Nitrogen and Phosphorus Resorption in Planted Forests Worldwide
by
 ,
,
Dalong Jiang
1,2 ,
,
Qinghong Geng
1,
Qian Li
3,
Yiqi Luo
4,
Jason Vogel
2,
Zheng Shi
1,
Honghua Ruan
1 and
Xia Xu
1,* 1
College of Biology and the Environment, Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
2
School of Forest Resources and Conservation, University of Florida, Gainesville, FL 32608, USA
3
Advanced Analysis and Testing Center, Nanjing Forestry University, Nanjing 210037, China
4
Center for Ecosystem Science and Society, Northern Arizona University, Flagstaff, AZ 86011, USA
*
Author to whom correspondence should be addressed.
Forests 2019, 10(3), 201; https://0-doi-org.brum.beds.ac.uk/10.3390/f10030201
Submission received: 31 January 2019
/
Revised: 18 February 2019
/
Accepted: 25 February 2019
/
Published: 26 February 2019
(This article belongs to the Special Issue Nutrient Cycling in Forest Ecosystems)
Abstract
:Nutrient resorption from senescing leaves is one of the plants’ essential nutrient conservation strategies. Parameters associated with resorption are important nutrient-cycling constraints for accurate predictions of long-term primary productivity in forest ecosystems. However, we know little about the spatial patterns and drivers of leaf nutrient resorption in planted forests worldwide. By synthesizing results of 146 studies, we explored nitrogen (N) and phosphorus (P) resorption efficiency (NRE and PRE) among climate zones and tree functional types, as well as the factors that play dominant roles in nutrient resorption in plantations globally. Our results showed that the mean NRE and PRE were 58.98% ± 0.53% and 60.21% ± 0.77%, respectively. NRE significantly increased from tropical to boreal zones, while PRE did not significantly differ among climate zones, suggesting differential impacts of climates on NRE and PRE. Plant functional types exert a strong influence on nutrient resorption. Conifer trees had higher PRE than broadleaf trees, reflecting the adaptation of the coniferous trees to oligotrophic habitats. Deciduous trees had lower PRE than evergreen trees that are commonly planted in P-limited low latitudes and have long leaf longevity with high nutrient use efficiency. While non-N-fixing trees had higher NRE than N-fixing trees, the PRE of non-N-fixing trees was lower than that of N-fixing trees, indicating significant impact of the N-fixing ability on the resorption of N and P. Our multivariate regression analyses showed that variations in NRE were mainly regulated by climates (mean annual precipitation and latitude), while variations in PRE were dominantly controlled by green leaf nutrient concentrations (N and P). Our results, in general, suggest that the predicted global warming and changed precipitation regimes may profoundly affect N cycling in planted forests. In addition, green leaf nutrient concentrations may be good indicators for PRE in planted forests.
1. Introduction
Nutrient availability is a critical constraint in plant productivity and carbon (C) stocks in terrestrial ecosystems [1,2]. Nutrient resorption, through which nutrients are reabsorbed before leaf senescence and reused for plant growth directly, is an essential component of nutrient conservation strategies [3,4]. It affects key ecosystem processes such as nutrient uptake [5,6], plant competition [7,8], C cycling and resource-use efficiency [9,10], and hence productivity [1,11]. From a physiological perspective, resorption efficiency is an important issue in nutrient conservation [6,12]. In ecosystem modeling, reliable estimates of resorption efficiency are of key importance for modeling nutrient cycling and for quantifying ecosystem productivity, particularly in the new generation of coupled global models [13,14]. While previous synthesis studies mainly focus on the resorption efficiency of natural forests [1,4,5,9], we know little about nutrient resorption and its associated drivers in planted forests worldwide.
Increasing demands for timber products have promoted more research on plantations, which require more nutrients for their rapid growth [15,16]. Nutrient resorption in planted forests is less studied, since many synthesis studies excluded data of nutrient resorption from plantations [5,7,17]. Additionally, few nutrient-resorption studies have separated planted forests from other forest types [18,19,20,21]. Planted forests may have different nutrient resorption efficiencies in comparison to natural forests, since they are commonly pioneer tree species and grow on nutrient-poor soils [4,22]. Recent studies show that plantations had significantly higher nitrogen resorption efficiency (NRE) than the adjacent natural forests in northeast China and inland Hokkaido, Japan [16,23]. Phosphorous resorption efficiency (PRE) was found to be lower in plantations than in natural forests [24]. Plant productivity is largely constrained by nutrient availability [25,26], making exploration of nutrient resorption efficiency of high necessity in planted forests worldwide.
Empirical and regional studies have undoubtedly advanced our understanding of nutrient resorption in planted forests [4,27,28,29,30]. A synthesis study showed that while phosphorous resorption efficiency (PRE) was higher in subtropical than in temperate zones, NRE did not differ between the two climate zones in China’s planted forests [24]. In contrary, Zhang et al. found that NRE increased and PRE first increased and then decreased along latitude in Metasequoia glyptostroboides forests in the east coast of China [27]. In terms of plant functional types, while Lal et al. found that deciduous species had higher NRE and PRE than evergreen species in a dry tropical environment [31], Machado et al. showed that evergreen species presented higher PRE, but not NRE, than deciduous species in plantations in the Brazilian Amazon [29]. Few studies have explored NRE in N-fixing trees vs. non-N-fixing trees in planted forests, though Yuan et al. found lower NRE in N-fixing trees than in the non-N-fixing trees (39% vs. 50%) in a semi-arid region of northern China [32]. A site study in the Karst ecosystem of southwestern China showed higher NRE and lower PRE for the conifer Pinus yunnanensis Franch. than for the broadleaf Eucalyptus maideni F. Muell. [33]. Much uncertainty still surrounds the global patterns of nutrient resorption efficiency of plantations across different climate zones and functional groups. This lack of clear understanding limits the accuracy in predicting long-term primary productivity in forest ecosystems. Therefore, it is urgent to examine nutrient resorption of planted forests regarding spatial patterns and drivers.
Planted forests, accounting for 7% of forest areas globally in 2015, are one of the essential components of terrestrial forest ecosystems providing us economic and social benefits [34,35]. Moreover, plantations have been advocated as important C sinks to mitigate future climate change [36,37]. Nutrient resorption enables plants to store reabsorbed nutrients for rapid and sustained growth at the beginning of the next growing season [38,39]. Understanding the nutrient resorption patterns in plantations is thus beneficial for improving nutrient conservation and management of planted forests. In this systematic review, we aimed to: (1) explore the global patterns of nutrient resorption (N and P) in planted forests and (2) identify various factors that play important roles in nutrient resorption worldwide.
2. Materials and Methods
2.1. Data Description
In this study, we compiled a dataset of 149 independent sampling sites based on 146 peer-reviewed papers about NRE and PRE in planted forests worldwide (Figure 1, Supplementary materials 1 and 2). In total, 643 observations for NRE and 539 observations for PRE were included in the dataset. We utilized the searching tools of ISI Web of Science and Google Scholar for retrieving articles on nutrient resorption, and the China National Knowledge Infrastructure (CNKI) to search for papers published in Chinese. Both methods included combinations of the terms ‘nutrient resorption’ or ‘nutrient retranslocation’ or ‘nutrient reabsorption’ and ‘concentration’, ‘forest’, and ‘tree’ as the searching keywords. We selected articles and extracted data (using Graph Digitizer 2.24, http://getdata-graph-digitizer.com/) with the following criteria: (1) N and P concentrations based on dry mass in green and senesced leaves were directly available or could be calculated based on presented tables or graphs; (2) we only selected data of trees from plots identified as planted forests; (3) any data from greenhouse, nursery, fertilized, and polluted sites were excluded; and (4) any data from cases of possible ‘premature senescence’ (e.g., drought, pests,…) were also eliminated.
NRE and PRE were estimated by percent reduction between green and senesced leaves [5,7,40,41]. However, to eliminate the underestimations of nutrient resorption, we used a mass loss correction factor (MLCF) with the calculation as follows [1,42]:
where NuRE is nutrient resorption efficiency; Nugreen and Nusenesced are nutrient concentrations in green and senesced leaves, respectively. The MLCF was calculated from the mass of green and senesced leaves or from the percentage of leaf mass loss during senescence. If green and senesced leaf mass and percentage of leaf mass loss were not available from a specific study, we used 0.745 for conifers, 0.780 for evergreen broad-leaved species, and 0.784 for deciduous broad-leaved species [1].
For studies that reported leaf nutrient concentrations during the growing season, we used maximum values for green leaf nutrient concentrations [5]. We also collected climate variables including latitude (°, LAT), longitude (°), mean annual temperature (°C, MAT), and mean annual precipitation (mm, MAP) and forest properties (e.g., tree species, functional groups, and stand ages) from those papers. Climate zones were subdivided into boreal, temperate, subtropical, and tropical regions [13]. Functional groups were separated into coniferous vs. broadleaf trees, deciduous vs. evergreen trees, and non-N-fixing vs. N-fixing trees.
2.2. Statistical Analysis
Before statistical analysis, the normality and homoscedasticity of the NRE and PRE were verified by the Kolmogorov–Smirnov test and Levene’s test, respectively. One-way analysis of variance (ANOVA) was performed to analyze the differences between NRE and PRE. Linear mixed-effects models (LMEs) were applied to quantify the effects of climate zones and different functional groups on NRE and PRE worldwide. Climate zones and functional groups were treated as fixed factors, and studies were treated as a random factor. The significance of fixed effects terms was assessed via ANOVA with F tests. If the differences were significant, post hoc multiple comparisons were subsequently conducted using Duncan’s test (p = 0.05). Stepwise regression analyses were used to explore the controls on the variation of NRE and PRE associated with climate variables, leaf nutrient status, and stand ages. All statistical analyses were performed using SPSS 22.0 for Windows (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Nutrient Resorption Patterns in Planted Forests
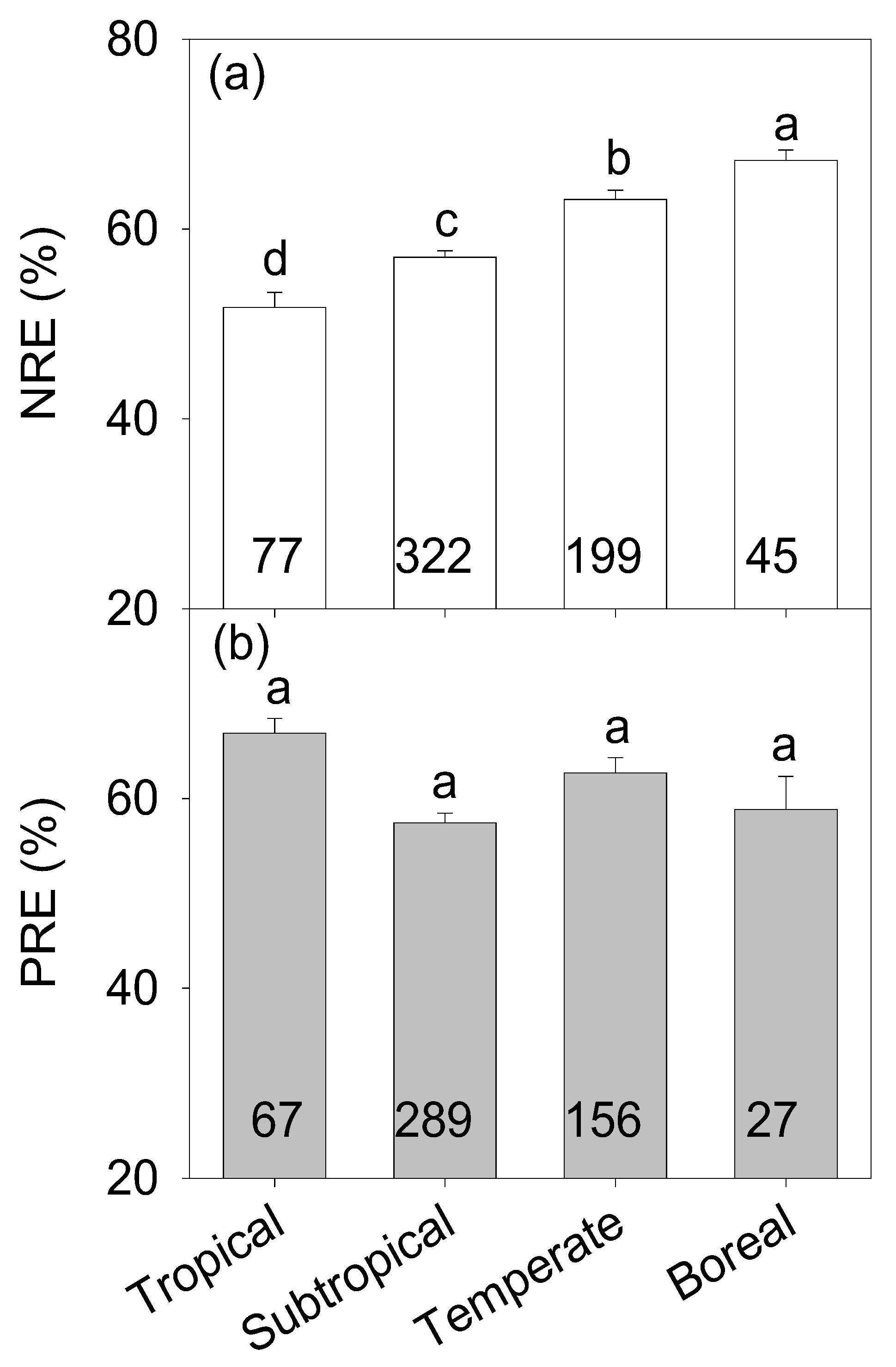
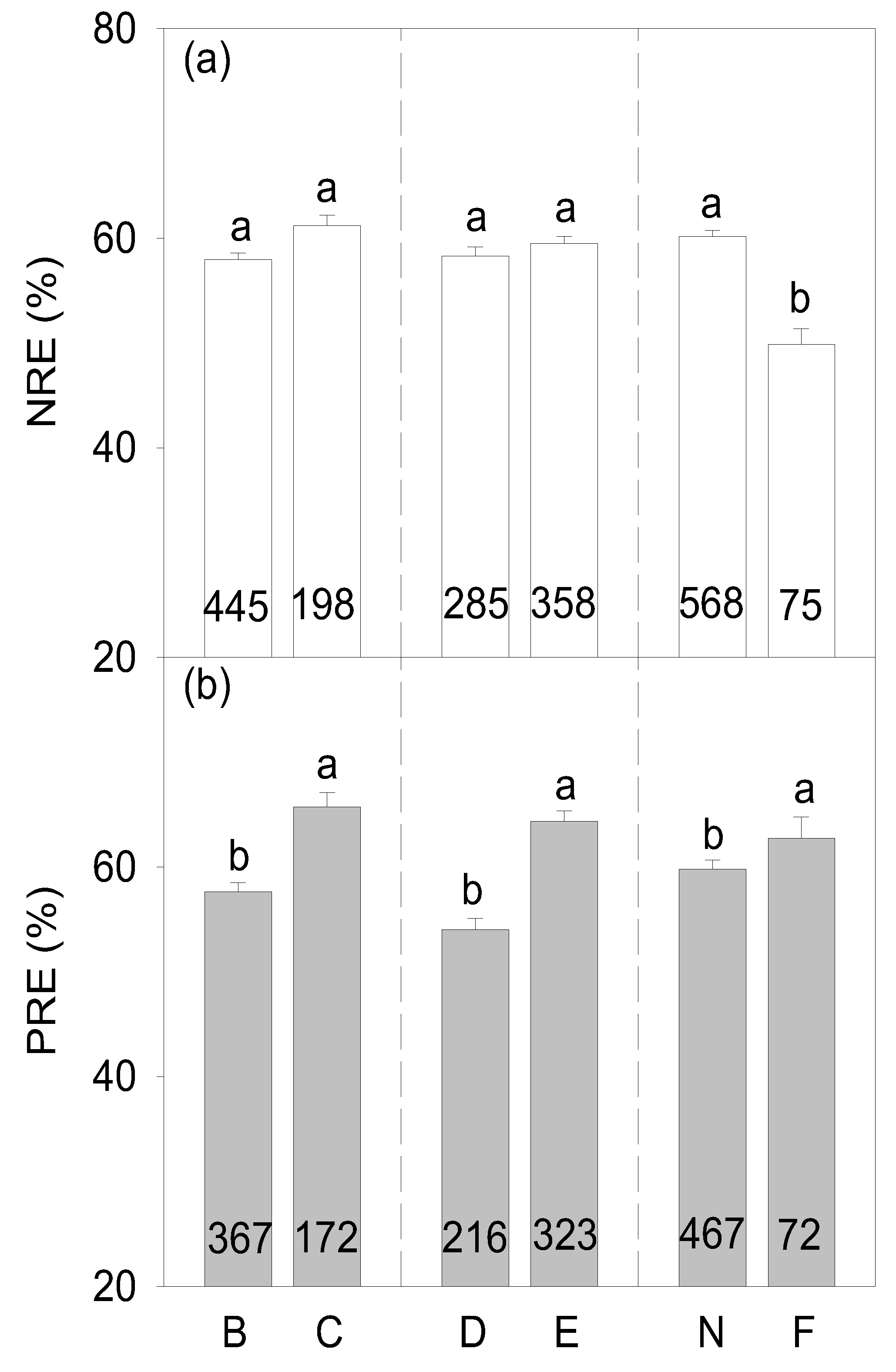
Globally, mean values of NRE and PRE in the planted forests were 58.98% ± 0.53% and 60.21% ± 0.77%, respectively, with no significant differences between them (p > 0.05, Figure 2). Generally, variations in both NRE and PRE showed remarkable differences among different climate zones and tree functional groups. While NRE significantly increased with increasing latitude (p < 0.05, Table 1, Figure 3a), PRE did not differ among climate zones (p > 0.05, Table 1, Figure 3b). Additionally, NRE did not differ between conifer and broadleaf trees (p > 0.05, Table 1, Figure 4a) and the PRE of conifer trees was higher than that of broadleaf trees (p < 0.05, Table 1, Figure 4b). Though NRE did not differ between deciduous and evergreen trees (p > 0.05, Table 1, Figure 4a), PRE was significantly higher in evergreen than deciduous trees (p < 0.001, Table 1, Figure 4b). The NRE of N-fixing trees was much lower than that of non-N-fixing trees (p < 0.001, Table 1, Figure 4a) and the PRE of N-fixing trees was much higher than that of non-N-fixing trees (p < 0.05, Table 1, Figure 4b).
3.2. Controls of Nutrient Resorption in Planted Forests
Variations in NRE and PRE at the global scale were not controlled by the same explanatory variables. Our multivariate regression analyses showed that NRE was regulated negatively by MAP and positively by green leaf P concentration (Pgreen) and latitude (LAT) (p < 0.001, Table 2). For PRE, green leaf N concentration (Ngreen) and Pgreen had positively and negatively impacted on it, respectively (p < 0.001, Table 2). The stand ages of forests had little impact on both NRE and PRE.
4. Discussion
4.1. Nutrient Resorption Patterns in Planted Forests
Our results showed that the mean values of NRE and RPE were 58.98% ± 0.53% and 60.21% ± 0.77%, respectively, for the planted forests worldwide—much higher than the assumed 50% for both NRE and PRE in most models [1]. Consistent with Zhang et al. that the NRE of Metasequoia glyptostroboides increased along latitude at the regional scale [27], we found that NRE significantly increased from tropical to boreal zones with increasing latitude. This pattern may result from three main reasons associated with low temperatures at high latitudes. First, decomposition and nutrient mineralization are high at low latitudes, which may subsequently enhance the availability of soil N [43,44]. Second, warmer temperatures and higher water availability could stimulate nutrient movement as well as root N uptake at low latitudes [45]. In high-latitude regions, however, the delivery of mass-flow nutrients from the soil to plant tissues is inhibited, which in turn, may reduce the process of nutrient resorption [20,46]. Third, the metabolic activity of plants may be inhibited at high latitudes [27]. Nutrient resorption is recognized as one of the efficient nutrient conservation mechanisms for plants to adapt to environmental conditions [12]. Plants at high latitudes usually grow quickly in order to finish development in relatively short growing seasons via high nutrient retrieval strategies that can enhance growth rate and lower dependence on the supply of soil nutrients [1,5,47]. Interestingly, PRE did not linearly increase or decrease from tropical to boreal zones, which is in line with the findings of Zhang et al., who observed a curved correlation of PRE with latitude [27]. PRE may largely depend on P availability in soils [13,48]. For example, a negative correlation between PRE and soil P was found in planted forests [27,33,49]. Those results suggest that NRE and PRE are differentially regulated by climate and soil nutrient availability.
The resorption of N and P varied among functional groups. First, the PREs of coniferous trees were significantly higher than those of broadleaf trees. Similar observations were also found in natural forests [18,20]. Coniferous trees are usually planted in nutrient-poor environments. The higher nutrient resorption could thus help them survive via reducing their dependence on soil nutrient supplies [16,50]. High nutrient resorption reflects the adaptation of plants to oligotrophic habitats [10,51]. Interestingly, NRE did not differ between coniferous and broadleaf trees. This may result from the limited number of NRE observations in coniferous planted forests, as also reported by Yuan and Chen [5]. Second, we also found that evergreen tree species presented higher PRE, whereas the NRE of deciduous species did not differ from that of the evergreen species, which is completely in line with the findings in the forest plantations in the Brazilian Amazon [29]. Possible reasons may include that evergreen species (1) are mostly planted in P-limited low latitudes [3,20] and (2) have high nutrient use efficiency and thus low nutrient loss rates associated with prolonged nutrient retention time in leaf biomass and leaf longevity [12,52]. Third, our results showed that N-fixing trees had significantly lower NRE but higher PRE than non-N-fixing trees. N-fixing trees can acquire N from the atmosphere, and they are thus less dependent on the internal N cycling process [53,54]. However, N-fixing trees may contain higher leaf N and may thus enhance PRE in order to maintain N to P stoichiometric homeostasis. These findings indicate that plant functional types exert a strong influence on nutrient resorption.
4.2. Controls of Nutrient Resorption in Planted Forests
In this study, the global-scale NRE and PRE patterns were not mainly regulated by the same ecological factors. Our multivariate regression analyses showed that NRE was primarily controlled by climate variables, supporting the observations of some previous studies [1,20,27,48]. Specially, we found that NRE was regulated negatively by MAP and positively by LAT. Climates (e.g., temperature and precipitation) most commonly affect soil N availability via microbial activities [44]. That is, soil N availability is lower under hostile climates (e.g., dry and cold at high latitude) than under climates that are favored by microbes (e.g., wet and warm at low latitude). Soil nutrient availability usually has a negative impact on nutrient resorption efficiency [41,55], leading to the phenomena that NRE correlated positively with LAT and negatively with MAP. Contrary to what we found here, Achat et al. showed that climates had only minor effects on the NRE of forests in 102 permanent forest sites across France [9]. The gap may result from the rather narrow range of climatic conditions in the study by Achat et al. [9] compared with the global climate scale considered in our study. In addition, NRE was also regulated positively by green leaf P (Pgreen). N and P in plants are usually closely related, which means changes of one nutrient could alter the other [56,57]. For example, plants may elevate NRE to maintain their N and P stoichiometric homeostasis by increasing Pgreen, and vice versa. Generally, our results suggest that the predicted global warming and changed precipitation regimes may profoundly affect N cycling in planted forests.
For the resorption of P, Ngreen had positive impacts on it, which is in accordance with the results of Wang et al. [58] and Yan et al. [4], suggesting that coupled relationships between N and P may exist in planted forests. Furthermore, we found that Pgreen had negative effects on PRE, which is supported by the findings by Kobe et al. [59]. Green leaf nutrient status is supposed to represent soil nutrient availability, whose negative impacts on nutrient resorption efficiency is widely accepted [41,59,60]. Therefore, green leaf nutrient status may negatively relate to nutrient resorption efficiency. We also found that climate variables were not the main explanatory factors for PRE, supporting the idea that soil properties and soil parent materials other than climate had more impact on soil P availability [48]. Green leaf nutrient concentration may be a good indicator for PRE in planted forests.
5. Conclusions
To our best knowledge, this synthesis provides the first comprehensive analysis of NRE and PRE in planted forests worldwide. Generally, NRE significantly increased along climate zones, while no significant differences were observed for PRE among climate zones. These results suggest differential impacts of climates on NRE and PRE. In terms of plant functional groups, the PRE of conifer trees was higher than that of broadleaf trees; evergreen trees had higher PRE than deciduous trees; non-N-fixing trees had higher NRE but lower PRE than N-fixing trees. These findings indicate that plant functional types exert strong impacts on nutrient resorption. Additionally, multivariate regression analyses showed that variations in NRE were mainly regulated by mean annual precipitation (MAP) and latitude, indicating that the predicted global warming and changed precipitation regimes may profoundly affect N cycling in planted forests. Variations in PRE were dominantly controlled by green leaf nutrient concentrations (N and P), suggesting green leaf nutrient concentrations may be good indicators for PRE in planted forests.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1999-4907/10/3/201/s1.
Author Contributions
Conceptualization: X.X.; Data curation: D.J.; Formal analysis: D.J.; Funding acquisition: D.J., H.R., and X.X.; Project administration: X.X.; Supervision: X.X.; Writing—original draft: D.J.; Writing—review and editing: Q.G., Q.L., Y.L., J.V., Z.S., H.R., and X.X.
Funding
This study was financially supported by the National Key Research and Development Program of China (2016YFD0600204), the Recruitment Program for Young Professionals (Thousand Youth Talents Plan), the Jiangsu Specially-Appointed Professors Program, the Six Talent Peaks Program of Jiangsu Province (JY-041& TD-XYDXX-006), the Nanjing Forestry University Science Fund for Distinguished Young Scholars (the “5151” Talent Program), the Jiangsu Collegiate Science and Technology Fund for the Excellent Innovative Research Teams, the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), the Doctorate Fellowship Foundation of Nanjing Forestry University, the Research Innovation Program for College Graduates of Jiangsu Province (KYLX16_0833), and the Scientific and Technological Innovation Program for College Students of Nanjing Forestry University (DXSKC-201617).
Acknowledgments
We thank many lab members for their help with the manuscript preparation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, A.P.; Kicklighter, D.W.; Melillo, J.M.; Felzer, B.S.; Schlosser, C.A.; Cronin, T.W. Consequences of considering carbon-nitrogen interactions on the feedbacks between climate and the terrestrial carbon cycle. J. Clim. 2008, 21, 3776–3796. [Google Scholar] [CrossRef]
- Aerts, R. Nutrient Resorption from Senescing Leaves of Perennials: Are there General Patterns? J. Ecol. 1996, 84, 597–608. [Google Scholar] [CrossRef]
- Yan, T.; Lu, X.T.; Zhu, J.J.; Yang, K.; Yu, L.Z.; Gao, T. Changes in nitrogen and phosphorus cycling suggest a transition to phosphorus limitation with the stand development of larch plantations. Plant Soil 2018, 422, 385–396. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Fan, Z.X.; Zhao, Q.; Wang, M.C.; Ran, J.Z.; Huang, H.; Niklas, K.J. Global data analysis shows that soil nutrient levels dominate foliar nutrient resorption efficiency in herbaceous species. Front. Plant Sci. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Killingbeck, K.T. Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Schmidt, M.; Veldkamp, E.; Corre, M.D. Tree-microbial biomass competition for nutrients in a temperate deciduous forest, central Germany. Plant Soil 2016, 408, 227–242. [Google Scholar] [CrossRef]
- Achat, D.L.; Pousse, N.; Nicolas, M.; Augusto, L. Nutrient remobilization in tree foliage as affected by soil nutrients and leaf life span. Ecol. Monogr. 2018, 88, 408–428. [Google Scholar] [CrossRef] [Green Version]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar] [CrossRef]
- Deng, M.F.; Liu, L.L.; Jiang, L.; Liu, W.X.; Wang, X.; Li, S.P.; Yang, S.; Wang, B. Ecosystem scale trade-off in nitrogen acquisition pathways. Nat. Ecol. Evol. 2018, 2, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Brant, A.N.; Chen, H.Y.H. Patterns and mechanisms of nutrient resorption in plants. Crit. Rev. Plant Sci. 2015, 34, 471–486. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Negative effects of fertilization on plant nutrient resorption. Ecology 2015, 96, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Z.; Liu, L.L.; Peng, S.S.; Penuelas, J.; Zeng, H.; Piao, S.L. Age-related modulation of the nitrogen resorption efficiency response to growth requirements and soil nitrogen availability in a temperate Pine plantation. Ecosystems 2016, 19, 698–709. [Google Scholar] [CrossRef]
- Sedjo, R.A. The potential of High-Yield Plantation Forestry for Meeting Timber Needs. In Planted Forests: Contributions to the Quest for Sustainable Societies; Boyle, J.R., Winjum, J.K., Kavanagh, K., Jensen, E.C., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 339–359. [Google Scholar]
- Yan, T.; Lu, X.T.; Yang, K.; Zhu, J.J. Leaf nutrient dynamics and nutrient resorption: A comparison between larch plantations and adjacent secondary forests in Northeast China. J. Plant Ecol. 2016, 9, 165–173. [Google Scholar] [CrossRef]
- Sun, X.; Kang, H.; Chen, H.Y.H.; Bjorn, B.; Samuel, B.F.; Liu, C. Biogeographic patterns of nutrient resorption from Quercus variabilis Blume leaves across China. Plant Biol. 2016, 18, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.Y.; Han, W.X.; Chen, Y.H.; Fang, J.Y. Resorption proficiency and efficiency of leaf nutrients in woody plants in eastern China. J. Plant Ecol. 2013, 6, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.Y.; Lu, X.T.; Hartmann, H.; Keller, A.; Han, X.G.; Trumbore, S.; Phillips, R.P. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales. Glob. Ecol. Biogeogr. 2018, 27, 875–885. [Google Scholar] [CrossRef]
- Yan, T.; Zhu, J.J.; Yang, K. Leaf nitrogen and phosphorus resorption of woody species in response to climatic conditions and soil nutrients: A meta-analysis. J. For. Res. 2018, 29, 905–913. [Google Scholar] [CrossRef]
- Han, W.X.; Tang, L.Y.; Chen, Y.H.; Fang, J.Y. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants. PLoS ONE 2013, 8, e83366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.N.; Wang, J.Y.; Guo, Z.W.; Wang, G.G.; Zeng, D.H.; Wu, T.G. Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. For. Ecol. Manag. 2018, 430, 445–450. [Google Scholar] [CrossRef]
- Maeda, Y.; Tashiro, N.; Enoki, T.; Urakawa, R.; Hishi, T. Effects of species replacement on the relationship between net primary production and soil nitrogen availability along a topographical gradient: Comparison of belowground allocation and nitrogen use efficiency between natural forests and plantations. For. Ecol. Manag. 2018, 422, 214–222. [Google Scholar] [CrossRef]
- Xu, S.; Zhou, G.; Tang, X.; Wang, W.; Wang, G.; Ma, K.; Han, S.; Du, S.; Li, S.; Yan, J.; et al. Different spatial patterns of nitrogen and phosphorus resorption efficiencies in China’s forests. Sci. Rep. 2017, 7, 10584. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Li, L.H. Soil water status influences plant nitrogen use: A case study. Plant Soil 2007, 301, 303–313. [Google Scholar] [CrossRef]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, W.H.; Yu, M.K.; Wang, G.G.; Wu, T.G. Latitudinal patterns of leaf N, P stoichiometry and nutrient resorption of Metasequoia glyptostroboides along the eastern coastline of China. Sci. Total Environ. 2018, 618, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.L.; Addo-Danso, S.D.; Wu, P.F.; Li, S.B.; Zou, X.H.; Zhang, Y.; Ma, X.Q. Leaf resorption efficiency in relation to foliar and soil nutrient concentrations and stoichiometry of Cunninghamia lanceolata with stand development in southern China. J. Soils Sediments 2016, 16, 1448–1459. [Google Scholar] [CrossRef]
- Machado, M.R.; Sampaio, P.D.; Ferraz, J.; Camara, R.; Pereira, M.G. Nutrient retranslocation in forest species in the Brazilian Amazon. Acta Sci.-Agron. 2016, 38, 93–101. [Google Scholar] [CrossRef]
- Ye, G.F.; Zhang, S.J.; Zhang, L.H.; Lin, Y.M.; Wei, S.D.; Liao, M.M.; Lin, G.H. Age-related changes in nutrient resorption patterns and tannin concentration of Casuarina equisetifolia plantations. J. Trop. For. Sci. 2012, 24, 546–556. [Google Scholar] [CrossRef]
- Lal, C.B.; Annapurna, C.; Raghubanshi, A.S.; Singh, J.S. Foliar demand and resource economy of nutrients in dry tropical forest species. J. Veg. Sci. 2001, 12, 5–14. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Jiang, G.M.; Wan, S.Q.; Zhang, W.H.; Chen, Q.S. Nitrogen resorption from senescing leaves in 28 plant species in a semi-arid region of northern China. J. Arid Environ. 2005, 63, 191–202. [Google Scholar] [CrossRef]
- Pang, D.B.; Wang, G.Z.; Li, G.J.; Sun, Y.L.; Liu, Y.G.; Zhou, J.X. Ecological stoichiometric characteristics of two typical plantations in the Karst ecosystem of southwestern China. Forests 2018, 9, 56. [Google Scholar] [CrossRef]
- MacDicken, K.; Jonsson, Ö.; Piña, L.; Marklund, L.; Maulo, S.; Contessa, V.; Adikari, Y.; Garzuglia, M.; Lindquist, E.; Reams, G.; et al. Global Forest Resources Assessment 2015: How Are the World’S Forests Changing? Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2015; pp. 1–54. [Google Scholar]
- Van Dijk, A.I.J.M.; Keenan, R.J. Planted forests and water in perspective. For. Ecol. Manag. 2007, 251, 1–9. [Google Scholar] [CrossRef]
- Fang, J.Y.; Chen, A.P.; Peng, C.H.; Zhao, S.Q.; Ci, L.J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Goodale, C.L.; Apps, M.J.; Birdsey, R.A.; Field, C.B.; Heath, L.S.; Houghton, R.A.; Jenkins, J.C.; Kohlmaier, G.H.; Kurz, W.; Liu, S.; et al. Forest carbon sinks in the northern Hemisphere. Ecol. Appl. 2002, 12, 891–899. [Google Scholar] [CrossRef]
- Netzer, F.; Schmid, C.; Herschbach, C.; Rennenberg, H. Phosphorus-nutrition of European beech (Fagus sylvatica L.) during annual growth depends on tree age and P-availability in the soil. Environ. Exp. Bot. 2017, 137, 194–207. [Google Scholar] [CrossRef]
- Weatherall, A.; Proe, M.F.; Craig, J.; Cameron, A.D.; Midwood, A.J. Internal cycling of nitrogen, potassium and magnesium in young Sitka spruce. Tree Physiol. 2006, 26, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.S.; Niklas, K.J.; Liu, Y.; Fang, X.M.; Wan, S.Z.; Wang, H. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 2015, 35, 1106–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.C.; Townsend, A.R.; Davidson, E.A.; Cleveland, C.C. Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytol. 2012, 196, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Heerwaarden, L.M.; Toet, S.; Aerts, R. Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: Facts and solutions. Oikos 2003, 101, 664–669. [Google Scholar] [CrossRef]
- Oleksyn, J.; Reich, P.B.; Zytkowiak, R.; Karolewski, P.; Tjoelker, M.G. Nutrient conservation increases with latitude of origin in European Pinus sylvestris populations. Oecologia 2003, 136, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, P.; Falkengren-Grerup, U.; Bengtsson, G. Relieving substrate limitation-soil moisture and temperature determine gross N transformation rates. Oikos 2005, 111, 81–90. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramer, M.D.; Hawkins, H.J.; Verboom, G.A. The importance of nutritional regulation of plant water flux. Oecologia 2009, 161, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Westoby, M. Nutrient concentration, resorption and lifespan leaf traits of Australian sclerophyll species. Funct. Ecol. 2003, 17, 10–19. [Google Scholar] [CrossRef]
- Augusto, L.; Achat, D.L.; Jonard, M.; Vidal, D.; Ringeval, B. Soil parent material-A major driver of plant nutrient limitations in terrestrial ecosystems. Glob. Chang. Biol. 2017, 23, 3808–3824. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.P.; Chen, X.; Huang, B.; Chi, G.Y. Distribution changes of phosphorus in soil-plant systems of larch plantations across the chronosequence. Forests 2018, 9, 563. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Wan, S.Q. Foliar nitrogen dynamics and nitrogen resorption of a sandy shrub Salix gordejevii in northern China. Plant Soil 2005, 278, 183–193. [Google Scholar] [CrossRef]
- Liu, J.T.; Gu, Z.J.; Shao, H.B.; Zhou, F.; Peng, S.Y. N–P stoichiometry in soil and leaves of Pinus massoniana forest at different stand ages in the subtropical soil erosion area of China. Environ. Earth Sci. 2016, 75, 1091. [Google Scholar] [CrossRef]
- Escudero, A.; Del Arco, J.M.; Sanz, I.C.; Ayala, J. Effects of leaf longevity and retranslocation efficiency on the retention time of nutrients in the leaf biomass of different woody species. Oecologia 1992, 90, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.D.; Reich, P.B. Modest enhancement of nitrogen conservation via retranslocation in response to gradients in N supply and leaf N status. Plant Soil 2009, 316, 193–204. [Google Scholar] [CrossRef]
- Zhao, G.S.; Shi, P.L.; Wu, J.S.; Xiong, D.P.; Zong, N.; Zhang, X.Z. Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the Tibetan Changtang Plateau. Ecol. Evol. 2017, 7, 7201–7212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, P.; Turner, B.L.; Lambers, H.; Laliberte, E. Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence. J. Ecol. 2014, 102, 396–410. [Google Scholar] [CrossRef]
- Deng, M.F.; Liu, L.L.; Sun, Z.Z.; Piao, S.L.; Ma, Y.C.; Chen, Y.W.; Wang, J.; Qiao, C.L.; Wang, X.; Li, P. Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-rupprechtii plantations. New Phytol. 2016, 212, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Chen, Q.; Elser, J.J.; He, N.; Wu, H.; Zhang, G.; Wu, J.; Bai, Y.; Han, X. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol. Lett. 2010, 13, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.N.; Lu, J.Y.; Yang, H.M.; Zhang, X.; Luo, C.L.; Zhao, Y.X. Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil 2014, 383, 301–312. [Google Scholar] [CrossRef]
- Kobe, R.K.; Lepczyk, C.A.; Iyer, M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 2005, 86, 2780–2792. [Google Scholar] [CrossRef]
- Tsujii, Y.; Onoda, Y.; Kitayama, K. Phosphorus and nitrogen resorption from different chemical fractions in senescing leaves of tropical tree species on Mount Kinabalu, Borneo. Oecologia 2017, 185, 171–180. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Global distribution of 149 independent sampling sites included in this global data synthesis.
Figure 1.
Global distribution of 149 independent sampling sites included in this global data synthesis.

Figure 2.
N and P resorption efficiency (%, NRE and PRE) in planted forests (mean ± SE). Values in the bars represent the number of observations. Different lowercase letters represent statistical differences at p = 0.05 between NRE and PRE.
Figure 2.
N and P resorption efficiency (%, NRE and PRE) in planted forests (mean ± SE). Values in the bars represent the number of observations. Different lowercase letters represent statistical differences at p = 0.05 between NRE and PRE.

Figure 3.
N and P resorption efficiency (%, NRE (a) and PRE (b)) among different climate zones in planted forests (mean ± SE). Values in the bars represent the number of observations. Different lowercase letters represent statistical differences at p = 0.05 among the climate zones.
Figure 3.
N and P resorption efficiency (%, NRE (a) and PRE (b)) among different climate zones in planted forests (mean ± SE). Values in the bars represent the number of observations. Different lowercase letters represent statistical differences at p = 0.05 among the climate zones.

Figure 4.
N and P resorption efficiency (%, NRE (a) and PRE (b)) between broadleaf (B) vs. conifer (C), deciduous (D) vs. evergreen (E), and non-N-fixing (N) vs. N-fixing (F) tree groups in planted forests (mean ± SE). Values in the bars represent the number of observations. Different lowercase letters represent statistical differences at p = 0.05 between the different functional types.
Figure 4.
N and P resorption efficiency (%, NRE (a) and PRE (b)) between broadleaf (B) vs. conifer (C), deciduous (D) vs. evergreen (E), and non-N-fixing (N) vs. N-fixing (F) tree groups in planted forests (mean ± SE). Values in the bars represent the number of observations. Different lowercase letters represent statistical differences at p = 0.05 between the different functional types.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
ANOVA table of the linear mixed-effects models for N and P resorption efficiency (%, NRE and PRE). *: <0.05, ***: <0.001.
Table 1.
ANOVA table of the linear mixed-effects models for N and P resorption efficiency (%, NRE and PRE). *: <0.05, ***: <0.001.
| Resorption | Fixed Term | df | F, p |
|---|---|---|---|
| NRE | Climate zones | 3 | 3.56 * |
| Broadleaf vs. Conifer | 1 | 0.56 | |
| Deciduous vs. Evergreen | 1 | 2.48 | |
| Non-N-fixing tree vs. N-fixing tree | 1 | 15.85 *** | |
| PRE | Climate zones | 3 | 0.79 |
| Broadleaf vs. Conifer | 1 | 4.74 * | |
| Deciduous vs. Evergreen | 1 | 12.93 *** | |
| Non-N-fixing tree vs. N-fixing tree | 1 | 5.06 * |
Table 2.
Stepwise regressions of N and P resorption efficiency (%, NRE and PRE) with stand age (year, Age), absolute latitude (°, LAT), mean annual precipitation (mm, MAP), and green leaf N and P concentrations (mg g−1, Ngreen and Pgreen, respectively) in planted forests. ***: <0.001.
Table 2.
Stepwise regressions of N and P resorption efficiency (%, NRE and PRE) with stand age (year, Age), absolute latitude (°, LAT), mean annual precipitation (mm, MAP), and green leaf N and P concentrations (mg g−1, Ngreen and Pgreen, respectively) in planted forests. ***: <0.001.
| Variables | Regression | n | r2 | Excluded Variables | |
|---|---|---|---|---|---|
| NRE | MAP | y = 67.48 − 0.01 × MAP | 476 | 0.09 *** | Age, Ngreen, Pgreen, LAT |
| MAP, Pgreen | y = 62.65 − 0.01 × MAP + 2.70 × Pgreen | 476 | 0.12 *** | Age, Ngreen, LAT | |
| MAP, Pgreen, LAT | y = 54.68 − 0.01 × MAP + 2.36 × Pgreen + 0.21 × LAT | 476 | 0.14 *** | Age, Ngreen | |
| PRE | Pgreen | y = 66.11 − 4.12 × Pgreen | 476 | 0.05 *** | Age, Ngreen, LAT, MAP |
| Pgreen, Ngreen | y = 60.31 − 5.33 × Pgreen + 0.41 × Ngreen | 476 | 0.07 *** | Age, LAT, MAP |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jiang, D.; Geng, Q.; Li, Q.; Luo, Y.; Vogel, J.; Shi, Z.; Ruan, H.; Xu, X. Nitrogen and Phosphorus Resorption in Planted Forests Worldwide. Forests 2019, 10, 201. https://0-doi-org.brum.beds.ac.uk/10.3390/f10030201
AMA Style
Jiang D, Geng Q, Li Q, Luo Y, Vogel J, Shi Z, Ruan H, Xu X. Nitrogen and Phosphorus Resorption in Planted Forests Worldwide. Forests. 2019; 10(3):201. https://0-doi-org.brum.beds.ac.uk/10.3390/f10030201
Chicago/Turabian StyleJiang, Dalong, Qinghong Geng, Qian Li, Yiqi Luo, Jason Vogel, Zheng Shi, Honghua Ruan, and Xia Xu. 2019. "Nitrogen and Phosphorus Resorption in Planted Forests Worldwide" Forests 10, no. 3: 201. https://0-doi-org.brum.beds.ac.uk/10.3390/f10030201
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.