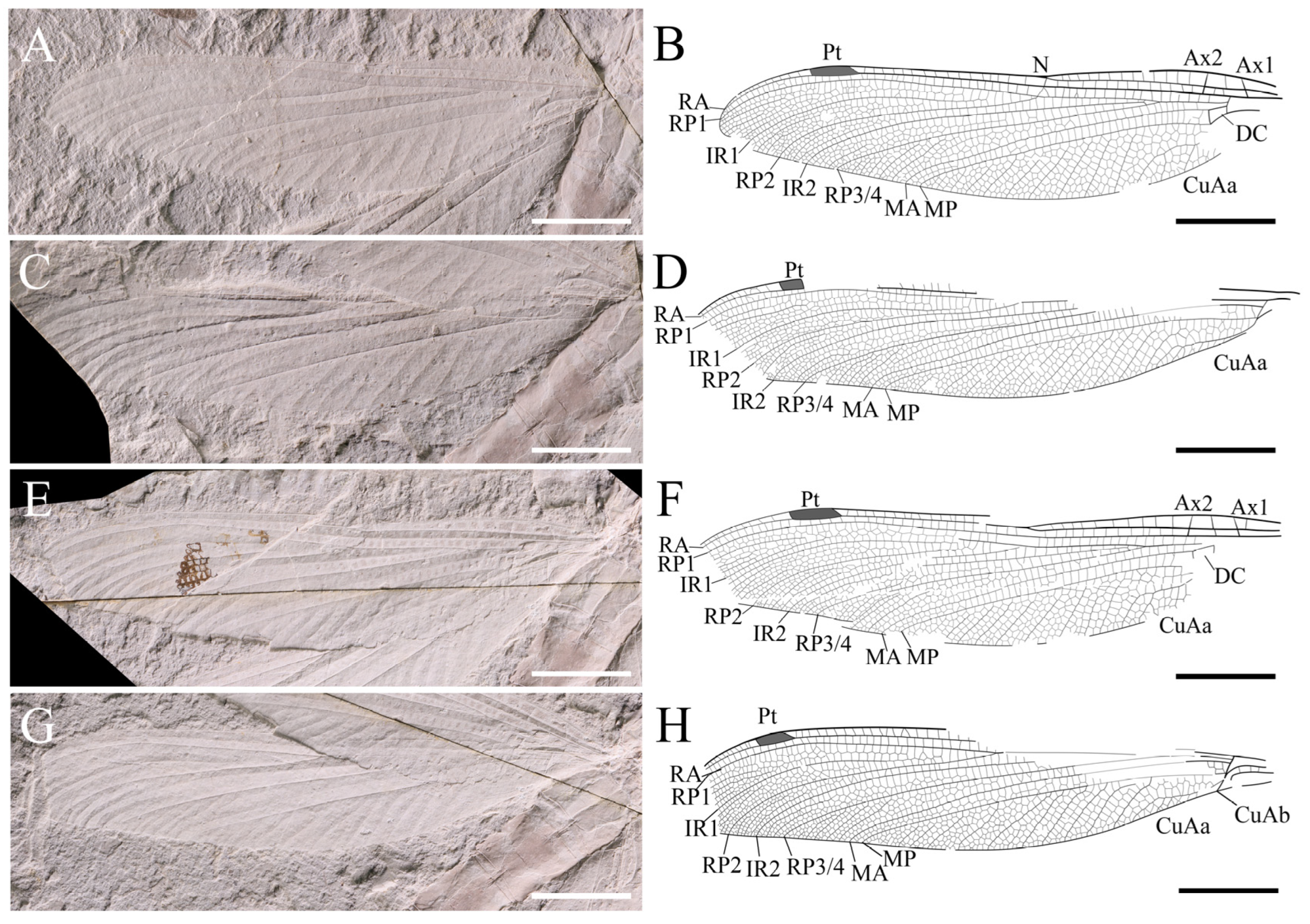
Forewing (left and right wings combined). Wing without petiolates. 117.2 mm long and 24.3 mm wide; ratio of width to length: 0.21; nodus invisible; from pterostigma to apex 14.9 mm; pterostigma 7.0 mm long, 1.8 mm wide. Discoidal space basally opened. Antenodal cross-veins out of sight, while not less than 20 posterior cross-veins before pterostigma. Pt shorter, covering four cells. IR1 long, originating from RP1, zigzagged at base, smooth distally, more or less parallel to RP1; RP2 originating at subnodus; RP2 and IR2 gently curved and parallel in basal part, with only one row of intervening cells, about seven or eight rows of cells in distal part. Area between RP2 and IR2 somewhat widened distally near posterior wing margin; area between RP3/4 and MA separated by only one row of cells at base and then broadened distally, with 14 rows of cells near posterior wing margin; zigzagged intercalary veins present in area between IRl and RP1, IR1 and RP2, RP2 and IR2, IR2 and RP3/4, as well as RP3/4 and MA; post-discoidal area with one row of cells in basal, becoming two rows of cells distally; MA fluctuates slightly at the origin of IR1, causing area between MA and MP to contract near wing margins. MAb 2.2 mm long and partially aligned with basal part of ‘gaff’. MP straight area between MP and CuA with 2–3 rows of cells; and area between MP and wing margin wide, with the widest space not less than seven rows of cells and about 10 intercalary veins originating from MP. CuA divided into CuAa and CuAb. CuAb points towards the posterior margin of the wing and short, CuAa runs almost parallel to the wing margin for some distance, forming a short and narrow cubito-anal area; CuAa branches towards the posterior end, with more than three posterior branches and two rows of cells at its broadest part. Area between MP and posterior wing margin broad, with about eight rows of cells at the widest part, and 10 almost parallel intercalary veins in this area.
Hind wing (left and right wings combined). Wing without short petiole; preserved length 114.2 mm, width 26.2 mm; ratio of width to length: 0.23. Distance from base to nodus 46.5 mm, distance from wing base to arculus 10.3 mm, from arculus to nodus 37.3 mm, from nodus to apex 67.8 mm, from nodus to pterostigma 39.0 mm, from pterostigma to apex 21.2 mm, from nodus to arculus 36.2 mm. Pterostigma 9.2 mm long, 1.7 mm wide. Primary antenodal cross-veins strong; Ax0 not preserved; Ax1 2.9 mm basal of arculus, disposed obliquely to ScP and R + MA; and Ax2 5.3 mm distal of arculus, with inverted obliquity. R + MA nearly straight; RP and MA parallel at base, separated at a slightly obtuse angle from RA in arculus; 14/12 secondary antenodal cross-veins present between C and ScP (left/right hind wing, respectively); thirteen secondary antenodal cross-veins present between ScP and RA distal of Ax2; and 19 antesubnodal cross-veins present between RA and RP, extending from Arc to Sn. Arculus, distinctly angled. Discoidal cells basally closed and free of cross-veins. MP nearly straight, slightly curved upwards two cells away from Ax2, and spaced two–three rows of cells away from CuAa. CuAa short and zigzagged, retaining only three branches and basally somewhat parallel to the posterior wing margin, with three or four rows of cells between. Area between the first and second branches of the left hind wing, CuAa wider than that of the right hind wing. The stem of the second branch of left hind wing, CuAa more curved, while part of right hind wing straight. Nodal well preserved; Sn fluctuates in the middle of the vein and aligns with the slightly tilted Cr. 16 postnodal cross-veins present between C and RA, not aligned with 21 cross-veins between RA and RP1 before Pterostigma. Area between RA and RP1 with one row of cells, two rows of cells exist after Pt. Pterostigma more elongated than the forewing and with a weak Pt-brace. IR1 long, smooth, and slightly zigzagged, originating at RP1 at a distance of six cells from Sn and more or less parallel to RP1. Area between RP1 and IR1, RP2 and IR2, and IR2 and RP3/4 strongly expanded in distal part, and more intercalary veins are present in these areas. RP2 aligned with subnodus; RP2 and IR2 smoothly curved, with only one row of intervening cells in basal part and about eight rows of cells in distal part. IR2 with an apparent origin in RP3/4; base of IR2 with two cells and 3.1 mm distal of base of RP3/4 in right hind wing. Area between RP3/4 and MA, separated by one row of cells basally and distally, strongly widened and presented with five subparallel intercalary veins of different lengths. MA slightly undulated in the middle of N and Pt. Post -discoidal area narrow distally, with one row of cells at the base between MA and MP and three rows of cells at the distal. Area between MP and wing margin board with approximately nine rows of cells spaced apart at the widest point, and about nine or 10 near-parallel intercalary veins in this area.
Body (part and counterpart combined). Total body length (from head to end of abdomen, including anal appendages) 172.0 mm. Head 17.1 mm long and 22.3 mm wide, with compound eyes large, preserved clypeus, post clypeus, and frons. Thorax 24.2 mm long and 20.2 mm wide, relatively well preserved, showing ventral view. Legs elongated, retaining four in part. Forelegs relatively well preserved, femur about 16.3 mm long, tibia 17.4 mm long, preserving two rows of tibial comb; tarsus retain only two-segmented, 1.8 mm long, in which basitarsus 0.84 mm long in counterpart; meso-leg partly preserved, femur about 16.9 mm long, tibia 16.2 mm long; meta-leg party preserved as well, femur more complete, about 18.4 mm, while tibia partially preserved (
Figure 1D,E). The abdomen long and cylindrical, with 122.9 mm long, 18.0 mm at the widest, and 10.4 mm at the narrowest. The first, eighth, ninth, and tenth abdominal segments are shorter than the others. The second abdominal segment is 13.2 mm long, the third 15.5 mm long, and the third to seventh abdominal segments approximately the same length, decreasing in width. The eighth abdominal segment 8.2 mm long, the ninth 9.2 mm long, and the tenth 3.9 mm long. The presence of gonapophyses in the counterpart (valvulae of the cone-shaped ovipositor) between the eighth and ninth abdominal segments 22.7 mm long. The gonapophyses strongly sclerotized, only weakly curved, and extremely elongated, with the ovipositor extending far beyond the end of the abdomen. In addition, on the ventral margin of the last abdominal segment, one tiny structure visible, maybe the cercus (
Figure 1F–H).




{kind=link}
{kind=link}

