
Identification of MicroRNAs as Potential Blood-Based Biomarkers for Diagnosis and Therapeutic Monitoring of Active Tuberculosis

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
- ATB: patients with ATB were diagnosed based on microbiological and pathological data. A positive MTB culture result from respiratory specimens or the presence of caseous granulomas in lung tissue was used to confirm the diagnosis of ATB. Cases of clinical ATB, which was defined as negative mycobacterial culture findings but favorable clinical and radiological response to anti-TB medication, were also included.
- LTBI: recent contact with ATB patients testing positive for the QFT-GIT were enrolled. The participants had no history of TB and no suggestive symptoms of TB.
- HC: the HCs had to be TB-negative based on the QFT-GIT assay and chest x-rays, and no history of TB. They were free of ATB symptoms, and had no recent contact with patients with ATB.
2.2. AFB Smear and Culture
2.3. QFT-GIT
2.4. miRNA Extraction
2.5. miRNA Sequence Analysis
2.6. miRNA Expression Profiles
2.7. qRT-PCR Validation
2.8. miRNA-Target Gene Network Construction and Gene Ontology (GO) Enrichment Analysis
2.9. Statistical Analysis
3. Results
3.1. Characteristics of Study Participants
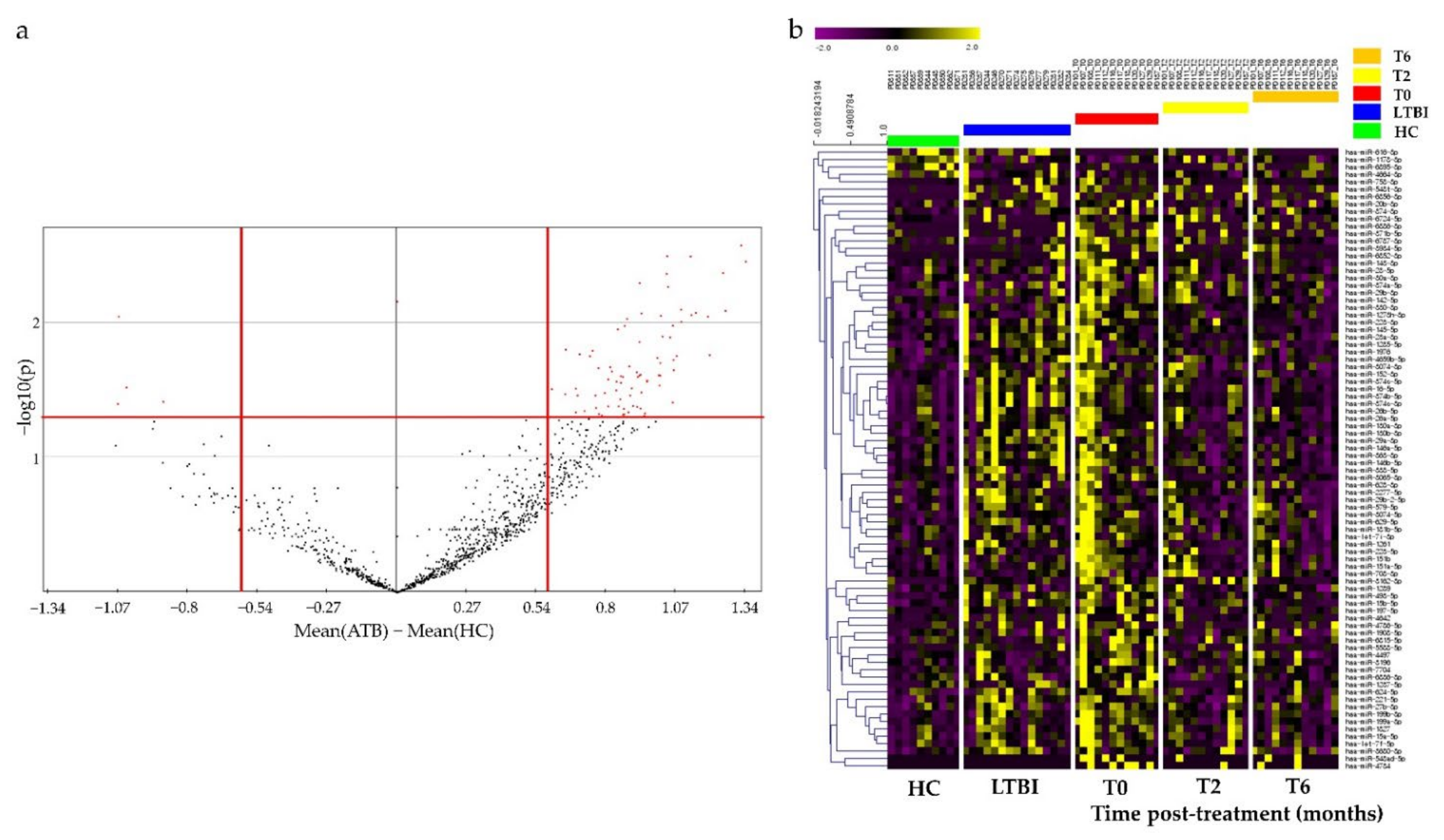
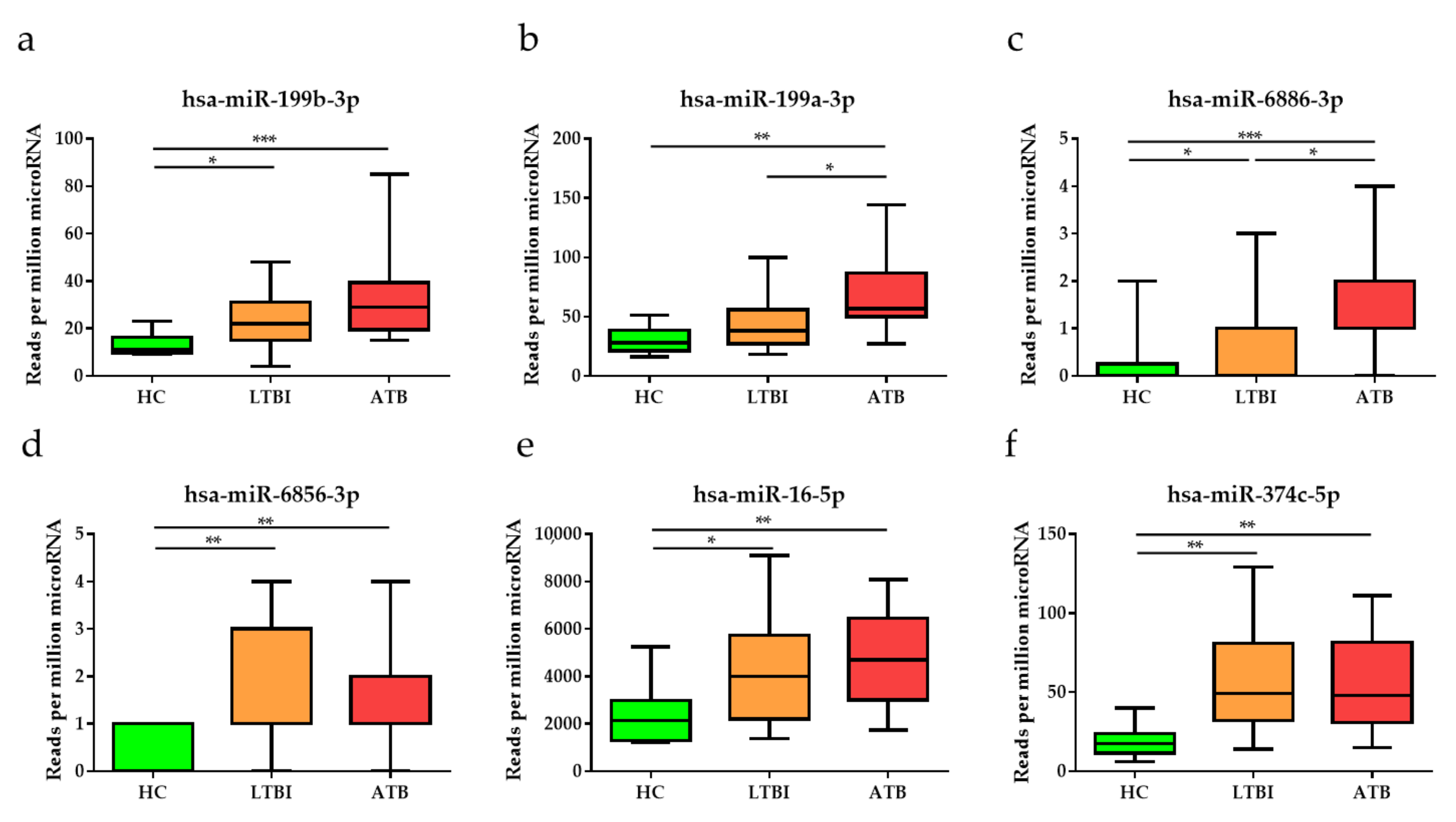
3.2. Identification of Potential miRNAs as Diagnostic Biomarker for TB
3.3. Diagnostic Performance of the Top Six DE miRNAs
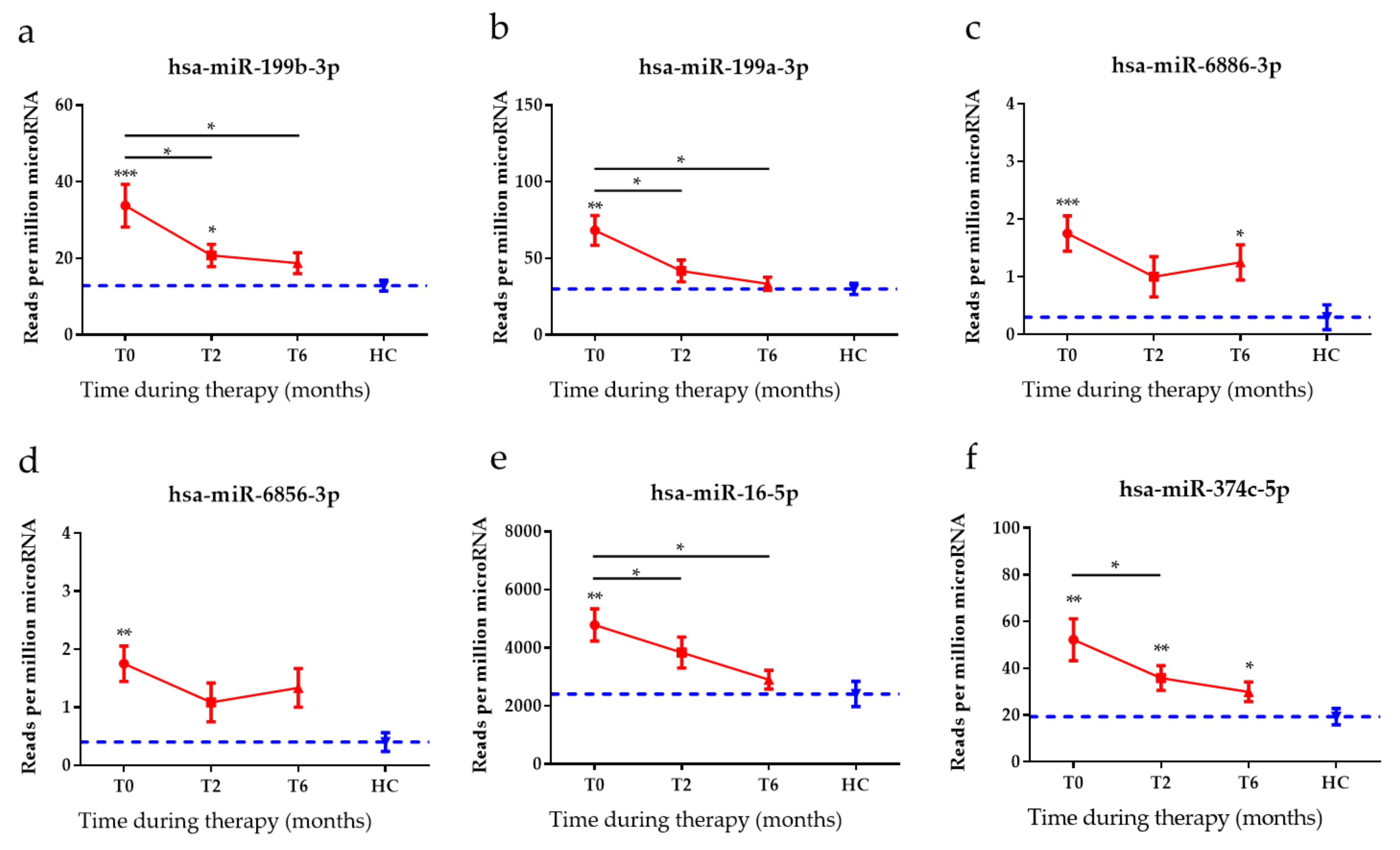
3.4. Anti-TB Treatment Response Profiles of Top Six DE miRNAs
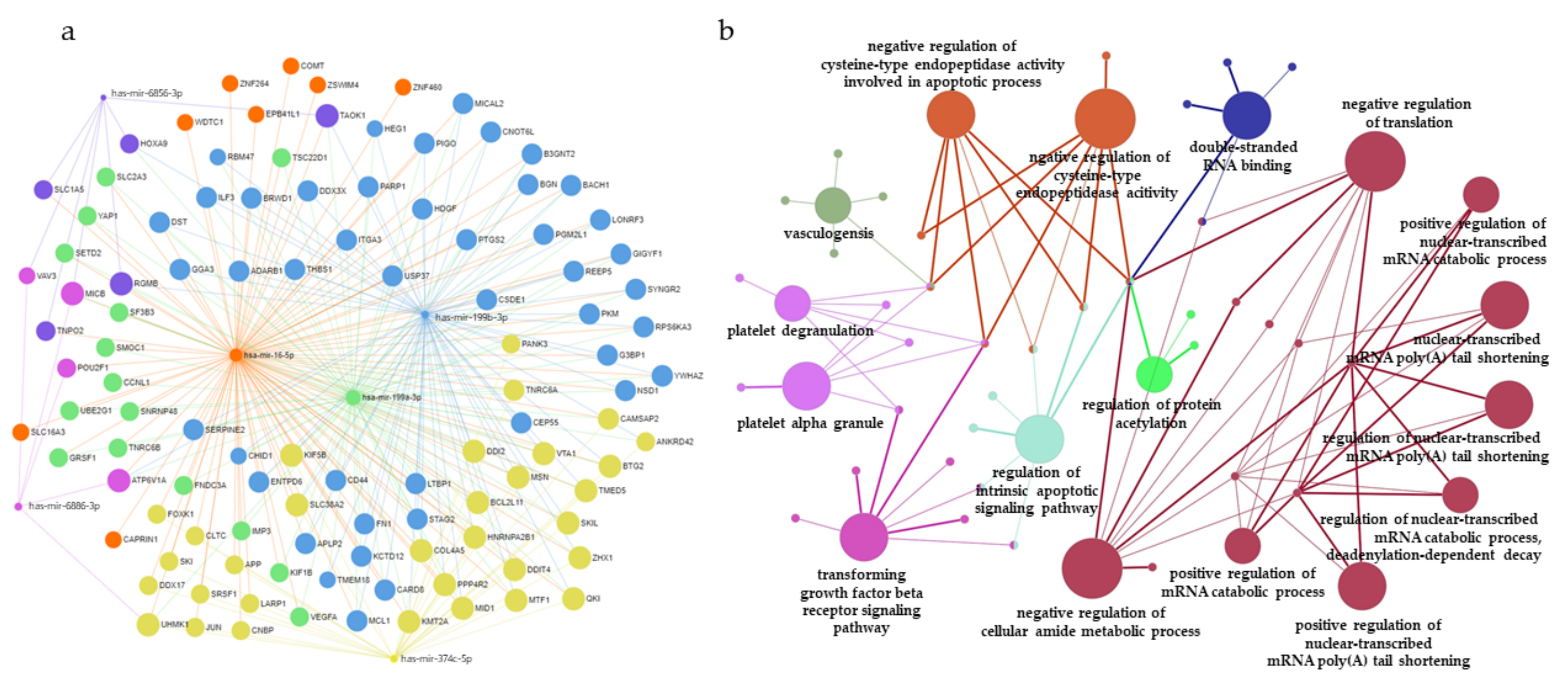
3.5. Construction of miRNAs and Target Gene Networks and GO Analysis of Target Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Konstantinos, A. Testing for tuberculosis. Aust. Prescr. 2010, 33, 12–18. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Tuberculosis Report 2019; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- WHO. Global Tuberculosis Report 2021; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Maitra, A.; Kamil, T.K.; Shaik, M.; Danquah, C.A.; Chrzastek, A.; Bhakta, S. Early diagnosis and effective treatment regimens are the keys to tackle antimicrobial resistance in tuberculosis (TB): A report from Euroscicon’s international TB Summit 2016. Virulence 2017, 8, 1005–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.L.; Bokil, N.J.; Saunders, B.M. Developing new TB biomarkers, are miRNA the answer? Tuberculosis 2019, 118, 101860. [Google Scholar] [CrossRef] [PubMed]
- Khader, S.A.; Divangahi, M.; Hanekom, W.; Hill, P.C.; Maeurer, M.; Makar, K.W.; Mayer-Barber, K.D.; Mhlanga, M.M.; Nemes, E.; Schlesinger, L.S.; et al. Targeting innate immunity for tuberculosis vaccination. J. Clin. Investig. 2019, 129, 3482–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, L.M.; Somoskovi, A.; Gutierrez, C.; Lee, E.; Paramasivan, C.N.; Abimiku, A.; Spector, S.; Roscigno, G.; Nkengasong, J. Laboratory diagnosis of tuberculosis in resource-poor countries: Challenges and opportunities. Clin. Microbiol. Rev. 2011, 24, 314–350. [Google Scholar] [CrossRef] [Green Version]
- Bozkanat, E.; Kaya, H.; Sezer, O.; Caliskan, T.; Kilic, E.; Ciftci, F.; Gumus, S.; Kartaloglu, Z. Comparison of tuberculin skin test and quantiferon-TB gold in tube test for diagnosis of latent tuberculosis infection in health care workers: A cross sectional study. J. Pak. Med. Assoc. 2016, 66, 270–274. [Google Scholar]
- Qiu, X.; Tang, Y.; Zou, R.; Zeng, Y.; Yue, Y.; Li, W.; Qu, Y.; Mu, D. Diagnostic accuracy of interferon-gamma-induced protein 10 for differentiating active tuberculosis from latent tuberculosis: A meta-analysis. Sci. Rep. 2019, 9, 11408. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Acuna, S.M.; Floeter-Winter, L.M.; Muxel, S.M. MicroRNAs: Biological Regulators in Pathogen-Host Interactions. Cells 2020, 9, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.A.; Jhun, B.W.; Kim, S.Y.; Moon, S.M.; Yang, B.; Kwon, O.J.; Daley, C.L.; Shin, S.J.; Koh, W.J. miRNA Expression Profiles and Potential as Biomarkers in Nontuberculous Mycobacterial Pulmonary Disease. Sci. Rep. 2020, 10, 3178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Prasla, Z.; Lee, F.E.; Bedi, B.; Sutliff, R.L.; Sadikot, R.T. MicroRNA-155 Modulates Macrophages’ Response to Non-Tuberculous Mycobacteria through COX-2/PGE2 Signaling. Pathogens 2021, 10, 920. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, M.V.; Ni, B.; Morris, J.D.; Brooks, M.N.; Carlson, T.K.; Bakthavachalu, B.; Schoenberg, D.R.; Torrelles, J.B.; Schlesinger, L.S. Mycobacterium tuberculosis lipomannan blocks TNF biosynthesis by regulating macrophage MAPK-activated protein kinase 2 (MK2) and microRNA miR-125b. Proc. Natl. Acad. Sci. USA 2011, 108, 17408–17413. [Google Scholar] [CrossRef] [Green Version]
- Afum-Adjei Awuah, A.; Ueberberg, B.; Owusu-Dabo, E.; Frempong, M.; Jacobsen, M. Dynamics of T-cell IFN-gamma and miR-29a expression during active pulmonary tuberculosis. Int. Immunol. 2014, 26, 579–582. [Google Scholar] [CrossRef]
- Huang, J.; Jiao, J.; Xu, W.; Zhao, H.; Zhang, C.; Shi, Y.; Xiao, Z. MiR-155 is upregulated in patients with active tuberculosis and inhibits apoptosis of monocytes by targeting FOXO3. Mol. Med. Rep. 2015, 12, 7102–7108. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Li, H.; Shao, H.; Li, C.; Lu, X. MicroRNA-365 in macrophages regulates Mycobacterium tuberculosis-induced active pulmonary tuberculosis via interleukin-6. Int. J. Clin. Exp. Med. 2015, 8, 15458–15465. [Google Scholar]
- Ouimet, M.; Koster, S.; Sakowski, E.; Ramkhelawon, B.; van Solingen, C.; Oldebeken, S.; Karunakaran, D.; Portal-Celhay, C.; Sheedy, F.J.; Ray, T.D.; et al. Mycobacterium tuberculosis induces the miR-33 locus to reprogram autophagy and host lipid metabolism. Nat. Immunol. 2016, 17, 677–686. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Zhang, A.R.; Xu, M.; Lou, J.; Qiu, W.Q. TLR-4/miRNA-32-5p/FSTL1 signaling regulates mycobacterial survival and inflammatory responses in Mycobacterium tuberculosis-infected macrophages. Exp. Cell Res. 2017, 352, 313–321. [Google Scholar] [CrossRef]
- Zonghai, C.; Tao, L.; Pengjiao, M.; Liang, G.; Rongchuan, Z.; Xinyan, W.; Wenyi, N.; Wei, L.; Yi, W.; Lang, B. Mycobacterium tuberculosis ESAT6 modulates host innate immunity by downregulating miR-222-3p target PTEN. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166292. [Google Scholar] [CrossRef]
- Abdalla, A.E.; Duan, X.; Deng, W.; Zeng, J.; Xie, J. MicroRNAs play big roles in modulating macrophages response toward mycobacteria infection. Infect. Genet. Evol. 2016, 45, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. Identification of MicroRNAs as Diagnostic Biomarkers for Breast Cancer Based on the Cancer Genome Atlas. Diagnostics 2021, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Gurtan, A.M.; Sharp, P.A. The role of miRNAs in regulating gene expression networks. J. Mol. Biol. 2013, 425, 3582–3600. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.P.; Graham, C.M.; McNab, F.W.; Xu, Z.; Bloch, S.A.; Oni, T.; Wilkinson, K.A.; Banchereau, R.; Skinner, J.; Wilkinson, R.J.; et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature 2010, 466, 973–977. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Park, Y.; Sim, B.; Kim, J.; Lee, H.; Cho, S.N.; Kang, Y.A.; Lee, S.G. Identification of serum biomarkers for active pulmonary tuberculosis using a targeted metabolomics approach. Sci. Rep. 2020, 10, 3825. [Google Scholar] [CrossRef]
- Yates, M.D.; Pozniak, A.; Uttley, A.H.; Clarke, R.; Grange, J.M. Isolation of environmental mycobacteria from clinical specimens in south-east England: 1973–1993. Int. J. Tuberc. Lung Dis. 1997, 1, 75–80. [Google Scholar]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef]
- Kim, S.; Lee, H.; Kim, H.; Kim, Y.; Cho, J.E.; Jin, H.; Kim, D.Y.; Ha, S.J.; Kang, Y.A.; Cho, S.N.; et al. Diagnostic performance of a cytokine and IFN-gamma-induced chemokine mRNA assay after Mycobacterium tuberculosis-specific antigen stimulation in whole blood from infected individuals. J. Mol. Diagn. 2015, 17, 90–99. [Google Scholar] [CrossRef]
- Lee, S.; Hong, N.; Kim, Y.; Park, S.; Kim, K.J.; Jeong, J.; Jung, H.I.; Rhee, Y. Circulating miR-122-5p and miR-375 as Potential Biomarkers for Bone Mass Recovery after Parathyroidectomy in Patients with Primary Hyperparathyroidism: A Proof-of-Concept Study. Diagnostics 2021, 11, 1704. [Google Scholar] [CrossRef]
- Torres, A.; Torres, K.; Wdowiak, P.; Paszkowski, T.; Maciejewski, R. Selection and validation of endogenous controls for microRNA expression studies in endometrioid endometrial cancer tissues. Gynecol. Oncol. 2013, 130, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, B.; Acharya, A.; Gautam, S.; Ghimire, S.P.; Mishra, G.; Parajuli, N.; Sapkota, B. Advances in diagnosis of Tuberculosis: An update into molecular diagnosis of Mycobacterium tuberculosis. Mol. Biol. Rep. 2020, 47, 4065–4075. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, A.; Beena, P.M.; Devnikar, A.V.; Mali, S. A systemic review on tuberculosis. Indian J. Tuberc. 2020, 67, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Suarez, I.; Funger, S.M.; Kroger, S.; Rademacher, J.; Fatkenheuer, G.; Rybniker, J. The Diagnosis and Treatment of Tuberculosis. Dtsch. Arztebl. Int. 2019, 116, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Sinigaglia, A.; Peta, E.; Riccetti, S.; Venkateswaran, S.; Manganelli, R.; Barzon, L. Tuberculosis-Associated MicroRNAs: From Pathogenesis to Disease Biomarkers. Cells 2020, 9, 2160. [Google Scholar] [CrossRef]
- Yang, T.; Ge, B. miRNAs in immune responses to Mycobacterium tuberculosis infection. Cancer Lett. 2018, 431, 22–30. [Google Scholar] [CrossRef]
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the development of miRNA bioinformatics tools. Brief. Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef] [Green Version]
- Alipoor, S.D.; Adcock, I.M.; Tabarsi, P.; Folkerts, G.; Mortaz, E. MiRNAs in tuberculosis: Their decisive role in the fate of TB. Eur. J. Pharmacol. 2020, 886, 173529. [Google Scholar] [CrossRef]
- Van Rensburg, I.C.; du Toit, L.; Walzl, G.; du Plessis, N.; Loxton, A.G. Decreased neutrophil-associated miRNA and increased B-cell associated miRNA expression during tuberculosis. Gene 2018, 655, 35–41. [Google Scholar] [CrossRef]
- Zhou, S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y.; Cai, L. miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta Pharmacol. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Zhang, S.; Liu, P.; Wang, J.; Du, H. Potential role of microRNAs in the treatment and diagnosis of cervical cancer. Cancer Genet. 2020, 248–249, 25–30. [Google Scholar] [CrossRef]
- Tian, X.; Liu, Y.; Wang, H.; Zhang, J.; Xie, L.; Huo, Y.; Ma, W.; Li, H.; Chen, X.; Shi, P. The role of miR-199b-3p in regulating Nrf2 pathway by dihydromyricetin to alleviate septic acute kidney injury. Free Radic. Res. 2021, 55, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, S.; Gallo, M.G.; Santoro, A.; Brugaletta, R.; Nijhawan, P.; Russo, C.; La Rosa, P.; Raponi, M.; Palamara, A.T.; Bertini, E.S.; et al. Personalized profiles of antioxidant signaling pathway in patients with tuberculosis. J. Microbiol. Immunol. Infect. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Wagh, V.; Urhekar, A.; Modi, D. Levels of microRNA miR-16 and miR-155 are altered in serum of patients with tuberculosis and associate with responses to therapy. Tuberculosis 2017, 102, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Xiong, A.; Wang, J.; Mao, X.L.; Jiang, Y.; Fan, Y. MiR-199a-3p modulates the function of dendritic cells involved in transplantation tolerance by targeting CD86. HLA 2019, 94, 493–503. [Google Scholar] [CrossRef]
- Xiong, H.; Ni, Z.; He, J.; Jiang, S.; Li, X.; He, J.; Gong, W.; Zheng, L.; Chen, S.; Li, B.; et al. LncRNA HULC triggers autophagy via stabilizing Sirt1 and attenuates the chemosensitivity of HCC cells. Oncogene 2017, 36, 3528–3540. [Google Scholar] [CrossRef]
- Hao, S.; Tian, W.; Chen, Y.; Wang, L.; Jiang, Y.; Gao, B.; Luo, D. MicroRNA-374c-5p inhibits the development of breast cancer through TATA-box binding protein associated factor 7-mediated transcriptional regulation of DEP domain containing 1. J. Cell Biochem. 2019, 120, 15360–15368. [Google Scholar] [CrossRef] [Green Version]
- Jee, Y.H.; Wang, J.; Yue, S.; Jennings, M.; Clokie, S.J.; Nilsson, O.; Lui, J.C.; Baron, J. mir-374-5p, mir-379-5p, and mir-503-5p Regulate Proliferation and Hypertrophic Differentiation of Growth Plate Chondrocytes in Male Rats. Endocrinology 2018, 159, 1469–1478. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Cui, L.; Ge, Y.; Shi, Z.; Zhao, K.; Guo, X.; Yang, D.; Yu, H.; Cui, L.; Shan, Y.; et al. Altered serum microRNAs as biomarkers for the early diagnosis of pulmonary tuberculosis infection. BMC Infect. Dis. 2012, 12, 384. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.H.; Chen, C.G.; Zhuang, C.B.; Zeng, Y.L.; Zeng, Z.Z.; Wen, P.H.; Yu, Y.M.; Ming, L.; Zhao, J.W. Integrating serum microRNAs and electronic health records improved the diagnosis of tuberculosis. J. Clin. Lab. Anal. 2021, 35, e23871. [Google Scholar] [CrossRef]
- Cui, J.Y.; Liang, H.W.; Pan, X.L.; Li, D.; Jiao, N.; Liu, Y.H.; Fu, J.; He, X.Y.; Sun, G.X.; Zhang, C.L.; et al. Characterization of a novel panel of plasma microRNAs that discriminates between Mycobacterium tuberculosis infection and healthy individuals. PLoS ONE 2017, 12, e0184113. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, J.; Kim, D.R.; Kang, Y.A.; Bong, S.; Lee, J.; Kim, S.; Lee, N.S.; Sim, B.; Cho, S.N.; et al. Urine IP-10 as a biomarker of therapeutic response in patients with active pulmonary tuberculosis. BMC Infect. Dis. 2018, 18, 240. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | ATB (n = 12) | LTBI (n = 15) | HC (n = 10) | p Value |
|---|---|---|---|---|
| Gender (male, %) | 8 (66.7%) | 6 (40.0%) | 6 (60.0%) | 0.36 |
| Age (mean ± SD) | 41.0 ± 19.6 | 42.1 ± 15.1 | 34.4 ± 7.2 | 0.44 |
| BCG scar (%) | 9 (75.0%) | 8 (53.3%) | 9 (90.0%) | 0.15 |
| AFB stain positive | 2 (16.7%) | |||
| Culture positive | 10 (83.3%) | |||
| QFT-GIT result | ||||
| Positive | 12 (100.0%) | 15 (100.0%) | 0 (0.0%) | |
| Intermediate | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | |
| Negative | 0 (0.0%) | 0 (0.0%) | 10 (100.0%) |
| miRNAs | AUC (95% CI) | Cutoff | Sensitivity (%) (95% CI) | Specificity (%) (95% CI) | p Value |
|---|---|---|---|---|---|
| miR-199b-3p | 0.96 (0.88–1.03) | >17.50 | 91.67 (61.52–99.79) | 90.00% (55.50–99.75) | <0.001 |
| miR-199a-3p | 0.90 (0.78–1.03) | >48.00 | 83.33 (51.59–97.91) | 90.00% (55.50–99.75) | <0.001 |
| miR-6886-3p | 0.88 (0.72–1.03) | >0.50 | 91.67 (61.52–99.79) | 80.00% (44.39–97.48) | <0.001 |
| miR-6856-3p | 0.87 (0.73–1.02) | >0.50 | 91.67 (61.52–99.79) | 60.00% (26.24–87.84) | <0.001 |
| miR-16-5p | 0.87 (0.71–1.02) | >2571.00 | 91.67 (61.52–99.79) | 80.00% (44.39–97.48) | <0.001 |
| miR-374c-5p | 0.86 (0.71–1.01) | >24.50 | 83.33 (51.59–97.91) | 80.00% (44.39–97.48) | <0.001 |
| GO ID | GO Terms | No. of Genes | p Value |
|---|---|---|---|
| GO:0034249 | negative regulation of cellular amide metabolic process | 9 | <0.001 |
| GO:0007179 | transforming growth factor beta receptor signaling pathway | 8 | <0.001 |
| GO:0017148 | negative regulation of translation | 8 | <0.001 |
| GO:2001242 | regulation of intrinsic apoptotic signaling pathway | 7 | <0.001 |
| GO:2000117 | negative regulation of cysteine-type endopeptidase activity | 7 | <0.001 |
| GO:0043154 | negative regulation of cysteine-type endopeptidase activity involved in apoptotic process | 6 | <0.001 |
| GO:0002576 | platelet degranulation | 6 | <0.001 |
| GO:0031091 | platelet alpha granule | 6 | <0.001 |
| GO:0001570 | vasculogenesis | 5 | <0.001 |
| GO:0003725 | double-stranded RNA binding | 5 | <0.001 |
| GO:0061014 | positive regulation of mRNA catabolic process | 4 | <0.001 |
| GO:0000289 | nuclear-transcribed mRNA poly(A) tail shortening | 4 | <0.001 |
| GO:1901983 | regulation of protein acetylation | 3 | <0.001 |
| GO:0060213 | positive regulation of nuclear-transcribed mRNA poly(A) tail shortening | 3 | <0.001 |
| GO:1900151 | regulation of nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay | 3 | <0.001 |
| GO:1900153 | positive regulation of nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay | 3 | <0.001 |
| GO:0060211 | regulation of nuclear-transcribed mRNA poly(A) tail shortening | 3 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Park, H.; Park, S.-B.; Lee, E.J.; Je, M.-A.; Ahn, E.; Sim, B.; Lee, J.; Jin, H.; Lee, K.E.; et al. Identification of MicroRNAs as Potential Blood-Based Biomarkers for Diagnosis and Therapeutic Monitoring of Active Tuberculosis. Diagnostics 2022, 12, 369. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics12020369
Kim J, Park H, Park S-B, Lee EJ, Je M-A, Ahn E, Sim B, Lee J, Jin H, Lee KE, et al. Identification of MicroRNAs as Potential Blood-Based Biomarkers for Diagnosis and Therapeutic Monitoring of Active Tuberculosis. Diagnostics. 2022; 12(2):369. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics12020369
Chicago/Turabian StyleKim, Junseong, Heechul Park, Sung-Bae Park, Eun Ju Lee, Min-A Je, Eunsol Ahn, Bora Sim, Jiyoung Lee, Hyunwoo Jin, Kyung Eun Lee, and et al. 2022. "Identification of MicroRNAs as Potential Blood-Based Biomarkers for Diagnosis and Therapeutic Monitoring of Active Tuberculosis" Diagnostics 12, no. 2: 369. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics12020369