Diversity Estimation and Antimicrobial Activity of Culturable Endophytic Fungi from Litsea cubeba (Lour.) Pers. in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Samples and Isolation of Endophytic Fungi
2.2. Genomic DNA Extraction, PCR Amplification and Molecular Identification
2.3. Estimation and Quantification of Fungal Diversity
2.4. Antimicrobial Activity of Endophytic Fungi
2.5. Statistical Analyses
3. Results
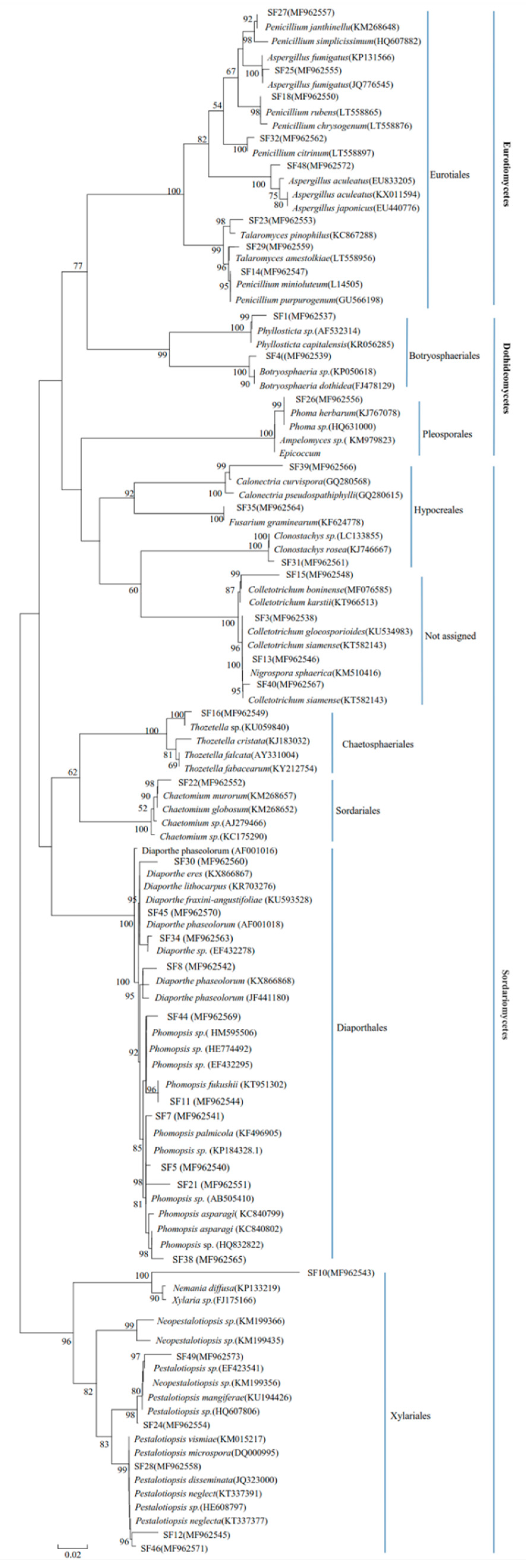
3.1. Identification and Composition of Endophyte Assemblage
3.2. Diversity Estimation of Endophytic Fungi
3.3. In Vitro Antimicrobial Activity of Endophytic Fungi
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marr, A.K.; Gooderham, W.J.; Hancock, R.E. Antibacterial Peptides for Therapeutic Use: Obstacles and Realistic Outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Daisy, B. Bioprospecting for Microbial Endophytes and Their Natural Products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, P.J.; Wang, T.; Dushin, R.G.; Bradford, P.A. Comparative in Vitro Activities of Ac98-6446, a Novel Semisynthetic Glycopeptide Derivative of the Natural Product Mannopeptimycin A, and Other Antimicrobial Agents against Gram-Positive Clinical Isolates. Antimicrob. Agents Chemother. 2004, 48, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Forest Microbiome: Diversity, Complexity and Dynamics. Fems Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef]
- Lewis, S.L.; Edwards, D.P.; Galbraith, D. Increasing Human Dominance of Tropical Forests. Science 2015, 349, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Porrasalfaro, A.; Bayman, P. Hidden Fungi, Emergent Properties: Endophytes and Microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Suryanarayanan, T.S.; Thirunavukkarasu, N.; Rajulu, M.B.G.; Venkatesan, G.; Sukumar, R. Endophytic Fungal Communities in Woody Perennials of Three Tropical Forest Types of the Western Ghats, Southern India. Biodivers. Conserv. 2011, 20, 913–928. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal Endophytes from Higher Plants: A Prolific Source of Phytochemicals and Other Bioactive Natural Products. Fungal Divers. 2010, 41, 1–16. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic Fungal Entomopathogens with Activity against Plant Pathogens: Ecology and Evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Rubini, M.R.; Silva-Ribeiro, R.T.; Pomella, A.W.V.; Maki, C.S.; Araújo, W.L.; Santos, D.R.D.; Azevedo, J.L. Diversity of Endophytic Fungal Community of Cacao (Theobroma cacao L.) and Biological Control of Crinipellis perniciosa,Causal Agent of Witches’ Broom Disease. Int. J. Biol. Sci. 2005, 1, 24–33. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and Taxane Production by Taxomyces Andreanae, an Endophytic Fungus of Pacific Yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Strobel, G.; Stierle, D.; Grothaus, P.; Bignami, G. The Search for a Taxol-Producing Microorganism among the Endophytic Fungi of the Pacific Yew, Taxus Brevifolia. J. Nat. Prod. 1995, 58, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Kour, A.; Shawl, A.S.; Rehman, S.; Sultan, P.; Qazi, P.H.; Suden, P.; Khajuria, R.K.; Verma, V. Isolation and Identification of an Endophytic Strain of Fusarium Oxysporum Producing Podophyllotoxin from Juniperus Recurva. World J. Microbiol. Biotechnol. 2008, 24, 1115–1121. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive Microbial Metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganley, R.J.; Brunsfeld, S.J.; Newcombe, G. A Community of Unknown, Endophytic Fungi in Western White Pine. Proc. Natl. Acad. Sci. USA 2004, 101, 10107–10112. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Li, D.; Liu, T.; Yu, F.; Zhang, Z.; Su, C.; Wang, Y.; Guo, Q.; Liu, Z. Cultural Endophytic Fungi Associated with Dendrobium Officinale: Identification, Diversity Estimation and Their Antimicrobial Potential. Curr. Sci. 2017, 112, 1690–1697. [Google Scholar] [CrossRef]
- Wang, F.; Chang, Z.; Duan, P.; Yan, W.; Xu, Y.; Zhang, L.; Miao, J.; Fan, Y. Hydrothermal Liquefaction of Litsea Cubeba Seed to Produce Bio-Oils. Bioresour. Technol. 2013, 149, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Xu, Y.; Cai, X.H.; Du, Z.Z.; Luo, S.D. Antimicrobially Active Isoquinoline Alkaloids from Litsea Cubeba. Planta Med. 2009, 75, 76–79. [Google Scholar] [CrossRef]
- Lin, B.; Zhang, H.; Zhao, X.X.; Rahman, K.; Wang, Y.; Ma, X.Q.; Zheng, C.J.; Zhang, Q.Y.; Han, T.; Qin, L.P. Inhibitory Effects of the Root Extract of Litsea cubeba (Lour.) Pers. on Adjuvant Arthritis in Rats. J. Ethnopharmacol. 2013, 147, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Liu, Y.Q. Chemical Composition and Antibacterial Activity of Essential Oils from Different Parts of Litsea cubeba. Chem. Biodivers. 2010, 7, 229–235. [Google Scholar] [CrossRef]
- Ho, C.L.; Jie-Ping, O.; Liu, Y.C.; Hung, C.P.; Tsai, M.C.; Liao, P.C.; Wang, E.I.; Chen, Y.L.; Su, Y.C. Compositions and in Vitro Anticancer Activities of the Leaf and Fruit Oils of Litsea cubeba from Taiwan. Nat. Prod. Commun. 2010, 5, 617–620. [Google Scholar] [PubMed]
- Wang, Y.; Chen, J.; Li, D.W.; Zheng, L.; Huang, J. CglCUT1 gene required for cutinase activity and pathogenicity of Colletotrichum gloeosporioides causing anthracnose of Camellia oleifera. Eur. J. Plant Pathol. 2017, 147, 103–114. [Google Scholar] [CrossRef]
- Wulff, E.G.; Sorensen, J.L.; Lubeck, M.; Nielsen, K.F.; Thrane, U.; Torp, J. Fusarium spp. associated with rice Bakanae: Ecology, genetic diversity, pathogenicity and toxigenicity. Environ. Microbiol. 2010, 12, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Shiotani, H.; Yamamoto, M.; Tsuge, T. Insertional mutagenesis and cloning of the genes required for biosynthesis of the host-specific AK-toxin in the Japanese pear pathotype of Alternaria alternata. Mol. Plant Microb. Interact. 1999, 12, 691–702. [Google Scholar] [CrossRef]
- Jaegoon, L.; Donghyun, L.; Sookyoung, P.; Jaeseoun, H.; Youngjin, K. First report of Diaporthe actinidiae, the causal organism of stem-end rot of kiwifruit in Korea. Plant Pathol. J. 2001, 17, 110–113. [Google Scholar]
- Boa, E.R. Recent Research on Bamboos; The Chinese Academy of Forestry: Hanzhou, China, 1985. [Google Scholar]
- Nagarajkumar, M.; Bhaskaran, R.; Velazhahan, R. Involvement of secondary metabolites and extracellular lytic enzymes produced by Pseudomonas fluorescens in inhibition of Rhizoctonia solani, the rice sheath blight pathogen. Microbiol. Res. 2004, 159, 73–81. [Google Scholar] [CrossRef]
- Foster, J.M.; Hausbeck, M.K. Managing Phytophthora Crown and Root Rot in Bell Pepper Using Fungicides and Host Resistance. Plant Dis. 2010, 94, 697–702. [Google Scholar] [CrossRef]
- Jin, Z.; Gao, L.; Zhang, L.; Liu, T.; Yu, F.; Zhang, Z.; Guo, Q.; Wang, B. Antimicrobial Activity of Saponins Produced by Two Novel Endophytic Fungi from Panax notoginseng. Nat. Prod. Res. 2017, 31, 2700–2703. [Google Scholar] [CrossRef]
- Morakotkarn, D.; Kawasaki, H.; Seki, T. Molecular Diversityof Bamboo-Associated Fungi Isolated from Japan. FEMS Microbiol. Lett. 2007, 266, 10–19. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal Rna Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Nascimento, T.L.; Oki, Y.; Lima, D.M.M.; Almeida-Cortez, J.S.; Fernandes, G.W.; Souza-Motta, C.M. Biodiversity of Endophytic Fungi in Different Leaf Ages of Calotropis procera and Their Antimicrobial Activity. Fungal Ecol. 2015, 14, 79–86. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Zhang, C.L.; Lin, F.C.; Kubicek, C.P. Identity, Diversity, and Molecular Phylogeny of the Endophytic Mycobiota in the Roots of Rare Wild Rice (Oryza granulate) from a Nature Reserve in Yunnan, China. Appl. Environ. Microbiol. 2010, 76, 1642–1652. [Google Scholar] [CrossRef]
- Kusari, P.; Kusari, S.; Spiteller, M.; Kayser, O. Endophytic Fungi Harbored in Cannabis sativa L.: Diversity and Potential;as Biocontrol Agents against Host Plant-Specific Phytopathogens. Fungal Divers. 2013, 60, 137–151. [Google Scholar] [CrossRef]
- Gond, S.K.; Mishra, A.; Sharma, V.K.; Verma, S.K.; Kumar, J.; Kharwar, R.N.; Kumar, A. Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Nyctanthes arbor-tristis, a Well-Known Medicinal Plant of India. Mycoscience 2012, 53, 113–121. [Google Scholar] [CrossRef]
- Katoch, M.; Phull, S.; Vaid, S.; Singh, S. Diversity, Phylogeny, Anticancer and Antimicrobial Potential of Fungal Endophytes Associated with Monarda citriodora L. BMC Microbiol. 2017, 17, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Xing, Y.; Chen, J.; Zhang, D.; Guo, S.; Wang, C. Antimicrobial Activities of Endophytic Fungi Isolated from Ophiopogon japonicus (Liliaceae). BMC Complement. Altern. Med. 2012, 12, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Gherbawy, Y.A.; Gashgari, R.M. Molecular Characterization of Fungal Endophytes from Calotropis procera Plants in Taif Region (Saudi Arabia) and Their Antifungal Activities. Plant Biosyst. 2014, 148, 1085–1092. [Google Scholar] [CrossRef]
- Schoch, C.L.; Gi-Ho, S.; Francesc, L.G.; Townsend, J.P.; Jolanta, M.; Valérie, H.; Barbara, R.; P Brandon, M.; Frank, K.; Zheng, W. The Ascomycota Tree of Life: A Phylum-Wide Phylogeny Clarifies the Origin and Evolution of Fundamental Reproductive and Ecological Traits. Syst. Biol. 2009, 58, 224–239. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K. Massively Parallel 454 Sequencing Indicates Hyperdiverse Fungal Communities in Temperate Quercus Macrocarpa phyllosphere. New Phytol. 2009, 184, 438–448. [Google Scholar] [CrossRef]
- Ho, M.Y.; Chung, W.C.; Huang, H.C.; Chung, W.H.; Chung, W.H. Identification of Endophytic Fungi of Medicinal Herbs of Lauraceae and Rutaceae with Antimicrobial Property. Taiwania 2012, 57, 229–241. [Google Scholar]
- Šamonil, P.; Valtera, M.; Bek, S.; Šebková, B.; Vrška, T.; Houška, J. Soil Variability through Spatial Scales in a Permanently Disturbed Natural Spruce-Fir-Beech Forest. Eur. J. For. Res. 2011, 130, 1075–1091. [Google Scholar] [CrossRef]
- Lebeis, S.L. Greater Than the Sum of Their Parts: Characterizing Plant Microbiomes at the Community-Level. Curr. Opin. Plant Biol. 2015, 24, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Vorholt, J.A. Microbial Life in the Phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.X.; Zou, W.X. Endophytes: A Rich Source of Functional Metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Meng, J.; Lu, H.; Chen, G.; Shi, G.; Zhang, T.; Tan, R. Metabolites of Colletotrichum gloeosporioides, an Endophytic Fungus in Artemisia Mongolica. J. Nat. Prod. 2000, 63, 1529–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, F.; Qin, J.; Wang, D.; Zhang, J.; Zhang, Y.; Zhang, S.; Pan, H. Efficacy Assessment of Antifungal Metabolites from Chaetomium globosum No. 05, a New Biocontrol Agent, against Setosphaeria turcica. Biol. Control 2013, 64, 90–98. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, G.J.; Jang, K.S.; Lim, H.K.; Kim, H.T.; Cho, K.Y.; Kim, J.C. Antifungal Activity against Plant Pathogenic Fungi of Chaetoviridins Isolated from Chaetomium globosum. FEMS Microbiol. Lett. 2005, 252, 309–313. [Google Scholar] [CrossRef]
- Yang, L.; Xie, J.; Jiang, D.; Fu, Y.; Li, G.; Lin, F. Antifungal Substances Produced by Penicillium oxalicum Strain Py-1—Potential Antibiotics against Plant Pathogenic Fungi. World J. Microbiol. Biotechnol. 2008, 24, 909–915. [Google Scholar] [CrossRef]
- Sahoo, S.; Sarangi, S.; Kerry, R.G. Bioprospecting of Endophytes for Agricultural and Environmental Sustainability. Microb. Biotechnol. 2017, 429–458. [Google Scholar]


{kind=link}
| Endophytic Fungal Taxon | Isolate Codes | Accession Numbers | The Closet Genbank Taxa | Similarity (%) | Numbers of Isolates from Plant Tissues | Total Abundance (Percentage Recovery) | |||
|---|---|---|---|---|---|---|---|---|---|
| Roots | Stems | Leaves | Fruits | ||||||
| Aspergillus fumigatus 1 | SF25 | MF962555 | Aspergillus fumigatus (KP131566.1) | 99 | 6 | 0 | 0 | 0 | 0.62 |
| Aspergillus fumigatus 2 | SF48 | MF962572 | Aspergillus fumigatus (EU833205.1) | 99 | 1 | 0 | 0 | 0 | 0.10 |
| Botryosphaeria dothidea | SF4 | MF962539 | Botryosphaeria dothidea (FJ478129.1) | 99 | 1 | 46 | 11 | 6 | 6.60 |
| Clonostachys sp. | SF31 | MF962561 | Clonostachys sp. (LC133855.1) | 99 | 5 | 2 | 3 | 0 | 1.03 |
| Calonectria curvispora | SF39 | MF962566 | Calonectria curvispora (GQ280568.1) | 99 | 25 | 2 | 0 | 0 | 2.78 |
| Chaetomium globosum | SF22 | MF962552 | Chaetomium globosum (KM268652.1) | 99 | 1 | 0 | 1 | 0 | 0.21 |
| Colletotrichum boninense | SF15 | MF962548 | Colletotrichum boninense (MF076585.1) | 99 | 0 | 1 | 3 | 0 | 0.41 |
| Colletotrichum gloeosporioides 1 | SF40 | MF962567 | Colletotrichum gloeosporioides (EU552111.1) | 99 | 2 | 32 | 284 | 68 | 39.79 |
| Colletotrichum gloeosporioides 2 | SF3 | MF962538 | Colletotrichum gloeosporioides (KU534983.1) | 99 | 4 | 0 | 41 | 4 | 5.06 |
| Diaporthe phaseolorum 1 | SF8 | MF962542 | Diaporthe phaseolorum (KX866868.1) | 99 | 7 | 37 | 18 | 4 | 6.80 |
| Diaporthe phaseolorum 2 | SF45 | MF962570 | Diaporthe phaseolorum (AF001018.2) | 99 | 0 | 0 | 0 | 9 | 0.93 |
| Diaporthe eres | SF30 | MF962560 | Diaporthe eres (KX866867.1) | 97 | 0 | 11 | 0 | 0 | 1.13 |
| Diaporthe sp. | SF34 | MF962563 | Diaporthe sp. (EF42278.1) | 97 | 0 | 7 | 0 | 0 | 0.72 |
| Fusarium graminearum | SF35 | MF962564 | Fusarium graminearum (KF624778.1) | 99 | 5 | 4 | 11 | 10 | 3.09 |
| Nigrospora sphaerica | SF13 | MF962546 | Nigrospora sphaerica (KM510416.1) | 100 | 0 | 0 | 1 | 6 | 0.72 |
| Nemania diffusa | SF10 | MF962543 | Nemania diffusa (KP133219.1) | 99 | 3 | 4 | 0 | 0 | 0.72 |
| Phomopsis sp.1 | SF5 | MF962540 | Phomopsis sp. (KP184328.1) | 99 | 0 | 8 | 29 | 0 | 3.82 |
| Phomopsis sp.2 | SF7 | MF962541 | Phomopsis sp. (JX436795.1) | 98 | 14 | 12 | 10 | 13 | 5.05 |
| Phomopsis sp.3 | SF21 | MF962551 | Phomopsis sp. (AB505410.1) | 97 | 3 | 3 | 0 | 4 | 1.03 |
| Phomopsis sp.4 | SF38 | MF962565 | Phomopsis sp. (HQ832822.1) | 99 | 1 | 22 | 0 | 0 | 2.37 |
| Phomopsis sp.5 | SF44 | MF962569 | Phomopsis sp. (HM595506.1) | 99 | 0 | 0 | 0 | 9 | 0.93 |
| Phomopsis fukushii | SF11 | MF962544 | Phomopsis fukushii (KT951302.1) | 97 | 1 | 0 | 2 | 1 | 0.41 |
| Phyllosticta capitalensis | SF1 | MF962537 | Phyllosticta capitalensis (KR056285.1) | 100 | 1 | 15 | 9 | 9 | 3.51 |
| Pestalotiopsis sp.1 | SF24 | MF962554 | Pestalotiopsis sp. (HQ607806.1) | 99 | 0 | 9 | 0 | 0 | 0.93 |
| Pestalotiopsis sp.2 | SF46 | MF962571 | Pestalotiopsis sp. (HE608797.1) | 99 | 2 | 0 | 0 | 0 | 0.21 |
| Pestalotiopsis sp.3 | SF49 | MF962573 | Pestalotiopsis sp. (EF423541.1) | 100 | 1 | 0 | 0 | 5 | 0.62 |
| Pestalotiopsis disseminata | SF28 | MF962558 | Pestalotiopsis disseminate (JQ323000.1) | 99 | 3 | 13 | 0 | 1 | 0.62 |
| Pestalotiopsis vismiae | SF12 | MF962545 | Pestalotiopsis vismiae (KM015217.1) | 99 | 2 | 1 | 3 | 0 | 1.75 |
| Penicillium rubens | SF18 | MF962550 | Penicillium rubens (LT558865.1) | 100 | 0 | 1 | 0 | 0 | 0.10 |
| Penicillium janthinellum | SF27 | MF962557 | Penicillium janthinellum (KM268648.1) | 99 | 29 | 0 | 0 | 0 | 2.99 |
| Penicillium citrinum | SF32 | MF962562 | Penicillium citrinum (LT558897.1) | 100 | 5 | 0 | 0 | 0 | 0.52 |
| Penicillium minioluteum | SF14 | MF962547 | Penicillium minioluteum (L14505.1) | 99 | 3 | 0 | 0 | 0 | 0.31 |
| Phoma sp. | SF26 | MF962556 | Phoma sp. (HQ631000.1) | 99 | 5 | 9 | 11 | 0 | 2.58 |
| Thozetella sp. | SF16 | MF962549 | Thozetella sp. (KU059840.1) | 96 | 1 | 1 | 0 | 0 | 0.21 |
| Talaromyces sp. | SF23 | MF962553 | Talaromyces sp. (KU556510.1) | 99 | 3 | 0 | 0 | 0 | 0.31 |
| Talaromyces amestolkiae | SF29 | MF962559 | Talaromyces amestolkiae (LT558956.1) | 99 | 8 | 1 | 1 | 0 | 1.03 |
| Total | 142 | 241 | 438 | 149 | 100.00 | ||||
| Parts | No. of Tissue | No. of Fungi | No. of Strains | No. of Genus | CR % | IR % | H’ | Ds | E |
|---|---|---|---|---|---|---|---|---|---|
| Root | 450 | 142 | 142 | 27 | 32.22 | 31.56 | 2.74 | 0.90 | 0.83 |
| Stem | 450 | 232 | 241 | 21 | 51.56 | 53.56 | 2.56 | 0.90 | 0.84 |
| Leaf | 450 | 413 | 438 | 16 | 91.78 | 97.33 | 1.43 | 0.56 | 0.51 |
| Fruit | 450 | 139 | 149 | 14 | 30.89 | 33.11 | 1.99 | 0.76 | 0.75 |
| Total | 1800 | 926 | 970 | 36 | 51.44 | 53.89 | 2.52 | 0.82 | 0.70 |
| No. | Endophytic fungi | Inhibition Ratio of Pathogen Mycelium Growth (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| ① | ② | ③ | ④ | ⑤ | ⑥ | ⑦ | ||
| SF11 | Phomopsis fukushii | 46.15 ± 0.33 W− | 55.56 ± 0.53 W− | 45.91 ± 0.71 W+ | 29.52 ± 0.32 W+ | 46.56 ± 0.52 W− | 55.35 ± 1.61 W+ | — |
| SF14 | Penicillium minioluteum | 39.23 ± 0.27 W+ | 58.82 ± 0.28 W+ | 52.20 ± 0.21 W+ | 60.95 ± 0.41 W− | 52.67 ± 0.47 W+ | 53.46 ± 0.31 W+ | 55.49 ± 0.37 W+ |
| SF22 | Chaetomium globosum | 58.61 ± 0.44 W+ | 73.43 ± 0.43 W+ | 53.12 ± 0.14 W+ | 70.23 ± 0.30 W+ | 73.20 ± 0.24 W+ | 78.43 ± 0.49 W+ | 56.60 ± 0.35 W+ |
| SF23 | Talaromyces sp. | 42.31 ± 0.27 W+ | 64.05 ± 0.18 W+ | 49.06 ± 0.21 W+ | 42.86 ± 0.24 W+ | 55.73 ± 0.27 W+ | 52.83 ± 0.28 W+ | 52.44 ± 0.34 W− |
| SF24 | Pestalotiopsis sp.1 | 46.92 ± 0.25 W+ | 52.94 ± 0.31 W− | 56.60 ± 0.29 W+ | 68.57 ± 0.43 W− | 57.25 ± 0.14 W+ | 50.31 ± 0.28 W+ | 39.63 ± 0.39 W− |
| SF27 | Penicillium janthinellum | 38.46 ± 0.41 W+ | 39.22 ± 0.20 W+ | 47.17 ± 0.41 W− | 31.43 ± 0.40 W− | 53.44 ± 0.32 W+ | 40.88 ± 0.37 W+ | 35.37 ± 0.21 W+ |
| SF28 | Pestalotiopsis disseminata | 50.00 ± 0.23 W+ | 58.17 ± 0.21 W+ | 57.23 ± 0.27 W+ | 39.05 ± 0.21 W− | 45.04 ± 0.21 W− | 48.43 ± 0.31 W− | 25.61 ± 0.37 W− |
| SF29 | Talaromyces amestolkiae | 60.77 ± 0.43 W− | 78.43 ± 0.18 W− | 60.38 ± 0.37 W− | 59.05 ± 0.19W+ | 73.28 ± 0.27 W− | 64.78 ± 0.42 W+ | 51.22 ± 0.43 W− |
| SF31 | Clonostachys sp. | 33.08 ± 0.38 W+ | 47.06 ± 0.35 W+ | 48.43 ± 0.41 W+ | 28.57 ± 0.33 W− | 32.06 ± 0.25 W− | — | — |
| SF32 | Penicillium citrinum | 50.77 ± 0.45 W+ | 62.75 ± 0.29 W+ | 61.01 ± 0.32 W+ | 39.05 ± 0.21 W+ | — | — | 80.49 ± 0.27 W− |
| SF35 | Fusarium graminearum | 36.92 ± 0.43 W+ | 49.67 ± 0.33 W+ | 53.46 ± 0.31 W+ | 37.14 ± 0.26 W+ | 49.62 ± 0.28 W+ | 40.88 ± 0.36 W− | 32.93 ± 0.17 W− |
| SF39 | Calonectria curvispora | 45.38 ± 0.25 W+ | — | — | — | 70.13 ± 0.19 W+ | 59.12 ± 0.18 W− | — |
| SF44 | Phomopsis sp.5 | 50.77 ± 0.40 W+ | 71.42 ± 0.25 W+ | 57.86 ± 0.32 W− | 40.95 ± 0.44 W− | 49.62 ± 0.36 W+ | 55.35 ± 0.23 W+ | — |
| SF49 | Pestalotiopsis sp.3 | 46.92 ± 0.24 W+ | 70.21 ± 0.35 W+ | 56.60 ± 0.35 W+ | 52.38 ± 0.33 W− | 51.91 ± 0.39 W+ | 57.23 ± 0.31 W+ | — |
| No. | Endophytic fungi | Inhibition Ratio of Pathogen Mycelium Growth (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| ① | ② | ③ | ④ | ⑤ | ⑥ | ⑦ | ||
| SF14 | Penicillium minioluteum | 76.32 ± 1.35 aB | 75.21 ± 1.63 bBC | 69.00 ± 2.40 aC | 61.08 ± 1.85 bD | 87.87 ± 1.97 aA | 84.73 ± 4.03 aA | 60.01 ± 3.03 bD |
| SF22 | Chaetomium globosum | 66.18 ± 3.98 bC | 91.73 ± 1.67 aA | 66.67 ± 3.39 aC | 77.60 ± 2.72 aB | 80.00 ± 6.25 abB | 93.24 ± 2.10 aA | 61.00 ± 3.82 bC |
| SF23 | Talaromyces sp. | 79.34 ± 3.22 aAB | 55.10 ± 0.85 dD | 37.33 ± 1.31 bF | 45.21 ± 3.25 cE | 80.63 ± 3.55 abA | 65.00 ± 2.19 bC | 73.60 ± 1.36 aB |
| SF27 | Penicillium janthinellum | 80.66 ± 2.80 aA | 71.07 ± 3.87 bcAB | 29.87 ± 6.01 bcE | 42.16 ± 3.55 cDE | 74.80 ± 4.02 bAB | 56.08 ± 8.85 bcCD | 64.78 ± 3.76 bBC |
| SF29 | Talaromyces amestolkiae | 27.37 ± 4.30 dCD | 64.19 ± 6.58 cdA | 25.33 ± 6.73 cD | 46.73 ± 6.43 cB | 50.65 ± 4.27 cAB | 45.00 ± 5.53 cdB | 42.14 ± 3.30 cBC |
| SF32 | Penicillium citrinum | 50.34 ± 3.70 cB | 60.24 ± 5.04 dA | 61.23 ± 2.53 aA | 42.23 ± 3.57 cBC | 32.76 ± 1.55 dD | 33.78 ± 2.70 dCD | 45.12 ± 2.91 cB |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Yang, D.; Zhang, L.; Chen, Y.; Hu, X.; Li, L.; Liang, J. Diversity Estimation and Antimicrobial Activity of Culturable Endophytic Fungi from Litsea cubeba (Lour.) Pers. in China. Forests 2019, 10, 33. https://0-doi-org.brum.beds.ac.uk/10.3390/f10010033
Wu F, Yang D, Zhang L, Chen Y, Hu X, Li L, Liang J. Diversity Estimation and Antimicrobial Activity of Culturable Endophytic Fungi from Litsea cubeba (Lour.) Pers. in China. Forests. 2019; 10(1):33. https://0-doi-org.brum.beds.ac.uk/10.3390/f10010033
Chicago/Turabian StyleWu, Fei, Dingchao Yang, Linping Zhang, Yanliu Chen, Xiaokang Hu, Lei Li, and Junsheng Liang. 2019. "Diversity Estimation and Antimicrobial Activity of Culturable Endophytic Fungi from Litsea cubeba (Lour.) Pers. in China" Forests 10, no. 1: 33. https://0-doi-org.brum.beds.ac.uk/10.3390/f10010033