

Diversity of Flea Species (Siphonaptera) and Their Vector-Borne Pathogens from Bats (Chiroptera) in Lithuania
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Flea Collection and Species Identification
2.2. Pathogen DNA Amplification
2.3. Sequencing
3. Results
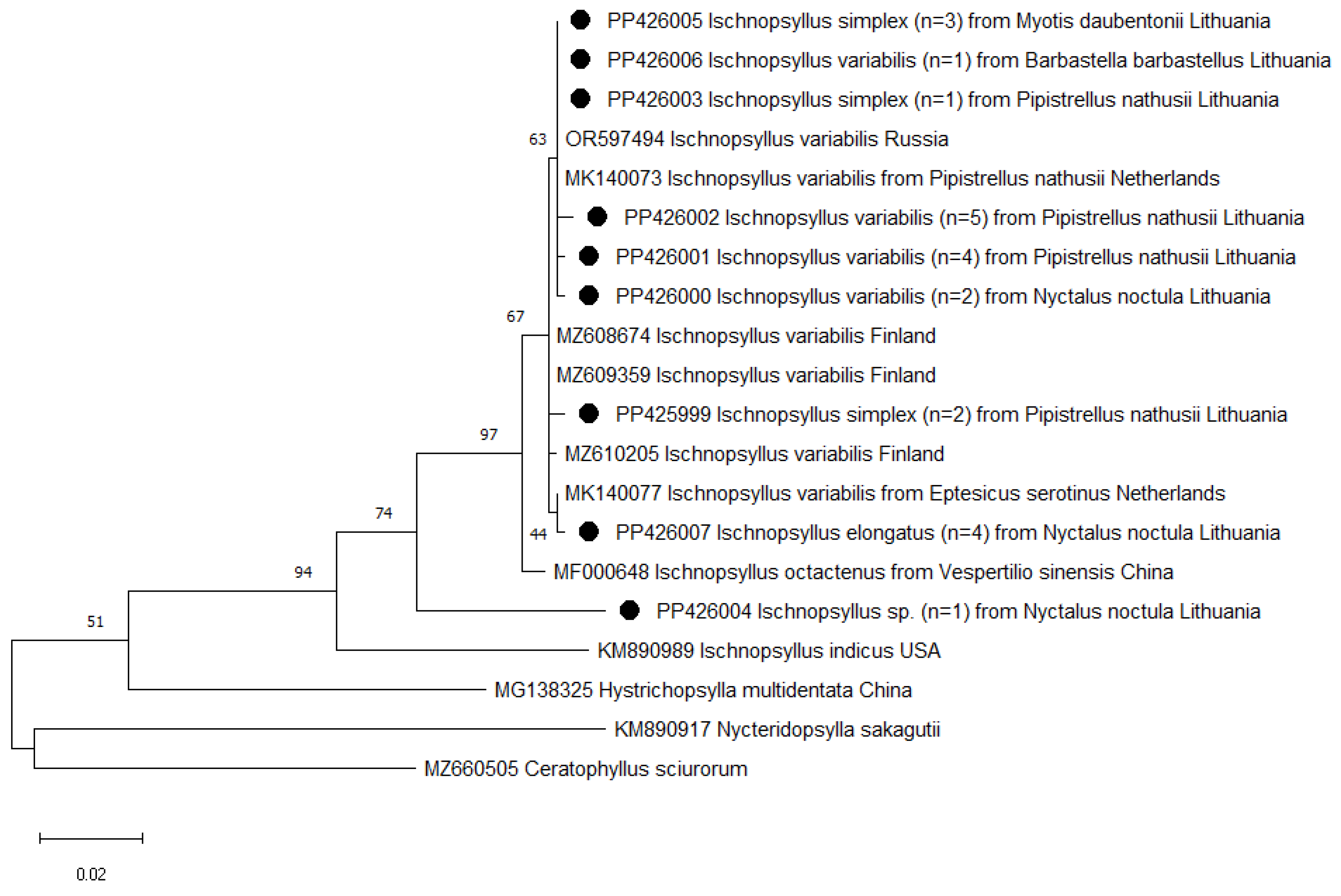
3.1. Identification of Flea Species
3.2. Detection of Pathogens
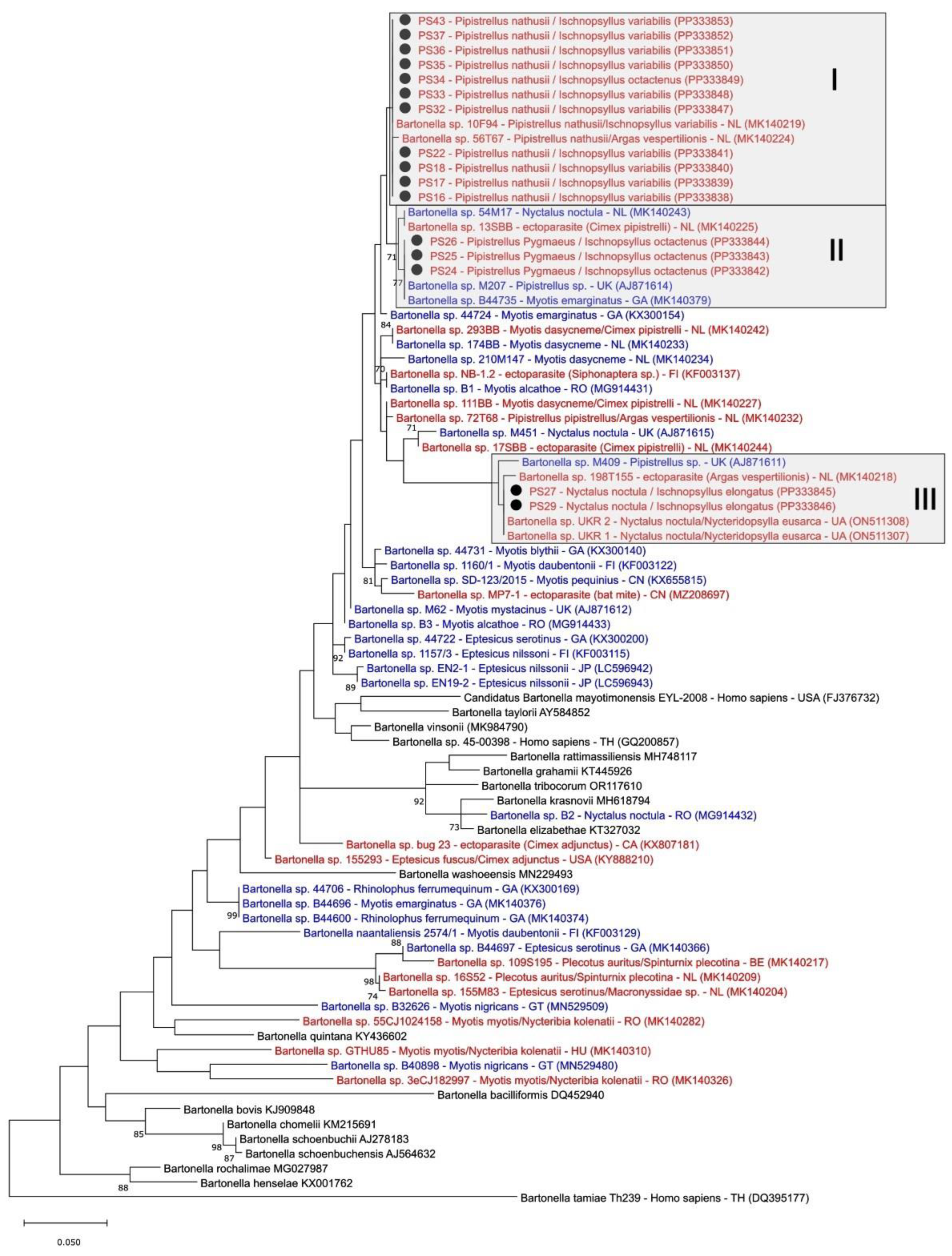
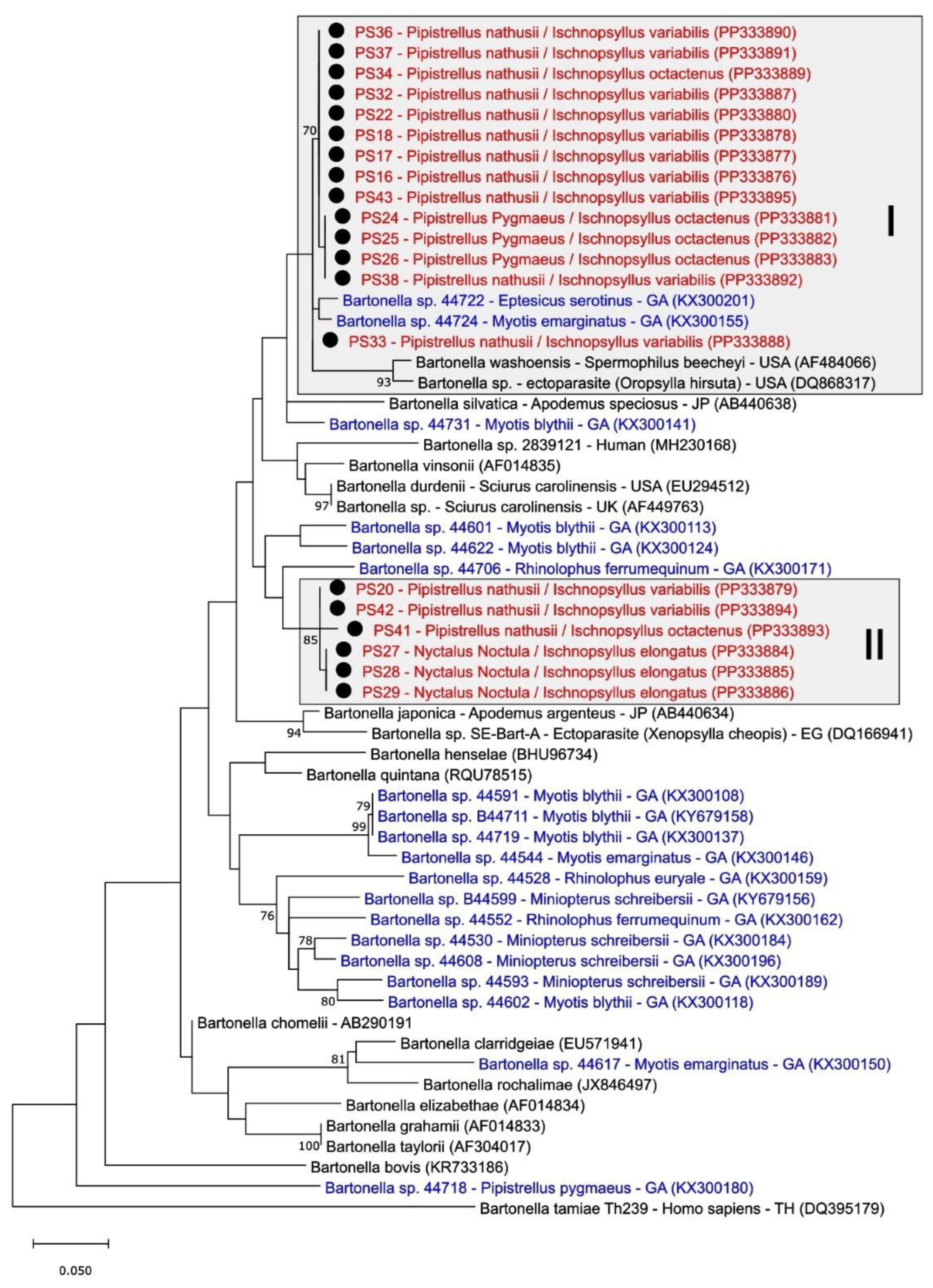
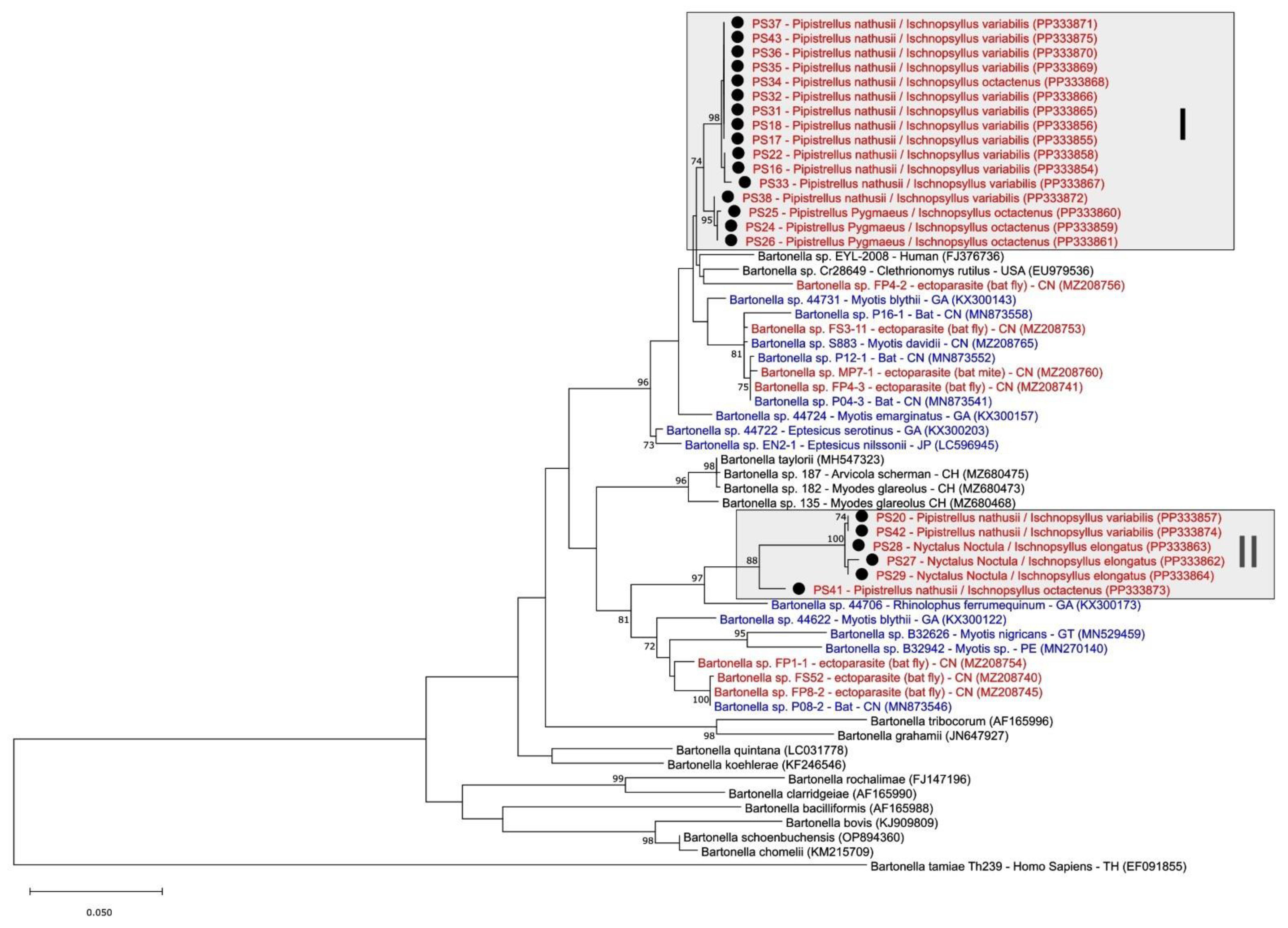
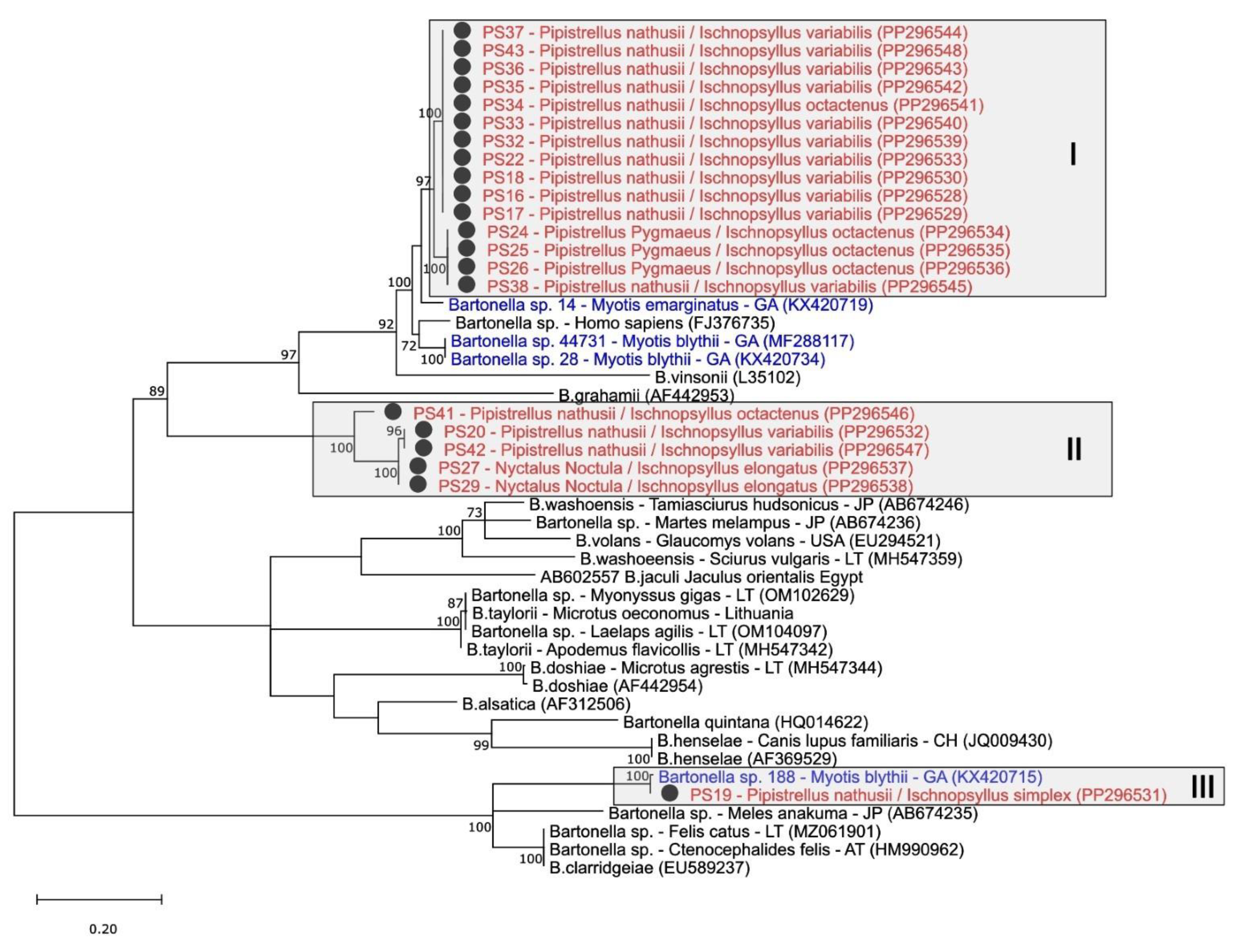
3.3. Phylogenetic Analysis of Bartonella spp. in Fleas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bitam, I.; Dittmar, K.; Parola, P.; Whiting, M.F.; Raoult, D. Fleas and flea-borne diseases. Int. J. Infect. Dis. 2010, 14, e667–e676. [Google Scholar] [CrossRef]
- Whiting, M.F.; Whiting, A.S.; Hastriter, M.W.; Dittmar, K. A molecular phylogeny of fleas (Insecta: Siphonaptera): Origins and host associations. Cladistics 2008, 24, 677–707. [Google Scholar] [CrossRef]
- Whitaker, A.P. Fleas (Siphonaptera) (Handbooks for the Identification of British Insects), 2nd ed.; Royal Entomological Society: London, UK, 2007; pp. 44–57. [Google Scholar]
- Bejec, G.A.; Bucol, L.A.; Ancog, A.B.; Pagente, A.C.; Panerio, J.J.M.; Bejec, A.L.N.; Belanizo, J.D.; Tuastomban, D.J.S.; Jose, R.P. Diversity of bat ectoparasites from the caves of selected key biodiversity areas in Central Visayas, Philippines. Biodiversitas 2023, 24, 1693–1703. [Google Scholar] [CrossRef]
- Szentiványi, T.; Heintz, A.C.; Markotter, W.; Wassef, J.; Christe, P.; Glaizot, O. Vector-borne protozoan and bacterial pathogen occurrence and diversity in ectoparasites of the Egyptian Rousette bat. Med. Vet. Entomol. 2023, 37, 189–194. [Google Scholar] [CrossRef]
- Munshi-South, J.; Wilkinson, G.S. Bats and birds: Exceptional longevity despite high metabolic rates. Ageing. Res. Rev. 2010, 9, 12–19. [Google Scholar] [CrossRef]
- Szentivanyi, T.; McKee, C.; Jones, G.; Foster, J.T. Trends in bacterial pathogens of bats: Global distribution and knowledge gaps. Transbound. Emerg. Dis. 2023, 2023, 9285855. [Google Scholar] [CrossRef]
- Ahn, M.; Wang, L.F. Translation from bats to humans beyond infectious diseases. J. Exp. Med. 2021, 218, e20211223. [Google Scholar] [CrossRef]
- Han, H.J.; Li, Z.M.; Li, X.; Liu, J.X.; Peng, Q.M.; Wang, R.; Gu, X.L.; Jiang, Y.; Zhou, C.M.; Li, D.; et al. Bats and their ectoparasites (Nycteribiidae and Spinturnicidae) carry diverse novel Bartonella genotypes, China. Transbound. Emerg. Dis. 2022, 69, e845–e858. [Google Scholar] [CrossRef]
- Pauza, D.H.; Pauziene, A.N. Bats of Lithuania: Distribution, status and protection. Mammal. Rev. 1998, 28, 53–68. [Google Scholar] [CrossRef]
- Lithuanian Ornithological Society. New Bat Species Recorded in Lithuania—Greater Mouse-Eared Bat (Myotis myotis). Available online: https://www.birdlife.lt/new-bat-species-recorded-in-lithuania-greater-mous (accessed on 5 December 2023).
- Baranauskas, K. Bats (Chiroptera) found in bat boxes in Southeastern Lithuania. Ekologija 2007, 53, 34–37. [Google Scholar]
- Jaunbauere, G.; Salmane, I.; Spungis, V. Occurrence of bat ectoparasites in Latvia. Latv. Entomol. 2008, 45, 38–42. [Google Scholar]
- Burazerovic, J.; Orlova, M.; Obradovic, M.; Cirovic, D.; Tomanovic, S. Patterns of abundance and host specificity of bat ectoparasites in the Central Balkans. J. Med. Entomol. 2018, 55, 20–28. [Google Scholar] [CrossRef]
- Léger, C. Bat parasites (Acari, Anoplura, Cestoda, Diptera, Hemiptera, Nematoda, Siphonaptera, Trematoda) in France (1762–2018): A literature review and contribution to a checklist. Parasite 2020, 27, 61. [Google Scholar] [CrossRef]
- Ivanova-Aleksandrova, N.; Dundarova, H.; Neov, B.; Emilova, R.; Georgieva, I.; Antova, R.; Kirov, K.; Pikula, J.; Zukalová, K.; Zukal, J. Ectoparasites of cave-dwelling bat species in Bulgaria. Proc. Zool. Soc. 2022, 75, 463–468. [Google Scholar] [CrossRef]
- Veikkolainen, V.; Vesterinen, E.J.; Lilley, T.M.; Pulliainen, A.T. Bats as reservoir hosts of human bacterial pathogen, Bartonella mayotimonensis. Emerg. Infect. Dis. 2014, 20, 960–967. [Google Scholar] [CrossRef]
- Vlaschenko, A.; Răileanu, C.; Tauchmann, O.; Muzyka, D.; Bohodist, V.; Filatov, S.; Rodenko, O.; Tovstukha, I.; Silaghi, C. First data on bacteria associated with bat ectoparasites collected in Kharkiv oblast, Northeastern Ukraine. Parasit. Vectors. 2022, 15, 443. [Google Scholar] [CrossRef]
- Rosicky, B. Fauna ČSR Blechy—Aphaniptera; ČSAV: Praha, Czech Republic, 1957. (In Czech) [Google Scholar]
- Rijpkema, S.; Golubic, D.; Molkenboer, M.; Verbeek-De Kruif, N.; Schellekens, J. Identification of four genomic groups of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks collected in a Lyme borreliosis endemic region of Northern Croatia. Exp. Appl. Acarol. 1996, 20, 23–30. [Google Scholar] [CrossRef]
- Brinck-Lindroth, G.; Smit, F.G.A.M. The Fleas (Siphonaptera) of Fennoscandia and Denmark; Brill, Leiden: Boston, MA, USA, 2007. [Google Scholar]
- Whiting, M.F. Mecoptera is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zool. Scr. 2002, 31, 93–104. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ražanskė, I.; Rosef, O.; Radzijevskaja, J.; Bratchikov, M.; Griciuvienė, L.; Paulauskas, A. Prevalence and co-infection with tick-borne Anaplasma phagocytophilum and Babesia spp. in red deer (Cervus elaphus) and roe deer (Capreolus capreolus) in Southern Norway. Int. J. Parasitol. 2019, 8, 127–134. [Google Scholar]
- de la Fuente, J.; Massung, R.F.; Wong, S.J.; Chu, F.K.; Lutz, H.; Meli, M.; von Loewenich, F.D.; Grzeszczuk, A.; Torina, A.; Caracappa, S.; et al. Sequence analysis of the msp4 gene of Anaplasma phagocytophilum strains. J. Clin. Microbiol. 2005, 43, 1309–1317. [Google Scholar] [CrossRef]
- Bown, K.J.; Lambin, X.; Ogden, N.H.; Petrovec, M.; Shaw, S.E.; Woldehiwet, Z.; Birtles, R.J. High-resolution genetic fingerprinting of European strains of Anaplasma phagocytophilum by use of multilocus variable-number tandem-repeat analysis. J. Clin. Microbiol. 2007, 45, 1771–1776. [Google Scholar] [CrossRef]
- Massung, R.F.; Slater, K.; Owens, J.H.; Nicholson, W.L.; Mather, T.N.; Solberg, V.B.; Olson, J.G. Nested PCR assay for detection of granulocytic ehrlichiae. J. Clin. Microbiol. 1998, 36, 1090–1095. [Google Scholar] [CrossRef]
- Sumner, J.W.; Nicholson, W.L.; Massung, R.F. PCR amplification and comparison of nucleotide sequences from the groESL heat shock operon of Ehrlichia species. J. Clin. Microbiol. 1997, 35, 2087–2092. [Google Scholar] [CrossRef]
- Liz, J.S.; Sumner, J.W.; Pfister, K.; Brossard, M. PCR detection and serological evidence of granulocytic ehrlichial infection in roe deer (Capreolus capreolus) and chamois (Rupicapra rupicapra). J Clin Microbiol. 2002, 40, 892–897. [Google Scholar] [CrossRef]
- Rar, V.A.; Epikhina, T.I.; Livanova, N.N.; Panov, V.V. Genetic diversity of Babesia in Ixodes persulcatus and small mammals from North Ural and West Siberia, Russia. Parasitol. 2011, 138, 175–182. [Google Scholar] [CrossRef]
- Mardosaitė-Busaitienė, D.; Radzijevskaja, J.; Balčiauskas, L.; Bratchikov, M.; Jurgelevičius, V.; Paulauskas, A. Prevalence and diversity of Bartonella species in small rodents from coastal and continental areas. Sci. Rep. 2019, 9, 9285855. [Google Scholar] [CrossRef]
- Kaewmongkol, G.; Kaewmongkol, S.; Burmej, H.; Bennett, M.D.; Fleming, P.A.; Adams, P.J.; Wayne, A.F.; Ryan, U.; Irwin, P.J.; Fenwick, S.G. Diversity of Bartonella species detected in arthropod vectors from animals in Australia. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 411–417. [Google Scholar] [CrossRef]
- Norman, A.F.; Regnery, R.; Jameson, P.; Greene, C.; Krause, D.C. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J Clin Microbiol. 1995, 33, 1797–1803. [Google Scholar] [CrossRef]
- Kim, C.M.; Kim, J.Y.; Yi, Y.H.; Lee, M.J.; Cho, M.R.; Shah, D.H.; Klein, T.A.; Kim, H.C.; Song, J.W.; Chong, S.T.; et al. Detection of Bartonella species from ticks, mites and small mammals in Korea. J. Vet. Sci. 2005, 6, 327–334. [Google Scholar] [CrossRef]
- Kaewmongkol, G. Detection and Characterization of Bartonella Species in Western Australia. Ph.D. Thesis, Murdoch University, Perth, Australia, 2012. [Google Scholar]
- Sakalauskas, P.; Lipatova, I.; Radzijevskaja, J.; Paulauskas, A. Pathogen screeningin the red fox (Vulpes vulpes) from lithuania. Biologija 2020, 65, 283–295. [Google Scholar] [CrossRef]
- Demaerschalck, I.; Ben Messaoud, A.; De Kesel, M.; Hoyois, B.; Lobet, Y.; Hoet, P.; Bigaignon, G.; Bollen, A.; Godfroid, E. Simultaneous presence of different Borrelia burgdorferi genospecies in biological fluids of Lyme disease patients. J. Clin. Microbiol. 1995, 33, 602–608. [Google Scholar] [CrossRef]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiol. 2004, 150, 1741–1755. [Google Scholar] [CrossRef]
- Wodecka, B.; Rymaszewska, A.; Sawczuk, M.; Skotarczak, B. Detectability of tick borne agents DNA in the blood of dogs, undergoing treatment for borreliosis. Ann. Agric. Environ. Med. 2009, 16, 914. [Google Scholar]
- Miťková, K.; Berthová, L.; Kalúz, S.; Kazimírová, M.; Burdová, L.; Kocianová, E. First detections of Rickettsia helvetica and R. monacensis in ectoparasitic mites (Laelapidae and Trombiculidae) infesting rodents in south-western Slovakia. Parasitol. Res. 2015, 114, 2465–2472. [Google Scholar] [CrossRef]
- Varanat, M.; Maggi, R.G.; Linder, K.E.; Breitschwerdt, E.B. Molecular prevalence of Bartonella, Babesia, and hemotropic Mycoplasma sp. in dogs with splenic disease. J. Vet. Intern. Med. 2011, 25, 1284–1291. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Krištofík, J.; Danko, Š. Arthropod ectoparasites (Acarina, Heteroptera, Diptera, Siphonaptera) of bats in Slovakia. Vespertilio 2012, 16, 167–189. [Google Scholar]
- Azrizal-Wahid, N.; Sofian-Azirun, M.; Low, V.L. New insights into the haplotype diversity of the cosmopolitan cat flea Ctenocephalides felis (Siphonaptera: Pulicidae). Vet. Parasitol. 2020, 281, 109102. [Google Scholar] [CrossRef]
- Zurita, A.; García-Sánchez, Á.M.; Cutillas, C. Comparative molecular and morphological study of Stenoponia tripectinata tripectinata (Siphonaptera: Stenoponiidae) from the Canary Islands and Corsica. Bull. Entomol. Res. 2022, 112, 681–690. [Google Scholar] [CrossRef]
- Hornok, S.; Beck, R.; Farkas, R.; Grima, A.; Otranto, D.; Kontschán, J.; Takács, N.; Horváth, G.; Szőke, K.; Szekeres, S.; et al. High mitochondrial sequence divergence in synanthropic flea species (Insecta: Siphonaptera) from Europe and the Mediterranean. Parasit. Vectors. 2018, 11, 221. [Google Scholar] [CrossRef]
- Bossard, R.L.; Lareschi, M.; Urdapilleta, M.; Cutillas, C.; Zurita, A. Flea (Insecta: Siphonaptera) family diversity. Diversity 2023, 15, 1096. [Google Scholar] [CrossRef]
- Gerth, M.; Saeed, A.; White, J.A.; Bleidorn, C. Extensive screen for bacterial endosymbionts reveals taxon-specific distribution patterns among bees (Hymenoptera, Anthophila). FEMS Microbiol. Microb. Ecol. 2015, 91, fiv047. [Google Scholar] [CrossRef]
- Bleidorn, C.; Henze, K. A new primer pair for barcoding of bees (Hymenoptera: Anthophila) without amplifying the orthologous coxA gene of Wolbachia bacteria. BMC Res. Notes. 2021, 14, 427. [Google Scholar] [CrossRef]
- Dittmar, K.; Whiting, M.F. New Wolbachia endosymbionts from Nearctic and Neotropical fleas (Siphonaptera). J. Parasitol. 2004, 90, 953–957. [Google Scholar] [CrossRef]
- Li, K.; Stanojević, M.; Stamenković, G.; Ilić, B.; Paunović, M.; Lu, M.; Pešić, B.; Đurić Maslovara, I.; Siljic, M.; Cirkovic, V.; et al. Insight into diversity of bacteria belonging to the order Rickettsiales in 9 arthropods species collected in Serbia. Sci. Rep. 2019, 9, 18680. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Kovács, R.; Meli, M.L.; Gönczi, E.; Hofmann-Lehmann, R.; Kontschán, J.; Gyuranecz, M.; Dán, A.; Molnár, V. First detection of bartonellae in a broad range of bat ectoparasites. Vet. Microbiol. 2012, 159, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Urushadze, L.; Bai, Y.; Osikowicz, L.; McKee, C.; Sidamonidze, K.; Putkaradze, D.; Imnadze, P.; Kandaurov, A.; Kuzmin, I.; Kosoy, M. Prevalence, diversity, and host associations of Bartonella strains in bats from Georgia (Caucasus). PLoS Negl. Trop. Dis. 2017, 11, e0005428. [Google Scholar] [CrossRef]
- Reeves, W.K.; Rogers, T.E.; Durden, L.A.; Dasch, G.A. Association of Bartonella with the fleas (Siphonaptera) of rodents and bats using molecular techniques. J. Vector Ecol. 2007, 32, 118–122. [Google Scholar] [CrossRef]
- Nabeshima, K.; Sato, S.; Brinkerhoff, R.J.; Amano, M.; Kabeya, H.; Itou, T.; Maruyama, S. Prevalence and Genetic Diversity of Bartonella Spp. in Northern Bats (Eptesicus nilssonii) and Their Blood-Sucking Ectoparasites in Hokkaido, Japan. Microb. Ecol. 2023, 85, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Kuang, G.; Zhang, J.; Yang, W.; Pan, H.; Han, X.; Yang, L.; Wang, J.; Yang, T.; Song, Z.; Feng, Y.; et al. Molecular Detection and Phylogenetic Analyses of Diverse Bartonella Species in Bat Ectoparasites Collected from Yunnan Province, China. Pathogens. 2022, 11, 1283. [Google Scholar] [CrossRef]
- Han, H.J.; Wen, H.L.; Zhao, L.; Liu, J.W.; Luo, L.M.; Zhou, C.M.; Qin, X.R.; Zhu, Y.L.; Zheng, X.X.; Yu, X.J. Novel Bartonella Species in Insectivorous Bats, Northern China. PLoS ONE 2017, 12, e0167915. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.D.; Kosoy, M.Y.; Bai, Y.; Osikowicz, L.M.; Franka, R.; Gilbert, A.T.; Boonmar, S.; Rupprecht, C.E.; Peruski, L.F. Diversity and phylogenetic relationships among Bartonella strains from Thai bats. PLoS ONE 2017, 12, e0181696. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, P.; Marinho Torres, J.; Perles, L.; Lourenço, E.C.; Herrera, H.M.; de Oliveira, C.E.; Zacarias Machado, R.; André, M.R. Intra- and Inter-Host Assessment of Bartonella Diversity with Focus on Non-Hematophagous Bats and Associated Ectoparasites from Brazil. Microorganisms 2020, 8, 1822. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bat Species | Flea Number, Sex, and Species |
|---|---|
| P. nathusii | 3 ♂, 10 ♀ I. octactenus |
| 3 ♂, 17 ♀ I. variabilis | |
| 2 ♂, 1 ♀ I. simplex | |
| P. pygmaeus | 0 ♂, 3 ♀ I. octactenus |
| N. noctula | 1 ♂, 0 ♀ N. eusarca |
| 1 ♂, 3 ♀ I. elongatus | |
| 1 ♂, 2 ♀ I. variabilis | |
| 0 ♂, 1 ♀ Ischnopsyllus sp. | |
| E. nilssonii | 2 ♂, 3 ♀ N. pentactena |
| M. daubentonii | 2 ♂, 1 ♀ I. simplex |
| B. barbastellus | 1 ♂, 0 ♀ I. hexactenus |
| 0 ♂, 1 ♀ I. variabilis | |
| Total number (female/male ratio) | 16 ♂, 42 ♀ (2.625, χ2 = 58.34) |
| Pathogen | Gene | I.v. * n/N | I.o. * n/N | I.s. * n/N | I.e. * n/N | I.h. * n/N | N.p. * n/N | N.e. * n/N | I.sp. * n/N | Total n/N |
|---|---|---|---|---|---|---|---|---|---|---|
| Anaplasma | RT-PCR msp2 | 0/24 | 1/16 | 0/6 | 0/4 | 0/1 | 1/5 | 0/1 | 0/1 | 2/58 |
| msp4 | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| 16S rRNA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| groESL | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| Borrelia | RT-PCR 23S rRNA | 1/24 | 0/16 | 0/6 | 0/4 | 0/1 | 1/5 | 0/1 | 0/1 | 2/58 |
| ospA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| 16S-23S rrs-rrlA ITS | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| flA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| Babesia | RT-PCR 18S rRNA | 3/24 | 0/16 | 1/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 4/58 |
| 18S rRNA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| Bartonella | RT-PCR ssrA | 14/24 | 6/16 | 1/6 | 3/4 | 0/1 | 0/5 | 0/1 | 0/1 | 24/58 |
| 16S-23S rRNA ITS | 14/24 | 6/16 | 1/6 | 2/4 | 0/1 | 0/5 | 0/1 | 0/1 | 23/58 | |
| gltA | 13/24 | 5/16 | 0/6 | 2/4 | 0/1 | 0/5 | 0/1 | 0/1 | 20/58 | |
| rpoB | 14/24 | 6/16 | 0/6 | 3/4 | 0/1 | 0/5 | 0/1 | 0/1 | 23/58 | |
| groEL | 14/24 | 6/16 | 0/6 | 3/4 | 0/1 | 0/5 | 0/1 | 0/1 | 23/58 | |
| Rickettsia | RT-PCR gltA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 |
| gltA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 | |
| Mycoplasma | 16S rRNA | 0/24 | 0/16 | 0/6 | 0/4 | 0/1 | 0/5 | 0/1 | 0/1 | 0/58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakalauskas, P.; Lipatova, I.; Griciuvienė, L.; Ražanskė, I.; Snegiriovaitė, J.; Paulauskas, A. Diversity of Flea Species (Siphonaptera) and Their Vector-Borne Pathogens from Bats (Chiroptera) in Lithuania. Diversity 2024, 16, 192. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040192
Sakalauskas P, Lipatova I, Griciuvienė L, Ražanskė I, Snegiriovaitė J, Paulauskas A. Diversity of Flea Species (Siphonaptera) and Their Vector-Borne Pathogens from Bats (Chiroptera) in Lithuania. Diversity. 2024; 16(4):192. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040192
Chicago/Turabian StyleSakalauskas, Povilas, Indrė Lipatova, Loreta Griciuvienė, Irma Ražanskė, Justina Snegiriovaitė, and Algimantas Paulauskas. 2024. "Diversity of Flea Species (Siphonaptera) and Their Vector-Borne Pathogens from Bats (Chiroptera) in Lithuania" Diversity 16, no. 4: 192. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040192