

Failed Repurposing of Lysosomotropic Drugs for COVID-19 Treatment or Prevention


Axe Microbiologie-Infectiologie et Immunologie, Research Center, CHU de Québec-Université Laval, Québec, QC G1V 4G2, Canada
Drugs Drug Candidates 2022, 1(1), 22-28; https://0-doi-org.brum.beds.ac.uk/10.3390/ddc1010003
Submission received: 27 October 2022
/
Revised: 15 November 2022
/
Accepted: 22 November 2022
/
Published: 2 December 2022
(This article belongs to the Special Issue Fighting SARS-CoV-2 and Related Viruses)
{kind=link}
Abstract
:The hope for the rapid discovery of an effective drug therapy for COVID-19 has led to several efforts to repurpose drugs approved for other indications. Lysosomotropic drugs, organic amines such as chloroquine, hydroxychloroquine, amiodarone and many others, were found to interfere with the viral life cycle in vitro but have failed in clinical trials. The properties of lysosomotropic drugs and the vacuolar cytopathology induced by them are briefly reviewed, including the critical role of lipophilicity, the central role of vacuolar (V)-ATPase for their concentration in acidic organelles, the altered function of these organelles including impaired endocytosis and secretion, macroautophagic accumulation and secondary phospholipidosis. The apparent preferential uptake of lysosomotropic drugs by phagocytic leukocytes (macrophages, neutrophils) and the high concentrations needed for a sustained disruption of vacuolar trafficking may have contributed to the failure of lysosomotropic drug repurposing for COVID-19.
1. Introduction
The coronarivus disease of 2019 (COVID-19), caused by the virus SARS-CoV-2, has had and continues to have a huge global impact on public health, with important morbidity and mortality. The hope for the rapid discovery of an effective drug therapy has led to several efforts to repurpose drugs approved for other indications to treat the viral pneumonia [1]. For instance, as of May 2021, 276 clinical trials of the anti-malarial drug hydroxychloroquine had been initiated [2]. Along this line, many drugs that have been identified in vitro as inhibitors of SARS-CoV-2 replication are false positive that fall into a category of agents that induce phospholipidosis in cultured cells [3]. These drugs are typically secondary or tertiary amines and significantly lipophilic; they transcend therapeutic caterogies and include antimalarials (chloroquine, hydroxychloroquine–Figure 1), antiarrhythmics (procainamide, amiodarone–Figure 1), psychotropic agents (haloperidol, etc.), estrogen receptor modulators (tamoxifen), antihistamines (clemastine) and many other classes and show antiviral action in cultured cells [3,4,5,6]. Tummino et al. [3] established that the threshold concentration for inducing phospholipidosis in cultured cells is correlated to the antiviral activity. Several drugs from the list, including chloroquine, hydroxycholoquine and amiodarone, have been investigated in clinical trials for COVID-19 treatment or prevention, but without success [2,3,7,8,9,10,11].
The phospholipidosis induced by cationic amphiphilic drugs is in fact a protracted consequence of their sequestration in acidic cell organelles. A lysosomotropic drug, defined as a weak base that accumulates in acidic cell compartment, is synonymous to the definition of cationic amphiphilic drug used by Tummino et al. [3]. In fact, there is evidence that these drugs are concentrated in all acidic cell compartments that express the proton pump vacuolar (V)-ATPase, including endosomes [12]. Thus, the hope was that such drugs could interfere with the endocytosis of the SARS-causing coronaviruses, a necessary step for their multiplication and release [1,13]. The author, a basic science investigator of the cellular effects of lysosomotropic drugs, offers here a personal perspective that may contribute to clarify the debate.
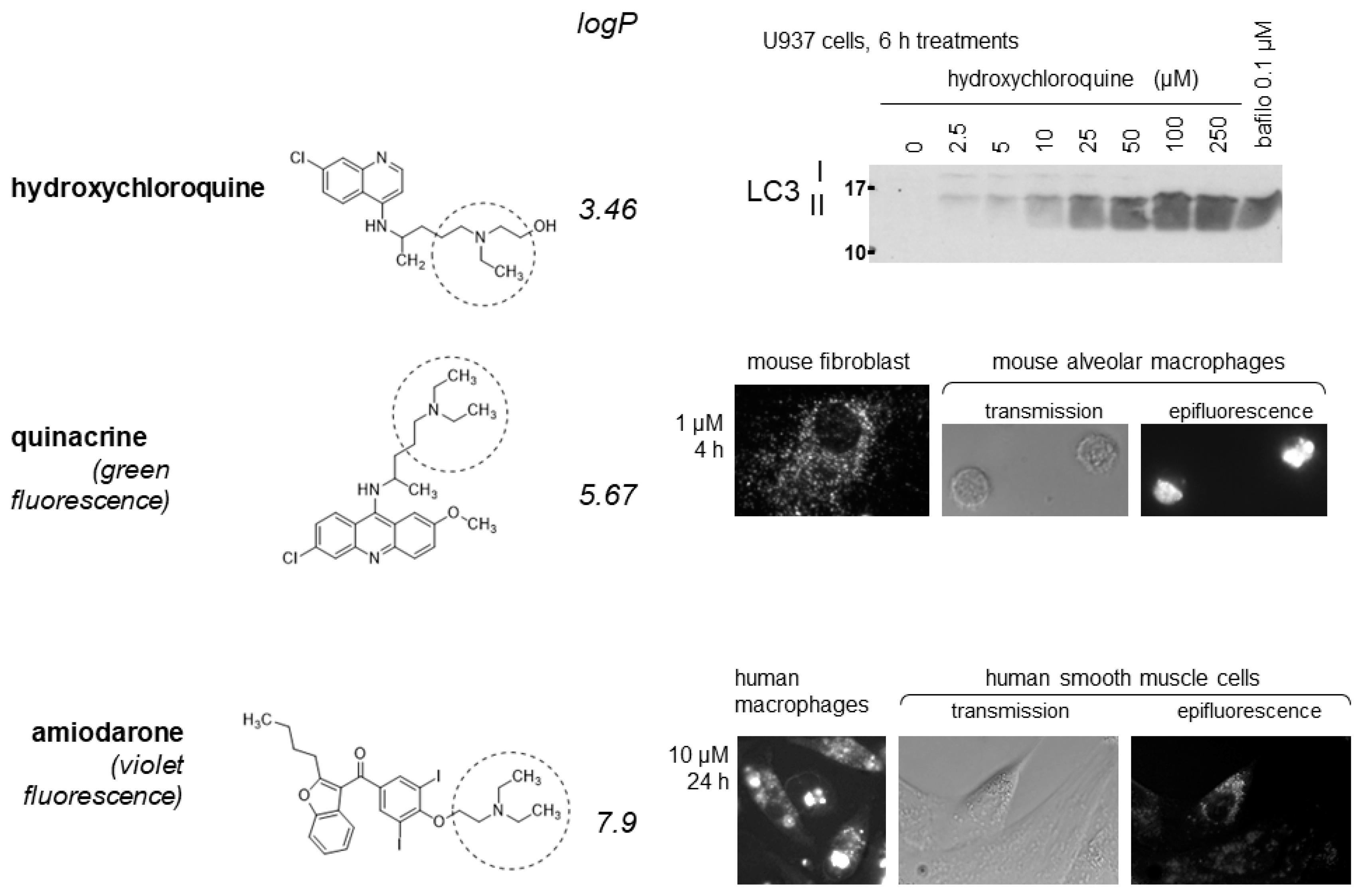
Figure 1.
Selected effects of some substituted triethylamines (the Et3N moiety is indicated by dotted circles). (Top) hydroxychloroquine, like several other substituted Et3Ns, blocks the autophagic clearance of LC3 II in U937 cells; the V-ATPase inhibitor bafilomyin A1 has a similar effect by inhibiting the acidification of intracellular organelles. (Middle) The more lipophilic drug quinacrine accumulates in perinuclear granules in murine cells, more so in freshly isolated pulmonary alveolar macrophages than in cultured fibroblasts. (Bottom) similarly, amiodarone granular uptake is more intense in human macrophages derived from peripheral blood mononuclear cells than in smooth muscle cells. The granular uptake of both quinacrine and amiodarone is completely prevented by cell pretreatment with bafilomycin A1 (not shown). Reproduced from [14,15], with permission, and [16].
Figure 1.
Selected effects of some substituted triethylamines (the Et3N moiety is indicated by dotted circles). (Top) hydroxychloroquine, like several other substituted Et3Ns, blocks the autophagic clearance of LC3 II in U937 cells; the V-ATPase inhibitor bafilomyin A1 has a similar effect by inhibiting the acidification of intracellular organelles. (Middle) The more lipophilic drug quinacrine accumulates in perinuclear granules in murine cells, more so in freshly isolated pulmonary alveolar macrophages than in cultured fibroblasts. (Bottom) similarly, amiodarone granular uptake is more intense in human macrophages derived from peripheral blood mononuclear cells than in smooth muscle cells. The granular uptake of both quinacrine and amiodarone is completely prevented by cell pretreatment with bafilomycin A1 (not shown). Reproduced from [14,15], with permission, and [16].

2. Discussion
2.1. Lysosomotropic Drugs
Lysosomotropic drugs are organic amines in equilibrium between a protonated charged form and a diffusible, uncharged form; their concentration in acidic cell compartments via ion trapping (lack of retrodiffusion of the charged form at low pH) was described several decades ago [17]. More recent work has shown that the intracellular proton pump V-ATPase provides the energy for drug concentration and retention [18]; a constellation of cellular effects of lysosomotropic drugs has also emerged: a general interference with vacuolar trafficking (secretion, endocytosis), the inhibition of basal macroautophagic clearance, a vacuolar cytopathology and the slow development of phospholipidosis [12].
The amines that are the best lysosomotropic agents possess a secondary or tertiary amine function that is not resonant with an unsaturated bond system and, critically, possess a high liposolubility because these molecules must diffuse through 2 membranes to reach the interior of acidic vacuoles from the extracellular fluid. These chemicals are indeed distributed in many apparently unrelated drug classes, don’t follow strict structure-activity rules but have in common a large apparent volume of distribution, consistent with rapid tissue uptake from the blood. Thus, chloroquine and hydroxychloroquine can be considered as substituted triethylamines (Et3N) with a fair liposolubility (logP values of 4.35 and 3.46, respectively), but there is a full scale of lipid solubility in this series. For instance, lidocaine is a more hydrosoluble Et3N (logP 2.63) and will cause a transient vacuolar cytopathology at millimolar concentrations only at sites of injection when used as a local anesthetic [19,20]. The antiarrhythmic agent amiodarone is much more lipophilic (logP 7.9), active at low micromolar levels in cultured cells, and has attracted attention in the past due to its accumulation in tissues and occasional production of related side effects such as skin discoloration and a drug accumulation disease in the lung, kidney, and other organs [14,21,22,23,24]. There is ample evidence that the lysosomotropic constellation of effects is observed following oral chronic dosing with amiodarone.
2.2. The Transport and Cellular Retention of Lysosomotropic Drugs Is Driven by V-ATPase
Vacuolar (V)-ATPase is a membrane-bound molecular rotary machine analogous to ATP synthase, but working in reverse mode: it maintains proton transport into specific organelles by consuming ATP [25]. V-ATPase is expressed in trans-Golgi-derived organelles: endosomes, secretory granules, late endosomes, lysosomes, that possess a more or less acidic content. This acidity is important for the clearance of lysosomal contents, but also for more subtle functions such as the dissociation of ligands from receptors that are later recycled at the cell surface from endosomes. Gene knock-out of critical V-ATPase subunits is embryologically lethal in mice and drosophila [26,27]. A powerful tool to analyze the function of V-ATPase is its specific and potent inhibitor bafilomycin A1 [18], but it is tolerated by cells for a few hours only.
2.3. Lysosomotropic Drug Accumulation Precedes the Cytopathology
Various mammalian cell types, either cultured or freshly isolated, concentrate lysosomotropic drugs in enlarged hollow vacuoles that are well visible in light or electron microscopy within 4 h of treatment [12,28]. When the drug is fluorescent, like quinacrine (strong green fluorescence) or amiodarone (weaker violet fluorescence), the vacuole contents is strongly fluorescent, showing that it is a concentrated solution of the amine (Figure 1). This hints at an osmotic mechanism for vacuole enlargement. Quantitative cell uptake and release experiments have been conducted with 2 substituted Et3Ns, procainamide (logP 1.13) and the much more lipophilic quinacrine (logP 5.67). Drug uptakes with apparent hyperbolic, Michaelis-Menten kinetics KM values of 4.69 mM and 9–10 µM were observed, respectively, in cultured fibroblasts or smooth muscle cells [16,28,29]. Relatively to their volume, cells concentrate quinacrine ∼5000-fold relative to the drug concentration in the culture medium [16]. Bafilomycin A1 pretreatment completely inhibit the uptake of procainamide, quinacrine or amiodarone; further, V-ATPase function is necessary for drug retention, as shown by the quick release of the lysosomotropic drugs in response to the addition of bafilomycin A1 to the medium [16,28]. Cell washout with fresh medium also rapidly releases the cationic amphiphilic drugs, with a parallel regression of the large vacuoles. While absence of evidence is not evidence of absence, plasma cell transporters have not been shown to influence the uptake or retention of quinacrine when investigating organic cation transporters or the extrusion pump P-glycoprotein [16]. Thus, the uptake of lysosomotropic drugs may be determined by their capacity to traverse membranes, consistent with the major effect of lipophilicity in their apparent affinity. The membranes of large vacuoles formed in response to lysosomotropic agents are strongly labeled with markers of late endosomes/lysosomes (Rab7, LAMP1, CD63, LC3 labeled with fluorescent proteins), occasionally with markers of early endosomes (Rab5, GTP-locked Rab5), but not with mitochondrial of endoplasmic reticulum markers [16,19,28].
2.4. The Cytopathology Induced by Lysosomotropic Drugs Is Complex and Evolving over Time
Early (2–4 h) following the treatment of a sufficient concentration of a lysosomotropic drug, there is evidence of the inhibition of macroautophagic process [28,30]. Thus, cells treated with procainamide, hydroxychloroquine, quinacrine or several other lysosomotropic drugs accumulate LC3-II (immunoblots, Figure 1) and p62/SQSTM1 [16,28], a sign of blocked clearance of autophagy in acidic vacuoles presumably buffered by the amines. Bafilomycin A1 treatment itself, by inhibiting vacuolar acidification, has the same effect consisting of LC3 II accumulation (Figure 1). However, spautin-1, an inhibitor of autophagy, did not alter quinacrine transport in cultured cells [16], suggesting that interference with autophagy is a secondary process. Interestingly, cells loaded with quinacrine undergo a protracted lysosomogenesis in an apparent ineffective feedback mechanism, as evidenced by increased cellular content of lysosomal proteins LAMP1 and LAMP2 and by the nuclear translocation of TFEB, a transcription factor that controls the synthesis of many lysosomal proteins [16].
Lysosomotropic drug-induced phospholipidosis is the reversible accumulation of polar phospholipids in intracellular bodies that are often multilamellar (‘onionoid’) and has been observed to occur in vivo following drug dosing [31]. In cultured cells, light microscopy shows that the enlarged vesicle induced by lysosomotropic drug treatment are initially filled with clear liquid (e.g., up to 4 h) [12], but later become filled with lipids (as shown by staining with Nile red, 24–72 h) [28]. In such organelles, there is a qualitative change of lipid composition as a function of time due to the inhibition of several lipid hydrolases functioning at acid pH [4,32]. Electron microscope observation showed the presence of amorphous debris in large vacuoles of cells treated with millimolar procainamide for 24 h with a hint of concentric membrane structures [12]. The membrane-associated autophagic effector LC3 is initially visualized at the surface of the enlarged vacuoles, but the latter become filled with it as a function of time in cells treated with amiodarone [14]. Hypothetically, this may result from multiple cycles of macroautophagic envelopment determined by expanding organelle damage [12] and may be the source of the concentric morphology.
Less well mechanistically defined aspect of the cytopathology induced by lysosomotropic drugs is the non-specific inhibition of endocytosis as shown by the reduced uptake of bovine serum albumin and transferrin in vacuolar cells [12,28]. Secretion is inhibited as well in a non-specific manner [12]. Some lysosomotropic drugs alter lipid metabolism in a complex manner [4,33]. Thus, cholesterol ablation reduced the antiproliferative effect of 6 Et3N drugs on U937 cells; these agents produced a desynchronized mitotic arrest that evolved into apoptosis only at extreme drug concentrations [15].
2.5. Why Was the Life Cycle of SARS-CoV-2 Expected to Be Susceptible to Lysosomotropic Drugs?
The coronavirus SARS-CoV-2 possess a single-stranded RNA genome within a membrane that is associated with several types of proteins encoded by this genome [34]. The spike protein protruding from the viral surface binds to the ectopeptidase ACE2 expressed at the surface of pulmonary epithelial cells leading to the endocytosis of the viral particle [34]. Proteolytic cleavage of the S protein is needed to permit endocytosis. The serine protease TMPRSS2 participates to this step at the host cell surface, as well as cathepsins B and L within the endosome This is one step of the viral cycle that is potentially sensitive to lysosomotropic drugs, as they depress endocytosis. From the endosomes, the uncoated RNA is released into the cytosol where the viral genome is replicated and multiple types of viral proteins are synthetized. Viral replication organelles are then formed from the endoplasmic reticulum, interact with the Golgi and are released by exocytosis [34]. The pathway of cellular egress for SARS-CoV-2 and other β-coronaviruses is peculiar: the releasing organelles have been identified as deacidified lysosomes [35]. Their susceptibility to lysosomotropic drugs is therefore doubtful.
2.6. Drug Accumulation Has Some Cell Type Specificity In Vitro and In Vivo
Mature red blood cells, devoid of endocytic and lysosomal vacuoles, do not concentrate the model lysosomotropic drug quinacrine [16]. Phagocytic cells may concentrate such drugs with a higher apparent affinity than other nucleated cell types, but are still dependent of a V-ATPase-dependent mechanism: in all the in vitro experiments reported below, bafilomycin A1 prevented the drug uptake. Blood-derived human monocytic cells, treated to functionally differentiate into macrophages, were shown to concentrate a low concentration of amiodarone in vacuoles more intensely and at a lower concentration than did smooth muscle cells, based on drug fluorescence (Figure 1) [14]. Similar observations were based on quinacrine uptake by murine cells: while cultured fibroblasts exhibited the vacuolar quinacrine uptake in the micromolar concentration range, freshly isolated lung alveolar macrophages did it more intensely (Figure 1) [16]. This comparison was motivated by the examination of lung tissue using fluorescence dissection microscopy: in mice treated for 48 h with a well tolerated dose of quinacrine, only intensely isolated fluorescent dots, corresponding to alveolar macrophages, were observed while the alveolar epithelium was not significantly fluorescent [16]. The lung distribution of the model lysosomotropic drug is troublesome for COVID-19 prevention or therapy, for which an epithelial site of action would be desirable for interference with the viral life cycle. Such a cell type preference for the accumulation of lysosomotropic drugs is a plausible cause for “foamy macrophages” observed in the lungs of patients chronically dosed with amiodarone [24]. Further, a study of freshly isolated human peripheral blood leukocytes has shown that polymorphonuclear leukocytes (PMLs; essentially neutrophils) capture most of the quinacrine added at micromolar concentrations as opposed to lymphocytes and monocytes (cytofluorometry) [30]. This study also showed a higher uptake affinity for quinacrine in PMLs than in lymphocytes (KM 1.1 vs. 6.3 µM, respectively), possibly related to a higher pinocytosis capability in the former phagocytic cell type.
3. Conclusions: Are the Effects of Lysosomotropic Drugs Clinically Exploitable?
Is the threshold concentration inducing the vacuolar cytopathology described above reached with clinical dosage of lysosomotropic drugs? Probably for a local and transient response to millimolar solutions of local anesthetics, but generally the threshold for triggering the vacuolar enlargement is above the therapeutic concentration for various other drugs. It is of possible interest that drugs that have been used as antimalarials (chloroquine, hydroxychloroquine, quinacrine) are lysosomotropic as this may indicate a special vulnerability of the parasite [36]. The anti-arrhythmic drug amiodarone is at the high end of lipophilicity in the Et3N series (Figure 1) and vacuolar sequestration is a source of side effects in some chronically dosed patients: notably skin discoloration involving mainly macrophages filled with autophagic LC3 immunoreativity, lung disease associated with “foamy” macrophages [14,23]. The tropism of lysosmotropic drugs for phagocytic leukocytes is not particularly relevant for the physiopathology of COVID-19, but may be so for the use of hydroxychloroquine as immunomodulator agent [37].
Funding
This research received no external funding. Work cited from our previous publications has been supported by the Canadian Institutes of Health Research (operating grant MOP-74448 to F.M., Canada Graduate Scholarships Doctoral Award to Guillaume Morissette) and the Natural Sciences and Engineering Research Council of Canada (operating grant to F.M.).
Institutional Review Board Statement
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Kupferschmidt, K.; Cohen, J. Race to find COVID-19 treatments accelerates. Science 2020, 367, 1412–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanden Eynde, J.J. COVID-19: Failure of the DisCoVeRy Clinical Trial, and Now-New Hopes? Pharmaceuticals 2021, 14, 664. [Google Scholar] [CrossRef] [PubMed]
- Tummino, T.A.; Rezelj, V.V.; Fischer, B.; Fischer, A.; O’Meara, M.J.; Monel, B.; Vallet, T.; White, K.M.; Zhang, Z.; Alon, A.; et al. Drug-induced phospholipidosis confounds drug repurposing for SARS-CoV-2. Science 2021, 373, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Salata, C.; Calistri, A.; Parolin, C.; Baritussio, A.; Palù, G. Antiviral activity of cationic amphiphilic drugs. Expert Rev. Anti Infect. Ther. 2017, 15, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Sauvat, A.; Ciccosanti, F.; Colavita, F.; Di Rienzo, M.; Castilletti, C.; Capobianchi, M.R.; Keep, O.; Zitvogel, L.; Fimia, G.M.; Piacentini, M.; et al. On-target versus off-target effects of drugs inhibiting the replication of SARS-CoV-2. Cell Death Dis. 2020, 11, 656. [Google Scholar] [CrossRef]
- Yang, L.; Pei, R.J.; Li, H.; Ma, X.N.; Zhou, Y.; Zhu, F.H.; He, P.L.; Tang, W.; Zhang, Y.C.; Xiong, J.; et al. Identification of SARS-CoV-2 entry inhibitors among already approved drugs. Acta Pharmacol. Sin. 2021, 42, 1347–1353. [Google Scholar] [CrossRef]
- Magagnoli, J.; Narendran, S.; Pereira, F.; Cummings, T.H.; Hardin, J.W.; Sutton, S.S.; Ambati, J. Outcomes of Hydroxychloroquine Usage in United States Veterans Hospitalized with COVID-19. Medcine 2020, 18, 114–127. [Google Scholar] [CrossRef]
- Mitjà, O.; Corbacho-Monné, M.; Ubals, M.; Alemany, A.; Suñer, C.; Tebé, C.; Tobias, A.; Peñafiel, J.; Ballana, E.; Pérez, C.A.; et al. BCN-PEP-CoV2 Research Group. A Cluster-Randomized Trial of Hydroxychloroquine for Prevention of COVID-19. N. Engl. J. Med. 2021, 384, 417–427. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Agoramoorthy, G.; Lee, S.S. The Drug Repurposing for COVID-19 Clinical Trials Provide Very Effective Therapeutic Combinations: Lessons Learned From Major Clinical Studies. Front. Pharmacol. 2021, 12, 704205. [Google Scholar] [CrossRef]
- Navarese, E.P.; Podhajski, P.; Andreotti, F.; La Torre, G.; Gajda, R.; Radziwanowski, A.; Nowicka, M.; Bukowski, P.; Gajda, J.; Omyła, M.; et al. Ion channel inhibition with amiodarone or verapamil in symptomatic hospitalized nonintensive-care COVID-19 patients: The ReCOVery-SIRIO randomized trial. Cardiol. J. 2022, 29, 739–750. [Google Scholar] [CrossRef]
- Tirupakuzhi Vijayaraghavan, B.K.; Jha, V.; Rajbhandari, D.; Myatra, S.N.; Ghosh, A.; Bhattacharya, A.; Arfin, S.; Bassi, A.; Donaldson, L.H.; Hammond, N.E.; et al. Hydroxychloroquine plus personal protective equipment versus personal protective equipment alone for the prevention of laboratory-confirmed COVID-19 infections among healthcare workers: A multicentre, parallel-group randomised controlled trial from India. BMJ Open 2022, 12, 059540. [Google Scholar] [CrossRef] [PubMed]
- Marceau, F.; Bawolak, M.T.; Lodge, R.; Bouthillier, J.; Gagné-Henley, A.; Gaudreault, R.C.; Morissette, G. Cation trapping by cellular acidic compartments: Beyond the concept of lysosomotropic drugs. Toxicol. Appl. Pharmacol. 2012, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stadler, K.; Ha, H.R.; Ciminale, V.; Spirli, C.; Saletti, G.; Schiavon, M.; Bruttomesso, D.; Bigler, L.; Follath, F.; Pettenazzo, A.; et al. Amiodarone alters late endosomes and inhibits SARS coronavirus infection at a post-endosomal level. Am. J. Respir. Cell. Mol. Biol. 2008, 39, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Morissette, G.; Ammoury, A.; Rusu, D.; Marguery, M.C.; Lodge, R.; Poubelle, P.E.; Marceau, F. Intracellular sequestration of amiodarone: Role of vacuolar ATPase and macroautophagic transition of the resulting vacuolar cytopathology. Br. J. Pharmacol. 2009, 157, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Parks, A.; Marceau, F. Lysosomotropic cationic drugs induce cytostatic and cytotoxic effects: Role of liposolubility and autophagic flux and antagonism by cholesterol ablation. Toxicol. Appl. Pharmacol. 2016, 305, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, A.; Charest-Morin, X.; Boivin-Welch, M.; Bouthillier, J.; Marceau, F. Autophagic flux inhibition and lysosomogenesis ensuing cellular capture and retention of the cationic drug quinacrine in murine models. PeerJ 2015, 3, e1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Duve, C.; De Barsy, T.; Poole, B.; Trouet, A.; Tulkens, P.; Van Hoof, F. Commentary. Lysosomotropic agents. Biochem. Pharmacol. 1974, 23, 2495–2531. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, Y. Membrane energization by proton pumps is important for compartmentalization of drugs and toxins: A new type of active transport. J. Exp. Biol. 1996, 199, 1447–1454. [Google Scholar] [CrossRef]
- Bawolak, M.T.; Morissette, G.; Marceau, F. Vacuolar ATPase-mediated sequestration of local anesthetics in swollen macroautophagosomes. Can. J. Anaesth. 2010, 57, 230–239. [Google Scholar] [CrossRef]
- Atilla, H.; Tekeli, O.; Can, B.; Karel, F.; Saran, Y. Effects of intracameral lidocaine on ocular tissues. Clin. Exp. Ophthalmol. 2003, 31, 73–77. [Google Scholar] [CrossRef]
- Myers, J.L.; Kennedy, J.I.; Plumb, V.J. Amiodarone lung: Pathologic findings in clinically toxic patients. Hum. Pathol. 1987, 18, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Ammoury, A.; Michaud, S.; Paul, C.; Prost-Squarcioni, C.; Alvarez, F.; Lamant, L.; Launay, F.; Bazex, J.; Chouini-Lalanne, N.; Marguery, M.C. Photodistribution of blue-gray hyperpigmentation after amiodarone treatment: Molecular characterization of amiodarone in the skin. Arch. Dermatol. 2008, 144, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruangchira-Urai, R.; Colby, T.V.; Klein, J.; Nielsen, G.P.; Kradin, R.L.; Mark, E.J. Nodular amiodarone lung disease. Am. J. Surg. Pathol. 2008, 32, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Bedrossian, C.W.; Warren, C.J.; Ohar, J.; Bhan, R. Amiodarone pulmonary toxicity: Cytopathology, ultrastructure, and immunocytochemistry. Ann. Diagn. Pathol. 1997, 1, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Santos-Pereira, C.; Rodrigues, L.R.; Côrte-Real, M. Emerging insights on the role of V-ATPase in human diseases: Therapeutic challenges and opportunities. Med. Res. Rev. 2021, 41, 1927–1964. [Google Scholar] [CrossRef]
- Dow, J.A.T.; Davies, S.A.; Guo, Y.; Graham, S.; Finbow, M.E.; Kaiser, K. Molecular genetic analysis of V-ATPase functions in Drosophila melanogaster. J. Exp. Biol. 1997, 200, 237–245. [Google Scholar] [CrossRef]
- Inoue, H.; Noumi, T.; Nagata, M.; Murakami, H.; Kanazawa, H. Targeted disruption of the gene encoding the proteolipid subunit of mouse vacuolar H+-ATPase leads to early embryonic lethality. Biochim. Biophys. Acta 1999, 1413, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Morissette, G.; Lodge, R.; Marceau, F. Intense pseudotransport of a cationic drug mediated by vacuolar ATPase: Procainamide-induced autophagic cell vacuolization. Toxicol. Appl. Pharmacol. 2008, 228, 364–377. [Google Scholar] [CrossRef]
- Marceau, F.; Bawolak, M.T.; Bouthillier, J.; Morissette, G. Vacuolar ATPase-mediated cellular concentration and retention of quinacrine: A model for the distribution of lipophilic cationic drugs to autophagic vacuoles. Drug Metab. Dispos. 2009, 37, 2271–2274. [Google Scholar] [CrossRef] [Green Version]
- Roy, C.; Gagné, V.; Fernandes, M.J.G.; Marceau, F. High affinity capture and concentration of quinacrine in polymormonuclear neutrophils via vacuolar ATPase-mediated ion trapping: Comparison with other peripheral blood leukocytes and implications for the distribution of cationic drugs. Tox. Appl. Pharmacol. 2013, 270, 77–86. [Google Scholar] [CrossRef]
- Reasor, M.J.; Hastings, K.L.; Ulrich, R.G. Drug-induced phospholipidosis: Issues and future directions. Expert Opin. Drug Saf. 2006, 5, 567–583. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, E.; Nadai, M.; Caretta, C.M.; Bergonzini, V.; Del Vecchio, C.; Ha, H.R.; Bigler, L.; Dal Zoppo, D.; Faggin, E.; Pettenazzo, A.; et al. Amiodarone impairs trafficking through late endosomes inducing a Niemann-Pick C-like phenotype. Biochem. Pharmacol. 2011, 82, 1234–1249. [Google Scholar] [CrossRef] [PubMed]
- Kuzu, O.F.; Toprak, M.; Noory, M.A.; Robertson, G.P. Effect of lysosomotropic molecules on cellular homeostasis. Pharmacol. Res. 2017, 117, 177–184. [Google Scholar] [CrossRef] [PubMed]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Dellibovi-Ragheb, T.A.; Kerviel, A.; Pak, E.; Qiu, Q.; Fisher, M.; Takvorian, P.M.; Bleck, C.; Hsu, V.W.; Fehr, A.R.; et al. β-Coronaviruses Use Lysosomes for Egress Instead of the Biosynthetic Secretory Pathway. Cell 2020, 183, 1520–1535. [Google Scholar] [CrossRef] [PubMed]
- Warhurst, D.C.; Steele, J.C.; Adagu, I.S.; Craig, J.C.; Cullander, C. Hydroxychloroquine is much less active than chloroquine against chloroquine-resistant Plasmodium falciparum, in agreement with its physicochemical properties. J. Antimicrob. Chemother. 2003, 52, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Durcan, L.; Petri, M. Immunomodulators in SLE: Clinical evidence and immunologic actions. J. Autoimmun. 2016, 74, 73–84. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marceau, F. Failed Repurposing of Lysosomotropic Drugs for COVID-19 Treatment or Prevention. Drugs Drug Candidates 2022, 1, 22-28. https://0-doi-org.brum.beds.ac.uk/10.3390/ddc1010003
AMA Style
Marceau F. Failed Repurposing of Lysosomotropic Drugs for COVID-19 Treatment or Prevention. Drugs and Drug Candidates. 2022; 1(1):22-28. https://0-doi-org.brum.beds.ac.uk/10.3390/ddc1010003
Chicago/Turabian StyleMarceau, François. 2022. "Failed Repurposing of Lysosomotropic Drugs for COVID-19 Treatment or Prevention" Drugs and Drug Candidates 1, no. 1: 22-28. https://0-doi-org.brum.beds.ac.uk/10.3390/ddc1010003