2.1. Location and Study Design
The investigation was conducted in the Alpine biogeographic region in the northern part of the Eastern Intermediate Alps in Austria at approximately 47°26′ east and 15°05′ north. The mean annual temperature and mean annual precipitation are 5.2 °C and 1510 mm, respectively. The prevailing silvicultural management is strip clear cutting [
20,
21,
22], where after felling, the forest is regenerated naturally in three strips. In the inner strip, Norway spruce dominates, while in the outer strip the light-demanding European larch prevails. In the transitional zone, mixed stands of these two species develop (
Figure 1). The total strip width is approximately two to three times the height of the dominant trees.
At four locations, we established three plots: one of each forest type (inner, outer, and transitional strip). All plots were on steep Northwest-oriented slopes at altitudes between 900 and 1300 m asl The plot size was between 150 and 1600 m
2 and was chosen in such a way that each plot contained at least 100 trees of the dominant species. The selected stands can be regarded as nearly even-aged. The mean coefficient of variation of the tree age within the plots, estimated from the sample trees, was about ±5% for spruce and ±10% for larch. The age of the stands varied between 40 and 170 years, the dominant height ranged between 20 and 40 m, the basal area between 30 and 50 m
2 ha
−1, and the stocking degree between 0.7 and 1.0. For the plot characteristics, see
Table 1, for further details please refer to Dirnberger et al. [
15] and Sterba et al. [
23].
2.4. Leaf Area and Competition
For each individual single tree, the 24 sample branches were used to estimate the coefficients
and
of an equation
with
, the leaf area of the branch and
the branch diameter. This equation was then used to estimate the leaf area of all branches of the respective sample tree. These leaf areas had been used by Dirnberger et al. [
15] to develop an equation for estimating the whole leaf area of each tree in the stand, as it depended on the crown surface area, the diameter at breast height (dbh) and the stand’s stocking degree for spruce, and on crown surface area, the dbh and the proportion of spruce for larch. With these equations, the total leaf area of all trees in the stands was calculated. For quantifying the fraction of the stand area that was potentially available (
APA) for a tree, we used the rasterizing approach [
29], based on the circlebow model by Römisch [
30]. Thus, the whole plot was rasterized (pixel size = 10 cm × 10 cm) and the squared distance of every pixel to all trees, weighted by their leaf area was calculated (Equation (1)).
with
the distance from the
-th pixel to the
-th tree, and
, the leaf area of the
-th tree. The pixel was then assigned to the tree
, where
is minimal. Thus, a weighted Voronoi diagram of the stand was constructed, resulting in the areas potentially available (
) of the trees (
Figure 2). The leaf area per potentially available area yields the area exploitation index (
AEI) according to Gspaltl et al. [
29] (Equation (2)).
AEI can be understood as an individual tree density measure characterizing the small-scale competition.
The total boundary
of the
of a single tree can be separated into (i) the boundary to spruce neighbours
and (ii) the boundary to other species. The proportion of
to
can then be understood as the contribution of spruce to the overall competition on the respective tree
(
Figure 2, Equation (3)).
For our stands, containing exactly two species, the contribution of larch to the overall competition of a tree can also be expressed in terms of (), so that we decided only to use the latter one for further analysis.
2.5. Leaf Area Distribution
For every sample tree, we fitted a beta distribution to the data of the relative cumulative leaf area as a function of the relative distance
from the top of the tree. Relative means, that the distance is divided by the crown length and thus
is 0 at the top of the tree and 1 at the crown base. The corresponding probability density function is given in Equation (4),
is the the gamma function. Following the discussion of Fielitz and Myers [
31,
32] and Romesburg [
33] we decided to estimate the coefficients
and
of the beta distribution by the leaf area weighted relative mean distance of the branch from the top
, and the corresponding variance
, using Equations (5) and (6) for every tree, rather than using a maximum likelihood approach. Given that the measured distances and leaf areas are no sample, but all branches of a sample tree were observed, deviations between estimates and true parameters may originate from the fact that the beta distribution itself is only an approximation of the discrete empirical leaf area distribution in the crown, rather than from the method of estimation.
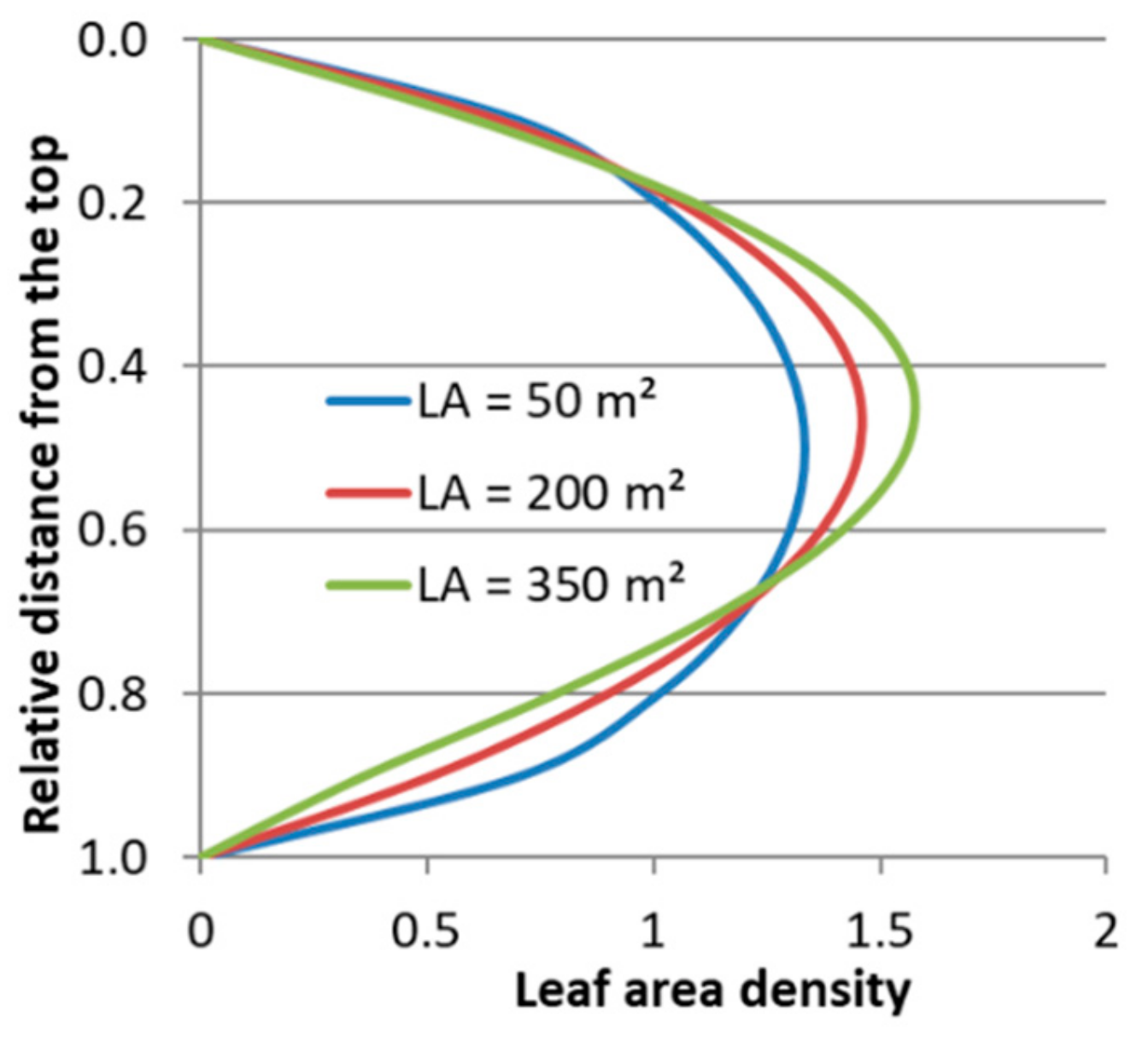
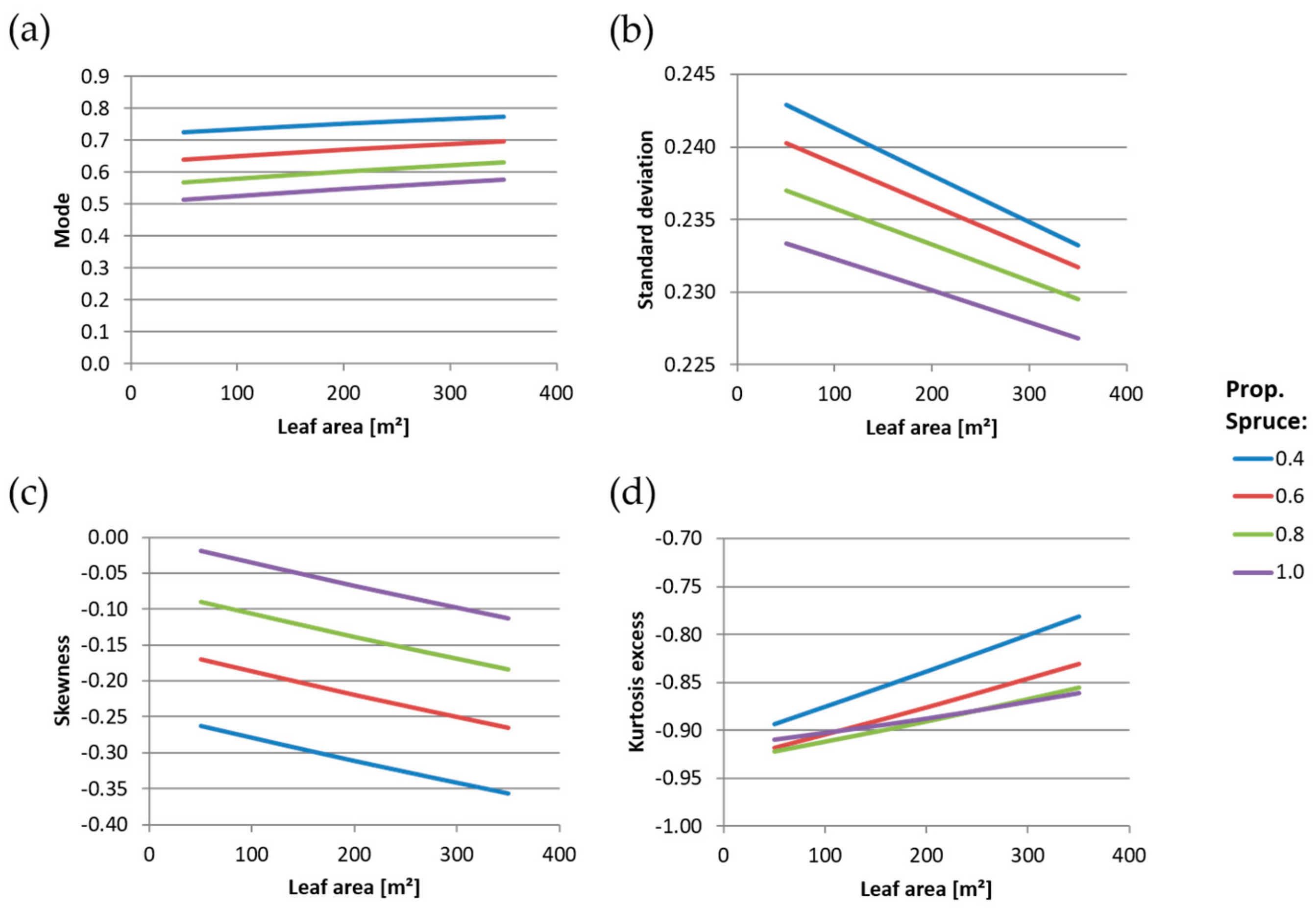
For further characterizing the distribution, the mode, the standard deviation, the skewness and the kurtosis excess were calculated from the parameters and .
The mode is defined as a value of
were the probability density function (Equation (4)) has its maximum. For our data that means the mode is the relative distance from the top of the tree to the height with the highest density within the crown.
The standard deviation results as:
The skewness is a measure of the asymmetry of the probability distribution function (pdf), a negative skewness indicates that the tail is on the left side of the pdf, while a positive skewness indicates that the tail is on the right. For our data that means a negative skewness indicates that the leaf area is predominantly located in the lower part of the crown, while a positive skewness indicates that the leaf area is predominantly located in the upper part of the crown.
The kurtosis excess is defined as kurtosis minus 3, so that an kurtosis excess of zero represents a mesokurtic distribution, while positive and negative kurtosis excess values stand for leptokurtic and platykurtic distributions respectively.
Then, an investigation into the dependency of the parameters
and
on the sample trees’ traits and their environment was conducted. Therefore, mixed models with individual tree measures and stand measures as fixed effects, and the plot as random effect, were evaluated using the R [
34] packages, lme4 [
35] and lmerTest [
36] the initial model is given by Equation (11).
is the value of the response variable (one of the two parameters
and
) for the
-th tree at sample plot
,
are the fixed-effect coefficients,
is the random effect coefficient for sample plot
, and
is the error for tree
at sample plot
. The fixed-effect regressors are
for the tree measures, and
for the stand characteristics,
is the random-effect regressor. The tree measures were the dbh, the tree height, crown length, crown width, leaf area, local density and the proportion of spruce in the competition. The local density was described by the
AEI (Equation (2)) and the proportion of spruce in the competition
(calculated as described by Equation (3) and
Figure 2). The stand characteristics were age, quadratic mean diameter, dominant height, stocking and the proportion of spruce. The latter were based on the relative stand density index (SDI), using the maximum density lines as evaluated by Vospernik and Sterba [
26] for the 95th percentile. Starting with the full initial model as defined in Equation (11), we used a backward variable selection to reduce model complexity and avoid overfitting.
For each tree in the stand, we then calculated its total leaf area using the equations of Dirnberger et al. [
15]. The equations finally determined for the relative leaf area distribution were then used for calculating the leaf area of each tree in two-meter sections, starting at the ground. Thus, we achieved the distribution of leaf area by species in two-meter sections for each plot. The difference between the modes of the stand’s leaf area distribution of larch and that of spruce was then investigated for its relationship with other stands’ characteristics, including spatial distribution characteristics, i.e., species intermingling and horizontal distribution. Species intermingling was described by Pielou’s segregation index [
37], based on the nearest neighbor principal. The index is bound between −1 and 1, positive values indicate that the species are segregated, while negative ones indicate association of the different species. An index value of zero means that the probability for a neighboring species is stochastically independent of the species of the subject tree.
The horizontal distribution was characterized by the Clark–Evans index [
38], indicating a clustered distribution with values <1 and more regular distributions with values >1. Thus, a CE
spruce < 1 would indicate a stronger horizontal separation of the species and vice versa.
For a detailed description of the indices, see Del Rio et al. [
39].













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}