
Biodiversity of Gelatinous Organisms in the Western Adriatic Sea and Identification of Their Echo Traces in Acoustic Data

 ,
,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
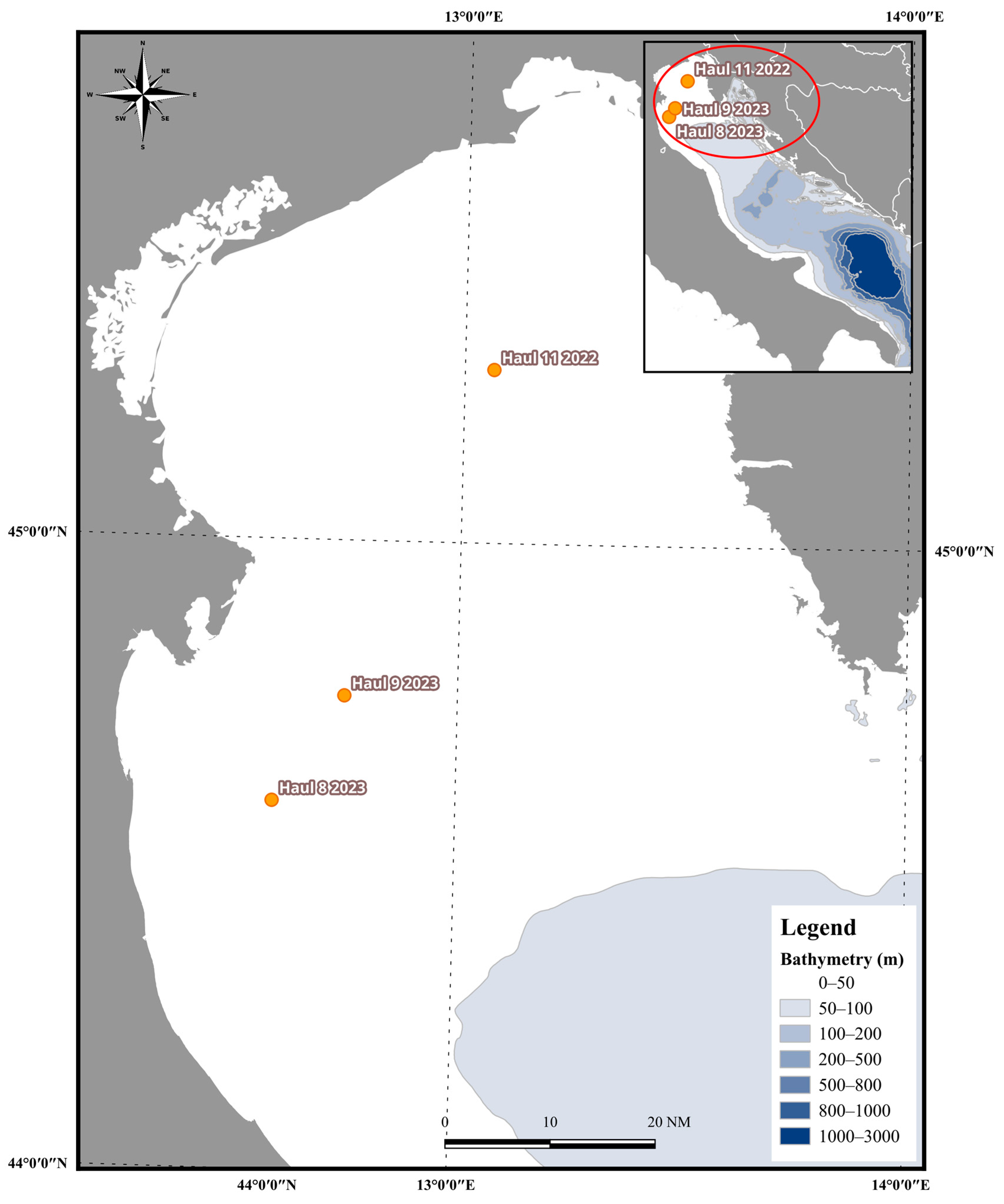
2. Materials and Methods
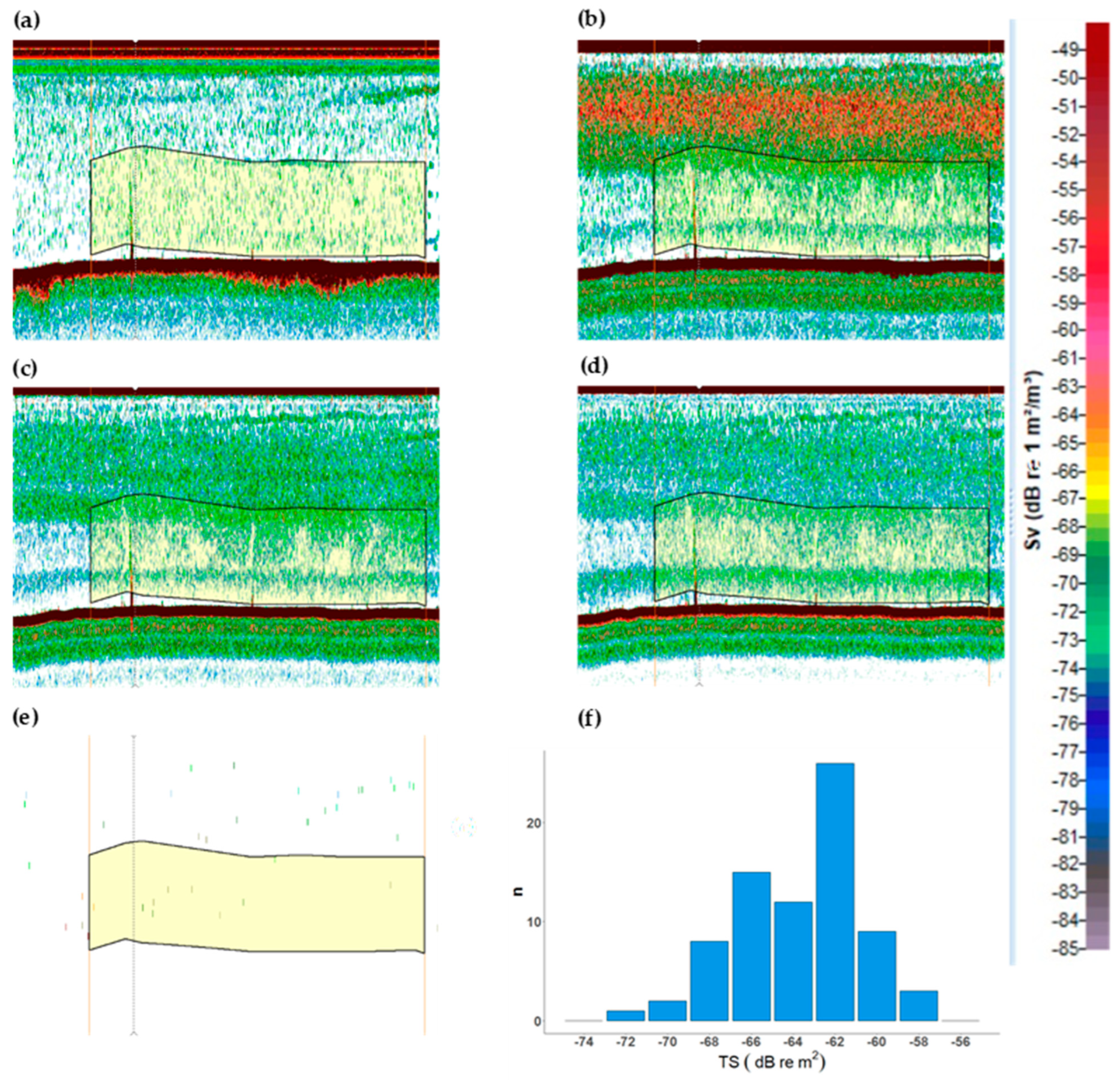
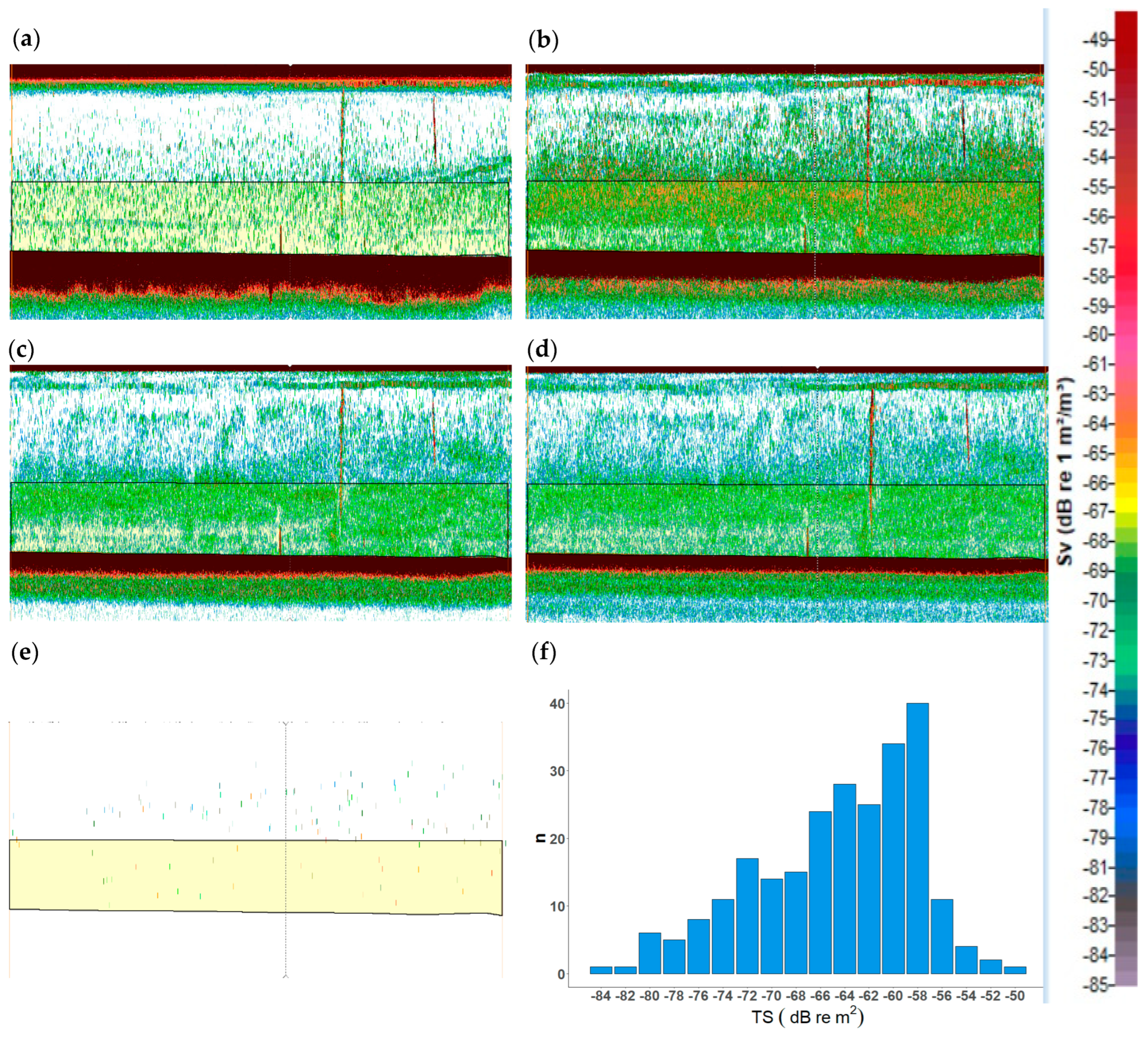
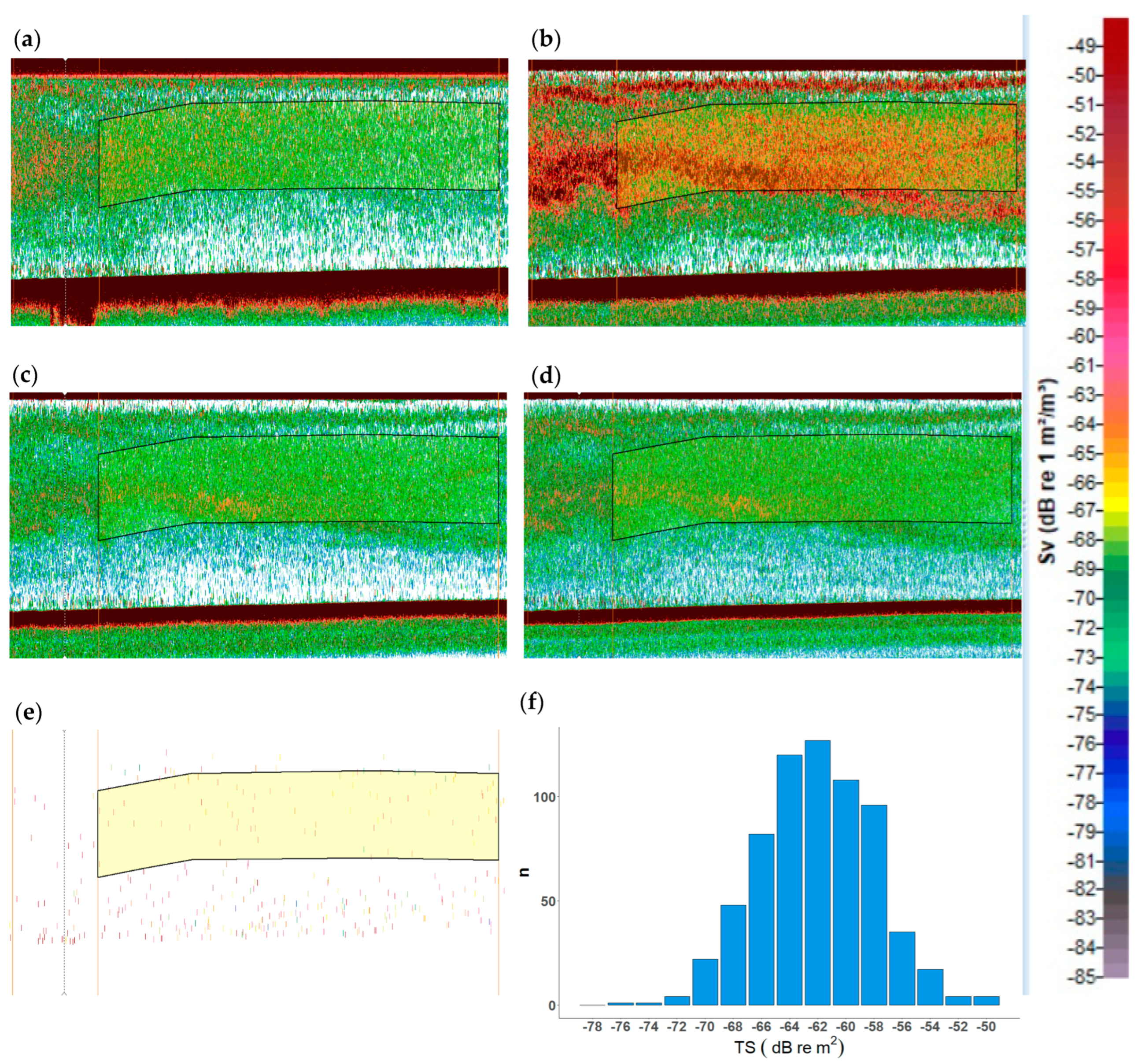
2.1. Echogram Analysis of Jellyfish Monospecific Hauls
2.2. Abundance of Salps and Jellyfish in Relation to Environmental Parameters
3. Results
3.1. Jellyfish Echo Trace Identification
3.2. Abundance of Salps and Jellyfish in Relation to Environmental Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brotz, L.; Cheung, W.W.L.; Kleisner, K.; Pakhomov, E.; Pauly, D. Increasing jellyfish populations: Trends in Large Marine Ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef]
- Condon, R.H.; Duarte, C.M.; Pitt, K.A.; Robinson, K.L.; Lucas, C.H.; Sutherland, K.R.; Mianzan, H.W.; Bogeberg, M.; Purcell, J.E.; Decker, M.B.; et al. Recurrent jellyfish blooms are a consequence of global oscillations. Proc. Natl. Acad. Sci. USA 2013, 110, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.E. Jellyfish blooms: Are populations increasing globally in response to changing ocean conditions? Hydrobiologia 2001, 451, 55–68. [Google Scholar] [CrossRef]
- Sun-Li, B. Impact of Hypoxia on the Vertical Distribution of Jellyfish in the Eastern Tropical Northern Pacific. Proceedings from the University of Washington School of Oceanography Senior Thesis, Academic Year 2011–2012. Available online: https://digital.lib.washington.edu/researchworks/bitstream/handle/1773/20062/444Thesis_Beatteay.pdf?sequence=1 (accessed on 11 August 2022).
- Wang, G.; Zhen, Y.; Yu, Z.; Shi, Y.; Zhao, Q.; Wang, J.; Mi, T. The physiological and molecular response of Aurelia sp.1 under hypoxia. Sci. Rep. 2017, 7, 1558. [Google Scholar] [CrossRef] [PubMed]
- Shoji, J.; Masuda, R.; Yamashita, Y.; Tanaka, M. Effect of low dissolved oxygen concentrations on behavior and predation rates on red sea bream Pagrus major larvae by the jellyfish Aurelia aurita and by juvenile Spanish mackerel Scomberomorus niphonius. Mar. Biol. 2005, 147, 863–868. [Google Scholar] [CrossRef]
- Møller, L.F.; Riisgård, H.U. Impact of jellyfish and mussels on algal blooms caused by seasonal oxygen depletion and nutrient release from the sediment in a Danish fjord. J. Exp. Mar. Biol. Ecol. 2007, 351, 92–105. [Google Scholar] [CrossRef]
- Boero, F. Review of Jellyfish Blooms in the Mediterranean and Black Sea. Studies and Reviews. General Fisheries Commission for the Mediterranean (GFCM); N. 92; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; 53p. [Google Scholar]
- Stoltenberg, I.; Dierking, J.; Müller-Navarra, D.C.; Javidpour, J. Review of jellyfish trophic interactions in the Baltic Sea. Mar. Biol. Res. 2021, 17, 311–326. [Google Scholar] [CrossRef]
- Milisenda, G.; Rosa, S.; Fuentes, V.L.; Boero, F.; Guglielmo, L.; Purcell, J.E.; Piraino, S. Jellyfish as Prey: Frequency of Predation and Selective Foraging of Boops boops (Vertebrata, Actinopterygii) on the Mauve Stinger Pelagia noctiluca (Cnidaria, Scyphozoa). PLoS ONE 2014, 9, e94600. [Google Scholar] [CrossRef] [PubMed]
- Schnedler-Meyer, N.A.; Mariani, P.; Kiørboe, T. The global susceptibility of coastal forage fish to competition by large jellyfish. Proc. R. Soc. B 2016, 283, 20161931. [Google Scholar] [CrossRef] [PubMed]
- Báez, J.C.; Pennino, M.G.; Albo-Puigserver, M.; Coll, M.; Giraldez, A.; Bellido, J.M. Effects of environmental conditions and jellyfish blooms on small pelagic fish and fisheries from the Western Mediterranean Sea. Estuar. Coast. Shelf Sci. 2022, 264, 107699. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Milisenda, G.; Basso, L.; Doyle, T.K.; Leone, A.; Piraino, S. Jellyfish Impacts on Marine Aquaculture and Fisheries. Rev. Fish. Sci. Aquac. 2021, 29, 242–259. [Google Scholar] [CrossRef]
- Kogovšek, T.; Bogunović, B.; Malej, A. Recurrence of bloom-forming scyphomedusae: Wavelet analysis of a 200-year time series. In Jellyfish Blooms: New Problems and Solutions; Purcell, J.E., Angel, D.L., Eds.; Developments in Hydrobiology 212; Springer: Dordrecht, The Netherlands, 2010; Volume 212. [Google Scholar] [CrossRef]
- Vodopivec, M.; Peliz, A.J.; Malej, A. Offshore marine constructions as propagators of moon jellyfish dispersal. Environ. Res. Lett. 2017, 12, 084003. [Google Scholar] [CrossRef]
- Leonori, I.; Ticina, V.; Giannoulaki, M.; Hattab, T.; Iglesias, M.; Bonanno, A.; Costantini, I.; Canduci, G.; Machias, A.; Ventero, A.; et al. History of hydroacoustic surveys of small pelagic fish species in the European Mediterranean Sea. Medit. Mar. Sci. 2021, 22, 751–768. [Google Scholar] [CrossRef]
- Yoon, E.-A.; Hwang, D.-J.; Chae, J.; Yoon, W.D.; Lee, K. Behavior and frequency analysis of Aurelia aurita by using in situ target strength at a port in southwestern Korea. Ocean Sci. J. 2018, 53, 53–62. [Google Scholar] [CrossRef]
- Mutlu, E. Target strength of the common jellyfish (Aurelia aurita): A preliminary experimental study with a dual-beam acoustic system. ICES J. Mar. Sci. 1996, 53, 309–311. [Google Scholar] [CrossRef]
- Brierley, A.S.; Axelsen, B.A.; Boyer, D.C.; Lynam, C.P.; Didcock, C.A.; Boyer, H.J.; Sparks, C.A.J.; Purcell, J.E.; Gibbons, M.J. Single-target echo detections of jellyfish. ICES J. Mar. Sci. 2004, 61, 383–393. [Google Scholar] [CrossRef]
- Colombo, G.A.; Mianzan, H.; Madirolas, A. Acoustic characterization of gelatinous-plankton aggregations: Four case studies from the Argentine continental shelf. ICES J. Mar. Sci. 2003, 60, 650–657. [Google Scholar] [CrossRef]
- Brosset, P.; Fromentin, J.M.; Van Beveren, E.; Lloret, J.; Marques, V.; Basilone, G.; Bonanno, A.; Carpi, P.; Donato, F.; Cikes Kec, V.; et al. Spatio-temporal patterns and environmental controls of small pelagic fish body condition from contrasted Mediterranean areas. Prog. Oceanogr. 2017, 151, 149–162. [Google Scholar] [CrossRef]
- Fanelli, E.; Da Ros, Z.; Menicucci, S.; Malavolti, S.; Biagiotti, I.; Canduci, G.; De Felice, A.; Leonori, I. The pelagic food web of the Western Adriatic Sea: A focus on the role of small pelagics. Sci. Rep. 2023, 13, 14554. [Google Scholar] [CrossRef]
- RES-GFCM/33/2009/2; Establishment of Geographical Sub-Areas in the GFCM Area Amending the Resolution GFCM/31/2007/2. GFCM, Food and Agriculture Organization of the United Nations: Rome, Italy, 2009.
- Machias, A.; Pyrounaki, M.M.; Leonori, I.; Basilone, G.; Iglesias, M.; De Felice, A.; Bonanno, A.; Giannoulaki, M. Catch of pelagic hauls in Mediterranean acoustic surveys: Is it the same between day and night? Sci. Mar. 2013, 77, 69–79. [Google Scholar] [CrossRef]
- Palermino, A.; De Felice, A.; Canduci, G.; Biagiotti, I.; Costantini, I.; Centurelli, M.; Leonori, I. Preliminary target strength measurement of Sprattus sprattus and its influence on biomass estimates in the Adriatic Sea (Mediterranean Sea). Fish. Res. 2023, 266, 106777. [Google Scholar] [CrossRef]
- De Robertis, A.; Higginbottom, I. A post-processing technique to estimate the signal-to-noise ratio and remove echosounder background noise. ICES J. Mar. Sci. 2007, 64, 1282–1291. [Google Scholar] [CrossRef]
- Ona, E. Methodology for Target Strength Measurements; ICES Cooperative Research Reports (CRR); International Council for the Exploration of the Sea (ICES): Copenhagen, Denmark, 1999; Volume 235, 65p. [Google Scholar]
- Sawada, K.; Furusawa, M.; Williamson, N.J. Conditions for the precise measurement of fish target strength in situ. J. Mar. Acous. Soc. Jpn. 1993, 20, 15–21. [Google Scholar] [CrossRef]
- Demer, D.A.; Soule, M.A.; Hewitt, R.P. A multiple-frequency method for potentially improving the accuracy and precision of in situ target strength measurements. J. Acous. Soc. Am. 1999, 105, 2359–2376. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 5 October 2023).
- Pierce, D. ncdf4: Interface to Unidata netCDF (Version 4 or Earlier) Format Data Files. R Package Version 1.21. 2023. Available online: https://CRAN.R-project.org/package=ncdf4 (accessed on 5 October 2023).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.6-23. 2023. Available online: https://CRAN.R-project.org/package=raster (accessed on 5 October 2023).
- Anderson, M.J. DISTLM Forward. Distance-Based Multivariate Analysis for a Linear Model Using forward Selection; A Computer Program; Department of Statistics, University of Auckland: Auckland, New Zealand, 2003. [Google Scholar]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Iglesias, M.; Miquel, J. Assessment of the Aphia minuta stock (Pisces: Gobiidae) by acoustic methods from the Bay of Alcudia (Mallorca, Western Mediterranean). Sci. Mar. 1998, 62, 19–25. [Google Scholar] [CrossRef]
- Kang, D.; Kim, J.; Lim, S. Acoustic target strength characteristics of two species of multiple jellyfishes, Aurelia aurita and Cyanea nozakii, in the southern coast of Korea. Ocean Polar Res. 2010, 32, 113–122. [Google Scholar] [CrossRef]
- Brierley, A.S.; Axelsen, B.E.; Buecher, E.; Sparks, C.A.J.; Boyer, H.; Gibbons, M.J. Acoustic observations of jellyfish in the Namibian Benguela. Mar. Ecol. Prog. Ser. 2001, 210, 55–66. [Google Scholar] [CrossRef]
- Monger, B.C.; Chinniah-Chandy, S.; Meir, E.; Billings, S.; Greene, C.H.; Wiebe, P.H. Sound scattering by the gelatinous zooplankters Aequorea victoria and Pleurobrachia bachei. Deep-Sea Res. II 1998, 45, 1255–1271. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, S.; Jin, X.; Li, C. Associations of large jellyfish distributions with temperature and salinity in the Yellow Sea and East China Sea. Hydrobiologia 2012, 690, 81–96. [Google Scholar] [CrossRef]
- Suchman, C.L.; Brodeur, R.D.; Daly, E.A.; Emmett, R.L. Large medusae in surface waters of the northern California Current: Variability in relation to environmental conditions. Hydrobiologia 2012, 690, 113–125. [Google Scholar] [CrossRef]
- Heim-Ballew, H.; Olsen, Z. Salinity and temperature influence on Scyphozoan jellyfish abundance in the Western Gulf of Mexico. Hydrobiologia 2019, 827, 247–262. [Google Scholar] [CrossRef]
- Yu, S.; Song, D.; Fan, M.; Xie, C. Effects of temperature and salinity on growth of Aurelia aurita. Ecol. Model. 2023, 476, 110229. [Google Scholar] [CrossRef]
- Schäfer, S.; Gueroun, S.K.M.; Andrade, C.; Canning-Clode, J. Combined Effects of Temperature and Salinity on Polyps and Ephyrae of Aurelia solida (Cnidaria: Scyphozoa). Diversity 2021, 13, 573. [Google Scholar] [CrossRef]
- Purcell, J.E. Successes and challenges in jellyfish ecology: Examples from Aequorea spp. Mar. Ecol. Prog. Ser. 2018, 591, 7–27. [Google Scholar] [CrossRef]
- Leoni, V.; Molinero, J.C.; Mefre, M.; Bonnet, D. Variability of growth rates and thermohaline niches of Rhizostoma pulmo’s pelagic stages (Cnidaria: Scyphozoa). Mar. Biol. 2021, 168, 107. [Google Scholar] [CrossRef]
- Purcell, J.E.; Atienza, D.; Fuentes, V.; Olariaga, A.; Tilves, U.; Colahan, C.; Gili, J.M. Temperature effects on asexual reproduction rates of scyphozoan species from the northwest Mediterranean Sea. Hydrobiologia 2012, 690, 169–180. [Google Scholar] [CrossRef]
- Doyle, T.K.; Houghton, J.D.R.; Buckley, S.M.; Hays, G.C.; Davenport, J. The broad-scale distribution of five jellyfish species across a temperate coastal environment. Hydrobiologia 2007, 579, 29–39. [Google Scholar] [CrossRef]
- Lee, S.H.; Tseng, L.C.; Yoon, Y.H.; Ramirez-Romero, E.; Hwang, J.S.; Molinero, J.C. The global spread of jellyfish hazards mirrors the pace of human imprint in the marine environment. Environ. Int. 2023, 171, 107699. [Google Scholar] [CrossRef]
- Lynam, C.P.; Hay, S.J.; Brierley, A.S. Jellyfish abundance and climatic variation: Contrasting responses in oceanographically distinct regions of the North Sea, and possible implications for fisheries. J. Mar. Biol. Assoc. U. K. 2005, 85, 435–450. [Google Scholar] [CrossRef]
- Purcell, J.E. Climate effects on formation of jellyfish and ctenophore blooms: A review. J. Mar. Biol. Assoc. U. K. 2005, 85, 461–476. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TS threshold | −85 dB |
| Pulse length determination level | 6 dB |
| Minimum normalized pulse length | 0.7 |
| Maximum normalized pulse length | 1.5 |
| Two-way maximum beam compensation | 4 dB |
| Maximum standard deviation minor-axis angle | 0.6° |
| Maximum standard deviation major-axis angle | 0.6° |
| Species | Total Catch (g) | Total Catch % |
|---|---|---|
| Rhizostoma pulmo (jellyfish) | 9500 | 56.63% |
| Aequorea aequorea (jellyfish) | 6900 | 41.13% |
| Chrysaora hysoscella (jellyfish) | 348 | 2.07% |
| Merlangius merlangus (fish) | 22 | 0.13% |
| Loligo vulgaris (squid) | 7 | 0.04% |
| Total | 16,776 | 100.00% |
| Species | Total Catch (g) | Total Catch % |
|---|---|---|
| Alloteuthis media (squid) | 6 | 0.01% |
| Trachurus mediterraneus (fish) | 38 | 0.05% |
| Merlangius merlangus (fish) | 34 | 0.04% |
| Engraulis encrasicolus (fish) | 1982 | 2.42% |
| Salpa maxima (salp) | 616 | 0.75% |
| Aequorea aequorea (jellyfish) | 13,640 | 16.65% |
| Sprattus sprattus (fish) | 87 | 0.11% |
| Aurelia aurita (jellyfish) | 65,031 | 79.39% |
| Chrysaora hysoscella (jellyfish) | 480 | 0.59% |
| Total | 81,914 | 100.00% |
| Species | Total Catch (g) | Total Catch % |
|---|---|---|
| Alloteuthis media (squid) | 48 | 0.30% |
| Aphia minuta (fish) | 1145 | 7.22% |
| Merlangius merlangus (fish) | 14 | 0.09% |
| Engraulis encrasicolus (fish) | 641 | 4.04% |
| Aequorea aequorea (jellyfish) | 13,300 | 83.86% |
| Sprattus sprattus (fish) | 159 | 1.00% |
| Aurelia aurita (jellyfish) | 537 | 3.38% |
| Sepiola sp (cuttlefish) | 13 | 0.08% |
| Lesueurigobius friesii (fish) | 2 | 0.01% |
| Total | 15,859 | 100.00% |
| Conditional (Sequential) Tests | |||||
|---|---|---|---|---|---|
| Variable | SS (Trace) | Pseudo-F | p | Prop | Cumulative |
| Smean (psu) | 10.7062 | 9.5938 | 0.0180 | 0.5782 | 0.5782 |
| Tmean (°C) | 1.5752 | 1.5155 | 0.2788 | 0.0851 | 0.6632 |
| Chlmean (mg/m3) | 0.6626 | 0.5944 | 0.4618 | 0.0358 | 0.6990 |
| BottomT (°C) | 0.6667 | 0.5434 | 0.4966 | 0.0360 | 0.7350 |
| Conditional (Sequential) Tests | |||||
|---|---|---|---|---|---|
| Variable | SS (Trace) | Pseudo-F | p | Prop | Cumulative |
| BottomT (°C) | 11.6175 | 6.1320 | 0.0398 | 0.4670 | 0.4670 |
| Chlmean (mg/m3) | 2.1992 | 1.1927 | 0.3696 | 0.0884 | 0.5553 |
| Tmean (°C) | 0.4699 | 0.2218 | 0.5804 | 0.0189 | 0.5742 |
| Smean (psu) | 0.0616 | 0.0234 | 0.8978 | 0.0025 | 0.5767 |
| Conditional (Sequential) Tests | |||||
|---|---|---|---|---|---|
| Variable | SS (Trace) | Pseudo-F | p | Prop | Cumulative |
| Tsup (°C) | 3.6761 | 5.1232 | 0.0564 | 0.4226 | 0.4226 |
| Smean (psu) | 1.9794 | 3.9025 | 0.1036 | 0.2276 | 0.6501 |
| Oxmean (mmol/m3) | 0.2726 | 0.4920 | 0.5236 | 0.0313 | 0.6815 |
| Chlmean (mg/m3) | 0.2884 | 0.4648 | 0.5408 | 0.0332 | 0.7146 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, A.; Biagiotti, I.; Canduci, G.; Costantini, I.; Palermino, A.; Centurelli, M.; Menicucci, S.; Leonori, I. Biodiversity of Gelatinous Organisms in the Western Adriatic Sea and Identification of Their Echo Traces in Acoustic Data. Diversity 2024, 16, 202. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040202
De Felice A, Biagiotti I, Canduci G, Costantini I, Palermino A, Centurelli M, Menicucci S, Leonori I. Biodiversity of Gelatinous Organisms in the Western Adriatic Sea and Identification of Their Echo Traces in Acoustic Data. Diversity. 2024; 16(4):202. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040202
Chicago/Turabian StyleDe Felice, Andrea, Ilaria Biagiotti, Giovanni Canduci, Ilaria Costantini, Antonio Palermino, Michele Centurelli, Samuele Menicucci, and Iole Leonori. 2024. "Biodiversity of Gelatinous Organisms in the Western Adriatic Sea and Identification of Their Echo Traces in Acoustic Data" Diversity 16, no. 4: 202. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040202