
Urinary mRNA Expression of Glomerular Podocyte Markers in Glomerular Disease and Renal Transplant

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Controls
2.2. Patients
- RTx characteristics:
- N characteristics:
2.3. Urine
2.4. Biochemical Analysis
2.5. Human Immortalized Podocytes Culture
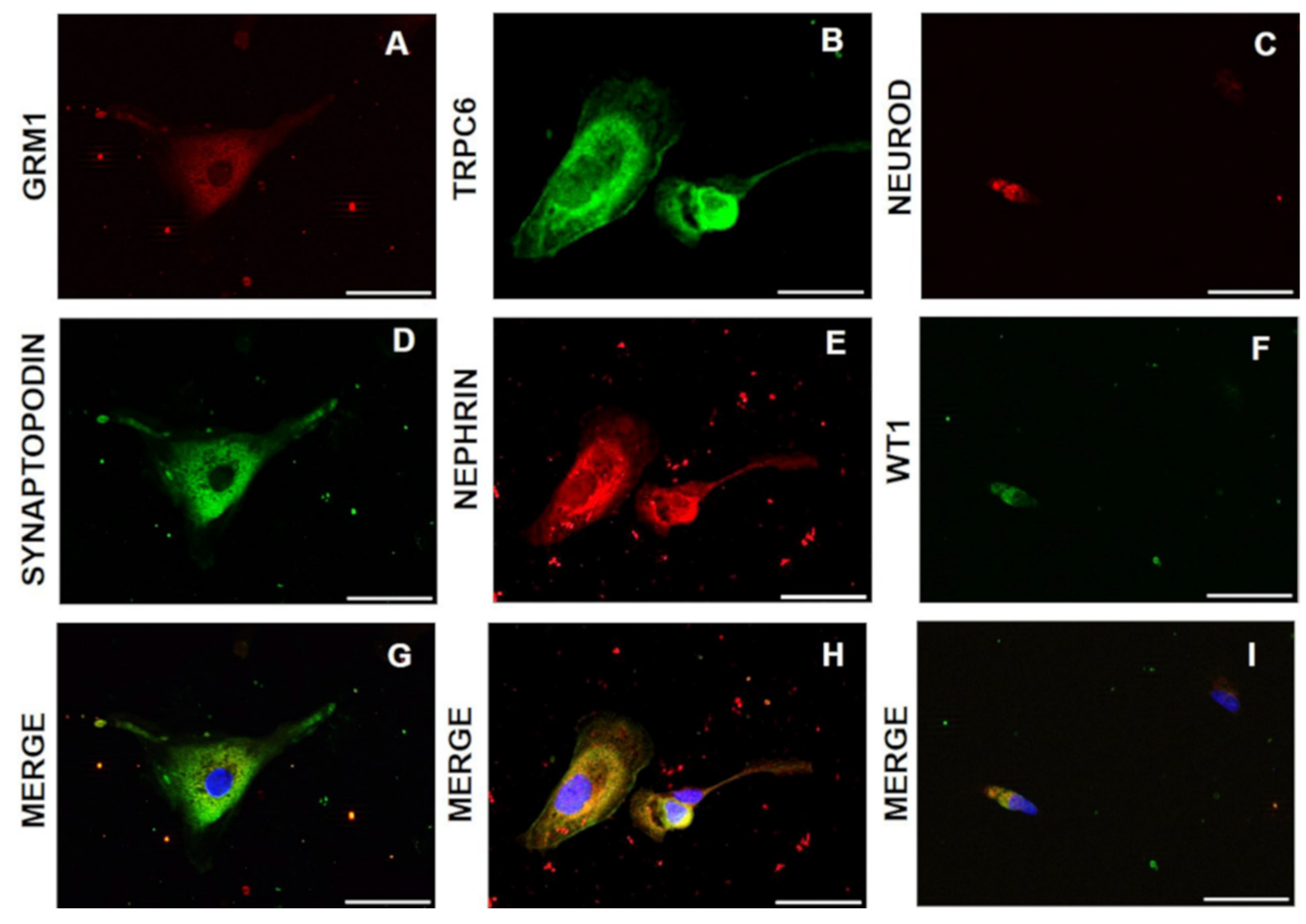
2.6. Immunofluorescence Staining
2.7. RNA Extraction
2.8. Urinary Cells Culture
2.9. RTqPCR
2.10. Statistical Analysis
3. Results
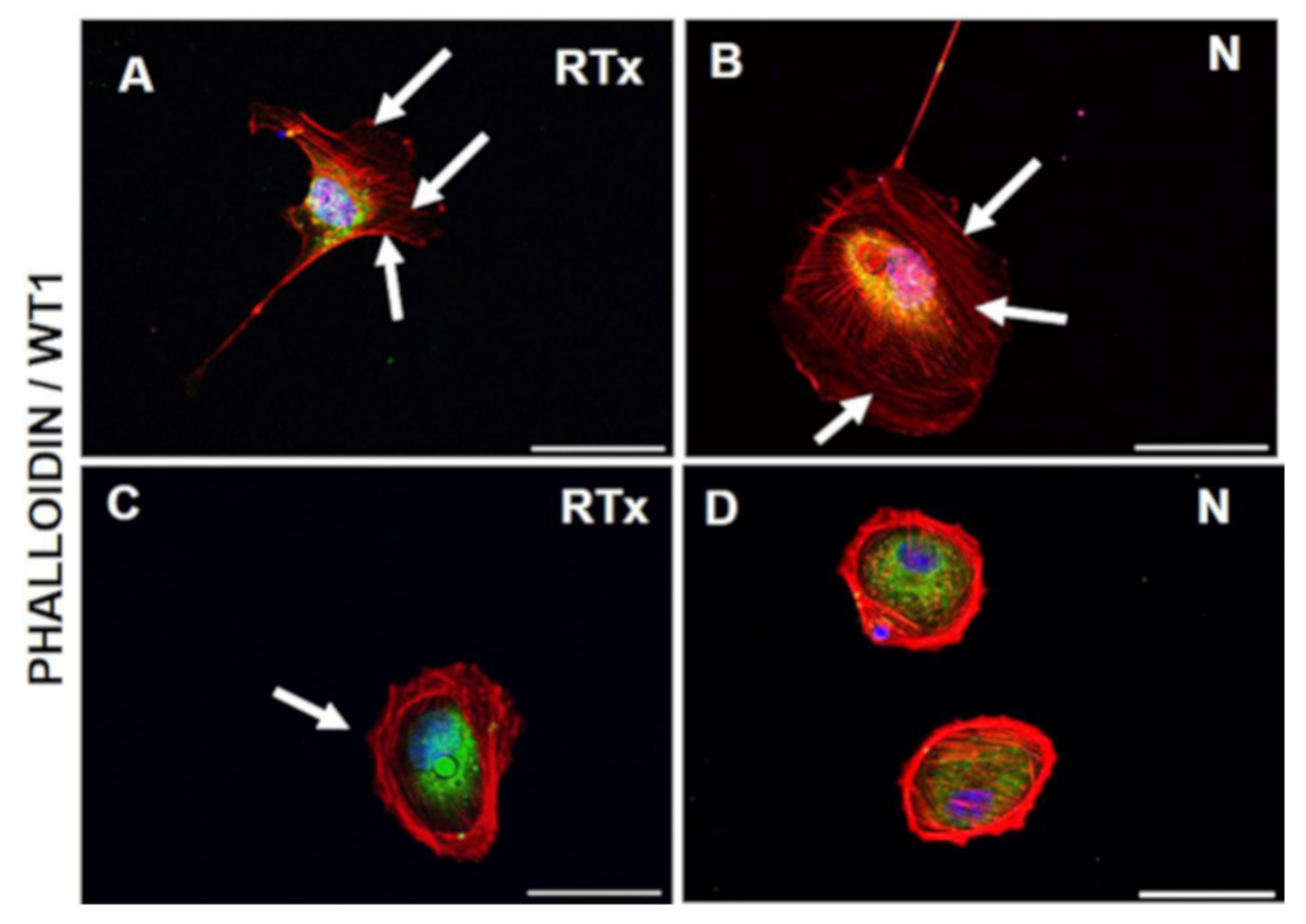
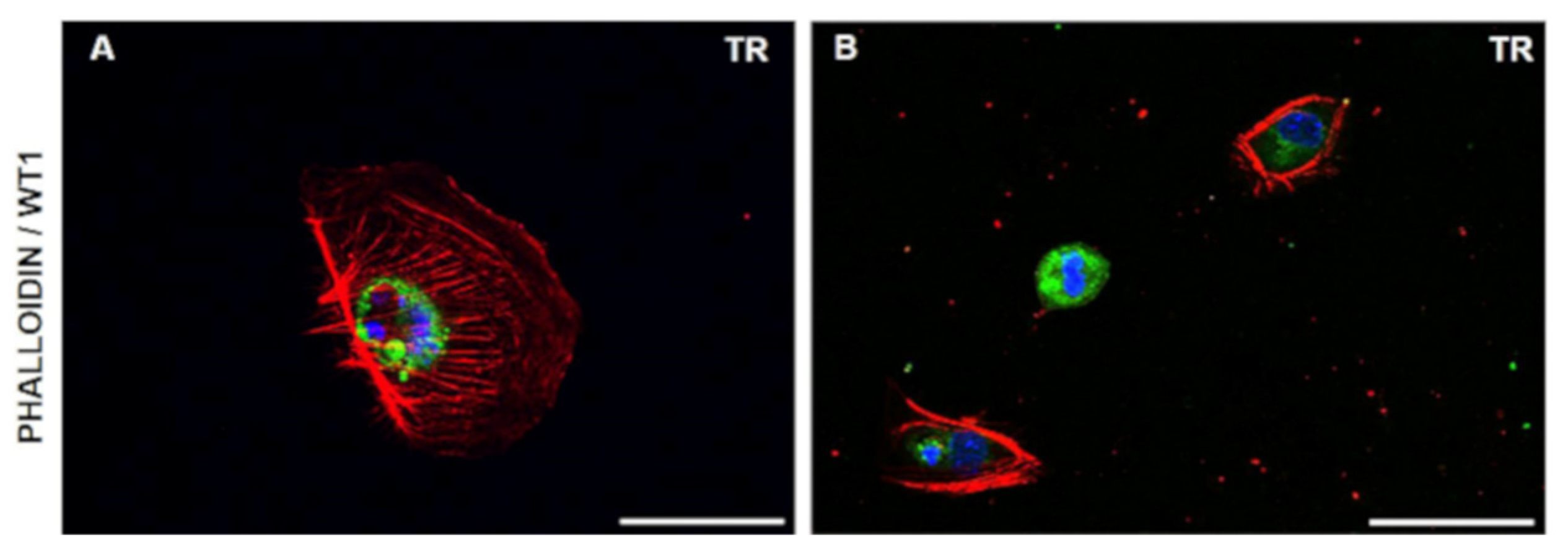
3.1. Cell Morphology and Growth
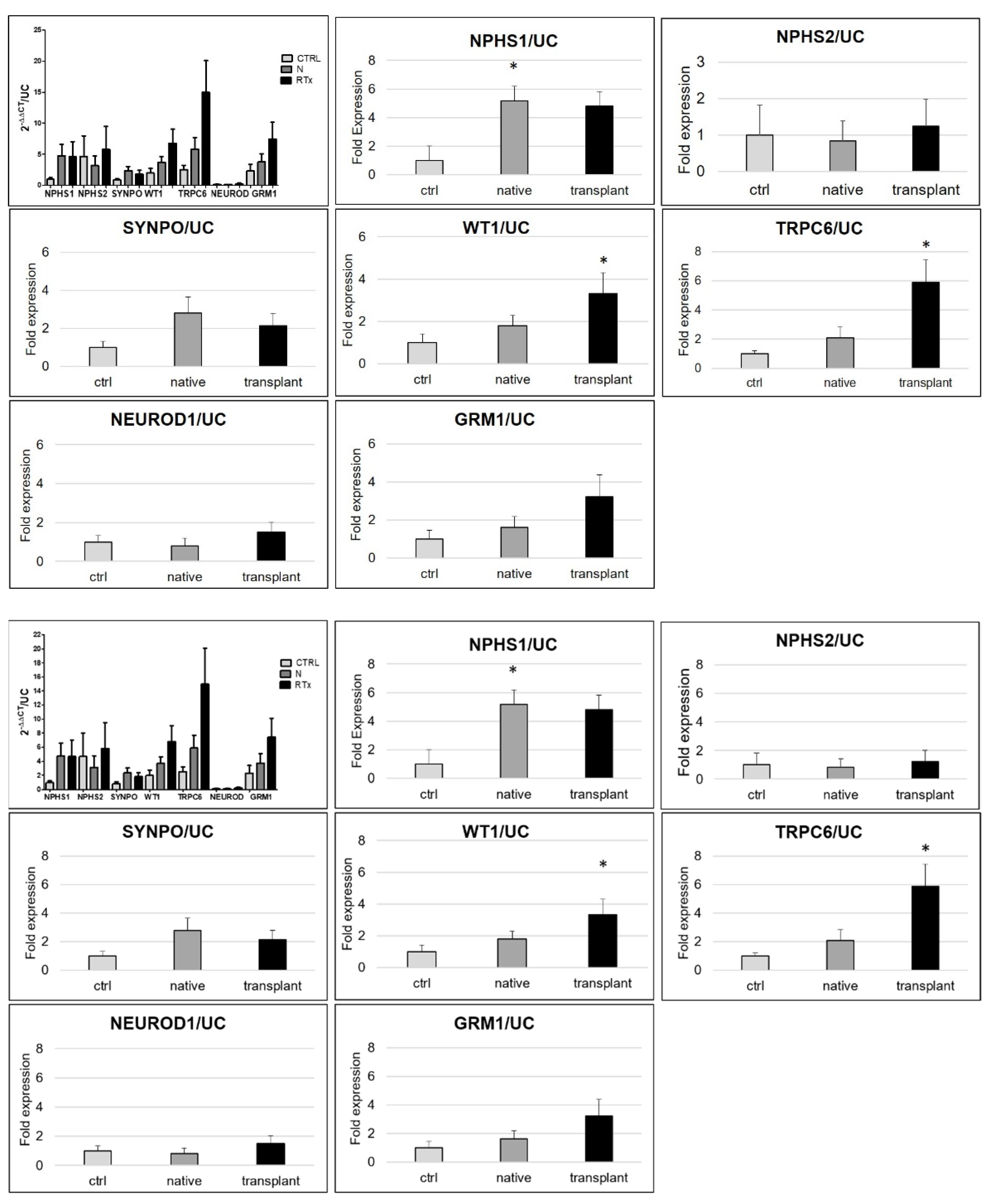
3.2. Urinary mRNA Expression
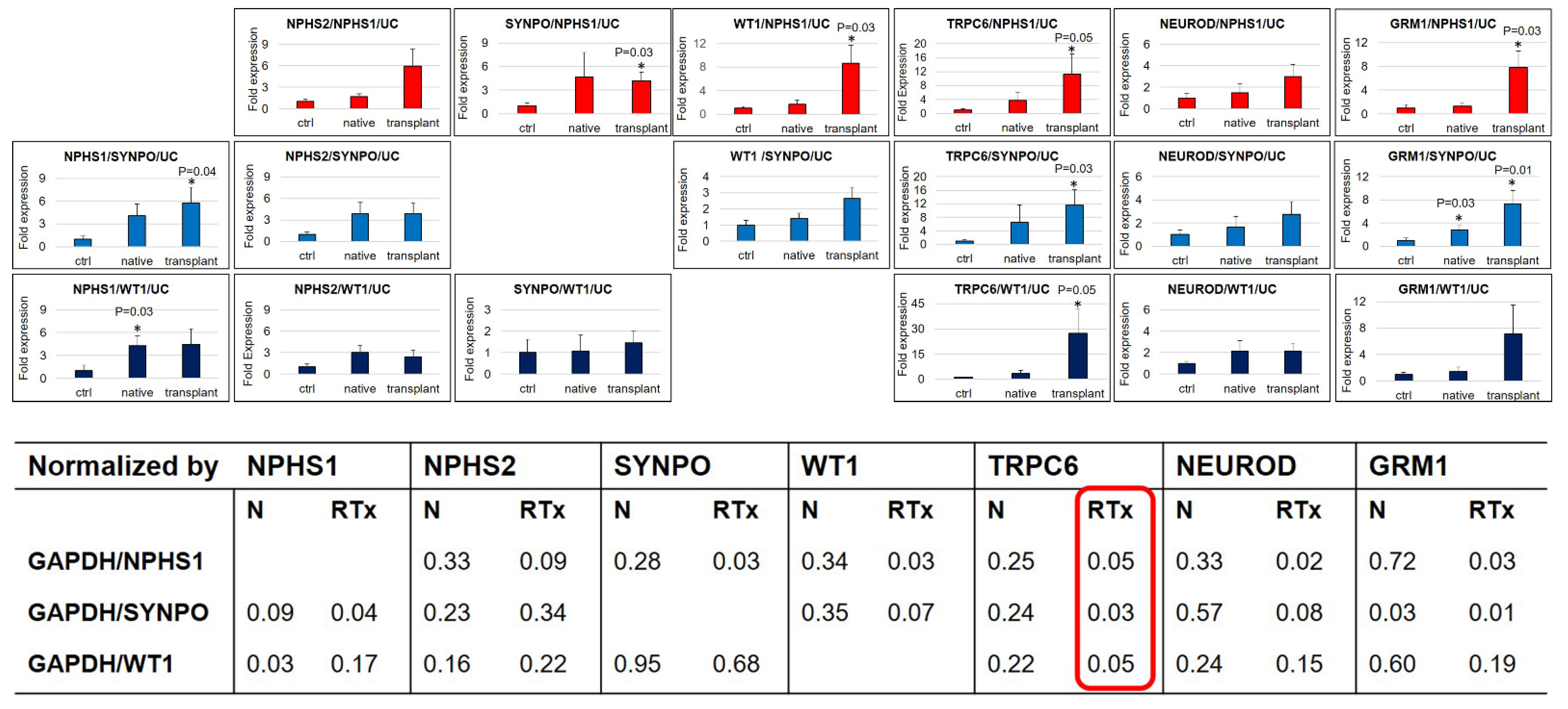
3.3. Correlation among the Different Markers after Normalization for Total Number of Cells
3.4. mRNA Expression Related to Urinary Podocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lorz, C.; Benito-Martín, A.; Boucherot, A.; Ucero, A.C.; Rastaldi, M.P.; Henger, A.; Armelloni, S.; Santamaría, B.; Berthier, C.C.; Kretzler, M.; et al. The Death Ligand TRAIL in Diabetic Nephropathy. J. Am. Soc. Nephrol. 2008, 19. [Google Scholar] [CrossRef] [Green Version]
- Armelloni, S.; Corbelli, A.; Giardino, L.; Li, M.; Ikehata, M.; Mattinzoli, D.; Messa, P.; Pignatari, C.; Watanabe, S.; Rastaldi, M.P. Podocytes: Recent biomolecular developments. Biomol. Concepts 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, C.; Kavvadas, P.; Simonini, P.; Ikehata, M.; Dussaule, J.C.; Chadjichristos, C.E.; Rastaldi, M.P.; Messa, P.; Chatziantoniou, C. Discoidin domain receptor-1 and periostin: New players in chronic kidney disease. Nephrol. Dial. Transplant. 2015, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfieri, C.; Regalia, A.; Moroni, G.; Cresseri, D.; Zanoni, F.; Ikehata, M.; Simonini, P.; Rastaldi, M.P.; Tripepi, G.; Zoccali, C.; et al. Novel markers of graft outcome in a cohort of kidney transplanted patients: A cohort observational study. J. Nephrol. 2019, 32. [Google Scholar] [CrossRef]
- Ju, W.; Nair, V.; Smith, S.; Zhu, L.; Shedden, K.; Song, P.X.K.; Mariani, L.H.; Eichinger, F.H.; Berthier, C.C.; Randolph, A.; et al. Tissue transcriptome-driven identification of epidermal growth factor as a chronic kidney disease biomarker. Sci. Transl. Med. 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Ye, Y.; Bao, W.; Yang, Q.; Wang, J.; Liu, Z.; Shi, S. Genome-wide identification of genes essential for podocyte cytoskeletons based on single-cell RNA sequencing. Kidney Int. 2017, 92. [Google Scholar] [CrossRef]
- Erpicum, P.; Hanssen, O.; Weekers, L.; Lovinfosse, P.; Meunier, P.; Tshibanda, L.; Krzesinski, J.-M.; Hustinx, R.; Jouret, F. Non-invasive approaches in the diagnosis of acute rejection in kidney transplant recipients, part II: Omics analyses of urine and blood samples. Clin. Kidney J. 2016. [Google Scholar] [CrossRef] [Green Version]
- Sigdel, T.K.; Gao, Y.; He, J.; Wang, A.; Nicora, C.D.; Fillmore, T.L.; Shi, T.; Webb-Robertson, B.-J.; Smith, R.D.; Qian, W.-J.; et al. Mining the human urine proteome for monitoring renal transplant injury. Kidney Int. 2016, 89. [Google Scholar] [CrossRef] [Green Version]
- Cassidy, H.; Slyne, J.; O’Kelly, P.; Traynor, C.; Conlon, P.J.; Johnston, O.; Slattery, C.; Ryan, M.P.; McMorrow, T. Urinary biomarkers of chronic allograft nephropathy. Proteom. Clin. Appl. 2015, 9. [Google Scholar] [CrossRef]
- Shahbaz, S.K.; Pourrezagholi, F.; Barabadi, M.; Foroughi, F.; Hosseinzadeh, M.; Ahmadpoor, P.; Nafar, M.; Yekaninejad, M.S.; Amirzargar, A. High expression of TIM-3 and KIM-1 in blood and urine of renal allograft rejection patients. Transpl. Immunol. 2017, 43–44. [Google Scholar] [CrossRef]
- Perez-Hernandez, J.; Olivares, M.D.; Forner, M.J.; Chaves, F.J.; Cortes, R.; Redon, J. Urinary dedifferentiated podocytes as a non-invasive biomarker of lupus nephritis. Nephrol. Dial. Transplant. 2016, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, M.; Bringhenti, R.N.; Rodrigues, P.G.; do Nascimento, J.F.; Pereira, S.V.; Zancan, R.; Monticielo, O.A.; Gasparin, A.A.; de Castro, W.P.; Veronese, F.V. Podocyte-associated mRNA profiles in kidney tissue and in urine of patients with active lupus nephritis. Int. J. Clin. Exp. Pathol. 2015, 8, 4600–4613. [Google Scholar]
- Mezni, I.; Galichon, P.; Mongi Bacha, M.; Xu-Dubois, Y.-C.; Sfar, I.; Buob, D.; Benbouzid, S.; Goucha, R.; Gorgi, Y.; Abderrahim, E.; et al. Urinary mRNA analysis of biomarkers to epithelial mesenchymal transition of renal allograft. Néphrologie Thérapeutique 2018, 14. [Google Scholar] [CrossRef]
- Suthanthiran, M.; Schwartz, J.E.; Ding, R.; Abecassis, M.; Dadhania, D.; Samstein, B.; Knechtle, S.J.; Friedewald, J.; Becker, Y.T.; Sharma, V.K.; et al. Urinary-Cell mRNA Profile and Acute Cellular Rejection in Kidney Allografts. N. Engl. J. Med. 2013, 369. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Hartono, C.; Ding, R.; Sharma, V.K.; Ramaswamy, R.; Qian, B.; Serur, D.; Mouradian, J.; Schwartz, J.E.; Suthanthiran, M. Noninvasive Diagnosis of Renal-Allograft Rejection by Measurement of Messenger RNA for Perforin and Granzyme B in Urine. N. Engl. J. Med. 2001, 344. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Kim, G.T.; Hyun, M.; Kim, S.; Seok, S.; Choi, R.; Lee, M.Y.; Chung, C.H. Peroxisome proliferator-activated receptor-δ activation ameliorates albuminuria by preventing nephrin loss and restoring podocyte integrity in type 2 diabetes. Nephrol. Dial. Transplant. 2012, 27, 4069–4079. [Google Scholar] [CrossRef] [Green Version]
- Naik, A.S.; Afshinnia, F.; Aqeel, J.; Cibrik, D.M.; Samaniego, M.; Wickman, L.; Wang, S.Q.; Chowdhury, M.; Wiggins, R.C. Accelerated podocyte detachment early after kidney transplantation is related to long-term allograft loss of function. Nephrol. Dial. Transplant. 2019, 34. [Google Scholar] [CrossRef]
- Fukuda, A.; Minakawa, A.; Sato, Y.; Iwakiri, T.; Iwatsubo, S.; Komatsu, H.; Kikuchi, M.; Kitamura, K.; Wiggins, R.C.; Fujimoto, S. Urinary podocyte and TGF-β1 mRNA as markers for disease activity and progression in anti-glomerular basement membrane nephritis. Nephrol. Dial. Transplant. 2017, 32. [Google Scholar] [CrossRef]
- Fukuda, A.; Sato, Y.; Iwakiri, T.; Komatsu, H.; Kikuchi, M.; Kitamura, K.; Wiggins, R.C.; Fujimoto, S. Urine podocyte mRNAs mark disease activity in IgA nephropathy. Nephrol. Dial. Transplant. 2015, 30. [Google Scholar] [CrossRef] [Green Version]
- Szeto, C.-C.; Lai, K.-B.; Chow, K.-M.; Szeto, C.Y.-K.; Yip, T.W.-C.; Woo, K.-S.; Li, P.K.-T.; Lai, F.M.-M. Messenger RNA expression of glomerular podocyte markers in the urinary sediment of acquired proteinuric diseases. Clin. Chim. Acta 2005, 361. [Google Scholar] [CrossRef]
- Sis, B.; Mengel, M.; Haas, M.; Colvin, R.B.; Halloran, P.F.; Racusen, L.C.; Solez, K.; Baldwin, W.M.; Bracamonte, E.R.; Broecker, V.; et al. Banff’09 Meeting Report: Antibody Mediated Graft Deterioration and Implementation of Banff Working Groups. Am. J. Transplant. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Solez, K.; Colvin, R.B.; Racusen, L.C.; Sis, B.; Halloran, P.F.; Birk, P.E.; Campbell, P.M.; Cascalho, M.; Collins, A.B.; Demetris, A.J.; et al. Banff’05 Meeting Report: Differential Diagnosis of Chronic Allograft Injury and Elimination of Chronic Allograft Nephropathy (‘CAN’). Am. J. Transplant. 2007, 7. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Wickman, L.; Wang, S.Q.; Zhang, Y.; Wang, F.; Afshinnia, F.; Hodgin, J.; Ding, J.; Wiggins, R.C. Accelerated podocyte detachment and progressive podocyte loss from glomeruli with age in Alport Syndrome. Kidney Int. 2017, 92. [Google Scholar] [CrossRef] [PubMed]
- Trimarchi, H. Mechanisms of Podocyte Detachment, Podocyturia, and Risk of Progression of Glomerulopathies. Kidney Dis. 2020, 6. [Google Scholar] [CrossRef]
- Galichon, P.; Amrouche, L.; Hertig, A.; Brocheriou, I.; Rabant, M.; Xu-Dubois, Y.-C.; Ouali, N.; Dahan, K.; Morin, L.; Terzi, F.; et al. Urinary mRNA for the Diagnosis of Renal Allograft Rejection: The Issue of Normalization. Am. J. Transplant. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lai, F.M.-M.; Lai, K.-B.; Chow, K.-M.; Kwan, B.C.-H.; Li, P.K.-T.; Szeto, C.-C. Urinary messenger RNA expression of podocyte-associated molecules in patients with diabetic nephropathy treated by angiotensin-converting enzyme inhibitor and angiotensin receptor blocker. Eur. J. Endocrinol. 2008, 158. [Google Scholar] [CrossRef]
- Fukuda, A.; Wickman, L.T.; Venkatareddy, M.P.; Wang, S.Q.; Chowdhury, M.A.; Wiggins, J.E.; Shedden, K.A.; Wiggins, R.C. Urine podocin:nephrin mRNA ratio (PNR) as a podocyte stress biomarker. Nephrol. Dial. Transplant. 2012, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Petermann, A.; Kunter, U.; Rong, S.; Shankland, S.J.; Floege, J. Urinary Podocyte Loss Is a More Specific Marker of Ongoing Glomerular Damage than Proteinuria. J. Am. Soc. Nephrol. 2005, 16. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Yanagihara, T.; Kihara, I. Cumulative Excretion of Urinary Podocytes Reflects Disease Progression in IgA Nephropathy and Schönlein-Henoch Purpura Nephritis. Clin. J. Am. Soc. Nephrol. 2007, 2. [Google Scholar] [CrossRef]
- Zhang, J.; Hansen, K.M.; Pippin, J.W.; Chang, A.M.; Taniguchi, Y.; Krofft, R.D.; Pickering, S.G.; Liu, Z.-H.; Abrass, C.K.; Shankland, S.J. De novo expression of podocyte proteins in parietal epithelial cells in experimental aging nephropathy. Am. J. Physiol. Physiol. 2012, 302. [Google Scholar] [CrossRef] [Green Version]
- Manonelles, A.; Guiteras, R.; Melilli, E.; Lazzeri, E.; Goma, M.; Crespo, E.; Bestard, O.; Sola, A.; Romagnani, P.; Cruzado, J.M. The Presence of Urinary Renal Progenitor Cells in Stable Kidney Transplant Recipients Anticipates Allograft Deterioration. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Nakamura, T.; Ushiyama, C.; Suzuki, S.; Hara, M.; Shimada, N.; Ebihara, I.; Koide, H. Urinary excretion of podocytes in patients with diabetic nephropathy. Nephrol. Dial. Transplant. 2000, 15. [Google Scholar] [CrossRef]
- Hara, M.; Yanagihara, T.; Takada, T.; Itoh, M.; Matsuno, M.; Yamamoto, T.; Kihara, I. Urinary excretion of podocytes reflects disease activity in children with glomerulonephritis. Am. J. Nephrol. 1998, 18, 35–41. [Google Scholar] [CrossRef]
- Kriz, W.; HÃhnel, B.; Hosser, H.; Rösener, S.; Waldherr, R. Structural Analysis of How Podocytes Detach from the Glomerular Basement Membrane Under Hypertrophic Stress. Front. Endocrinol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, S.U.; Nelson, W.J.; Myers, B.D.; Lemley, K.V. Urinary excretion of viable podocytes in health and renal disease. Am. J. Physiol. Physiol. 2003, 285. [Google Scholar] [CrossRef] [Green Version]
- Greka, A.; Mundel, P. Cell Biology and Pathology of Podocytes. Annu. Rev. Physiol. 2012, 74. [Google Scholar] [CrossRef] [Green Version]
- Tryggvason, K.; Patrakka, J.; Wartiovaara, J. Hereditary Proteinuria Syndromes and Mechanisms of Proteinuria. N. Engl. J. Med. 2006, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boute, N.; Gribouval, O.; Roselli, S.; Benessy, F.; Lee, H.; Fuchshuber, A.; Dahan, K.; Gubler, M.-C.; Niaudet, P.; Antignac, C. NPHS2, encoding the glomerular protein podocin, is mutated in autosomal recessive steroid-resistant nephrotic syndrome. Nat. Genet. 2000, 24. [Google Scholar] [CrossRef] [PubMed]
- Barisoni, L.; Schnaper, H.W.; Kopp, J.B. A Proposed Taxonomy for the Podocytopathies: A Reassessment of the Primary Nephrotic Diseases. Clin. J. Am. Soc. Nephrol. 2007, 2. [Google Scholar] [CrossRef] [PubMed]
- Schmid, H.; Henger, A.; Cohen, C.D. Gene expression profiles of podocyte-associated molecules as diagnostic markers in acquired proteinuric diseases. J. Am. Soc. Nephrol. 2003, 14, 2958–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pippin, J.; Kumar, V.; Stein, A.; Jablonski, P.; Shankland, S.J.; Davis, C.L. The Contribution of Podocytes to Chronic Allograft Nephropathy. Nephron Exp. Nephrol. 2009, 111. [Google Scholar] [CrossRef]
- Endlich, K.; Kliewe, F.; Endlich, N. Stressed podocytes—mechanical forces, sensors, signaling and response. Pflügers Arch. Eur. J. Physiol. 2017, 469. [Google Scholar] [CrossRef]
- Petrica, L.; Ursoniu, S.; Gadalean, F.; Vlad, A.; Gluhovschi, G.; Dumitrascu, V.; Vlad, D.; Gluhovschi, C.; Velciov, S.; Bob, F.; et al. Urinary podocyte-associated mRNA levels correlate with proximal tubule dysfunction in early diabetic nephropathy of type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Lai, F.M.-M.; Lai, K.-B.; Chow, K.-M.; Li, K.-T.P.; Szeto, C.-C. Messenger RNA Expression of Podocyte-Associated Molecules in the Urinary Sediment of Patients with Diabetic Nephropathy. Nephron Clin. Pract. 2007, 106. [Google Scholar] [CrossRef]
- Sato, Y.; Wharram, B.L.; Lee, S.K.; Wickman, L.; Goyal, M.; Venkatareddy, M.; Chang, J.W.; Wiggins, J.E.; Lienczewski, C.; Kretzler, M.; et al. Urine Podocyte mRNAs Mark Progression of Renal Disease. J. Am. Soc. Nephrol. 2009, 20. [Google Scholar] [CrossRef]
- Chugh, S.S. Transcriptional regulation of podocyte disease. Transl. Res. 2007, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastie, N.D. Wilms’ tumour 1 (WT1) in development, homeostasis and disease. Development 2017, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armelloni, S.; Ikehata, M.; Mattinzoli, D.; Li, M.; Alfieri, C.M.; Rastaldi, M.; Messa, P. NeuroD Expression in Podocytes and Interrelationships with Nephrin at Both Nuclear and Cytoplasmic Sites. Cell. Physiol. Biochem. 2018, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.; Kim, E.Y.; Hagmann, H.; Benzing, T.; Dryer, S.E. Opposing effects of podocin on the gating of podocyte TRPC6 channels evoked by membrane stretch or diacylglycerol. Am. J. Physiol. Physiol. 2013, 305. [Google Scholar] [CrossRef]
- Dryer, S.E.; Roshanravan, H.; Kim, E.Y. TRPC channels: Regulation, dysregulation and contributions to chronic kidney disease. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1041–1066. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Greka, A. Calcium-permeable ion channels in the kidney. Am. J. Physiol. Physiol. 2016, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greka, A.; Mundel, P. Calcium Regulates Podocyte Actin Dynamics. Semin. Nephrol. 2012, 32. [Google Scholar] [CrossRef] [Green Version]
- Greka, A.; Mundel, P. Balancing Calcium Signals through TRPC5 and TRPC6 in Podocytes. J. Am. Soc. Nephrol. 2011, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puliti, A.; Rossi, P.I.A.; Caridi, G.; Corbelli, A.; Ikehata, M.; Armelloni, S.; Li, M.; Zennaro, C.; Conti, V.; Vaccari, C.M.; et al. Albuminuria and Glomerular Damage in Mice Lacking the Metabotropic Glutamate Receptor 1. Am. J. Pathol. 2011, 178. [Google Scholar] [CrossRef]
- Reiser, J.; Polu, K.R.; Möller, C.C.; Kenlan, P.; Altintas, M.M.; Wei, C.; Faul, C.; Herbert, S.; Villegas, I.; Avila-Casado, C.; et al. TRPC6 is a glomerular slit diaphragm-associated channel required for normal renal function. Nat. Genet. 2005, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilatovskaya, D.V.; Staruschenko, A. TRPC6 channel as an emerging determinant of the podocyte injury susceptibility in kidney diseases. Am. J. Physiol. Physiol. 2015, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winn, M.P. A Mutation in the TRPC6 Cation Channel Causes Familial Focal Segmental Glomerulosclerosis. Science 2005, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.Y.; Roshanravan, H.; Dryer, S.E. Changes in podocyte TRPC channels evoked by plasma and sera from patients with recurrent FSGS and by putative glomerular permeability factors. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863. [Google Scholar] [CrossRef]
- Möller, C.C.; Wei, C.; Altintas, M.M.; Li, J.; Greka, A.; Ohse, T.; Pippin, J.W.; Rastaldi, M.P.; Wawersik, S.; Schiavi, S.; et al. Induction of TRPC6 Channel in Acquired Forms of Proteinuric Kidney Disease. J. Am. Soc. Nephrol. 2007, 18. [Google Scholar] [CrossRef]
- Topolnik, L.; Azzi, M.; Morin, F.; Kougioumoutzakis, A.; Lacaille, J.-C. mGluR1/5 subtype-specific calcium signalling and induction of long-term potentiation in rat hippocampal oriens/alveus interneurones. J. Physiol. 2006, 575. [Google Scholar] [CrossRef]
- Hartmann, J.; Henning, H.A.; Konnerth, A. mGluR1/TRPC3-mediated Synaptic Transmission and Calcium Signaling in Mammalian Central Neurons. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Watson, L.M.; Bamber, E.; Schnekenberg, R.P.; Williams, J.; Bettencourt, C.; Lickiss, J.; Jayawant, S.; Fawcett, K.; Clokie, S.; Wallis, Y.; et al. Dominant Mutations in GRM1 Cause Spinocerebellar Ataxia Type 44. Am. J. Hum. Genet. 2017, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | RTx | N |
|---|---|---|
| Number of patients | 20 | 18 |
| Age at RBx (years; mean ± SD) | 55 ± 16 | 51 ± 14 |
| Sex (n) (M/F) | 9/11 | 8/10 |
| Prot-U (g/L) | 0.71 ± 0.83 | 2.44 ± 2.94 |
| UC (g/L) | 0.69 ± 0.39 | 0.73 ± 0.25 |
| sCr (mg/dL) | 1.91 ± 0.78 | 2.13 ± 2.50 |
| mGFR (mL/min per 1.73 m2) | 57 ± 28 | 80 ± 47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armelloni, S.; Mattinzoli, D.; Ikehata, M.; Alfieri, C.; Belingheri, M.; Moroni, G.; Cresseri, D.; Passerini, P.; Cerutti, R.; Messa, P. Urinary mRNA Expression of Glomerular Podocyte Markers in Glomerular Disease and Renal Transplant. Diagnostics 2021, 11, 1499. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11081499
Armelloni S, Mattinzoli D, Ikehata M, Alfieri C, Belingheri M, Moroni G, Cresseri D, Passerini P, Cerutti R, Messa P. Urinary mRNA Expression of Glomerular Podocyte Markers in Glomerular Disease and Renal Transplant. Diagnostics. 2021; 11(8):1499. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11081499
Chicago/Turabian StyleArmelloni, Silvia, Deborah Mattinzoli, Masami Ikehata, Carlo Alfieri, Mirco Belingheri, Gabrilella Moroni, Donata Cresseri, Patrizia Passerini, Roberta Cerutti, and Piergiorgio Messa. 2021. "Urinary mRNA Expression of Glomerular Podocyte Markers in Glomerular Disease and Renal Transplant" Diagnostics 11, no. 8: 1499. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11081499